
Abstract
The cyclical arrival of endometrial cells into the abdominal cavity through retrograde flux at menstruation represents the etiopathogenetic basis of endometriosis. The endometrium has peculiar regenerative properties linked to the presence of adult stem cells similar to mesenchymal stem cells (MSCs). Once in the abdominal cavity, these MSCs could proliferate, invade, and differentiate into endometrial cells, finally generating ectopic implants. As only differentiated endometrial cells, and not endometrial MSCs, possess steroid hormone receptors, MSCs could be responsible for the high rate of persistence/recurrence of the disease after hypoestrogenism-inducing therapies. Even angiogenesis promoted by MSCs could play an important role, as survival and proliferation of endometriotic tissue depend on the formation of new blood vessels. Inhibition of angiogenesis represents, in fact, a new, promising therapeutic approach for the disease. Further, medications directly targeting endometriosis MSCs could be effective, alone or in association with hormonal treatments, in increasing the success of medical treatment.
Introduction
Endometriosis is a chronic, estrogen-dependent disease characterized by the presence of ectopic endometrial tissue, composed of both glands and stroma, outside the uterine cavity. It affects women in the reproductive age causing a wide spectrum of clinical pictures, often including dysmenorrhea, pelvic pain, dyspareunia, infertility, and finally having a strong impact on the quality of life. 1
Endometriosis may be treated surgically and/or with medications aimed at avoiding menstrual bleeding (eg, continuously administered contraceptive pill) or at inducing a chronic hypoestrogenic state able to cause atrophic changes in the ectopic endometrium (eg, gonadotropin-releasing hormone analogs). Unfortunately, all currently available treatments frequently fail to completely eradicate the disease, and recurrence is often observed after a free interval of variable length. 2
Although endometriosis is a benign disease, it is characterized by some of the adaptive properties of cancer cells, such as deregulated proliferation, invasiveness, and neoangiogenesis. Indeed, the establishment and persistence of endometriotic lesions strictly depend on the formation of new blood vessels, which provide oxygen and nutrient supply to the ectopic tissue. 3 –5 Accordingly, antiangiogenic treatment could represent a possible therapeutic strategy against the disease. 6,7
Many theories have been proposed to explain the causes of endometriosis, and several factors have been implicated in its onset; retrograde menstruation, genetic predisposition, peritoneal cells metaplasia, and altered immunological surveillance, among others, are potentially involved. Moreover, in the last decade, cell populations with markers and functional properties of stem cells have been identified in the endometrium, 8 –17 and their role in the pathogenesis of endometriosis has been hypothesized. 18 –21
Endometrial stem cells are physiologically implicated in the cyclic, monthly endometrial regeneration that occurs after menstruation. 9,10,15 –17,22 Through retrograde menstruation, they may reach the abdominal cavity, invade the peritoneum, and generate endometriotic implants.
Which are the mechanisms involved in stem cell proliferation, differentiation, and neoangiogenesis? Could endometrial stem cells explain why endometriosis so frequently persists or recurs after the therapies based on the induction of hypoestrogenism? Can these stem cells be counteracted (and the disease treated) using new pharmacological therapies?
The present review focuses on the available evidence to address these issues.
Pathogenesis of Endometriosis
Endometriosis is a frequent disease, it affects approximately 3% to 10% of women in the reproductive age, but its prevalence rises to 25% to 80% among those complaining of infertility and 40% to 80% among those affected by pelvic pain. 1,23 The endometrium-like, ectopic tissue consists of both glandular epithelium and stroma, with prevalent traits of benignity mixed to some features (eg, tissue invasion and neoangiogenesis) similar to malignant pathologies. 24
The main symptoms of endometriosis are chronic pelvic pain, dysmenorrhea, dyspareunia, and infertility; however, the clinical presentation is very variable among affected women, ranging from severely symptomatic to asymptomatic forms. 23,25 Two different conditions are traditionally distinguished, adenomyosis, characterized by the presence of endometrial glands and stroma within the myometrium, and “external” endometriosis, localized inside the pelvis or even in organs at a considerable distance from the anatomic site of origin (eg, the lung). Pelvic endometriosis is further subdivided into 3 prevalent clinical forms, (1) implants placed on the surface of the peritoneum (superficial endometriosis), (2) ovarian cysts delimited by ectopic endometrium (endometriomas), and (3) solid nodules of endometrial tissue, mixed with fat and fibromuscular tissue, placed more deeply under the peritoneum (deep endometriosis). 1
The pathogenesis of the disease has been extensively investigated in the last decades, and several hypotheses have been proposed 19 ; however, some mystery still surrounds its onset and some of its characteristic features, such as the ability to persist/recur after treatment. 26
The most popular, commonly accepted theory on endometriosis origin is Sampson’s theory, postulating that endometriotic implants arise from retrograde menstruation of endometrial tissue through the fallopian tubes into the peritoneal cavity. 23,27 –29 This has been confirmed by the finding that endometrial fragments are found in the pouch of Douglas. 30 The observation that endometriosis spontaneously occurs only in humans and primates supports the idea that the disease affects exclusively species that menstruate. 29 On the other side, retrograde menstruation is probably a very common event, but only a relatively small proportion of women develop endometriosis; this suggests that other factors, for example, immunological, genetic, and intraperitoneal biochemical alterations, must be involved. 1,8,10,27,31 –33 Furthermore, the alternative “embryonic rest” theory proposes that endometriosis could arise from Müllerian cells scattered inside the peritoneal cavity that could be induced to form endometrial tissue when submitted to specific biochemical stimuli. 34 Another hypothesis, also able to explain the extra-abdominal localizations of the disease, proposes that the genesis of endometriotic lesions could be peritoneal metaplasia and differentiation of mesothelial cells into endometrial cells. 33,35,36 Finally, it has also been postulated that blood or lymphatic vessels could be responsible for the dissemination of endometrial cells at distance. 37,38
Recently, a new putative pathogenetic mechanism for endometriosis has been proposed based on the finding that human endometrium hosts a small population of cells with functional properties of stem cells. 8 –17,22 These cells are postulated to be in charge of proliferation and cyclic regeneration of the endometrial tissue after menstruation. According to this hypothesis, endometriosis could be the result of migration of endometrial stem cells into the abdominal cavity, of their proliferation, peritoneal invasion, and differentiation into endometrial cells in ectopic sites. 18,19,22,39
Endometrial Regeneration and Endometrial Stem Cells
Endometrial cyclic regeneration
Along a woman’s reproductive life, the human endometrium undergoes approximately 400 cyclical processes of growth, differentiation, shedding, and regeneration. 40,41
Functionally, the endometrium is composed of 2 layers; the outer, functionalis layer and the inner, basalis layer. The first is composed of epithelial and stromal cells organized in glandular structures and is destined to shed at each menstruation. 42 Every month it grows from 0.5 to 1 mm to 7 to 10 mm in thickness under the influence of circulating estrogens and progesterone. 20,43 The basalis layer, instead, rests on the myometrium at the end of the breakdown period and after each menstruation acts in quality of germ cell compartment to rebuild the new functionalis layer. 40 The expression of estrogen and progesterone receptors was reported during the proliferative and secretory phases in both the layers. 44
Characteristics of stem cells
Stem cells are quite rare, undifferentiated cells defined by their functional ability to self-renew or differentiate into cells of different lineages. 45 They have a striking ability to generate identical daughter cells and proliferate generating a stem cell population. Alternatively, a stem cell can produce progenitor cells that further differentiate into transit-amplifying (TA) cells and still keep an active proliferating ability 46 (Figure 1). The TA cells may start to differentiate into more specialized cell types, acquiring specific surface markers and gradually losing the ability to self-renew. 20,47
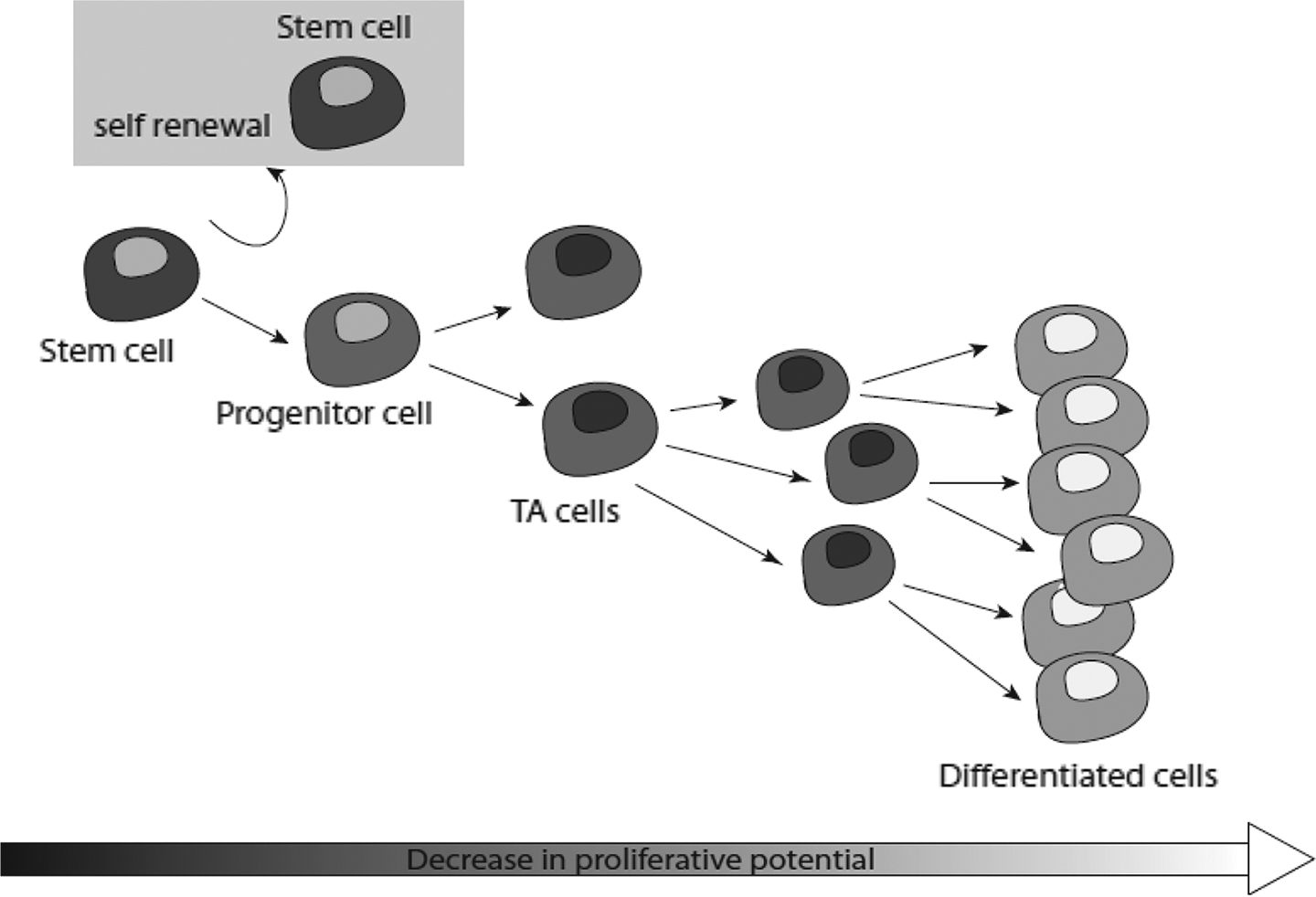
Stem cells self-renewal and differentiation. Stem cells are able to indefinitely self-renew or differentiate, giving rise to committed progenitors. At the next step, progenitors evolve into transit-amplifying (TA) cells that rapidly proliferate and differentiate to finally produce differentiated cells of a given cell lineage.
Stem cells may be found at each stage of embryonic development but are also detectable in most adult tissues. 45 Stem cells of the adult tissues are usually committed to generate differentiated cells of the organ in which they reside and participate in the organ homeostasis, as shown in the stomach, intestine, liver, prostate, and pancreas. 48 Stem cells with stromal characteristics, the so-called mesenchymal stem cells (MSCs), have also been detected in almost all organs and enriched in bone marrow, dental pulp, synovial fluid, adipose tissue, and umbilical cord blood. 49
Isolation, purification, and characterization of endometrial stem cells
The first evidence of the presence of stem cells, both of the epithelial and stromal type, in the human endometrium was provided in 2004. 8 Suspensions of purified cells obtained from human uterine tissue were cultured; rare epithelial (0.22% of all cells) and stromal stem cells (1.25% of all cells) with stem cell characteristics were isolated. 8 Clonogenicity of these cells, defined as the ability of a single cell to give rise to a colony when cultured at low densities, was verified through the formation of 2 types of cell colonies, large and small. Large colonies displayed a greater self-renewal ability than smaller ones, suggesting their derivation from endometrial progenitor stem cells; small colonies, instead, were more likely to derive from TA cells with lower proliferation potential. 8 The stemness of the cells isolated from the human uterus was also verified showing the expression of transcription factors such as Oct-4, crucial for the maintenance of pluripotency 50,51 and c-Kit (CD117), a receptor tyrosine kinase that binds to the stem cell factor. 8,16,52
The differentiative properties of endometrial stem cells were also shown by evaluation of the phenotype of endometrial colonies (colony-forming units [CFUs]). The CFUs of epithelial origin were found to generate gland-like, keratin-positive structures similar to endometrial glands. 14 Differently, endometrial stem cells of stromal origin were able to behave as multipotent stem cells and differentiate into 4 different mesenchymal cell lines when grown in specific culture media, myocytes (positively staining for α-smooth muscle actin and caldesmone), osteocytes (expressing alkaline phosphatase and parathyroid hormone receptor 1), adipocytes (positive for lipoprotein lipase), and chondrocytes (positive for collagen type II and Alcian blue stain). 14
It is well established that stem cells reside mainly in the basalis layer of the endometrium. Schwab and colleagues showed that endometrial samples collected either in the proliferative or in the secretory phase of the menstrual cycle, as well as from postmenopausal, inactive endometrium, contained epithelial and stromal stem cells with comparable clonogenicity. 9 This observation suggests that adult/progenitor stem cells reside in a uterine tissue that does not vary according to the menstrual cycle phase and is well preserved even in menopause, such as the basalis endometrium.
The search for label-retaining cells was also used to identify and purify progenitor stem cells in the endometrium. This technique is based on the cell ability to incorporate and retain the 5-bromo-2-deoxyuridine (BrdU) in DNA. Adult stem cells are characterized by rather infrequent cell division and therefore are able to retain the BrdU in their nucleus for a longer time in contrast to differentiated cells. In vivo experimental studies showed that BrdU was retained longer by stromal cells located in the perivascular site, near the junction endometrium-myometrial, an important site of remodeling both in human and murine endometrium, while in epithelial tissue, the retention was reduced. 10,11
Endometrial adult/progenitor stem cells in the basalis layer were also identified using the cytofluorimetric-based detection of the side-population cells (SP cells), defined by the unique ability of stem cells to extrude Hoechst 33342 dye. 53 Kato et al observed that 5.11% of the cells constituting human endometrium belong to the SP, and a higher proportion of these cells just appear at the end of menstruation, when only the basalis layer is well preserved. 12 In addition to this finding, endometrial MSCs were also found in the functionalis and in the menstrual blood, implicating a broader localization than the basalis layer. 54 Further, the similar gene expression profile of premenopausal basalis and postmenopausal basalis endometrium suggest that a population of putative endometrial stem/progenitor may reside in the basalis layer of endometrium, playing a key role in the cyclical regeneration of endometrial epithelium and stroma that occurs after every menstruation. 19,22,55,56
Masuda and colleagues 15 obtained a small population of cells from human endometrium, with the same phenotype of SP cells. This population exhibited the expression of several endothelial cell markers compared to endometrial main population of cells, and when transplanted under the kidney capsule of immunodeficient mice, it showed the ability to reconstitute organized endometrial tissue. 15
Which is the origin of endometrial adult/progenitor stem cells? Do they permanently reside in the basalis layer of the endometrium or, alternatively, do they arrive there from the bone marrow that contains both mesenchymal and hematopoietic stem cells? 57,58
Several data of the literature suggest a localization of bone marrow-derived cells in the endometrium. Indeed, the uterus of women previously submitted to bone marrow transplantation was found to contain a population of epithelial and stromal cells having the same human leukocyte antigens of the bone marrow donor. 59 In a similar experiment on female mice transplanted with bone marrow of male mice donors, the cytological analysis of uterine samples 6 months after the transplant showed the presence of the SRY gene, clearly of male origin. 59 However, the relevance of this observation is still undetermined.
The phenotype analysis of MSCs derived from the human endometrium showed that they possess some features in common with MSCs derived from other tissues, such as the expression of high levels of ITGB1 (CD29), CD44, NT5E (CD73), and Thy1 (CD90). 5,13,14,16,54,60,61 Approximately 20% of the endometrial MSCs resulted positive for endoglin (CD105) but were negative for platelet endothelial cell adhesion molecule 1 (CD31), CD34, and protein tyrosine phosphatase, receptor type, C (CD45; Table 1). 14 As MSCs in other tissues, endometrial MSCs are likely pericytes and express markers of perivascular cells such as platelet-derived growth factor β (PDGF-β) receptor and CD146. 54
Phenotypical Characterization of the Stem Cells Derived From Human Endometrium.a

Abbreviation: MSC, mesenchymal stem cell.
a The phenotype analysis of stem cells derived from human endometrium shows that they possess some features in common with adult MSCs, such as the expression of high levels of CD29, CD105, CD146, CD44, CD41, CD73, and CD90. Differently, endometrial stem cells result negative for CD31, CD34, and CD45.
Overall, the phenotype analysis confirms a marked similarity of endometrial MSCs with the MSC population detected in other organs. 5,14,54,61,62 As reported for other MSC sources, it is conceivable that MSCs are resident in the uterus. However, 2 populations of adult/progenitor stem cells could even coexist in the human endometrium, one permanent resident and the second continuously immigrating from the bone marrow.
Adult/Progenitor Stem Cells in Human Endometriosis
The existence of a population of adult/progenitor MSCs in endometriosis is strongly suggested by studies performed either on tissue samples or using animal models in which a disease similar to human endometriosis may be induced. 63 –66
Animal models
The use of animal models proved to be essential for the study of the pathogenesis and therapeutic approaches of endometriosis. Some researchers have used rodents as an in vivo assay to study endometriosis; however, the inoculation of human endometrial tissue in the peritoneal cavity of a rodent has shown several limits. 67 Rodents and humans, in fact, are quite different as rodents lack menstruation and never show spontaneous endometriosis; artificial endometriosis may be induced in them by xenotransplantation of endometrial tissue at ectopic intra-abdominal sites, but their reaction to the disease is quite different from that of humans. Nonhuman primates (baboons, Rhesus monkeys, and Cynomolgus monkeys) would undoubtedly represent the most appropriate animal model for endometriosis research in view of their marked phylogenetic similarity to humans; moreover, they can develop spontaneous endometriosis, macroscopically and histologically comparable to the human disease. 63 –65 Unfortunately research in nonhuman primates has enormous economical costs and is quite complicated to set up; for this reason, most studies available to date have been performed in rodents.
Masuda et al 66 demonstrated that human endometrial cells are able to generate foci of endometriosis when transplanted under the kidney capsule of ovariectomized, immunodeficient NOD/SCID/γcnull mice. After xenotransplantation at this site, the generation of a functional endometrium may be observed; this tissue is responsive to exogenous ovarian steroids (estrogens and progesterone) and shows the same morphological and functional changes in the eutopic endometrium when these hormones are given for at least 10 weeks. 66 An important neoangiogenesis occurs in the transplanted tissue, and the mouse kidney is invaded by human blood vessels that finally produce a functional circulatory system. 66 The induced implants of ectopic endometrial tissue derive blood supply from the surrounding vascular system; human vessels connect with mouse vessels forming a highly integrated network of neoformed blood vessels. 68,69
Isolation and characterization of ectopic endometrial MSCs
The coexpression of Oct-4 and c-Kit, typically expressed by cells with stem characteristics, was reported in the tissue samples of endometriosis that showed a positivity for both proteins in about 60% of the cases. 70 Furthermore, a high expression of markers as Musashi-1, Notch-1, and SALL4, all expressed by stem cells, was detected not only in eutopic endometrial cells but also in endometriosis tissue. 71
Moreover, 3 recent studies isolated and characterized MSC populations from endometriosis tissue. 5,61,72 Kao et al first isolated MSCs from endometriotic implants and showed their increased proliferation and matrix invasion in respect to the endometrial tissue. 5 Our group also isolated MSCs from ovarian and peritoneal endometriotic implants and reported their increased proliferation, migration, and angiogenic properties in respect to MSCs from eutopic tissue of the same patient as well as from normal endometrial tissue. Regarding the phenotype, ectopic MSCs were instead completely overlapping with eutopic MSCs. 61
Different hypothesis could explain the altered phenotype detected in ectopic MSCs. The MSCs, once located in the microenvironment of the ectopic lesion, may undergo a selection process that could lead to the survival of the MSCs/clones with enhanced migratory, proliferative, and angiogenic properties. Alternatively, it can be postulated that the extrauterine microenvironment found in ectopic sites might epigenetically modulate the endometrial MSCs, thus changing their characteristics. 61 Indeed, the importance of epigenetic changes induced by the microenvironment is gaining great consideration in the pathogenesis of cancer and several other human diseases. 73
Role of stem cells in the natural history of endometriosis
Stem cells could play a role in the natural history of endometriosis at least at the following steps: the initial onset of the disease, its persistence after hormonal treatment, and its recurrence.
Some data suggest that a small amount of adult/progenitor stem cells deriving from the basal endometrium could be contained in the menstrual blood. In fact, cell colonies with a classical MSC phenotype were detected after 15 days of in vitro culture of the menstrual blood of a healthy woman. 74 The protein analysis of the culture medium of such MSCs showed the presence of granulocyte macrophage colony-stimulating factor, PDGF-BB, and proangiogenic factors such as angiopoietin 2 and vascular endothelial growth factor (VEGF) 74 , factors that stimulate active neoangiogenesis able to feed the ectopic cell colony and to establish a novel endometriosis implant.
Retrograde menstruation is considered a rather common event that occurs in 90% of the menstruating women. It has been hypothesized that in 6% to 10% of women who develop endometriosis, endometrial stem/progenitor cells, inappropriately shed during menstruation, reach the peritoneal cavity where they implant, proliferate, and establish endometriotic implants. 20
In women with a high number of menstrual events (eg, those who delay motherhood after 35-38 years) the risk that retrograde menstruation occurs would be higher. Moreover, if the patient has some genetic predisposition to an insufficient local immune reaction, endometrial cells/stem cells could be saved from the immune response, and several endometriosis localizations could start to develop. 75,76
The persistence and/or recurrence of endometriosis could also be due to stem cells activity. Surgical and medical treatments often fail to eradicate and definitively eliminate the disease. Probably in some cases, the currently available treatments do not destroy all endometriosis tissues, either because not all tissues are identified and eliminated during surgery or because not all the cells in ectopic implants are sensitive to estrogen deprivation. Different from the fully differentiated endometrial cells, MSCs from normal and endometriosis tissue were reported to lack expression of estrogen receptors that were then acquired during differentiation, suggesting that their survival and growth were estrogen independent. 61 As a consequence, a prolonged medication-induced hypoestrogenic state would cause the atrophy and death of differentiated cells only, but would be useless against stem cell population, that will survive despite the treatment. After months or years, the subsequent proliferation of these stem cells, overcoming the resistance of the local immunity,75 could regenerate the ectopic endometrium, finally causing the clinical reappearance of the disease.
However, in the treatment of patients with endometriosis, prevention of the endometriosis lesion development only will not be sufficient as a therapy. Treatment options, aimed at interfering with established lesions, have to be developed.
Effect of antiangiogenic therapy on endometriosis and endometrial progenitor/stem cells: In vitro and in vivo results
Angiogenesis is of pivotal importance for the growth and invasion of solid tumors as well as for the development of other invasive diseases such as endometriosis. 77,78 It is well established that the blockage of the development of the blood supply to growing implants leads to tissue hypoxia, growth arrest, and necrosis. 3,5 –7 In addition, recent data based on the direct effect of some antiangiogenic agents on endometrial MSCs added a new perspective in the mechanisms of their action, suggesting a direct role of antiangiogenenic drugs in the development and recurrence of endometriosis.
The most important angiogenic agent in endometriosis is VEGF. 79 More than 80% of blood vessels observed in endometriosis implants are pericyte free and very responsive to VEGF. 6 Indeed, increased levels of VEGF were found in the peritoneal fluid of patients with endometriosis and in ectopic endometrial tissue. 78,80,81
Among the different antiangiogenic agents, tyrosine kinase inhibitors showed a broader spectrum of targets beside VEGF. In particular, we recently focused our attention on Sorafenib (BAY 43-9006), a multikinase inhibitor targeting Raf kinases, such as RAF-1 and B-RAF, as well as several growth factor receptors. 82 –84 In particular, Sorafenib targets CD117 (c-Kit), VEGF receptor 2 (VEGF-R2) and VEGF-R3, CD140 (platelet growth factor beta receptor) as well as of the Raf-1 downstream protein Ezrin. This protein was previously shown to be involved in migration of endometrial stromal cells in patients with endometriosis. 73 After isolation of MSCs from eutopic and ectopic endometrium, we evaluated the MSC expression of the different Sorafenib targets. Both eutopic and ectopic endometrial MSCs expressed CD140 and not c-Kit or VEGF-R2 and VEGF-R3. Moreover, the expression of the activated phosphorylated form of ezrin was higher in ectopic MSCs in respect to eutopic and control lines, indicating possible VEGF-independent targets of Sorafenib in MSCs. 61
Sorafenib treatment inhibited the increased phosphorylation of ezrin in ectopic MSCs, and consequently, it was also effective in limiting the increased migration of ectopic MSCs. Moreover, it reduced the increased proliferation and VEGF release found in ectopic MSC. The inhibitory effect of Sorafenib on proliferative and angiogenic properties of ectopic MSC may be explained by its effect on tyrosine kinases that are target of Sorafenib, such as CD140. This mechanism was reported for tumor cell lines, where the inhibition of receptor signaling pathways reduced the cytokine-induced proliferation and HIF-1α transcriptional activity. 85 Indeed, HIF-1α activation was impaired in endometrial MSCs by Sorafenib.
The use of the antiangiogenic therapy for the treatment of endometriosis in vivo was tested in several experimental models (Table 2). Although these in vivo studies cannot dissect the possible mechanisms of the antiangiogenic agents, nor their direct activity on endothelial cells or endometrial stem/progenitors themselves, the results appear promising.
Studies on Antiangiogenic Treatment in Endometriosis.a

Abbreviation: VEGF-R2, vascular endothelial growth factor receptor 2.
a The table summarizes some of most recent studies inherent to the use of antiangiogenic molecules, with the aim of searching new and more effective treatments against endometriosis.
Dabrosin and colleagues reported a successful treatment of endometriosis in estrogen-supplemented ovariectomized mice by transient overexpression of the gene for the natural angiogenesis inhibitor angiostatin. Results showed a significant reduction in the number and size of endometriosis implants after 2 to 18 days from the inoculation of human endometrial cells in nude mice. 89 In another experiment, rodents with surgically induced endometriosis were treated with SU5416 and SU6668, 2 tyrosine-kinase receptor inhibitors targeting VEGF-R2. 87 This treatment after 14 days reduced microvessel density inside the endometriosis implants as well as the size of lesions. Bevacizumab, a recombinant human monoclonal anti-VEGF antibody, was tested on nude mice with surgically induced endometriosis. 86 Treatment with Bevacizumab, started 2 weeks after the surgical implantation of endometriosis, induced a significant reduction in cell proliferation, increased apoptosis, and markedly decreased vascular density in the implanted tissue as well as a striking reduction in VEGF levels in the peritoneal fluid. 86
Recently, Ozer and colleagues showed that both Bevacizumab and Sorafenib were able to induce a significant decrease in endometriotic implants and microvessels density in rats with induced endometriosis, without compromising the ovarian reserve of viable follicles. 88 Finally, studies conducted on mice with peritoneal endometriosis showed that treatment with daily doses of cabergoline, a drug that causes inhibition of VEGF-R2 phosphorylation, was able to significantly increase necrotic areas in endometriosis implants and promote their regression. 90,91
Conclusions
Although extensively studied, the etiopathogenic mechanism of endometriosis remains partially unclear. The disease is quite frequent during the reproductive age, and in most cases its onset seems to be related to menstruation. The number of menstruations that a woman has during her life is likely to be one of the main determinants of the risk of developing pelvic endometriosis. Pregnancy and lactation are considered protective factors simply for consequent lack of menstruation. More pregnancies and less menstruations causes higher protection, while no pregnancy or delayed pregnancy and more menstruations increase the probability of developing endometriosis.
During menstruation some shed endometrial cells reach the abdominal cavity through retrograde flux. This phenomenon is probably more common than previously believed, but in most cases, the number of cells that reach the peritoneum is so small that the local immune reaction destroys them, avoiding further consequences.
The endometrium is a highly dynamic tissue that regenerates and grows each month under the influence of ovarian estrogens. This striking regenerative ability is linked to the presence, in its basal and functionalis layers, of a small population of adult/progenitor stem cells similar to MSCs that actively proliferate and differentiate rebuilding the functional endometrium after each menstruation. The MSCs, once arrived into the peritoneal cavity by menstrual retrograde flux, could have the ability of generating endometriosis implants using their ability to proliferate, invade, promote neoangiogenesis, and differentiate into endometrial cells. In addition, in the peritoneum, MSCs may undergo a selection process or may be modulated by the local environment and change their characteristics. In fact, their functional characterization shows that they are more aggressive than normal MSCs and are unresponsive to steroid hormones. 61 Endometriosis adult/progenitor stem cells could therefore be responsible for the partial resistance of endometriosis to the pharmacologically induced hypoestrogenic state and consequently for the high rate of persistence/recurrence of the disease after the current hormonal therapies.
Angiogenesis may play an important role in the onset and maintenance of endometriosis implants. Indeed, survival and proliferation of endometriosis lesions strictly depend on the formation of new blood vessels that provide oxygen and nutrient supply. Inhibition of angiogenesis represents a new, promising therapeutic approach for the disease. Further, the reported effects of Sorafenib on endometriosis MSC proliferation, invasion, and HIF-1 α activation offer a new perspective for the use of this class of drugs. Antiangiogenic agents could be effective, alone or in association with hormonal treatment, in eliciting a more complete and definitive success of the medical treatment of endometriosis.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.