
Abstract
The mesenchymal stem cells (MSCs) have awakened interest in regenerative medicine due to its high capability to proliferate and differentiate in multiple specialized lineages under defined conditions. The reproductive system is considered a valuable source of MSCs, which needs further investigations. Many factors have been reported as critical for these cell lineage specification and determination. In this review, we discuss the main effects of extracellular matrix or tissue environment and growth factors in the cell lineage commitment, including the reproductive stem cells. The MSCs responses to culture medium stimuli or to soluble factors probably occur through several intracellular activation pathways. However, the molecular mechanisms in which the cells respond to these mechanical or chemical perturbations remain elusive. Recent findings suggest a synergic effect of microenvironment and soluble cell culture factors affecting cell differentiation. For future applications in cell therapy, protocols of reproductive MSCs differentiation must be established.
Introduction
A considerable increase in chronic degenerative diseases has been recorded in the last decades, accounting for more than 50% of deaths worldwide. 1 Several studies on stem cells and their potential for tissue regeneration have presented good results encouraging the development of therapies to save lives. Reproductive diseases are inserted in this context as a target of many recent researches. 2 –10 Several studies investigate the role of mesenchymal stem cells (MSCs) in endometriosis. 2 –4,7 Bone marrow-derived MSCs contribute to endometrial growth and regeneration and play a role in endometriosis progression. 4 Participation of stem cells in infertility, 5,6 cancer 7 and other uterine disorders and the possibility of it use for future therapies are also focused by some researches. 4,10
Stem cells are commonly defined by their capability for extensive self-renewal and to differentiate into one or more specialized cell types. 11 –16 Based on the tissue of origin and potential to differentiate into one or more specific types of mature cells, two major groups are distinguished: (1) embryonic stem cells: pluripotent stem cells that can be isolated from the inner cell mass of preimplantation embryos, 14,17,18 and (2) adult stem cells: derived from fetal or adult tissues from specific organs.
The MSCs are pluripotent adult somatic stem cells derived from bone marrow 11,19 and also from wide variety of organs, 14,18,20,21 including in the reproductive system, like endometrium, 3,4,5,22,23 ovarians 6 and also placenta. 13
Unlike what was thought for many years, recent studies reported that MSCs can differentiate into various specific lineages, including adipocytes, chondrocytes, osteocytes, myocytes, astrocytes, neurons, 24 –28 vascular endothelial cells, 19,29 –31 hepatocytic lineage, 32,33 lung, gut, skin epithelia, 14,34,35 and endometrial cells. 3,36
The ability of MSCs to propagate and to differentiate in specific cell-matured phenotypes under defined culture conditions is termed “plasticity.” 19 ,37 Several studies describing MSCs differentiation in vitro and in vivo have been published and in most of them these cells were isolated from bone marrow aspirates, expanded in culture with high efficiency, and induced to differentiate to multiple lineages under controlled protocols. 19,21,31 –33,35,38 –40
As advantage of these cells, the MSCs can be used in the same patient (autograft), reducing the possibility of immune rejection. Moreover, differently from embryonic pluripotent stem cells, these cells can be used with minor ethical implications.
Satisfactory results have been achieved in some clinical trials using rodent and nonrodent mammal MSCs. 26 Mcbeath and coworkers 41 studied the differentiation potential of MSCs to specific connective tissue cells, specifically bone and adipose tissue, whereas Grassel and Ahmed 42 had worked with condrogenesis. Morelli et al 34 showed the differentiation in vitro of MSCs derived from human endometrium in smooth muscle cells, adipocytes, osteoblasts, 36 and chondrocytes. 34,36 These findings suggests that the uterus is an alternative source of mesenchymal stem cells. 4 The isolation of MSCs in the reproductive system clarifies the mechanism of reproductive diseases like endometriosis. 3
Both, in vivo and in vitro plasticity of MSCs greatly depends on the microenvironment. 39 –42 Moreover, it is also known that, in the absence of the native soluble and cell-contact signaling network like the bone marrow environment, reduced MSCs plasticity and proliferation capacity are observed on in vitro cultures. Such findings indicate that soluble growth factors, paracrine, autocrine or humoral signaling, and extracellular matrix microenvironment interact synergistically to regulate cell proliferation and lineage specification and also to maintain the differentiation state of MSCs. 20,39 However, the molecular mechanisms underlying the commitment of MSCs to a specific lineage demands more studies. Here we discuss the main factors that are supposed to be critical for stem cells differentiation process, including the reproductive cells, specifically the effects of matrix or tissue microenvironment and soluble growth factors.
The ECM or Tissue Microenvironment Directs MSCs Lineage Specification
It has already been shown that MSCs are frequently recruited to the sites of tissue injury or inflammation and differentiate into multiple specific lineages, 27,35,43 but the mechanisms underlying this process are not clear. The ECM elasticity or stiffness seems to be vital for cell lineage specification.
According to the matrix elasticity, tissues can be classified into 3 distinct groups: (1) softer tissues as the brain, (2) stiff tissues as the muscles, and (3) rigid tissues as the bones. 44 –46 It is believed that the great variation in tissue or ECM microenvironment provides specific conditions that guide cell differentiation. In this context, the molecular mechanism underlying the MSCs interaction with the tissue or matrix and how it will transduce those stimuli in cell morphological modification are not yet well understood. 45
Studies have been carried out mostly evaluating the contribution of ECM microenvironment in chondrogenesis, osteogenesis, and adipogenesis, 41 ,45 –48 and in cancer development. 49 In addition, Masuda et al 2 studied the effects of local environment in the endometrial regeneration by stem cells.
It has been hypothesized that during cell–matrix interaction, specific stimuli are produced and are transduced in chemical signaling events for distinct phenotype expression. 45,50 –52 In their niche or within tissue, cells are exposed to mechanical forces. When cells are submitted to these forces, a variety of physiological modifications occur such as cell motility, proliferation, and differentiation. 53 The mechanisms by which the cells recognize these mechanical perturbations and transduce them in specific chemical signaling remain obscure. 50,52 It is known that the matrix stiffness or elasticity plays a key role in cell lineage commitment 32,47,48 and influences focal-adhesion structure and the cytoskeleton. 45
The development of extracellular matrix models with variable degrees of elasticity based on inert polyacrylamide gels mimicking the consistency of the different tissues is necessary in order to evaluate the differentiation of MSCs. 45 They observed that in soft matrices, that mimics brain tissue, the MSCs showed a neuronal phenotype, in intermediate stiffness matrices that mimics striated muscle, the same cells differentiated into myoblast, and in rigid matrices resembling cartilage or bone, the cells became osteoblasts. This study provided great evidence that the matrix composition can drive cell differentiation. 45
Two key events are present in the described processes, the ability of the MSCs to recognize the extracellular matrix forces and, second, the mechanic stimuli transduction to generate specific signals that will drive the differentiation. 45 Multiple signaling pathways are activated in response to mechanical force stimulation. McBeath et al 41 observed that both matrix stiffness and soluble factors modulate MSC-specific lineage commitment via RhoA signaling and Rho-kinase (ROCK) activity, which regulates the actin–myosin contractility. It has been shown that inhibition of cytoplasmic myosin II using blebbistatin disrupts the influence of matrix elasticity in cell differentiation. 45,46 These findings suggested that during force transduction, specific signaling pathways are activated including Rho guanosine triphosphatase, mitogen-activated protein kinase, tyrosine kinases/phosphatases, 49,52,54 which can alter gene expression and promote cell differentiation. 41,45,46,55,56
Comparative studies of microenvironment and growth factors’ influence in MSCs differentiation were performed. Davis 56 investigating the influence of both bone morphogenetic protein 2 and three-dimension (3D) osteoconductive substrates in osteogenesis, observed enhanced effect in osteogenic response of MSCs due to multiple pathway activation mediated on both substrate and growth factor.
The ECM microenvironment seems to be more selective than other factors and can alter the phenotype in precommitted cells. 45 Although satisfactory in vitro results has been achieved in vitro, many molecular issues underlying the cell commitment must be clarified for future application in vivo.
Another key point in cell investigation is related to cell medium for culture. Many conclusions on ECMs culture influencing cell lineage commitment derive from studies on culture of cells in two dimension (2D) surfaces. A different behavior have been described when the cell culture is done on 3D approaches. 46,53,57,58 Notwithstanding, it was shown that cell types derived from in vivo settings quickly lose their differentiated phenotype when are plated onto 2D medium. 58 Besides, in vitro differentiation studies relies on biological intervention, like special media but scaffolds are important to avoid possible host reactions in vivo. 59
Recently, Jurgens 60 induced chondrogenesis in a collagen type II hydrogels mimicking the joint 3D microenvironment, Schneider et al 44 induced tenogenic differentiation in 3D high-density microenvironment, and Mohr et al 13 produced an osteogenic graft in a chorion-derived scaffold from MSC derived from human placenta. Together, these results demonstrated that the ECM stiffness or elasticity play a key role in cell differentiation. 46,47,49
The Effect of Soluble Growth Factors in Cell Differentiation
As was reported in several studies in vitro, the cell differentiation involves numerous extra and intracellular signaling pathways modulated by either cell–cell or cell–matrix contact as well as soluble growth factors. 2 The growth factors are peptides that can be produced by the target cell (autocrine) or released through the plasma membrane of adjacent cells (paracrine), modulating cellular activity. 61
The effects of several different growth factors in MSC lineage specification was extensively studied in vitro in chondrogenesis, 62 –64 osteogenesis, 64 –68 and adipogenesis. 69 –71 Many growth factors promoting MSCs proliferation and differentiation have been described, but the exact molecular mechanisms by which they act is still unclear. To better understand these mechanisms, it is first necessary to identify the factors that promote proliferation, differentiation, or retain the differentiation state of MSCs. 24
The influence of growth factor in cell differentiation varies not only in stem cell type but also in the strain that is sought. Many factors have been described as MSC differentiation inducers including platelet-derived growth factor (PDGF), basic fibroblast growth factor (bFGF), transforming growth factor (TGF) β, epidermal growth factor (EGF), and insulin-like growth factor (IGF). 27,72 –74 These factors were previously described by Kuznetsov et al 75 as essential for bone marrow stromal fibroblast colony formation in vitro. Beyond the cell differentiation, they also modulate the angiogenesis, ECM composition, chemotaxis of osteoprogenitor cells, and granulation or scar tissue deposition. 65,74 Recent studies showed the importance of stromal-epithelial cell interactions in the endometrial epithelial stem cell niche. They identified several growth factors, such as EGF or TGF-α or PDGF-BB , and fibroblast feeder, layers to establish clonal growth. 76,77
Subsequent studies 62 –65,68 –71 have found wide variation in MSCs response to growth factors in different cell lineage specification. Although the endometrial differentiation has been already described, 3,78 the factors involved in this process need to be further studied. Different factors can be required for cell specification depending on the stimuli (Table 1).
Wide Variation of Growth Factors Involved in MSCs Differentiation. Growth Factors Involved in MSCs Differentiation.a
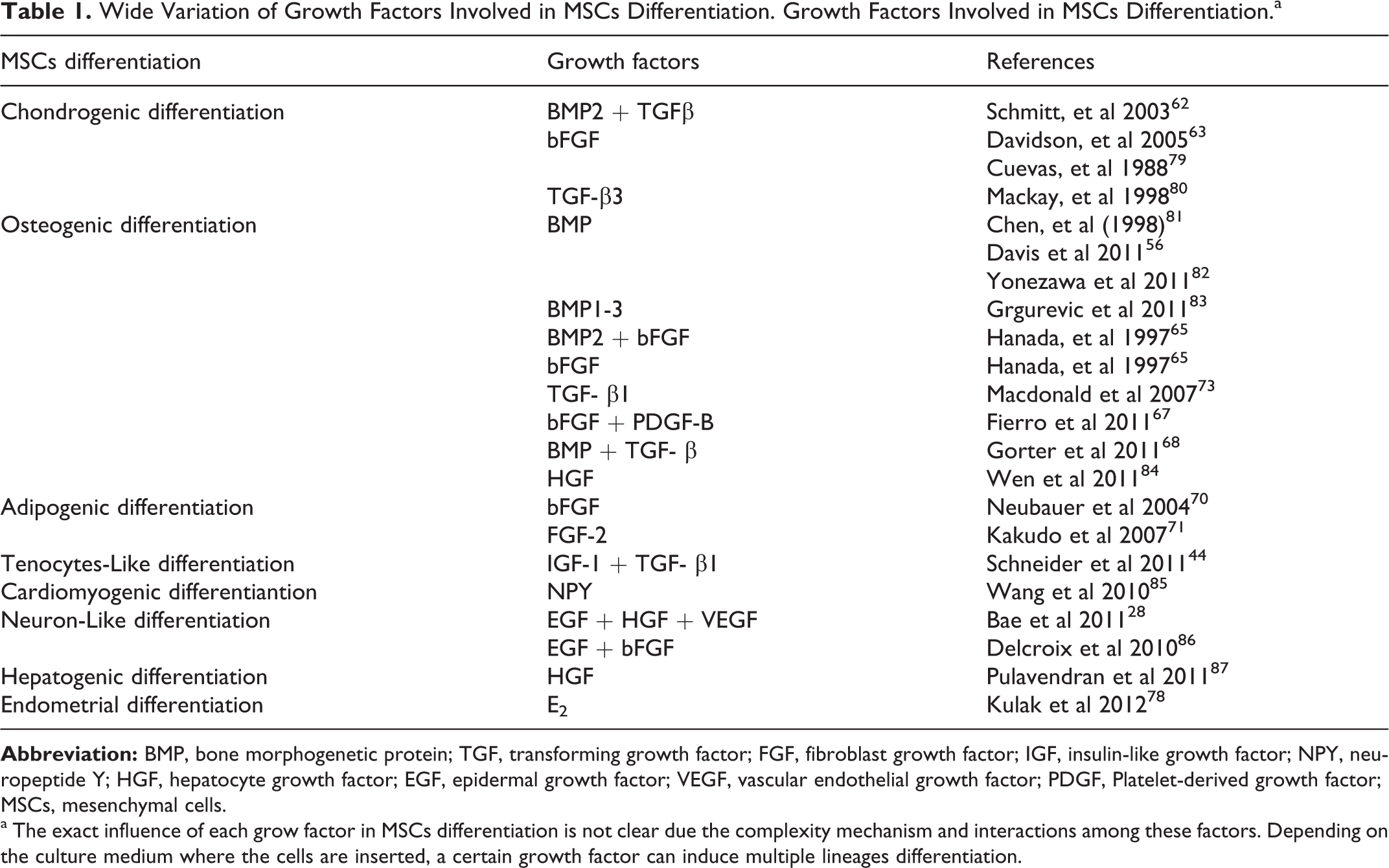
a The exact influence of each grow factor in MSCs differentiation is not clear due the complexity mechanism and interactions among these factors. Depending on the culture medium where the cells are inserted, a certain growth factor can induce multiple lineages differentiation.
The exact effect of each growth factor in cell differentiation is not clear because the mechanism underlying this process is complex. For example, it was observed in vitro that depending on the culture medium where the cells are inserted, a certain growth factor can induce chondrogenic, osteogenic, or adipogenic differentiation. It has been shown that FGFs are crucial for chondrogenesis, 63,79 adypogenesis, 70,71 and osteogenesis. 16 On the other hand, chondrogenic differentiation was observed to be inducible by bone morphogenetic protein (BMP) 62,64 or TGF and dexametazone. 80 It was believed that BMPs was only fundamental for upregulation of cartilage and bone phenotype expression. However, it have been demonstrated that BMP are also implicated in several cell type grow and specification. Specific lineages were obtained in several conditioned medium with BMPs including, bone, cartilage, fat, and nervous cells. 62 Interactions between one or more factors result in enhanced effect in MSCs differentiation. Hanada et al 65 and Deans and Moseley 24 reported that bFGF and BMP-2 synergistically enhanced the osteogenic differentiation of MSCs in vivo.
Basically, the growth factors interact with distinct cell receptors activating different intracellular signaling pathways which turn on transcriptional factors for specific gene expression depending on the stimuli.
Chen et al 81 reported that selective activation of BMP receptor type 1B (BMPR-1B) result in differentiation of MSC to osteoblastic lineage, whereas activating the BMBR-1A these cells differentiate into adipocyte lineage. This latter information was suggestive that the overexpression or loss of the receptors may be important in determining the response of MSCs to growth factor stimulation. 24 Most recently, Lin and Hankenson 16 studied the interactions of BMP, Wnt, Notch, hedgehog, and FGF signaling pathways in osteoblastogenesis. In all signaling pathways, the transcriptional factor Runx2 is activated and seems to play a key role in osteogenic differentiation. However, the more precise molecular understanding of these complex interactions is recommended.
Conclusions
Several studies have been carried out aiming to determine the exact mechanism by which the cell lineage specification occurs. However, few studies have been carried out regarding reproductive system stem cells, and no conclusive data were achieved. Recent studies refer the synergic effects of ECM compliance and soluble cell culture factors in cell differentiation. Comparative studies of microenvironment and growth factors influence in MSCs differentiation were performed. The MSCs’ differentiation depends on the environment where they are inserted and growth factor’s interaction as well as on the intensity and duration of produced stimulation (Figure 1). The molecular mechanism underlying ECM and growth factor’s interactions and cell specification must be well investigated.
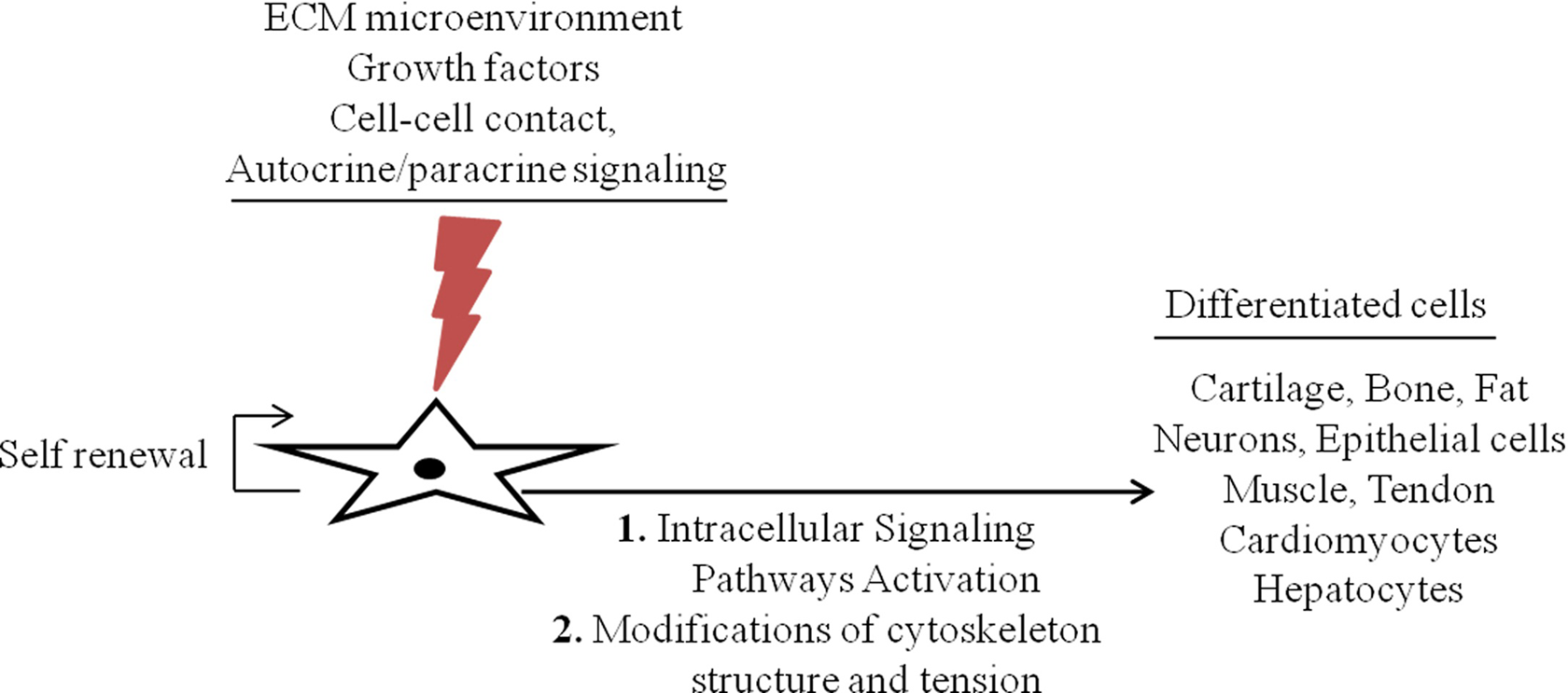
Schematic representation of the general mechanism of mesenchymal stem cell (MSC) differentiation mediated by matrix microenvironment and growth factors. These factors interact synergistically to regulate cell proliferation and cell differentiation, although the effects of extracellular matrix (ECM) seems to be crucial where the cells are inserted and more selective than others.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.