
Abstract
Introduction:
In all, 10% to 20% of all pregnant women smoke despite intentions to quit. Smoking cessation drugs such as nicotine replacement therapy (NRT) and bupropion are recommended for pregnant women. Our observation that developmental exposure to nicotine adversely affects metabolic and reproductive outcomes in rats has raised concerns about NRT’s safety during pregnancy. Conversely, the effect of bupropion has not been reported.
Objective:
The goal of this study was to examine the effect of fetal and neonatal exposure to bupropion on postnatal metabolic and reproductive outcomes.
Methods:
Dams (N = 5/group) were exposed to saline or bupropion (5 or 10 mg/kg per d) for 2 weeks prior to mating until weaning. We assessed weight, adiposity, and glucose homeostasis in all offspring until 26 weeks of age. Onset of puberty, fertility, and pregnancy outcomes in the female offspring were also assessed.
Results:
Fetal and neonatal exposure to bupropion did not cause metabolic derangement in the offspring despite a significant decrease in birth weight in the offspring of dams treated with 10/mg/kg per d bupropion (5.9 ± 0.2 g vs control 6.7 ± 0.2 g; P = .02). Moreover, with the exception of accelerated pubertal onset in F1 and F2 offspring, bupropion administration to pregnant dams had no impact on fertility or pregnancy outcomes for either the dam or the female offspring.
Conclusion:
Fetal and neonatal exposure to the smoking cessation drug bupropion, unlike NRT, does not appear to adversely affect metabolic outcomes or the fertility of the female offspring. However, bupropion does appear to alter pubertal onset through an as yet unknown mechanism.
Introduction
Maternal cigarette smoking is associated with a higher risk of poor pregnancy outcomes including spontaneous abortion, premature delivery, and fetal death, yet 10% to 20% of all women smoke while pregnant. 1 –3 Although many women express a desire to refrain from smoking during their pregnancy, the highly addictive nature of nicotine makes smoking cessation difficult. 4 Indeed most smokers who attempt to quit without pharmacologic smoking cessation aids are unsuccessful. 5 There are 3 smoking cessation drugs approved for use in North America, sustained release bupropion (Zyban®), nicotine replacement therapy (NRT), and varenicline (Champix®). Of these, NRT is generally recommended as the first-line pharmacotherapy for pregnant women who cannot quit smoking by other means. 6 However, the use of NRT during pregnancy continues to be controversial as animal models have clearly demonstrated that nicotine can have toxic effects on multiple organ systems in the fetus. 7 –9 To date there is limited clinical evidence available regarding the safety of bupropion or varenicline use during pregnancy. 10 –12 Despite this, studies have reported that 0.3% to 3% of pregnant smokers take bupropion at some point during their pregnancy. 13,14 Importantly, bupropion, under the trade name Wellbutrin®, is also the nonselective serotonin reuptake inhibitor antidepressant most commonly used during pregnancy, with 0.7% of all pregnant women reporting bupropion use during pregnancy. 15,16
In nonpregnant smokers, bupropion is highly effective for smoking cessation when compared to either placebo or NRT. 17,18 Similarly, a small cohort study reported a significantly higher rate of cessation in pregnant women taking bupropion versus those not on smoking cessation therapy. 19 Bupropion is a dopamine/norepinephrine reuptake inhibitor but its ability to reduce cravings and alleviate withdrawal symptoms may be due, in part, to the fact that bupropion also acts as a noncompetitive antagonist at the nicotinic acetylcholine receptor (nAChR). 20,21 Animal studies have reported adverse metabolic and reproductive health outcomes following fetal and/or neonatal exposure to nicotine; the effects are proposed to occur via nAChR-mediated pathways. 8,22 Therefore, we hypothesized that exposure to bupropion, a nAChR antagonist, during fetal and neonatal life would not cause the same metabolic and reproductive deficits that have been reported in nicotine-exposed animals.
Materials and Methods
Maintenance and Treatment of Animals
All animal experiments were approved by the Animal Research Ethics Board at McMaster University, in accordance with the guidelines of the Canadian Council for Animal Care. Nulliparous 200 to 250 g female Wistar rats (Harlan, Indianapolis, Indiana) were maintained under controlled lighting (12:12 L: D) and temperature (22°C) with ad libitum access to food and water. Two weeks prior to mating the dams were randomly assigned to receive saline or bupropion hydrochloride. Dams were injected with 5 or 10 mg/kg per d bupropion hydrochloride (BUP5, BUP10; Toronto Research Chemicals, North York, ON) or saline (N = 5) subcutaneously for 14 days prior to mating and during pregnancy until weaning (postnatal day 21; PND21). In rats, the 10 mg/kg dose yields maximum plasma levels of bupropion (ie, C max) of 96.4 ng/mL; reported C max values for bupropion in human pharmacokinetic studies range from 72 to 105 ng/mL. 23 –25 Maternal body weight and food consumption were monitored biweekly for the duration of the study. Two weeks after the initiation of treatment, dams were mated (1:1) with age-matched Wistar rats and were monitored daily for confirmation of breeding (ie, the presence of sperm in a vaginal flush). The day that a positive sign of copulation was observed was designated as gestational day 0 (GD0). The dams were allowed to deliver normally. For each dam, time to pregnancy, gestation length, litter size, birth weight, sex, and the number of stillbirths were recorded. From these data the mating success rate ([number of females mated/number of females cohabited] × 100), pregnancy success rate ([number of dams delivering a litter/number dams with a confirmed mating] × 100), the live birth index ([number of live offspring/number of offspring delivered] × 100), and the sex ratio (number of male offspring/number of female offspring) were calculated. At birth (PND1), litters were culled to 8 preferentially selecting for equal number of male and female offspring to ensure uniformity of litter size between treated and control litters. Pups were weighed weekly during lactation, and at 3 weeks of age (PND21) 1 male and 1 female pup per litter was euthanized by CO2 asphyxiation. Pancreas tissue and fat pads (mesenteric and perirenal) were collected, weighed, and processed for subsequent analysis. After weaning, we assessed metabolic outcomes in male and female offspring and fertility of the female offspring as described below.
Postnatal Weight Gain and Adiposity
Pups were weighed weekly from weaning until 26 weeks of age to assess postnatal growth. Food consumption was determined biweekly from 7 weeks of age until the end of the study. At 3, 7, and 26 weeks of age, 1 male and 1 female offspring from each litter were euthanized, and perirenal, mesenteric, and gonadal fat pads were collected to assess postnatal adiposity.
Glycemic Control
Glucose tolerance was investigated in bupropion-exposed and control rats at 2, 4, and 6 months of age (N = 5 per sex per group) using sequential intraperitoneal (ip) glucose tolerance tests (GTT). To avoid litter effects, only 1 animal from a single litter was tested. After an overnight fast, glucose was measured using the NovaMax glucometer (Nova Diabetes Care, Inc, Billerica, Massachusetts) in saphenous vein samples, collected by repeated puncture, at baseline (09:00 hours), 15, 30, 60, and 120 minutes after rats were given 2 g/kg glucose (Sigma-Aldrich, St Louis, Missouri) in water by ip injection. The total glucose response following the GTT was summarized as the area under the curve (AUC) calculated using the trapezoidal rule. Additional blood was taken at baseline for the determination of fasting serum insulin and the calculation of the homeostasis model of assessment of insulin resistance (HOMA-IR). Blood samples were allowed to clot at 4°C, centrifuged, and stored at −80°C until assayed. Serum insulin concentrations were measured by an ultra sensitive rat insulin enzyme-linked immunosorbent assay (Crystal Chem Inc., Downers Grove, Illinois). The homeostatic model assessment-insulin resistance (HOMA-IR) was calculated using HOMA calculator version 2.2.2 (http://www.dtu.ox.ac.uk). The IR was also evaluated by sequential insulin tolerance tests (ITTs) at 4 and 6 months of age. For the ITT, following an overnight fast, rats were given 1 IU/kg insulin (Novolinge Toronto, human biosynthetic insulin, Novo Nordisk, Mississauga, Ontario) in saline by subcutaneous injection. Glucose was measured using the NovaMax glucometer in saphenous vein samples, collected by repeated puncture, at baseline, 20, 40, and 60 minutes.
β-Cell Mass
All these analyses were conducted by a single researcher who was blinded to the treatment group. Immunohistochemistry and morphometric analyses were used to assess the effects of bupropion on β-cell mass at PND21; the time at which maximal exposure to the drug has occurred. Pancreata were weighed, fixed by immersion in 10% neutral-buffered formalin (EM Science, Gibbstown, New Jersey), and embedded in paraffin. Immunohistochemical detection of insulin was performed on 5 μm serial sections, separated by an average of 30 μm, for all groups (5 sections per animal; 5 animals per sex per group; maximum of 1 animal per litter). Tissue sections were deparaffinized in xylene, rehydrated, and washed in phosphate-buffered saline (PBS). Endogenous peroxidase activity was quenched in methanol, followed by antigen retrieval in 10 mmol/L citrate buffer (pH 3.0) and incubation with 10% normal goat serum and 1% bovine serum albumin (BSA). Sections were then incubated with the primary antibody, a polyclonal, guinea pig anti-swine insulin antibody (1:150 dilution; DakoCytomation, Carpinteria, California) which has been shown by the manufacturer to cross-react with rat insulin peptides, overnight at 4°C. Sections were then washed in PBS, and immunostaining was identified using the Vectastain kit (Vector Laboratories, Burlinghame, California) with diaminobenzadine as the chromogen. Tissue sections were counterstained, dehydrated, and mounted with Permount (Fisher Scientific, Fair Lawn, New Jersey). Control sections were incubated with 1% BSA in PBS in place of the primary antibody. All measurements were performed at ×10 magnification, and the whole pancreas was analyzed. Immunopositive cells were identified and the β-cell area was calculated as a percentage of the total area of the section using integrated morphometry software (Metamorph, Universal Imaging Corp, Downington, Pennsylvania). At least 25 fields were counted per animal. β-Cell mass was determined by quantifying the β-cell area as a percentage of the total pancreatic area and then multiplying the total pancreatic weight by this percentage.
Pubertal Onset and Fertility of F1 Offspring
Pubertal onset (ie, vaginal opening) was determined in control and bupropion-exposed offspring (F1), and fertility in these offspring was assessed at 6 months of age. Female offspring (F1; N = 5 per group) were housed 1:1 with an unexposed proven male and monitored daily for confirmation of breeding (ie, the presence of sperm in the vaginal swab). The day that a positive sign of copulation was observed was designated GD0. For each dam (F1), time to pregnancy (ie, days until detection of sperm in the vaginal swab), gestation length, litter size, birth weight, sex, and the number of stillbirths were recorded. Mating and pregnancy success rates, live birth index, sex ratio, survival to PND4, survival to weaning, and time of vaginal opening of the female (F2) offspring were also determined.
Statistical Analysis
All statistical analyses were performed using SigmaStat (v.3.1, SPSS, Chicago, Illinois). The results are expressed as mean ± standard error of the mean. For pregnancy and neonatal outcomes the litter was considered the statistical unit. β-Cell mass at PND21 was analyzed using 2-way analysis of variance (ANOVA) with sex and treatment as the independent variables. For all postnatal testing after weaning, results for male and female offspring were analyzed separately by using 1-way ANOVA (α = 0.05) followed by post hoc comparisons when significance was indicated. Data were checked for normality and equal variance, when data failed normality or equal variance test, data were reanalyzed using appropriate nonparametric tests.
Results
Pregnancy and Birth Outcomes: P Generation
There was no effect of either dose of bupropion on mating or pregnancy success, time to pregnancy, gestation length, the live birth index, litter size, sex ratio, survival to PND4, or survival to weaning (Table 1). However, birth weight of pups exposed to the highest dose of bupropion (ie, BUP10) was significantly reduced relative to controls, an effect that was not due to altered maternal food consumption or weight gain during pregnancy (Table 1).
Pregnancy and Neonatal Outcomes: P Generation.a
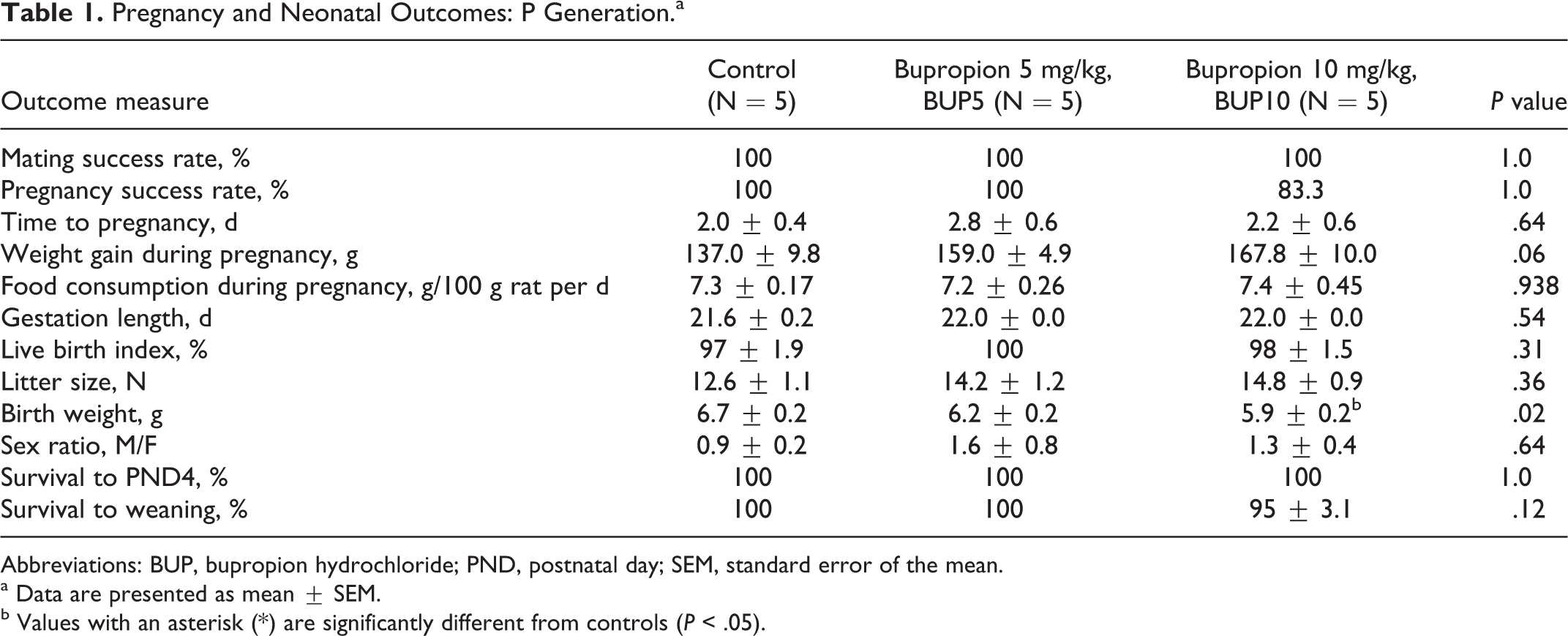
Abbreviations: BUP, bupropion hydrochloride; PND, postnatal day; SEM, standard error of the mean.
a Data are presented as mean ± SEM.
b Values with an asterisk (*) are significantly different from controls (P < .05).
Postnatal Weight Gain and Adiposity
There was no effect of bupropion on body weight, fat pad weight (absolute or relative), or total percentage of body fat in either male or female offspring at 3, 7 (all P > .05; data not shown), or 26 weeks of age (all P > .05; Supplemental Tables 1 and 2).
Glycemic Control
Fetal and neonatal exposure to bupropion did not significantly alter the fasting glucose levels, the total response to a glucose challenge (AUC glucose), fasting insulin, or HOMA-IR in either male or female offspring at 3, 7 (all P > .05; data not shown), or 26 weeks of age (all P > .05; Supplemental Tables 3 and 4). Similarly, there was no effect of perinatal exposure to bupropion in response to the ITT at any age examined (Supplemental Figure 1).
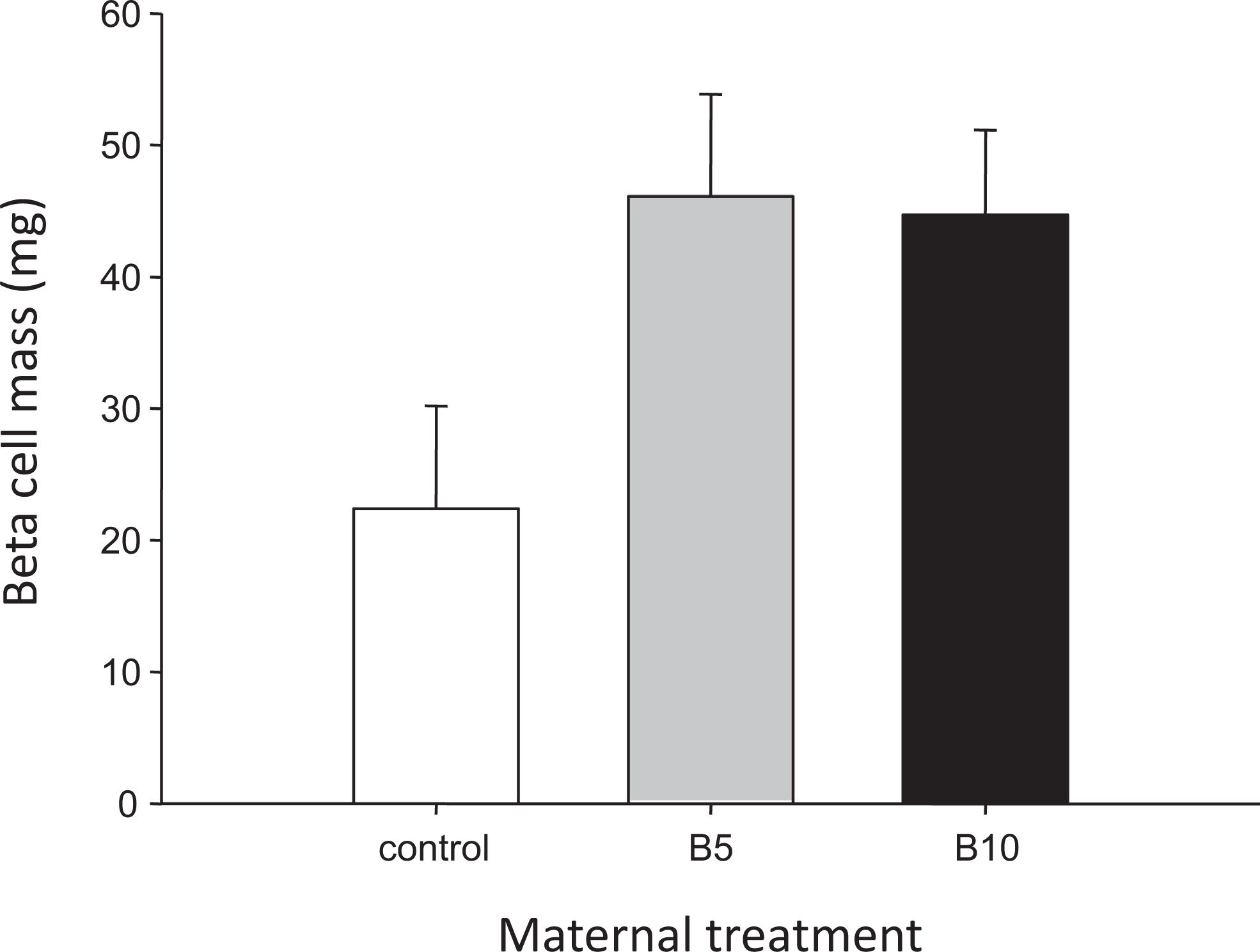
Pancreatic β-cell mass (mg) in postnatal day 21 (PND21) offspring of control and bupropion-exposed dams (B5 = bupropion 5 mg/kg per d; B10 = bupropion 10 mg/kg per d). Data are presented as mean ± standard error of the mean (SEM).
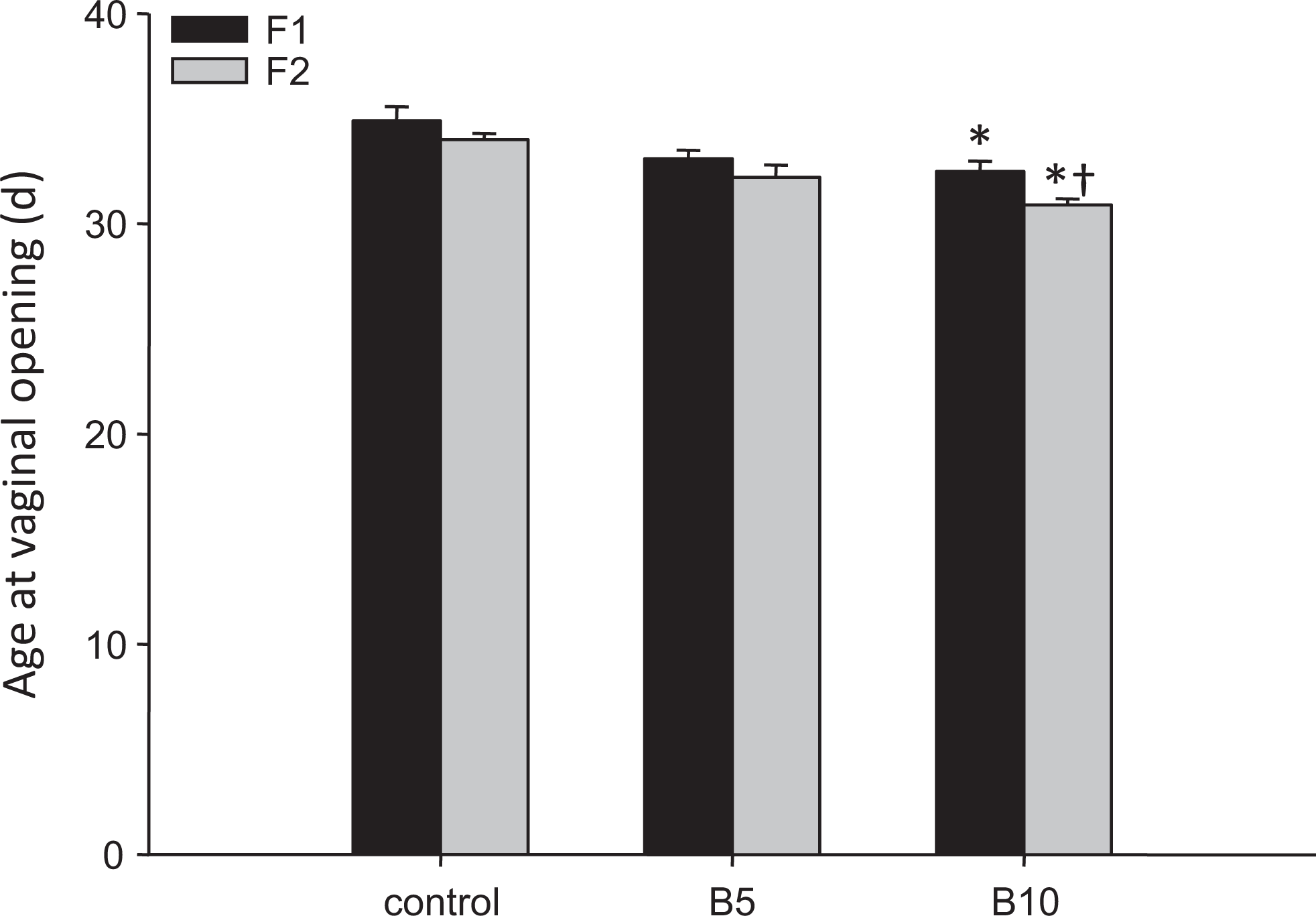
Age at vaginal opening F1 and F2 offspring. Data are presented as mean ± standard error of the mean (SEM). Data with an asterisk (*) are significantly different from control animals of the same generation, data marked with † are significantly different from their parents (ie, F1 generation).
β-Cell Mass
There was no main effect of either sex (P = .233) or bupropion exposure (P = .070) on β-cell mass and no significant interaction effect (P = .137; Figure 1).
Pubertal Onset and Fertility of F1 Offspring
Bupropion exposure during pregnancy resulted in advanced vaginal opening (ie, earlier pubertal onset) in the female F1 offspring but only at the highest dose tested (Figure 2). At 6 months of age there was no effect of fetal and neonatal exposure to bupropion on time to pregnancy, gestation length, mating or pregnancy success, the live birth index, litter size, birth weight, sex ratio, or postnatal survival to either PND4 or weaning (Supplemental Table 5). Interestingly, the F2 offspring of females exposed to 10 mg/kg bupropion in utero and during lactation also had an earlier onset of puberty relative to control animals (Figure 2). Moreover, the onset of puberty in the F2 BUP10 offspring was significantly earlier than in the F1 BUP10 females (F1 32.5 ± 0.5 vs F2 30.9 ± 0.3 days; P = .017), an effect only seen in this group.
Discussion
Cigarette smoking during pregnancy is associated with an increased risk of adverse health outcomes for the offspring both in the short (ie, intrauterine growth restriction, prematurity, fetal, and neonatal death) and long term (ie, increased risk for obesity, cognitive and behavioral abnormalities, and respiratory disorders). 1,2,9,26 Therefore, it is universally agreed that smoking cessation during pregnancy has significant health benefits for both the mother and her child. Although smoking cessation counseling is recommended as the first-line intervention for pregnant smokers many women continue to smoke. 27 For women who do not respond to counseling, clinical practice guidelines recommend that pharmacological interventions be considered for smoking cessation. 6
Of the 3 approved smoking cessation drugs (ie, NRT, bupropion, and varenicline), NRT is most commonly recommended for use during pregnancy. 13 However, there are still safety concerns related to the use of NRT during pregnancy and lactation. 7,28 In addition to the well-known consequences of fetal and neonatal exposure to nicotine on central nervous system development and long-term neurocognitive outcomes, more recently results from animal studies have demonstrated that perinatal exposure to nicotine results in metabolic abnormalities and reduced fertility in the offspring. 8,22,29 –31 The adverse effects of nicotine are thought to occur via the activation of nAChR present in fetal tissues. Indeed, the nAChR subunits α2-α4, α6, α7, and β2-β4 are present in the pancreas and ovary at birth (PND1 and nAChR subunits have been identified in pancreatic beta cells and adipocytes. 32 –35 Since bupropion exerts its actions by blocking activation of nAChRs, 20,21 we hypothesized that bupropion exposure during fetal and neonatal life would not cause the same metabolic and reproductive deficits that have been reported in nicotine-exposed animals.
Fetal and neonatal exposure to 10 mg/kg per d of bupropion resulted in a significant reduction in birth weight but had no impact on any other pregnancy or neonatal outcomes. This reduction in fetal growth may be attributable, at least in part, to reduced uterine blood flow, an outcome that has been reported in women taking bupropion during pregnancy. 36 Postnatally, there was no effect of bupropion on food consumption or body weight in either the mothers or their offspring despite the fact that bupropion treatment has been reported to cause decreased food consumption in nonpregnant rats and result in weight loss in humans. 37,38 The lack of any effect of developmental exposure to bupropion on pancreatic β-cell mass or postnatal glycemic control is interesting given that bupropion treatment caused cellular stress and an ablation of glucose-stimulated insulin secretion in vitro in a rat β-cell line. 39 However, the concentration of bupropion used in this study (ie, 1 μmol/L) is likely higher than the fetal exposure to the drug. Indeed, in rats the reported maximum plasma concentration of bupropion following a single 10 mg/kg ip injection is 96.4 ng/mL (ie, 350 nmol/L); of this, estimates of transplacental transfer suggest that only 20% of the drug enters the fetal circulation. 23,40 Therefore, higher doses of bupropion may have negative effects on pancreatic development or function. Although in our study fetal and neonatal exposure to bupropion did not result in any significant metabolic deficits in the offspring, it did have implications for the reproductive health of the female offspring.
The female offspring of dams exposed to 10 mg/kg per d bupropion (F1) and their daughters (F2) had earlier vaginal opening (ie, precocious puberty) relative to control offspring. The observed acceleration of puberty in bupropion-exposed offspring may be due to its action as an inhibitor of the dopaminetransporter (DAT). 20 Genetic ablation of the DAT results in increased extracellular dopamine levels; a similar effect is seen following the administration of bupropion to nonpregnant rats. 41 –43 This elevation in brain dopamine levels may negatively impact maternal care. Indeed, significant deficits in maternal care in DAT knockout mice have been noted; behaviors that, in rats, have been shown to result in accelerated puberty in the female offspring. 44,45 The decreased time to puberty in the BUP10 F2 offspring was unexpected; however, rat pups that have received impaired maternal care themselves show similar deficits in maternal behaviors that may explain the earlier pubertal onset in the F2 generation. 46 However, whether or not bupropion exposure alters maternal care remains to be determined.
In summary, we have demonstrated that, in rats, although the highest dose of bupropion did cause a significant reduction in birth weight, fetal and neonatal exposure to bupropion does not cause significant metabolic derangement in the offspring. Moreover, with the exception of accelerated pubertal onset in F1 and F2 offspring, bupropion administration to pregnant dams has no impact on fertility or pregnancy outcomes for either the dam or her female offspring. Results from this study suggest that bupropion exposure during pregnancy may have few adverse metabolic and reproductive consequences for the offspring. However, although these results in an animal model are reassuring, further clinical studies are required to evaluate the safety and efficacy of bupropion for smoking cessation during pregnancy.
Footnotes
Acknowledgments
We thank Bryce Poirier and the staff of the McMaster University Central Animal Facility for their help with the animal work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: an operating grant from the
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.