
Abstract
Objective:
In this study, we sought to characterize the tumor necrosis factor α (TNFα) baseline operational capacity in mature fetuses and their mothers prior to the onset of labor.
Materials and Methods:
We used an experimental pregnant nonhuman primate model to measure the plasma concentration of TNFα, TNF transmembrane receptor I (TNFRI), and TNFRII with validated enzyme-linked immunosorbent assays. Coefficients of correlations between the maternal and the fetal values and the soluble TNFα, TNFRI, or TNFRII concentrations and ratios were calculated.
Results:
The TNFα/TNFRI ratio was 3 times lower in fetal circulation than in maternal circulation. No correlations were noted between the maternal and the fetal TNFα, TNFRI, or TNFRII plasma concentrations.
Conclusions:
These findings suggest that the fetal and maternal baseline circulatory operational capacities of TNFα are independent of each other and tuned differently. This differential regulation of TNFα in fetal and maternal circulation at the end of pregnancy may be guided to protect the fetus from the systemic inflammatory response that is essential for the mechanisms of labor to proceed in the mother.
Keywords
The tumor necrosis factor α (TNFα) gene maps to 6p21.3 in the genetic region of the major histocompatibility complex. 1 Cachectin is the 157 amino acid residue protein product of this gene with a molecular weight of approximately 17 kDa. This protein is a member of the TNF superfamily which produces a vast range of effects including the expression of other cytokines, immune receptors, proteases, growth factors, and cell cycle genes that regulate basic cellular processes. 2 –7 The biological active native form of TNFα is a trimer 8 and acts using transmembrane cellular receptors TNF transmembrane receptor I (TNFRI; also called CD120a, p55, or htr antigen) and TNFRII (also called CD120b, p75, or utr antigen) that are constitutively but differentially expressed. 9 The TNFRI protein has 426 amino acid residues and TNFRII has 439 amino acid residues with similarities in the pattern of cysteine locations in 4 domains in the extracellular portions of the protein and with unrelated intracellular domains suggesting different signal transduction pathways. 6,10 To modulate systemic TNFα actions, the extracellular section of TNFRI and TNFRII can be cleaved from the transmembrane domain; these truncated soluble forms of the receptors can bind to TNFα and neutralize the engagement capacity of TNFα at the cellular level serving also as a reservoir for the controlled release of TNFα when needed. 10 –15 The serum levels of TNF binding protein 1 (sTNFRI) and TNF binding protein 2 (sTNFRII) vary among individuals but are stable over time for given individuals. 11
During early development, TNFα, TNFRI, and TNFRII are expressed by the placenta villous stroma cells and trophoblast cells 16 –23 contributing to early placenta morphogenesis 24 and to the maintenance of feto-maternal homeostasis. 17,18,25 The TNFα can bind to the endothelial cells TNFRI participating in the induction of apoptosis of the vascular smooth muscle cells surrounding the maternal spiral arteries. 23 Prior to 9 weeks gestation, the concentration of sTNFα is higher in maternal circulation than in the gestational sac extracelomic fluid while the concentration of sTNFRI is higher in the gestational sac celomic fluid than in maternal circulation. 26 The levels of sTNFRI and sTNFRII are higher in the urine of pregnant women than in control 27 and although racial differences may exist in cellular responses to TNFα and other cytokines that may explain variances in the prevalence of disorders caused by endothelial dysfunction, 28 presently, there is no TNFα gene polymorphism categorically associated with increased risk of intrauterine fetal demise, preeclampsia, preterm delivery, or small for gestational age newborns. 29 In an animal model, however, chronic TNFα administration to the mother caused reduced glucose tolerance and a relative macrosomia in the female offspring due to accelerated adipose tissue development. 30
After birth, TNFα in circulation is primarily produced by monocytes and macrophages and acts as a key player in the systemic inflammatory response syndrome (SIRS). 2,8,31,32 The SIRS is defined by the presence of high fever, tachycardia, tachypnea, and leukocytosis with more than 10% immature neutrophils in circulation. 33 When the TNFα response is followed by elevated interleukins 1 and 6, endothelial damage and activation of the coagulation cascade may become the genesis of severe sepsis with organ dysfunction. 2,3,8,31,32,34,35 The TNFα also acts in the neuroimmune crosstalk in the central nervous system diseases by providing signals to infiltrating immune cells. 36
In circulation, the concentration of TNFRI and TNFRII correlate with TNFα concentration suggesting that the stimuli which cause TNFα levels to rise also induce shedding of TNFRI and TNFRII. 19,37 –39 At the moment, the relationships between the concentrations of TNFα, TNFRI, and TNFRII in the fetal circulation remain unknown and thus the pathophysiology of the fetal inflammatory response syndrome is not completely understood. 40 –42 In this study, we sought to characterize the baseline TNFα operational capacity in fetal circulation prior to the onset of labor. We hypothesized that the maternal and fetal plasma concentrations of TNFα, TNFRI, and TNFRII would be similar and would correlate with each other.
Materials and Methods
We used a pregnant nonhuman primate model that has been validated as an experimental surrogate to human pregnancies. 43,44 A total of 10 time pregnant baboons weighing 18 to 20 kg were suitable for this study because of lack of previous surgical history and fetal viability on sonographic evaluation. All animals were obtained, operated, and housed at the breeding colony of the Biological Resource Laboratory at the University of Oklahoma. All animals had a sonographic evaluation with a maternal blood sample at 38 to 42 days from fertilization. One animal lost the pregnancy. The remaining 9 animals were acclimated to the indoor facilities 3 days prior to the planned delivery and maintained under a controlled photoperiod (lights on from 06:00 to 18:00 daily) and controlled room temperature (21°C -23°C). The animals were fed a daily diet of Purina 5045 High Protein Monkey Chow pellets (Ralston Purina Company, St Louis, Missouri) as well as fresh fruit. Water was available continuously. The study was conducted at the University of Oklahoma under an Animal Care Committee approved protocol. The American Association for Accreditation of Laboratory Animal Care accredited the facilities at Oklahoma University.
Surgical Intervention of Animals
Animals were operated at 175 to 180 days of gestation (spontaneous term delivery occurs in baboons at approximately 184 ± 2 days of gestation), after a 12-hour period of food deprivation. Animals were initially anesthetized with an intramuscular injection of ketamine (4 mg) and an intravenous injection of Thiamylal (2 mL intravenously [iv] of a 2.5% solution; Parke Davis, New Jersey). Animals were then placed in supine position in the operating table, intubated, and prepped in the usual manner for an obstetrical laparotomy following the primate research laboratory protocol. Anesthesia for surgery was achieved with isofluorane 1.5% and N2O on a one-to-one basis. The respiratory rate was maintained between 8 and 12, depending on the results obtained by pulse oximetry. Cesarean deliveries were achieved via a 10 cm midline laparotomy made just below the umbilicus. A transverse hysterotomy was then performed, and the fetal occiput was first delivered and then the entire body. The neonate was then handed to the awaiting doctor in veterinary medicine. The uterine incision and abdominal fascia were closed with absorbable sutures (0) and the skin incision with a subcutaneous 3-0 vicryl suture. All surgical procedures lasted less than 30 minutes. Animals were then placed in their cages and observed until complete recovery.
Determination of TNFα, TNFRI, and TNFRII Plasma Concentrations
A total of thirty-four 5 mL blood samples were collected (10 maternal blood samples at 38-42 days postmating; 9 maternal blood samples 30 minutes before the operation when an iv line connected to a normal saline solution was started; 9 umbilical cord venous samples after the cord was clamped and prior to delivery of the placenta from the uterus; and 6 maternal blood samples 30 minutes after the skin incision was closed, with the animal breathing on her own and at the time the Foley catheter and the iv line were removed). After collection, blood samples were transferred into chilled, heparinized polypropylene tubes and centrifuged at 4°C at 1200g for 5 minutes. The plasma was removed, divided into aliquots, frozen in liquid nitrogen, and stored at −80°C until assayed.
Sensitive enzyme-linked immunoassays designed to measure human sTNFα, sTNFRI, and sTNFRII were used to determine their concentrations in baboon plasma. Soluble TNFα, TNFRI, and TNFRII immunoassays were obtained from R&D System (Minneapolis, Minnesota,) validated by the manufacture of the assay for use with human serum and plasma. We have used these assays in our study because nonhuman primate-specific sTNFα, sTNFRI, and sTNFRII immunoassays are not commercially available. These 3 immunoassays were validated for use with baboon plasma before conduction of the study. Assay validation consisted of spike and recovery experiments at various concentrations in pooled baboon maternal plasma (Figure 1). These assays employ the quantitative sandwich immunoassay technique. Briefly, recombinant sTNFα, sTNFRI, and sTNFRII standards and baboon plasma specimens were incubated in duplicate wells of the microtiter plates precoated with monoclonal antibodies specific to the soluble TNFα and TNF receptors. During this incubation, the immobilized antibodies in the microtiter plates bound the specific TNFα and receptor antigen present in the standards and samples to form antigen–antibody complexes. After washing away the inbound substances, affinity-purified polyclonal antibodies to sTNFα, sTNFRI, and sTNFRII conjugated to an enzyme were added to the assay wells. Following an incubation period, the assay plates were washed repeatedly to remove the unbound antibody enzyme reagents. This step was immediately followed by the addition of a substrate solution (tetramethylbenzidine) to each well. Color developed in the assay plates was proportional to the amount of sTNFα, sTNFRI, and sTNFRII bound in the initial step. Color development was stopped after a defined period and the microtiter plates were read using a programmable spectrophotometer (Ceres 900 Micro plate Workstation; BIO-Tek Instruments, Winooski, Vermont). The reproducibility of the immunoassays in baboon plasma was calculated; inter- and intracoefficient of variants were less than 10%. The volume of plasma used for analysis was 0.5 mL. All samples were analyzed in 1 batch.
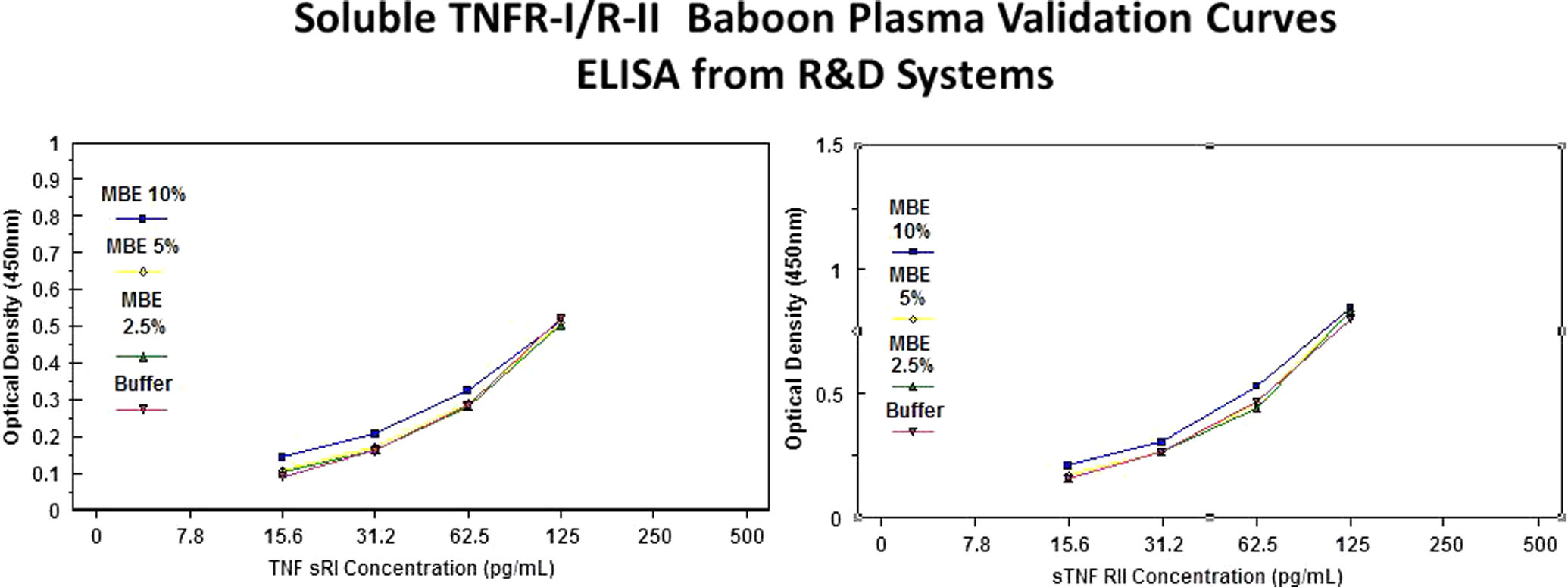
Spike and recovery experiments conducted in the absence (0%) and presence of varying concentrations (2.5%-10%) of normal pooled baboon maternal plasma at 4 different concentrations of serum levels of tumor necrosis factor α binding protein 1 (sTNFRI) and sTNFRII (enzyme-linked immunosorbent assay [ELISA], R&D System, Minneapolis, Minnesota). The validation curves worked well. Parallel curves indicate that there were no significant matrix effects. Baboon plasma TNFRI and TNFRII can be detected using human sTNFRI/RII immunoassays from R&D System even after 1:10 dilutions.
Statistical Analysis
The plasma concentrations of TNFα, TNFRI, and TNFRII in maternal blood samples obtained early in pregnancy, prior to surgery, and after surgery as well as in umbilical cord blood samples were measured and differences between the concentrations were determined using Wilcoxon signed ranks test. Pearson correlation coefficient was used to determine the possible relationships between the concentrations of sTNFα, sTNFRI, and sTNFRII in maternal and fetal plasma. A P value <.05 was considered significant.
Results
All newborns were vigorous and healthy at birth according to institutionally derived standards. We performed no necropsy studies. Of all, 5 of the newborns were female and 4 were males. Dr Wolf decided based on maternal behaviors that 4 newborns should be nursery reared while 5 could be reared by the mothers. All mothers—5 with their infants—returned to their colonies and outside corrals a few days after surgery. The TNFα, TNFRI, and TNFRII concentrations were measured in the 34 plasma samples. No significant correlation between maternal and cord plasma concentrations of TNFα, TNFRI, or TNFRII was observed.
The median concentration of sTNFα, sTNFRI, and sTNFRII expressed in pg/mL with interquartile ranges is shown in Table 1. Four relevant observations were noted, first, the maternal plasma concentration of TNFα and TNFRI were higher at the end of pregnancy than early in pregnancy; second, the maternal plasma TNFRII concentration and the TNFα/TNFRI ratio were stable throughout gestation; third, maternal plasma concentration of TNFα, TNFRI, and TNFRII before and after the delivery of the placenta were similar; fourth, there was a higher TNFRI in fetal circulation (627 pg/mL) compared to maternal circulation (227 pg/mL) with a TNFα/TNFRI ratio that was more than 3 times lower in fetal than in maternal circulation (0.025 in maternal plasma vs 0.009 in fetal plasma). In addition, the Wilcoxon test helped us to observe a tendency to a significant difference (ie, almost 300%/s, P= .07). An increase in the sample size would certainly reduce the Wilcoxon test values obtained.
The TNFα, TNFRI, and TNFRII in Maternal and Fetal Circulation Prior to the Onset of Labora
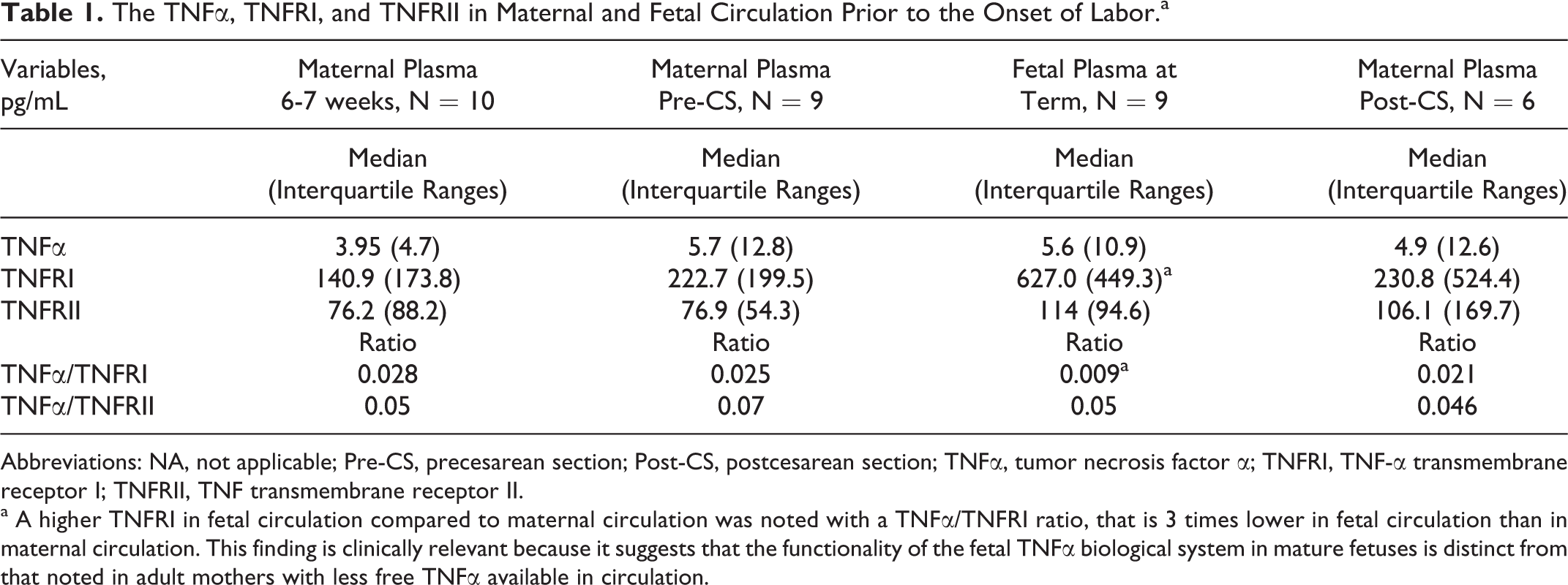
Abbreviations: NA, not applicable; Pre-CS, precesarean section; Post-CS, postcesarean section; TNFα, tumor necrosis factor α; TNFRI, TNF-α transmembrane receptor I; TNFRII, TNF transmembrane receptor II.
a A higher TNFRI in fetal circulation compared to maternal circulation was noted with a TNFα/TNFRI ratio, that is 3 times lower in fetal circulation than in maternal circulation. This finding is clinically relevant because it suggests that the functionality of the fetal TNFα biological system in mature fetuses is distinct from that noted in adult mothers with less free TNFα available in circulation.
Comment
The TNFα has bioactive actions in most cells and can cause tissue organization or tissue destruction depending on the time of action, cellular context, and the induced inflammatory stimulus. 45 We present an observational study in which high-sensitivity enzyme-linked immunosorbent assays were used to measure sTNFα, sTNFRI, and sTNFRII in a controlled experimental fashion. We compared the maternal and fetal baseline operational capacities to mount a TNFα inflammatory response in uncomplicated pregnancies avoiding the stress of labor. Our 4 principal observations are that (1) there is no correlation between the maternal and fetal plasma concentrations of TNFα, TNFRI, or TNFRII suggesting that in uncomplicated pregnancies the fetal and maternal circulatory TNFα biological systems are independent of each other; (2) maternal plasma concentration of TNFα and TNFRI are higher at the end of pregnancy suggesting that pregnancy is a “controlled proinflammatory state” because the TNFα/TNFRI ratio remained relatively stable throughout gestation; (3) the maternal plasma concentrations of TNFα, TNFRI, and TNFRII before and after the delivery of the placenta were similar suggesting that in uncomplicated pregnancies at term, the contribution of the placenta to the maternal plasma concentration of TNFα, TNFRI, and TNFRII must be small; (4) the functionality of the fetal TNFα biological system in circulation prior to delivery is distinct and independent of the mother; the fetus is perhaps more capable of blocking the systemic TNFα inflammatory responses that may occur during the labor process. Indeed, we found 3 times lower TNFα/TNFRI ratio in fetal plasma than in maternal plasma suggesting an enhanced natural inhibition of TNFα in the circulation of mature fetuses when compared to their mothers.
We are inclined to hypothesize on the reason for the presence of a repressive environment to the action of TNFα in fetal circulation prior to the onset of labor. Our observations together with other lines of reported evidences suggest that the developmentally regulated inflammatory process that allows for early placentation and placental morphological changes to meet fetal demands is different from the systemic inflammatory process occurring during parturition. In the latter, TNFα induces trophoblastic cyclooxygenase 2 expression, 46,47 decreases PG15-hydroxy dehydrogenase 48 leading to increasing PG levels, 49 and increases the expression of matrix metalloproteinase 2 (MMP-2) and MMP-9 50 that degrades the extracellular matrix of the chorio-amniotic membranes. In addition, the production of placenta corticotrophin-releasing hormone is also stimulated by TNFα 51 while the placental 11β-hydroxysteroid dehydrogenase which converts cortisol into its inactive derivate cortisone is inhibited resulting in a net TNFα-dependent elevation of cortisol that may trigger uterine contractions and labor. 52 However, the beneficial effects of induced inflammation during parturition could also predispose to neonatal injury. The differential regulation of the TNFα biological system in fetal and maternal circulation at the end of pregnancy may be guided to protect the fetus from the inflammatory response that is essential for the mechanisms of labor to proceed in the mother. Future studies in this experimental model could address this hypothesis. On the other hand, presently, the proteolitic enzyme that cleaves the transmembrane TNFRII to release sTNFRII has been identified 3 but not the one that allows for shedding of the sTNFRI and this matter could be one of the keys to the understanding of the regulation of this potent biological system in perinatal medicine. 11
Limitations of the Study
The small sample limits the statistical power of the study making our results preliminary observations. By design and because of the additional experimental challenges, we did not examine the effects of spontaneous labor directly on the fetus. Similarly, the mechanisms responsible for fetal protection against the maternal TNFα inflammatory process remain unknown. Factors that could be involved in deregulation include the duration of labor, microbial invasion of the amniotic cavity, and gestational age. This experimental method could be used to address these questions and to model therapeutic interventions aimed at preventing or ameliorating the excessive TNFα-dependent stimuli that has been linked to neonatal sepsis, long-term pulmonary disease, and chronic cerebral degeneration. 41,42,52 –59
In conclusion, we present a method to study the pathophysiology of systemic TNFα inflammatory responses in perinatal medicine. Our observations from a limited number of nonhuman primates suggest that mature fetuses have an independent and distinct baseline capacity to modulate TNFα actions in circulation when compared to their mothers.
Footnotes
Acknowledgments
The authors are grateful to Dr Sam Edwin and staff at his Wayne State University laboratory for validating and performing the immunoassays.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: in part by the