
Abstract
Objective:
This study examines whether silencing specific β-nerve growth factor small interfering RNA (β-NGF siRNA) can affect the growth of ectopic endometriotic implants, generalized hyperalgesia, and nerve fiber density in endometriosis.
Methods:
Four specific β-NGF siRNAs were detected by Western blot analysis, and the most efficient specific siRNA was transferred into rats with surgically induced endometriosis through gene transfer. The length × width × height of each ectopic transplant that survived from 2 groups were measured at pre-and postbombardment after 2 weeks. The transplants were collected 2 weeks after bombardment. Warm-water tail flick test was performed before the rats were sacrificed. The specimens were sectioned and stained immunohistochemically with antibodies against the types of nerve fibers to compare the presence of different nerve fibers in the treatment and control groups. The serums and supernatants of the peritoneal washings in the treatment and control groups were collected for enzyme-linked immunosorbent assay (ELISA) analysis. The extra rats were successfully induced with endometriosis and through gene transfer as described above. The spherical volumes of the transplants and tail flick latency post-bombardment after 4, 6, 8, and 10 weeks were measured.
Results:
The spherical volumes in the treatment group were much smaller than those in the control group, and tail flick latency significantly increased in the treatment group postbombardment after 2 weeks. The ELISA analysis showed that the concentrations of β-NGF in the serums and supernatants of the peritoneal fluid decreased in the treatment group unlike in the control group. Less sympathetic and sensory innervation was observed in the treatment group postbombardment after 2 weeks. The outcomes of the spherical volumes of the transplants and tail flick latency postbombardment after 4, 6, 8, and 10 weeks showed that the sizes of the transplants did not return to their previous size and that the treatment had some effects on generalized hyperalgesia.
Conclusion:
Specific siRNA-mediated silencing of the β-NGF gene expression after gene transfer suppressed the growth of ectopic endometriotic implants resulted in a significant improvement in generalized hyperalgesia as well as reduced sympathetic and sensory nerve fiber density in the treatment group.
Introduction
Endometriosis, the presence of endometrial tissues outside the uterus, is a common and often debilitating disease of women in the reproductive age. This disease is frequently associated with significant pain and has been recognized as a major cause of infertility among women in many industrialized nations. Although clinicians and surgeons have struggled to establish appropriate treatment strategies, present medical therapies for endometriosis are frequently inadequate. Many women ultimately face the prospect of multiple surgeries or hysterectomy. 1,2
The pathogenesis of endometriosis and the pathophysiological basis for endometriosis-associated pain are not well understood. Endometriosis is believed to be a chronic inflammatory state with disturbances in both cell-mediated and humoral immunity. 3 The high concentration of cytokines and prostaglandins found in the peritoneal fluid of patients with endometriosis may contribute to an increase in nociceptive stimuli on nerve fibers. 4 Proinflammatory cytokines contribute to the development of inflammatory pain and hyperalgesia 5,6 and may be involved in endometriosis pathophysiology. Several studies demonstrated an increased expression of the nerve growth factor (NGF) in eutopic and ectopic endometrium in endometriosis. 7 –9 The NGF is not only essential for the survival of small nociceptive sensory neurons and sympathetic neurons 10 but is also a key mediator of pain and inflammation. 11,12 The NGF regulates immune and inflammatory responses through direct and/or indirect effects on immune competent cells. 13 Moreover, NGF can directly modulate the function of nociceptive sensory neurons. The NGF administration causes hyperalgesia in humans and rodents, 14 and NGF inhibitors mitigate pain induced by inflammation in cutaneous and visceral tissues. 15 Sevcik et al 16 showed that anti-NGF therapy profoundly reduces bone cancer pain in rat models. However, knowledge on the application of anti-NGF to endometriosis is currently limited.
RNA interference is a sequence-specific posttranscriptional gene silencing process initiated by double-stranded RNA. 17 RNA interference can silence a specific cellular gene and provide a powerful reverse genetic approach to analyze gene functions both in vitro and in vivo. 18 The most widely utilized nucleic acid–based sequence-specific gene-silencing molecules at present are small interfering RNAs 19 called siRNAs, which consist of symmetrical duplexes with 19 to 21 base pairs. 20 The siRNA method can inhibit the target gene expression with specificity, efficiency, and endurance. 21
Several researchers have identified nerve fibers in the endometriotic lesions of women with endometriosis. 9,22,23 Tokushige et al 24 demonstrated the presence of multiple, small unmyelinated nerve fibers in the peritoneal endometriotic lesions of women with confirmed endometriosis; the density of the nerve fibers in the peritoneal endometriotic lesions was much greater than that in the peritoneum of women without endometriosis. Berkley et al 8,25 reported that endometriotic implants develop a sensory and sympathetic nerve supply both in rats and in women similar to that of a healthy rat uterus. The present study demonstrated the existence of a much greater density of nerve fibers in deep infiltrating endometriosis than in peritoneal endometriotic lesions. 26 These results indicate that nerve fibers in endometriotic lesions play an important role in the pathogenesis of endometriosis.
We applied specific NGF siRNA to a rat model with surgically induced endometriosis to investigate further the influence of siRNA on nerve fibers and pathogenesis of endometriosis.
Materials and Methods
Establishment of Animal Models
All animal procedures were conducted in accordance with the institutional guidelines of Qilu Hospital of Shandong University (Shandong, P. R. China). Thirty adult female Wistar rats weighing 180 to 210 g were housed in cages in an air-conditioned room at 25°C ± 1°C in a 12-hour dark–light cycle. All animals had free access to water and laboratory food. The estrous stage was monitored daily by vaginal smear every morning, beginning at least 2 weeks before surgery and continued until the day of sacrifice. Only rats with regular 4-day cycles both before and after surgery were utilized. Three rats were excluded for irregular estrous stage. A rat model was surgically induced with endometriosis as previously described. 25 Surgery was performed under aseptic precautions. Rats in the estrous stage were anesthetized with 3% pelltobarbitalum natricum (Solarbio; Beijing Solarbio Science & Technology Co, Ltd, China) at a dose of 0.2 mL per 200 g by intraperitoneal injection. A midline abdominal incision exposed the uterus. A 1 cm segment of the middle of the left uterine horn was removed and placed in warm sterile saline. Four pieces of uterine horn (∼2 × 2 mm) were cut from this segment and sewn with 4.0 nylon sutures around alternate cascade mesenteric arteries that supply the caudal small intestine. Sewing started from the cecum. The incision was closed in layers, and the rats were allowed to recover from the anesthesia under close observation. Three of the rats died because of complications related to surgery. The remaining rats were then randomly divided into 2 groups (12 rats per group), namely, the treatment group and the control group.
Specific siRNAs
The complementary DNA sequence of the β-NGF gene was obtained from Genbank (M27617.1). Four specific siRNAs against rat β-NGF were designed and synthesized (Shanghai GenePharma Co, Ltd, China) to interfere with the β-NGF expression. The 4 groups, including PGPU6/GFP/Neo-NGF-rat-307, PGPU6/GFP/Neo-NGF-rat-63, PGPU6/GFP/Neo-NGF-rat-251, and PGPU6/GFP/Neo-NGF-rat-1070, were designated as groups 1, 2, 3, and 4, respectively. The targeting and nucleotide sequences of the 4 groups were as follows: group 1, GCAGAACCGTACACAGATAGC, 5’-CACCGCAGAACCGTACAGATAGCTTCAAGAGAGCTATCTGTGTACGGTTCTGCTTTTTTG-3’ (sense), and 5’-GATCCAAAAAAGCAGAACCGTACACAGATAGCTCTCTTGAAGCTATCTGTGTACGGTTCTGC-3’ (anti-sense); group 2, GCGCTTCAAGTCAGTGAAACT, 5’-CACCGCGCTTCAAGTCAGTGAAACTTTCAAGAGAAGTTTCACTGACTTGAAGCGCTTTTTTG-3’ (sense), and 5’-GATCCAAAAAAGCGCTTCAAGTCAGTGAAACTTCTCTTGAAAGTTTCACTGACTTGAAGCGC-3’ (anti-sense); group 3, GCGTAATGTCCATGTTGTTCT, 5’-CACCGCGTAATGTCCATGTTGTTCTTTCAAGAAGAACAACATGGACATTACGCTTTTTTG-3’ (sense), 5’-GATCCAAAAAAGCGTAATGTCCATGTTGTTCTTCTCTTGAAAGAACAACATGGACATTACGC-3’ (anti-sense); and group 4, GGACTGCATGATAATTTATCG, 5’-CACCGGACTGCATGATAATTTATCGTTCAAGAGACGATAAATTATCATGCAGTCCTTTTTTG-3’ (sense), 5’-GATCCAAAAAAGCACTGCATGATAATTTATCGTCTCTTGAACGATAAATTATCATGCAGTCC-3’ (anti-sense). PGPU6/GFP/Neo was utilized as mock plasmid for negative control. The targeting sequence was GTTCTCCGAACGTGTCACGT.
Cell Culture and Transfection
Michigan Cancer Foundation’s 7 (MCF-7), which normally produce NGF proteins, 27,28 were cultivated in 25 mm2 flasks with a high-glucose Dulbecco-modified Eagle medium (Gibco-BRL, Grand Island, New York) and supplemented with 10% fetal calf serum (Gibco-BRL) and antibiotics containing 100 µg/mL of streptomycin (Gibco-BRL). The cultures were maintained at 37°C in a humidified atmosphere with 5% CO2. The media were changed every 2 days. When the cells reached 60% to 70% confluency, the 4 specific siRNAs were transfected into the culture cells with Lipofectamine 2000 (Invitrogen, Grand Island, New York) according to the manufacturer’s instructions. The MCF-7 cells without specific siRNA transfection served as positive control. The cells were washed with pre-cooled phosphate-buffered saline and then harvested for Western blot assay after transfection for 48 hours.
Western Blot Analysis
The proteins were extracted from the transfected cells with a protein extraction kit (Solarbio; Beijing Solarbio Science & Technology Co. Ltd.) according to the manufacturer’s directions.
Equal amounts of proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene difluoride membranes (Millipore, Billerica, Massachusetts). The membranes were blocked in 5% nonfat powdered milk in Tris-buffered saline tween (TBST) for 1 hour at room temperature and then stained overnight at 4°C with rabbit anti-NGF antibody (Abcam, Cambridge, UK; dilution 1: 1000) and mouse anti-β-actin monoclonal antibody (Boster, China, dilution 1:1000). After rinsing 3 times with TBST, the membranes were incubated at room temperature for 1 hour with a horseradish peroxidase–conjugated secondary antibody (Zhongshan Golden Bridge Biotecnology Co, Ltd, China; dilution 1:5000). Immunoblots were developed with ECL Plus Western Blot Detection reagents (Millipore) and exposed to an X-ray film (Kodak, Rochester, New York) from 30 seconds to 1 minute to allow optimal detection (all bands were visible but not overexposed). All samples were tested in triplicate. Western blot densitometric analysis was performed with the Image G analysis software. The band densities of NGF were normalized to the densities of β-actin.
Gene Transfer
The maximum interfering effectiveness of the specific siRNA-coated plasmid was screened out by Western blot from the 4 specific groups. Figure 1 shows that the β-NGF-specific siRNAs effectively inhibited protein expression. The protein levels of most of the specific siRNAs decreased and remained visible compared with the positive control. The most efficient specific siRNA was group 3 (F = 26.12; P < .001). Hence, we selected group 3 to perform the siRNA interfering experiment.
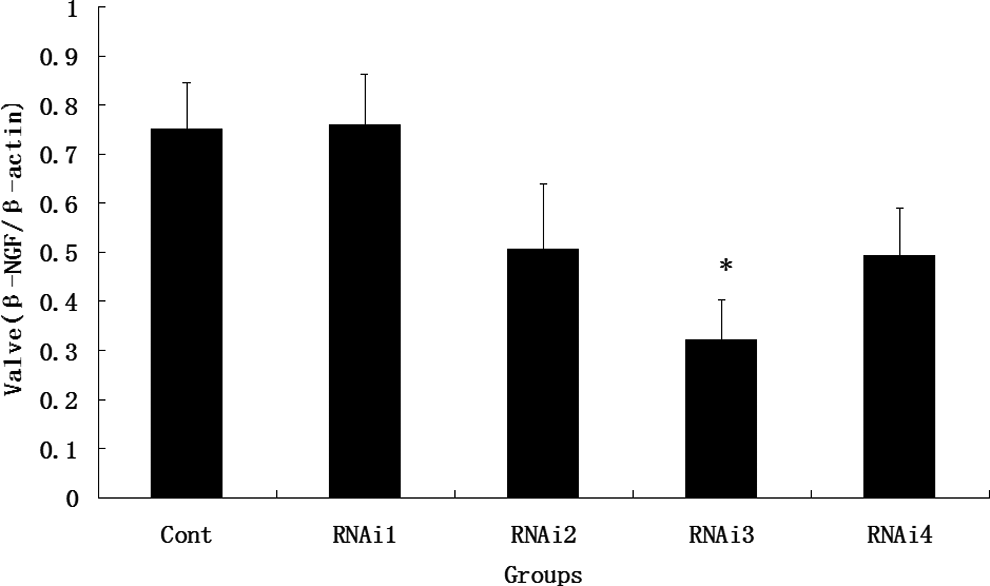
The β-NGF Western blotting analysis after siRNA transfection. The MCF-7 cells without specific siRNA transfection served as a positive control, namely, cont group. RNAi1, RNAi2, RNAi3, and RNAi4 represented group 1, group 2, group 3, and group 4, respectively (*P < .001). β-NGF indicates β-nerve growth factor; siRNA, small interfering RNA; MCF-7, Michigan Cancer Foundation’s 7.
Gold microparticles (25 mg) were placed in a centrifugation tube and shaken with 100 μL 0.05 mol/L spermidine. Specific siRNA-coated plasmid 1 μg/μL 50 μL was precipitated with 100 μL 1 mol/L CaCl2. The mixture was centrifuged and precipitated, and the supernatant was removed. After rinsing with dehydrated alcohol for 3 times, 200 μL of dehydrated alcohol were supplemented to prepare a DNA/gold microparticle suspension at a ratio of 1 μg DNA and 0.5 mg gold microparticles. The gold microparticles were placed in a sample tube, prepared in a gene gun bullet, and then preserved at 4°C. The microparticles of the mock plasmid were considered the control group as previously mentioned.
The endometriotic implants were exposed through intraperitoneal injection of anesthesia 2 weeks after laparotomies were performed on the rat models with surgically induced endometriosis. All rats were successfully induced (at least 1 from the 4 pieces of uterine horn that survived). The success rate of the uterine horn transplant was 55%, and the length × width × height of each surviving transplant were measured. The spherical volume of each surviving transplant was calculated through prolate ellipsoid formula V (mm3) = 0.52 × length × width × height. 29 When the implants that survived were valued, the researchers did not consider the group to which the implants belonged. Gene transfer into the implants for the treatment group was performed with a portable gene gun (ST-500; &Ningbo Xinzhi Biological Polytron Technologies Inc, China). The gene gun parameters (pressure, particle size or number, and distance) were optimized based on the instrument's manual. Each implant was subjected to 2 bombardments of bullets with a distance of 2 cm (each bullet contained 1 μg of plasmid and 0.5 mg of gold microparticles/2 cm2). Each implant in the control group was subjected to 2 bombardments of bullets containing mock plasmid microparticles. All the rats survived and were sacrificed by overanesthesia 2 weeks after the bombardments. Warmwater tail flick test was performed before the rats were sacrificed through the method proposed by Lashbrook et al. 30 Each rat was kept quiet in a special fixed case for 30 minutes, and the tail was immersed in warm water (temperature was fixed at 50°C ± 1°C). The time course between the immersion and flicking of the rat tail from the water surface was recorded as rat tail flick latency to assess endometriosis-associated pain. When tail flick latency was valued, the researchers did not consider the group to which the rats belonged. The measurements were performed manually. Each rat was kept quiet for 1 minute after the test and was tested thrice. The length × width × height of each ectopic transplant from the 2 groups were measured again, and the spherical volume of each ectopic transplant was calculated as mentioned previously. The specimens were collected and fixed in 10% neutral-buffered formalin for 18 to 24 hours. Blood samples and peritoneal washings were obtained from all the rats. Saline (2 mL) was instilled in the peritoneal cavity and then the abdomen was massaged for 5 minutes. Lastly, the peritoneal washing was retracted at the base of the abdominal cavity. 31 After centrifugation, the serums and supernatants of the peritoneal washings in the treatment and control groups were preserved at -80°C.
The rats with regular 4-day cycles were then randomly divided into 2 groups (32 rats per group), namely, the treatment group and the control group. The following procedures were performed as mentioned previously. The rats (8 rats per group) were sacrificed postbombardment after 4, 6, 8, and 10 weeks. Warmwater tail flick test was performed. The length × width × height of each surviving transplant were measured, and the spherical volume of each ectopic transplant was calculated again. None of the rats died of complications related to surgery.
Immunohistochemistry
The implants were fixed with formalin, processed, and embedded in paraffin according to standard protocol. The section was cut at 4 μm in each specimen and mounted onto slides. These sections were routinely stained with hematoxylin and eosin. The antigen retrieval for immunohistochemistry was performed in boiling citrate buffer (0.01 mol/L, pH 6.0) for 15 minutes with a microwave oven. Endogenous peroxidase activity was prevented by incubating in 0.3% hydrogen peroxide for 15 minutes. Nonspecific binding was blocked by 10% goat serum (Zhongshan Golden Bridge Biotecnology Co, Ltd) for 20 minutes at room temperature. The sections were immunostained overnight at 4°C with antibodies for monoclonal mouse anti-NF (dilution 1:150; Abcam), which is a highly specific marker for myelinated nerve fibers; polyclonal rabbit anti-NGF (dilution 1:200; Abcam); monoclonal mouse anti-NGFRp75 (dilution 1:200; Abcam); polyclonal rabbit anti-TrkA (dilution 1:500; Abcam); polyclonal rabbit anti–vasoactive intestinal polypeptide (VIP dilution 1:1000; Abcam), which is a specific marker for parasympathetic neurons; polyclonal rabbit anti-NPY (dilution 1:1000; Abcam), which is a specific marker for sympathetic neurons; polyclonal mouse anti-SP (dilution 1:250); polyclonal rabbit anti-CGRP (dilution 1:300; Abcam), which are sensory fiber markers that can be present in both Aδ and C nerve fibers; and monoclonal rabbit anti–growth associated protein 43 (GAP-43; dilution1:100; Abcam), which is a marker for new neurite outgrowth and regeneration. The slides were washed and incubated with horseradish peroxidase–conjugated secondary antibody at room temperature for 30 minutes. Peroxidase activity was visualized by exposure to a diaminobenzidine tetrahydrochloride solution (DAB kit; Zhongshan Golden Bridge Biotechnology Co, Ltd) for 3 to 5 minutes. The sections were then washed, counterstained with hematoxylin for 1 minute, dehydrated, and mounted with coverslips. Normal rat skin was utilized as positive control because it contains myelinated and unmyelinated nerve fibers expressing neurofilament (NF), NGF, NGF receptor p75 (NGFRp75), tyrosine kinase receptor-A (Trk-A), substance P (SP), calcitonin gene-related peptide (CGRP), VIP, neuropeptide Y (NPY), and GAP-43. We utilized the functional layer of the eutopic endometrium from the rat without endometriosis as negative control because of the absence of nerve fibers.
The β-NGF Enzyme-Linked Immunosorbent Assay
The β-NGF concentrations in the serums and supernatants of the peritoneal washings of both the treatment and control groups were measured by an enzyme-linked immunosorbent assay (ELISA) kit. This technique utilizes a goat anti-rat-β-NGF for capture antibody (R&D Systems, Minneapolis, Minnesota), a goat anti-rat-β-NGF antibody for detection (R&D Systems), streptavidin horseradish peroxidase-conjugated rabbit anti-goat antibody (R&D Systems), and a substrate solution (Zhongshan Golden Bridge Biotechnology Co, Ltd) in accordance with the manufacturer’s instructions. Optical density was measured at 450 nm, and NGF concentrations were extrapolated from a standard curve by recombinant rat β-NGF.
Quantification of the Nerve Fiber Density
The images were captured with an Olympus DP72 camera (Tokyo, Japan). The assessment of nerve fiber mean density was performed with the Image Pro Plus software (Media Cybernetics, Maryland). The sections were analyzed and the entire section was evaluated after immunostaining. The integrated optical density (IOD) and area of the images were calculated with the Image Pro Plus software. The area was then divided by IOD to obtain the mean density of the nerve fiber. All lighting conditions and magnifications were maintained constant. The procedure was performed twice by 2 independent observers (each blinded to the other) without any knowledge of the experimental groups from which the slices were obtained. The concordance rate between the 2 observers was more than 95%.
Statistical Analysis
The results were expressed as mean ± SD. All analyses were performed with the Statistical Package for Social Sciences (version 17.0). The comparison between the 2 groups was performed through nonparametric 2-tailed t test (Mann-Whitney test). One-way analysis of variance and Student Newman-Keuls test were conducted to compare the data of the multiple groups in the Western blot analysis. Statistical significance was defined as P value of less than .05.
Results
Spherical Volumes Pre- and Postbombardment After 2 Weeks in Rats With Induced Endometriosis and Tail Flick Latency Postbombardment After 2 Weeks
The endometriotic lesions in the rats were successfully induced through the protocol described previously in the study (Figure 2). The ectopic growths were embedded in a small amount of fat and connective tissue. When dissected freely, the growths appeared as oval fluid-filled cysts, each still attached to the blood vessel to which it had been sewn. The starting volumes of the control and treatment groups at prebombardment were evaluated, and no statistically significant difference was found between the 2 groups. The spherical volumes of the treatment group were smaller than those of the control group at postbombardment (*P < .05; Figure 3A). All rats were subjected to warmwater tail flick test. A significant difference existed in the tail flick latencies of the 2 groups (*P < .05; Figure 3B).
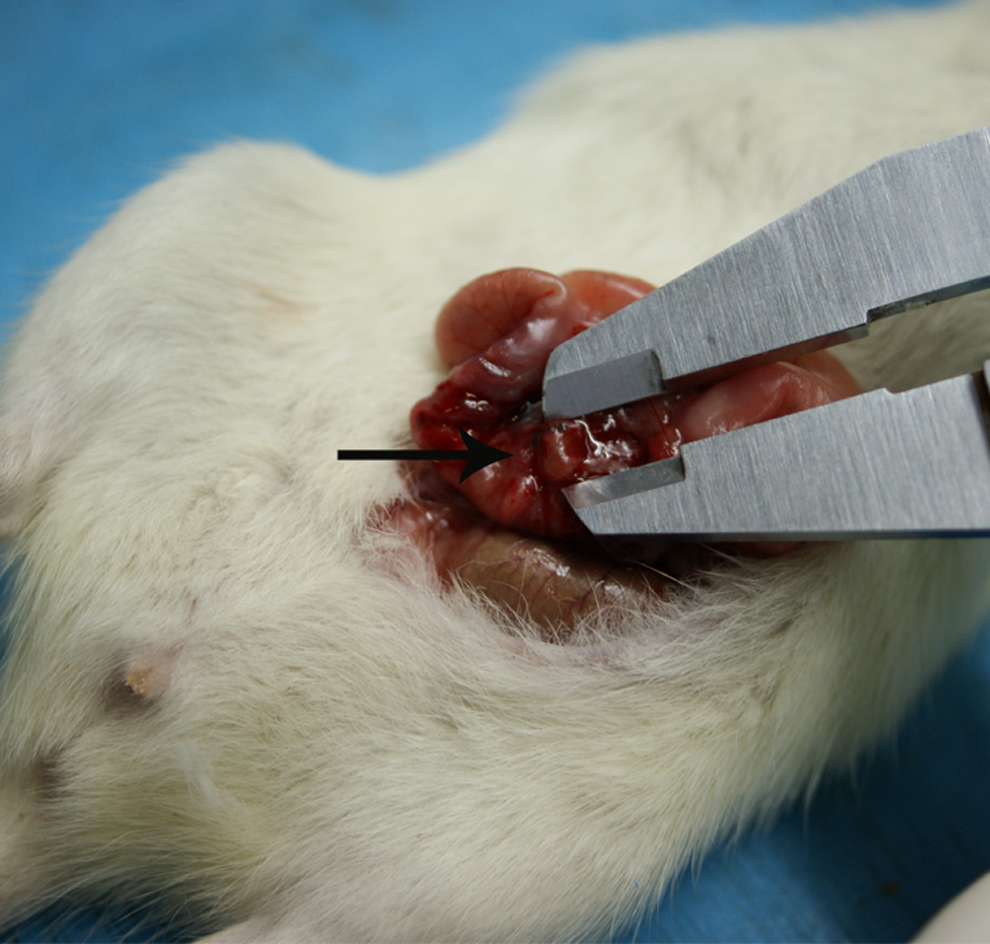
Morphologic appearance of induced endometriotic lesions (pointed by the arrow).
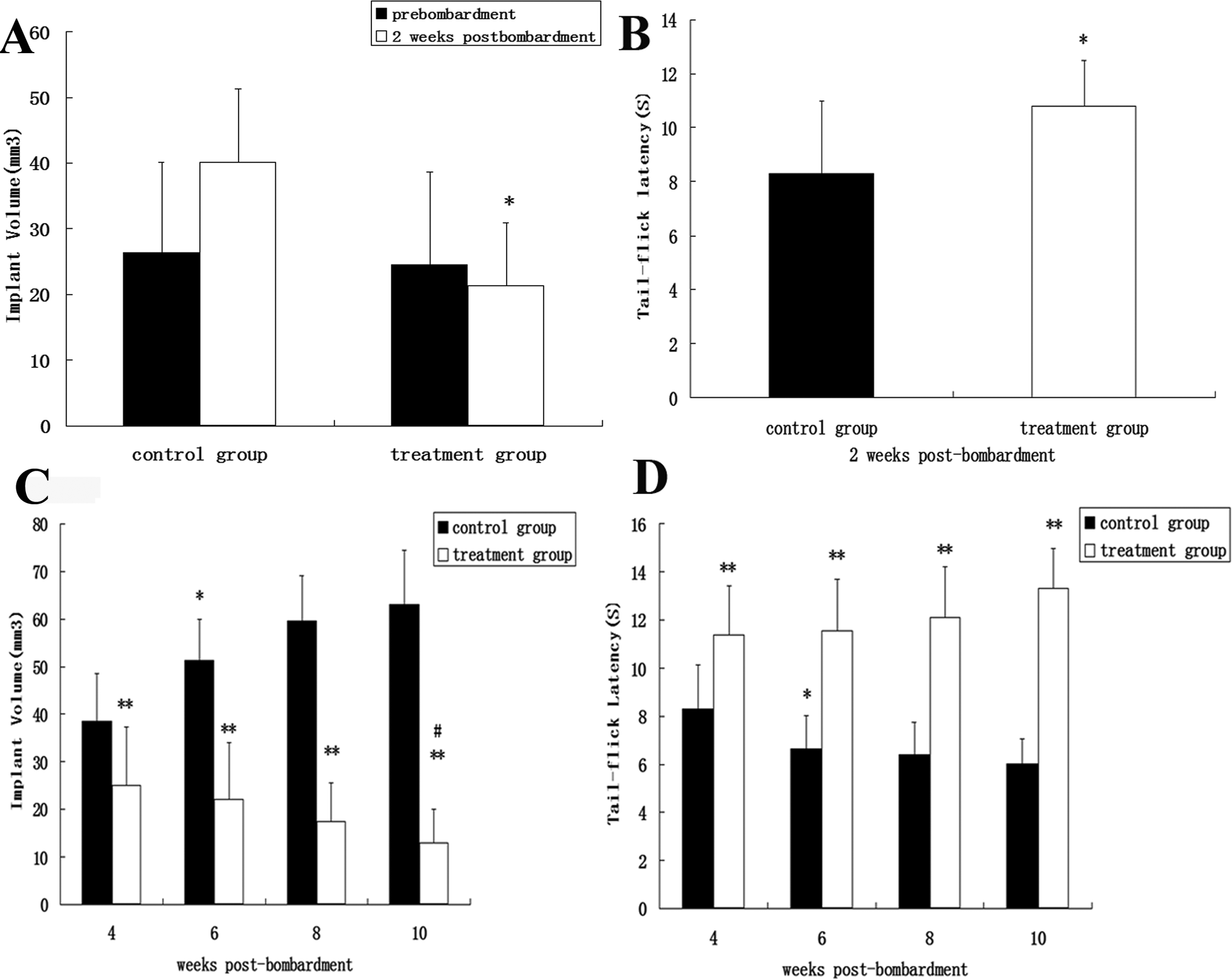
A, Pre-and postbombardment after 2 weeks’ volumes of endometriotic implants in the control and treatment group. There was a statistically difference in these volumes of implants between the control group and the treatment group postbombardment (*P < .05). B, The warmwater tail flick test 2 weeks postbombardment. A significant difference existed in the tail flick latencies between the control group and the treatment group postbombardment after 2 weeks (*P < .05). C, Spherical volumes of transplants postbombardment after different times. *Differs from the 4-week time period (P < .05); **differs from the control group at the same period (P < .05); #differs from the 4-week time period (P < .05). D, The tail flick latency postbombardment after different times. *Differs from the 4-week time period (P < .05); **differs from the control group at the same period (P < .05).
Spherical Volumes of Transplants and Tail Flick Latency Postbombardment After 4, 6, 8, and 10 Weeks
As shown in Figure 3C, the spherical volumes of the control groups postbombardment after 4 and 6 weeks were evaluated. A statistically significant difference was found between the 2 groups (*P < .05). Significant differences were also found in the volumes of the control and treatment groups postbombardment after 4, 6, 8, and 10 weeks (**P < .05). The volumes of the treatment groups postbombardment after 4 and 10 weeks were evaluated, and a statistically significant difference was found between the 2 groups (#P < .05). A significant difference was observed in the tail flick latencies between the control and treatment groups postbombardment after 4, 6, 8, and 10 weeks (**P < .05; Figure 3D).
The β-NGF Secretion Analysis Postbombardment After 2 Weeks
The β-NGF concentrations in the serums and supernatants of the peritoneal fluid were measured with an ELISA kit to further quantify the β-NGF expression in both the treatment and control groups postbombardment after 2 weeks. Figure 4 shows that the β-NGF protein concentrations in the serums were lower in the treatment group than in the control group (P < .05). The β-NGF protein concentrations in the supernatants of the peritoneal washing also decreased in the treatment group compared with the control group (P < .05). Analysis with the ELISA kit revealed that the siRNA-mediated silencing of the β-NGF gene expression after gene transfer had effects on the treatment group but not on the control group postbombardment after 2 weeks.
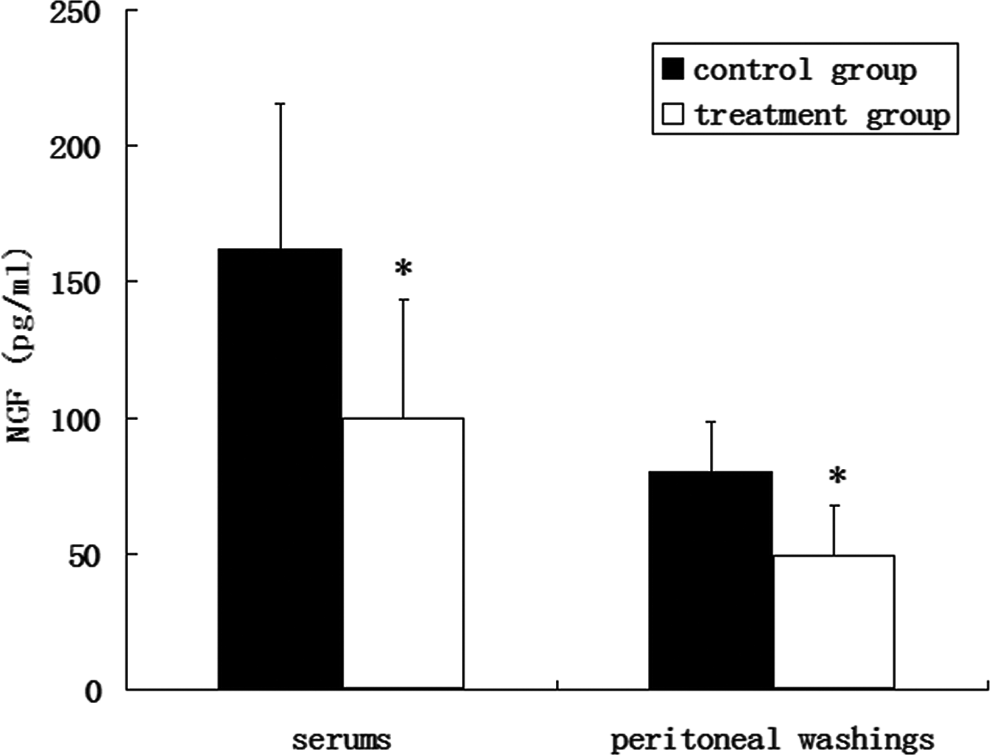
The β-NGF secretion analysis postbombardment after 2 weeks. The NGF protein levels in serums and supernatants of peritoneal washing from the control group and the treatment group were measured by ELISA postbombardment after 2 weeks. Both NGF levels in serums and supernatants of peritoneal washing were lower in the treatment group than that in the control group (*P < .05). β-NGF indicates β-nerve growth factor; ELISA, enzyme-linked immunosorbent assay.
Mean Density of Nerve Fibers Postbombardment After 2 Weeks
The mean density of the nerve fibers postbombardment after 2 weeks is provided in Table 1. The nerve fibers stained with different kinds of special markers in the ectopic endometriotic lesions are shown in Figure 5. Significant differences were found between the treatment and control groups in terms of the mean density of the nerve fibers in the endometriotic implants stained with most of the specific markers except in NF and VIP. The phenotype of the innervation and not its density was influenced by the treatment postbombardment after 2 weeks.
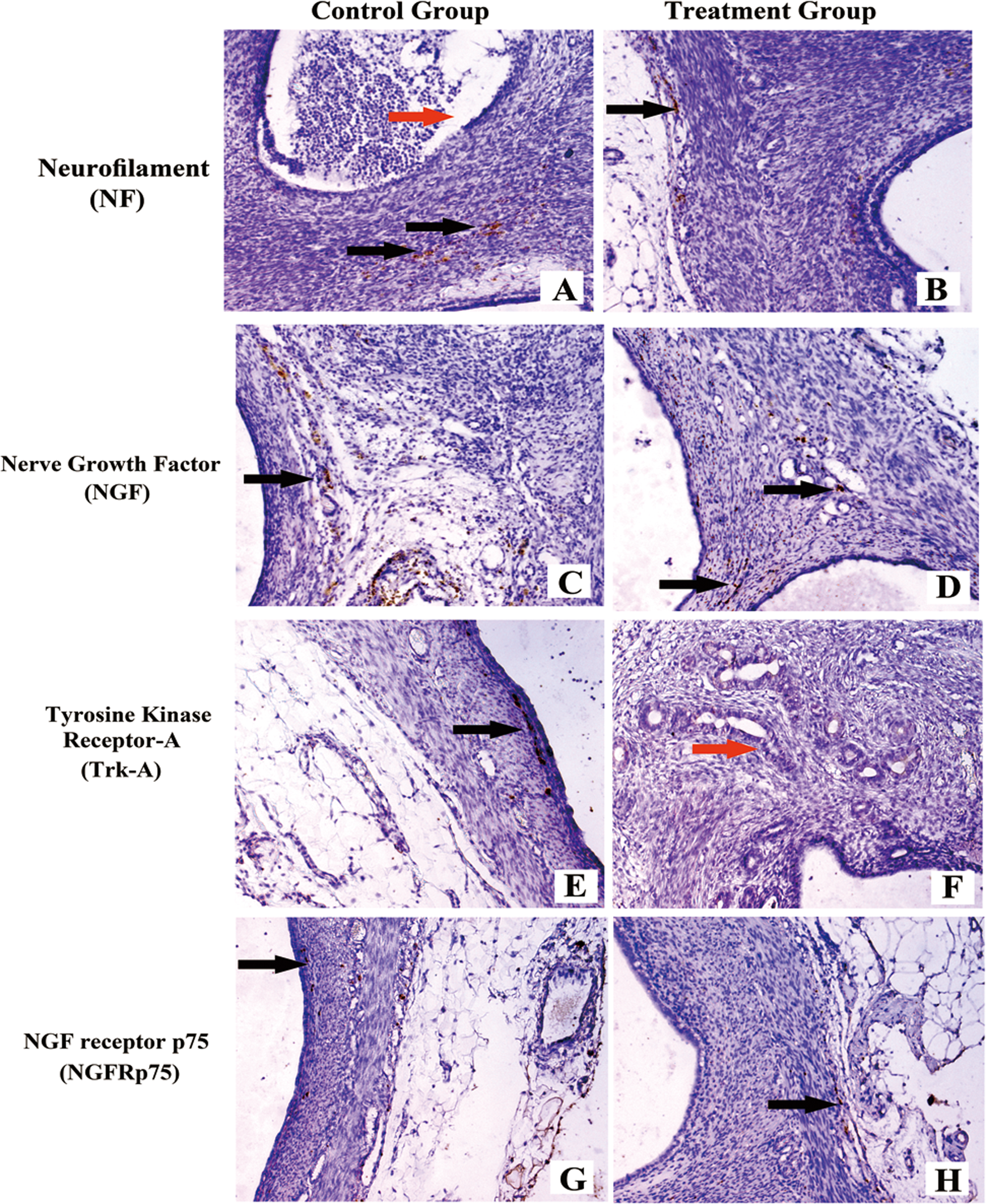
Nerve fibers in ectopic endometriotic lesions postbombardment after 2 weeks. A, Ectopic endometriotic lesion stained with NF from the control group with the mock plasmid by gene transfer. B, Ectopic endometriotic lesion stained with NF from the treatment group with the specific β-NGF siRNA by gene transfer fibers. C, Ectopic endometriotic lesion stained with NGF from the control group. D, Ectopic endometriotic lesion stained with NGF from the treatment group. E, Ectopic endometriotic lesion stained with Trk-A from the control group. F, Ectopic endometriotic lesion stained with Trk-A from the treatment group. G, Ectopic endometriotic lesion stained with NGFRp75 from the control group. H, Ectopic endometriotic lesion stained with NGFRp75 from the treatmen group.
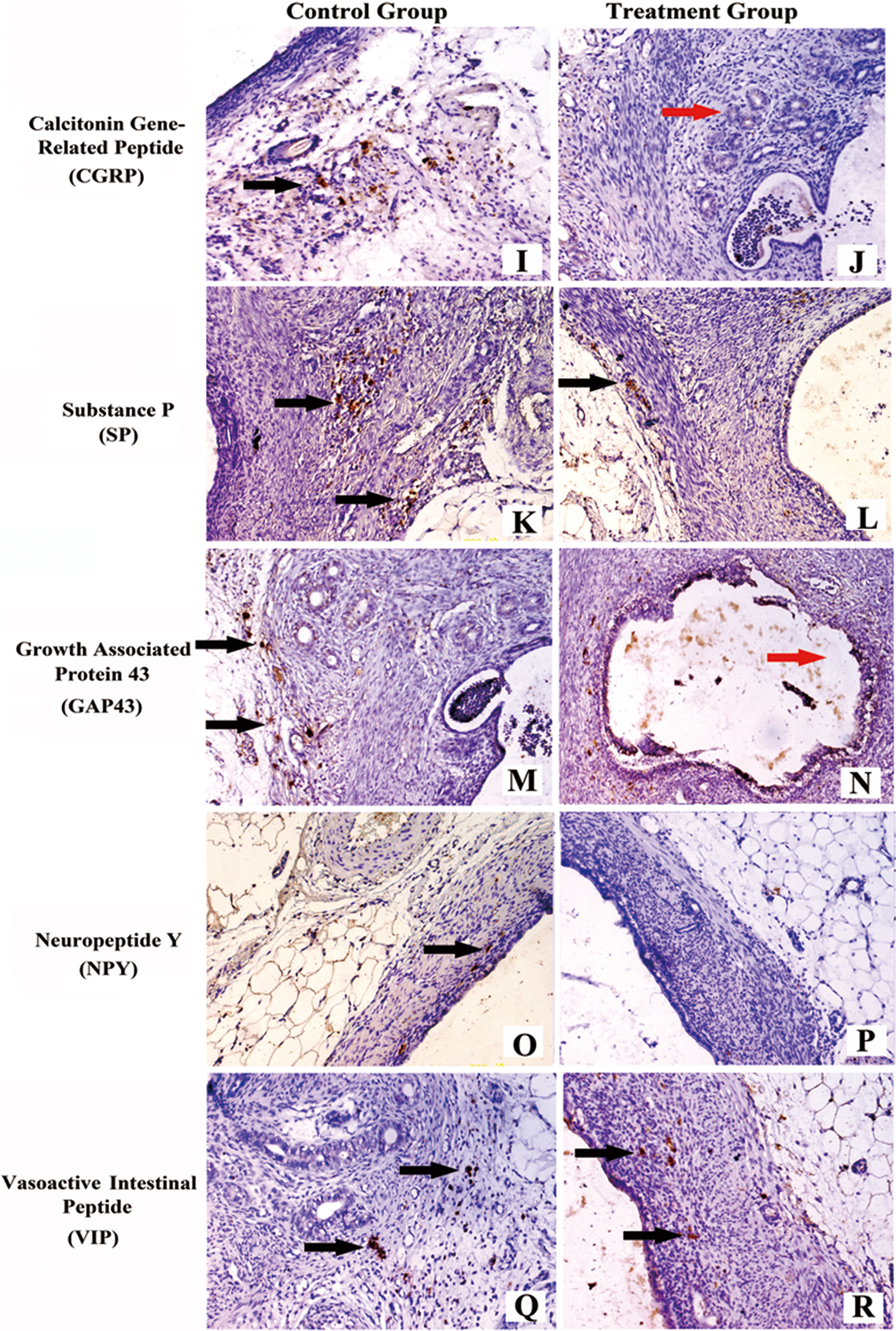
I, Ectopic endometriotic lesion stained with CGRP from the control group. J, Ectopic endometriotic lesion stained with CGRP from the treatment group. K, Ectopic endometriotic lesion stained with SP from the control group. L, Ectopic endometriotic lesion stained with SP from the treatment group. M, Ectopic endometriotic lesion stained with GAP-43 from the control group. N, Ectopic endometriotic lesion stained with GAP-43 from the treatment group. O, Ectopic endometriotic lesion stained with NPY from the control group. P, Ectopic endometriotic lesion stained with NPY from the treatment group. Q, Ectopic endometriotic lesion stained with VIP from the control group. R, Ectopic endometriotic lesion stained with VIP from the treatment group. Scale bars represent 50 μm in A-R (magnification ×200). Black arrows represent nerve fibers and red arrows represent endometriotic glands. β-NGF indicates β-nerve growth factor; siRNA, small interfering RNA; VIP, vasoactive intestinal polypeptide; NF, neurofilament; NGF, nerve growth factor; Trk-A, tyrosine kinase receptor-A; NGFRp75, NGF receptor p75; CGRP, calcitonin gene-related peptide; SP, substance P; GAP-43, growth-associated protein 43; NPY, neuropeptide Y.
Quantitative Assessment of the Endometrial Nerve Fiber Mean Density Stained Against Different Neural Markers Postbombardment After 2 Weeks in Model Rats With Endometriosis.a
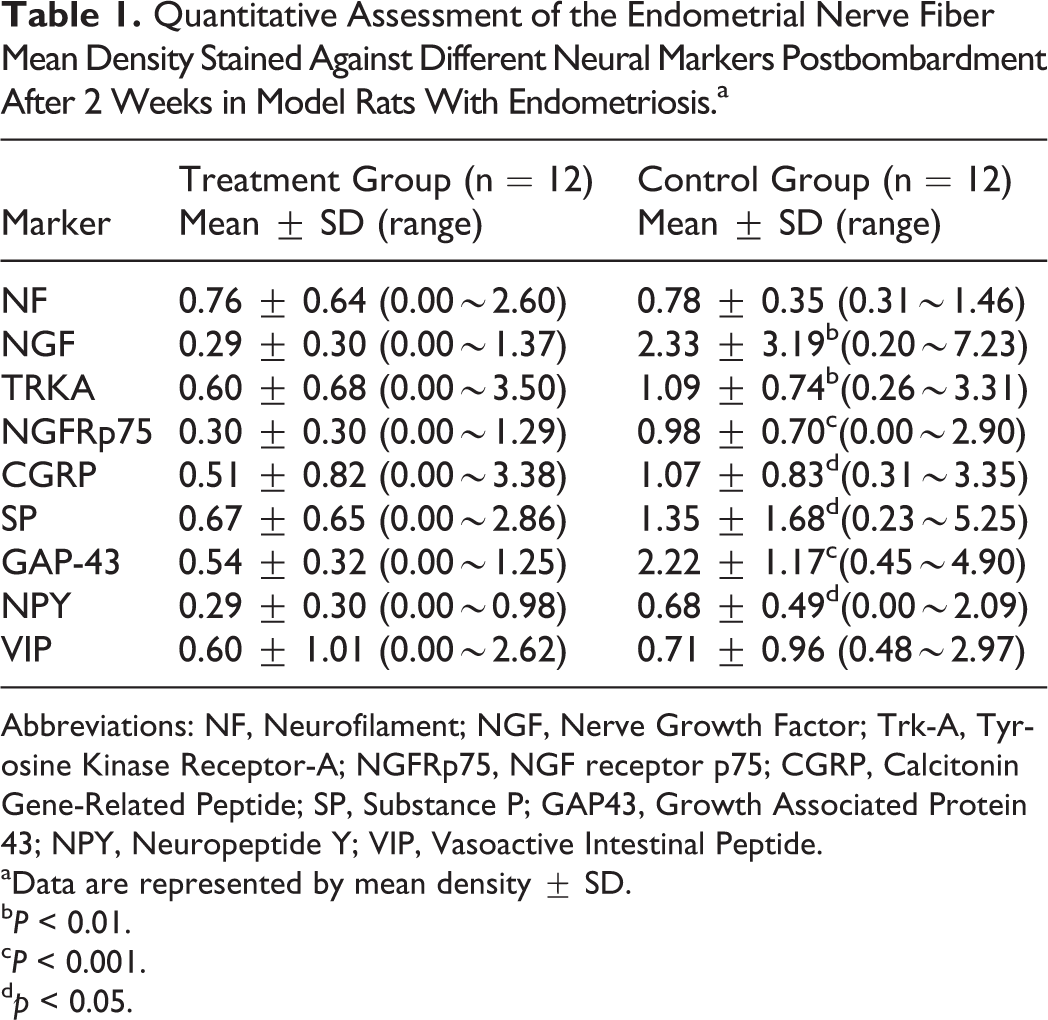
Abbreviations: NF, Neurofilament; NGF, Nerve Growth Factor; Trk-A, Tyrosine Kinase Receptor-A; NGFRp75, NGF receptor p75; CGRP, Calcitonin Gene-Related Peptide; SP, Substance P; GAP43, Growth Associated Protein 43; NPY, Neuropeptide Y; VIP, Vasoactive Intestinal Peptide.
aData are represented by mean density ± SD.
b P < 0.01.
c P < 0.001.
d p < 0.05.
Discussion
This research demonstrated that specific β-NGF siRNA suppressed the size of the endometriotic lesions and prolonged tail flick latency. Studies show that surgically induced endometriosis results in vaginal hyperalgesia 32 and hypersensitivity to noxious thermal stimulus in rodents. 33 Women with endometriosis exhibit generalized hyperalgesia in body areas beyond the site of the ectopic implants without any significant change in the pain sensory threshold. 34 The results of the present study suggest that the use of tail flick latency as a proxy measure for endometriosis-associated generalized hyperalgesia in rodents is justifiable. The treatment results in improved tail flick latency and may relieve pain symptoms in rats with induced endometriosis. The outcomes of the spherical volumes of transplants and tail flick latency at different times after postbombardment show that the sizes did not return to their previous size and that the treatment had a long-term effect on generalized hyperalgesia postbombardment after 4, 6, 8, and 10 weeks. This finding indicates that the effect of siRNA may not be temporary. The outcomes also suggest that increased vaginal nociception depended on the endometriotic lesions. 35 The ELISA performed postbombardment after 2 weeks indicates that the concentrations of β-NGF in the serums and supernatants of the peritoneal washings decreased more in the treatment group than in the control group. This result suggests that the specific β-NGF siRNA of the treatment group could have some effects. Both SP and CGRP are sensory nerve fiber markers that can be present in both Aδ and C nerve fibers. The mean densities of the CGRP and SP immunoreactive nerve fibers were low in the treatment group, which indicates that the sensory nerve fibers were reduced. The NPY stands for sympathetic nerve fibers. A significant difference was observed in the mean density of the nerve fibers stained with NPY in the 2 groups. Less nerve fibers stained with NGF, TrkA, and NGFRp75 existed in the treatment group than in the control group. However, no statistically significant difference was observed in the mean density of the NF-immunoactive nerve fibers in the treatment and control groups. These findings indicate that the phenotype of the innervation, not its total density, was influenced by the treatment postbombardment after 2 weeks.
Endometriosis was induced in cycling rats by autotransplanting pieces of the uterus onto the abdominal arteries where the pieces developed into endometriotic lesions, namely, cysts. The cysts contained growth factors and cytokines and developed their own sympathetic and sensory C- and Aδ-fiber innervation. 25 This condition suggests that each cyst’s innervation is a major contributor in the maintenance and modulation of pain in the established endometriosis. 36 A sprouted innervation in the uterine transplants contributes to the development of hyperalgesia in the rat model of endometriosis. 37 Therefore, less innervation may likely ameliorate the pain of the disease.
The NGF is known to have an influence in the occurrence of hyperalgesia. 38 The NGF itself can serve as a pain mediator and can induce the expression of neuropeptides, including SP and CGRP, which are involved in modulating the transmission of central pain. 39 Transgenic animals lacking the gene for NGF or its specific receptor TrkA are born with no small-diameter nociceptive sensory neurons and are hypoalgesic. In contrast, animals with overexpressed NGF are behaviorally hyperalgesic. 40 The NGF directly activates and sensitizes primary afferent nociceptors. 41 The presence of TrkA on CGRP-positive sensory fibers are demonstrated, which are likely to be C- or Aδ-fiber nociceptors. 42 The increase in NGF in the cysts in the proestrous stage when hyperalgesia severity is greatest suggests that NGF directly activates C-fibers in cysts, thereby contributing to endometriosis-induced hyperalgesia. 43 This conclusion is consistent with clinical results showing that pain severity correlates with NGF levels in diseased tissues or extracellular fluid in conditions such as chronic prostatitis 44 and painful intervertebral disc. 45 These results provide strong support for the hypothesis that the reduction or absence of NGF is likely to alleviate endometriosis-associated pain in both rats and women, which is in agreement with the result of the present study.
The NGF increases the pain response by binding with Trk-A. 46 TrkA can signal through at least 6 different pathways. The major pathways include a mitogen-activated protein kinase pathway involving extracellular signal-regulated protein kinase 1/2 (ERK), a phosphoinositide 3-kinase pathway involving the serine/threonine kinase Akt, and a phospholipase C-γ1 pathway. 47 The reason for the existence of numerous pathways is unknown, but evidence (in vitro) suggests that different pathways or a combination of pathways mediate different NGF effects and that these pathways differ according to cell type. Basal and NGF-induced activation of ERK occur primarily in nociceptors. 48 The ERK activation contributes to pain hypersensitivity. 49 These evidences suggest that ERK has an important function in nociceptive pathways, especially in conditions that involve inflammation where NGF is also known to play a key role. 50 A study shows that the basal levels of activated ERK in dorsal root ganglia (DRG) cells are low but can be selectively increased in CGRP/trkA cells by NGF treatment. 51 The NGF can increase the number of sensory neurons and is selectively trophic for small fiber sensory neurons and sympathetic ganglion neurons. 38 We demonstrated that the application of β-NGF siRNA can decrease the sensory and sympathetic innervation of the cysts in the treatment group, which is consistent with the findings of previous studies. The densities of the different kinds of nerve fibers may be influenced by other pathways related to NGF. This condition could be one of the reasons the phenotype of the innervation and not overall density was affected by the treatment. Another reason could be the existence of an appropriate treatment time. The same treatment could produce the desired effect if done in a proper time. 37 Further investigation is required to clarify this phenomenon.
A recent study has suggested that the overexpression of NGF in the peritoneal fluid (PF) of women with endometriosis may promote neurite outgrowth in endometriotic lesions. 52 The study showed that the NGF inhibitor (both anti-NGF and K252a) blocks neurite outgrowth in DRG and that NGF is an important neurotropic factor in the mediation of nerve growth and innervation in peritoneal endometriosis. Another study found that immunodeficient mice xenografted with human breast cancer cells and treated with either anti-NGF antibodies or siRNA against NGF exhibit inhibited tumor growth and metastasis. These treatments can induce a decrease in cell proliferation with a concomitant increase in the apoptosis of breast cancer cells and inhibition of tumor angiogenesis when directed against NGF. 28 Shelton et al 53 indicated that the blocking function of antibodies against NGF completely reverses established pain in rats with fully developed arthritis despite continuing joint destruction and inflammation. However, the β-NGF level of peripheral blood was not detected in the studies. 28,53
The present study showed that endometriosis regressed in size in the treatment group compared with the control group at postbombardment. The treatment led to ameliorated sensitivity to noxious thermal stimulus and less sympathetic and sensory innervation in the treatment group. The NGF addresses endometriosis-related symptoms and pathogenesis. Further investigation is required to clarify the influence of specific β-NGF-siRNA on endometriosis. However, the use of siRNA has some side effects, including the immunological off target effect that could activate the innate immune system and induce the huge production of inflammatory cytokines in mammals as well as nonspecific gene silencing and nonspecific toxic effect. Designing highly effective siRNAs or modifying siRNAs can retain or enhance the efficiency of gene silencing and reduce related side effects. 54 –57 No studies on the application of β-NGF siRNA in women with endometriosis have been assessed to date. Thus, the application of this treatment in women may have some limitations. For instance, the safety of siRNA should be investigated thoroughly before clinical application for therapeutic purposes.
In conclusion, the specific siRNA-mediated silencing of the β-NGF gene expression after gene transfer suppressed the growth of ectopic endometriotic implants, resulted in improved generalized hyperalgesia and reduced sympathetic and sensory nerve fiber density in the treatment group. Specific β-NGF siRNA-mediated silencing may be a novel therapeutic agent for endometriosis.
Footnotes
Authors’ Note
Yan Chen now works at the Department of Obstetrics and Gynecology of the Third Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan 450052, P. R. China.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was financially supported by the