
Abstract
Preimplantation culture of mouse embryos has been suggested to result in reduced anxiety-like behavior in adulthood. Here, we investigated the effects of in vitro fertilization (IVF), embryo culture, and different diets on anxiety-like behavior using the elevated plus maze (EPM). We hypothesized that exposure to suboptimal conditions during the preimplantation stage would interact with the suboptimal diet to alter behavior. The expression of genes related to anxiety was then assessed by quantitative real-time polymerase chain reaction in various brain regions. When fed a normal diet during gestation and a moderately high-fat Western diet (WD) postnatally, naturally conceived (NC) and IVF mice showed similar anxiety-like behavior on the EPM. However, when fed a low-protein diet prenatally and a high-fat diet postnatally (LP/HF), NC mice showed a modest increase in anxiety-like behavior, whereas IVF mice showed the opposite: a strongly reduced anxiety-like behavior on the EPM. The robust reduction in anxiety-like behavior in IVF males fed the LP/HF diets was, intriguingly, associated with reduced expression of MAO-A, CRFR2, and GABA markers in the hypothalamus and cortex. These findings are discussed in relation to the developmental origin of health and disease hypothesis and the 2-hit model, which suggests that 2 events, occurring at different times in development, can act synergistically with long-term consequences observed during adulthood.
Introduction
Gestation is a highly sensitive period and if gametes, embryos, and fetuses are exposed to stressful environments, poor health outcomes can appear later in life. 1 For example in the historical cohort of women pregnant during the Dutch famine of winter 1944 to 1945, exposure to caloric restriction during the first half of pregnancy resulted in a significant increase in obesity rate in their 19-year-old children, whereas exposure during the last trimester or first month of life produced a lower rate of obesity. 2
The Developmental Origin of Health and Disease (DOHaD) hypothesis, also known as the Barker hypothesis, proposes that susceptibility to several adult diseases derives from embryonic adaptation to suboptimal conditions in utero or during the preimplantation period. 3 Embryos are not lost, but adaptive metabolic programming occurs with functional consequences that can manifest later in life. 4,5 Etiologic events leading to obesity, hyperlipidemia, type 2 diabetes, hypertension, cardiovascular diseases, shortened life span, asthma, and other conditions have been linked to in utero insults. 4,6 –10 These harmful events can occur even before implantation. In rats, for example, a low-protein maternal diet experienced exclusively during the preimplantation period resulted in blastocysts exhibiting a slower rate of cell proliferation and offspring with lower birth weight, reduced postnatal growth rate, abnormal organ to body weight ratios, and higher systolic blood pressure. 3
The DOHaD hypothesis is particularly relevant, given the increasing use of assisted reproductive technologies (ARTs). Several studies in humans and animals have shown that in vitro fertilization (IVF) and embryo culture can be metabolically taxing on preimplantation embryos. Indeed, oxidative stress can have a considerable impact on embryo development and cultured preimplantation embryos show metabolic changes, 11 abnormal metabolic and ultrastructural configurations, 12 increased programmed cell death in cells in the inner cell mass, 13 abnormalities of placentation, 14 and altered expression of developmentally important genes. 15 –19 Culture media and/or nonphysiological oxygen tension contribute to these adaptive changes to increased oxidative stress. 18,20
The long-term developmental and behavioral consequences of these adaptive responses to subtle changes in the prenatal environment are poorly known. Recent studies have shown that, although mice derived from cultured embryos present only minor alterations in preweaning behavioral milestones, as adults they exhibit alterations in spontaneous anxiety, locomotor activity, spatial, and implicit memories; these differences are particularly evident in male mice. 21,22 In particular, the work of Ecker et al. showed that offspring that resulted from culture in vitro during the preimplantation period had decreased anxiety as adults. 21 Of note, reduced anxiety/fearfulness in rodents signify more risky behavior and, as a consequence, being more vulnerable to attacks by a predator. 23
Suboptimal conditions and adaptation during an early period of development (first hit) may set the stage for long-term vulnerability to a following adverse event (second hit) that may occur also in utero or later in life, resulting in a more extreme outcome or phenotype. This is known as the “2-hit” model. 23 Based on this model, the present study was devised to assess the combined effects of conception method (hit 1) and altered diet (hit 2) on anxiety-like behavior in the elevated plus maze (EPM) and the expression of selected genes in brain regions associated with stress response, anxiety, and emotional behaviors.
Altered diets may represent taxing conditions in utero, postnatally, or both. Mice naturally conceived (NC) and generated by IVF were exposed to 2 types of diets. The Western diet (WD) group, so called for its similarity to the daily diet used in most “Western” countries, was fed a control diet in utero and during the nursing period followed by a diet with moderately high-fat content after weaning. Mice fed more challenging dietary protocol were given a low-protein diet in utero, a normal diet until weaning and a high-fat diet postweaning (LP/HF), a well-established stressful nutrition paradigm. 24
We found that when compared to the other subgroups, IVF mice fed the LP/HF diet spent more time on the EPM open alleys, suggesting reduced levels of the anxiety-like behavior. Reduced levels of anxiety-like behavior were paralleled by alterations in gene expression of MAO-A, CRF, and GABA, in anxiety-related cortical and subcortical structures and in the cerebellum of male mice. Implications of these findings will be discussed in relation to the DOHaD hypothesis.
Material and Method
Animals
C57Bl/6J mice were used for all experiments. They were kept in the University of California San Francisco (UCSF) animal facility with controlled temperature conditions at 23°C, a 12:12 light–dark cycle and ad libitum access to water and food. Female mice were randomly assigned to an IVF group (oocyte collection) or to a natural breeding group. Animals were weighed at birth and weekly until behavioral testing and were maintained in-group housing. All experimental procedures described in the present study were approved by the Committee on Animal Research at the UCSF.
Preimplantation Embryo Production
The IVF groups were generated from 25 IVF experiments using 118 superovulated females, 25 sperm donors, and 35 transfer recipients. Oocytes were obtained from superovulated C57Bl/6J female mice as described previously. 14,25 Briefly, female mice were injected with 5 IU of pregnant mare serum gonadotropin and 46 to 48 hours later with 5 IU of human chorionic gonadotropin (hCG). The following morning, oocytes were collected from the ampullae. Oocytes were then incubated for 4 hours in Whitten Medium (WM) containing 30 mg/mL bovine serum albumin and sperm obtained from the cauda epididymis of C57Bl/6J males. After 4 hours, fertilized oocytes were rinsed and cultured in WM in humidified air at 37°C with 5% CO2 to the blastocyst stage. Embryos were transferred approximately 112 hours after hCG injection to the uterus of surrogate females (CD1) and kept in the same animal facility. Females assigned to the breeding group (control, NC) were bred overnight to C57Bl/6J males. Mice were tested for differences in behavior between the age of 10 and 28 weeks.
Diet
Pregnant mice from each group (NC and IVF) were randomly assigned to 1 of 2 dietary regimens (Figure 1). The WD groups (normally conceived and IVF) were assigned a control diet (D02041001; Research Diets, New Brunswick, New Jersey; Table S1) in utero and during nursing, followed by a diet with moderate fat content (D12266B) after weaning. Mice exposed to a stressful diet were fed a low-protein diet in utero (D02041002), control diet during nursing, and a high-fat diet (D12492) postweaning (LP/HF groups).
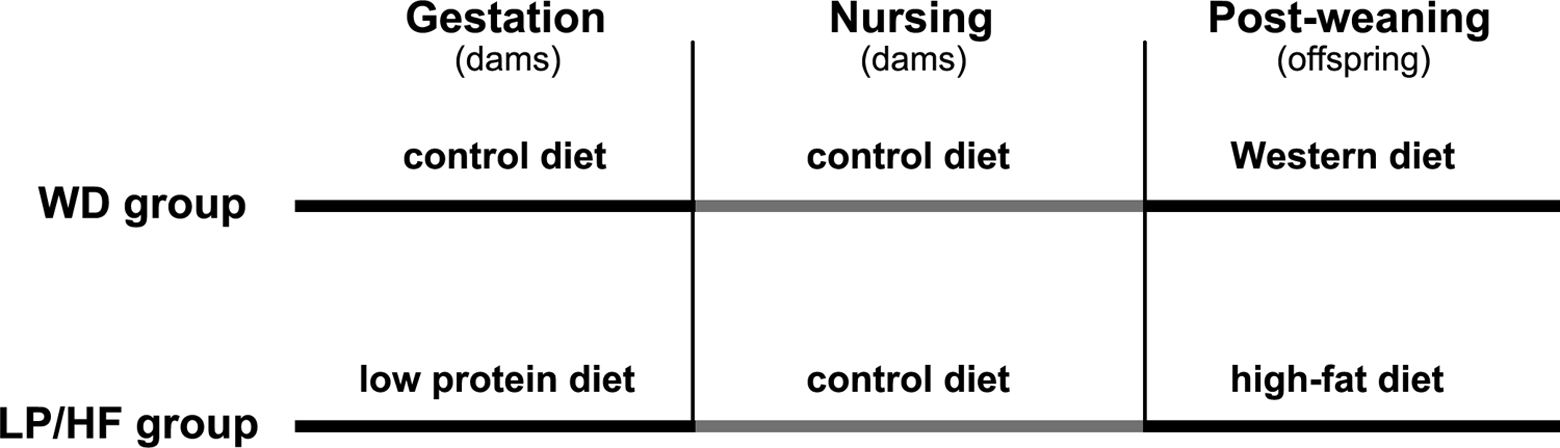
Diet protocol. Outline of the sequence and types of diets fed to mice in the Western diet (WD) and low-protein diet prenatally and a high-fat diet postnatally (LP/HF) groups.
Behavioral Assessment
Stress/anxiety-related behaviors were assessed with the EPM in 10- to 28-week-old adult mice. The EPM consisted of 2 opposing open arms (38 × 5 cm) and 2 opposing enclosed arms (38 × 5 cm, walls 15 cm high) connected by a central platform 5 × 5 cm and elevated 50 cm from the workbench. Arms were equipped with rows of infrared beams interfaced with a computer (Hamilton-Kinder, Poway, California). Mice were brought to the testing room 30 minutes before the beginning of the trials and allowed to habituate to dim (120 Lux) lighting conditions. 26 Trials started with individual mice placed in the central platform facing the enclosed arms and allowed to explore the plus-maze for 5 minutes. After every animal’s performance, the EPM was carefully rinsed with a diluted solution of sodium hypochlorite. Distance covered and time spent on open arms were recorded. Head poke-ins, number of entries on the open alleys, distance covered, and time spent on closed alleys were also recorded as measures of activity. It should be noted that time spent in the closed arms is the complementary measure of the time spent on open alleys. Distance covered was expressed in centimeters and time spent in seconds.
The data from male and female mice were evaluated separately: male NC-WD (n = 15, 4L), male NC-LP/HF (n = 8, 2L), male IVF-WD (n = 9, 4L), male IVF-LP/HF (n = 6, 3L), female NC-WD (n = 10, 4L), female NC-LP/HF (n = 4, 2L), female IVF-WD (n = 12, 3L) and female IVF-LP/HF (n = 5, 3L).
Brain Dissection and Quantitative Real-Time Polymerase Chain Reaction
After the behavioral sessions, animals were deeply anesthetized with CO2 and killed by decapitation. The brain was immediately removed and dissected at 4°C as described, 27 obtaining the following anatomical areas: amygdala, brain stem, caudate, cerebellum, entire cortex of 1 hemisphere, hippocampus, the entire hypothalamus, and septum. The same 2 individuals performed brain dissections in all the experimental groups to ensure consistency of method. Each anatomical specimen was immediately snap frozen and stored at −80°C until use. Two operators were working at any given time. One operator dissected the cerebellum, brain stem, and caudate, while the other operator removed the amygdala, cortex, hippocampus, and hypothalamus. The whole process took approximately 20 to 30 minutes.
Gene expression was examined in dissected brain regions. All gene transcripts were evaluated by quantitative real-time polymerase chain reaction (qPCR). Samples from the WD group (both NC and IVF) and LP/HF group were analyzed at separate times so that gene expression levels could only be compared between conception groups and not between diet groups. For each replicate, total RNA was isolated from each area using the “All Prep” DNA/RNA mini kit (Qiagen, Valencia, California) according to the manufacturer’s instructions.
19
Briefly, DNAse-treated RNA was obtained and converted to complementary DNA (cDNA). The q-PCR was performed using Bio-Rad universal PCR supermix (Sigma, Saint Louis, Missouri), gene specific-primers, (Bio-Rad, Hercules, California), and 1 ng of cDNA. Three replicates were used for each q-PCR reaction; data were normalized to GAPDH expression; a minus-template served as control. Primers used to detect the genes are presented in Table S2. Average fold-changes in gene expression were normalized so that expression in NC mice was equal to 1 and expression in the experimental groups was designed as relative expression (RE). The RE was calculated using the 2 −ΔΔCt method
28

where C t represents the average value of all samples.
Statistics
Data are presented as the mean ± standard deviation. Litter size and body weights at birth were compared using an analysis of variance, and pairwise comparisons were adjusted with the Bonferroni method.
Analysis of covariance models were used to test the effect of conception and diet on behavioral data while controlling for age, sex, and weight at testing. All pairwise comparisons were adjusted with the Bonferroni correction. The Student’s t-test was used to compare gene expression levels in the brain. All statistical testing was performed at .05 level of significance.
SAS version 9.3 (Cary, North Carolina) was used for statistical modeling.
Results
Body Weight
Litter sizes were as follows: NC-WD 6.4 ± 2.4 (n = 7), NC-LP/HF 6 ± 0 (n = 2) and IVF-WD 4 ± 1.7 (n = 9), IVF-LP/HF 2.8 ± 1.6 (n = 5). The NC litters from dams fed the WD diet were significantly larger than IVF litters from dams fed a low-protein diet. Newborn IVF pups weighed significantly (P < .01) more than the NC pups, but there was no difference between diet groups (NC-WD 1.3 ± .1 n = 45, NC-LP/HF 1.2 ± .1 n = 12, IVF-WD, 1.6 ± .2 n = 36, IVF-LP/HF 1.5 ± .2 n = 14).
Behavior
Stress/anxiety-like behavior was examined with the EPM. Higher anxiety is correlated with the mouse’s avoidance of the elevated open arms of the maze and reduced anxiety with a mouse’s confidence to enter and explore the open arms. Distance covered, time spent, number of head pokes, and entries in the open arms were recorded and compared in order to assess differences in stress/anxiety-like behavior. The distance covered in the closed arms of the EPM is an estimate of the general activity level of the mice. 29
The ANCOVA model (Table 1; Figure 2) revealed that, when controlled for weight and age, sex was the only significant predictor (P < 0.01) for distance covered in the closed arms, that is, general activity. Specifically males were less active, traveling 253.5 cm less in the closed arm of the EPM. For parameters that indicated reduced stress/anxiety-like behavior, open time, open distance, and open entry, the interaction between conception and diet was the only significant predictor (P < .05). When fed the WD, NC and IVF mice spent a similar amount of time and covered a similar distance in the open arms of the EPM. However, IVF mice fed the LP/HF diet covered a significantly greater distance (257 cm; P < .05) and spent more time (40.27 seconds; P < .05) in the open arms than NC mice fed the same LP/HF diet. Diet did not significantly affect the amount of time or distance spent by NC mice in the open arm of the EPM. In contrast, diet did affect the behavior of IVF mice. The IVF mice fed the LP/HF diet traveled 241 cm more (P < .01) and spent 34.3 seconds more (P < .05) in the open arm than the IVF mice fed the WD.
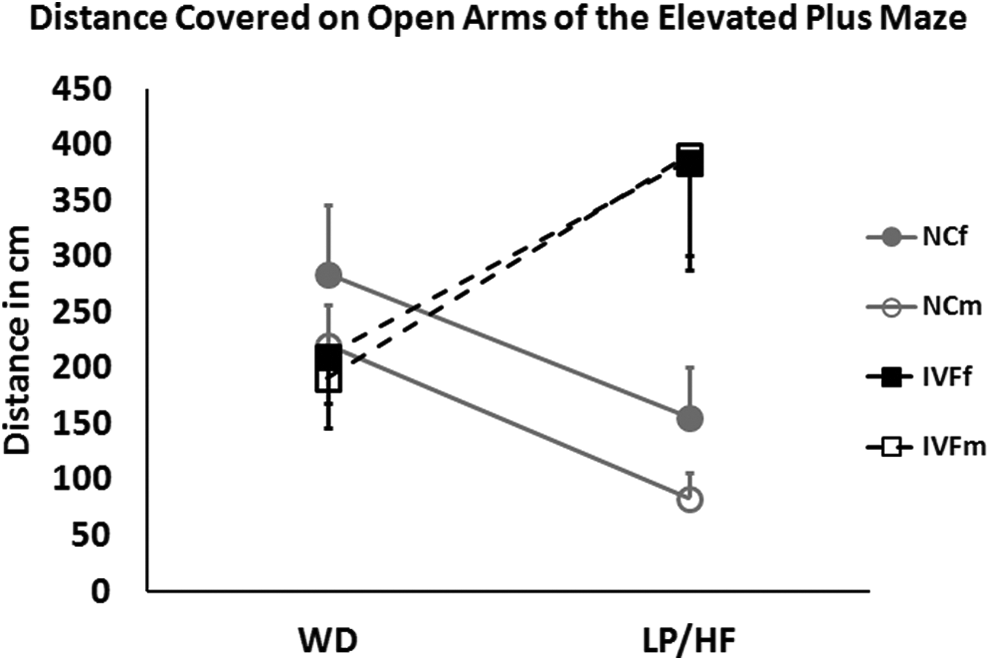
Behavior in the elevated plus maze (EPM). Distance covered on the EPM open arms by naturally conceived (NC) and in vitro fertilized (IVF) mice fed either the Western diet (WD; left) or the low-protein diet prenatally and a high-fat diet postnatally (LP/HF; right). NC and IVF mice covered a similar average distance on open arms when fed the WD. NC and IVF mice behaved in opposite ways when fed the LP/HF diet, with a trend toward less distance traveled for the NC mice and significantly greater distance traveled for the IVF mice. This interaction between conception and diet was highly significant (P < .0001). There was no difference between male and female mice within treatments, as illustrated by the parallel plots.
Results of ANCOVA Analysis.
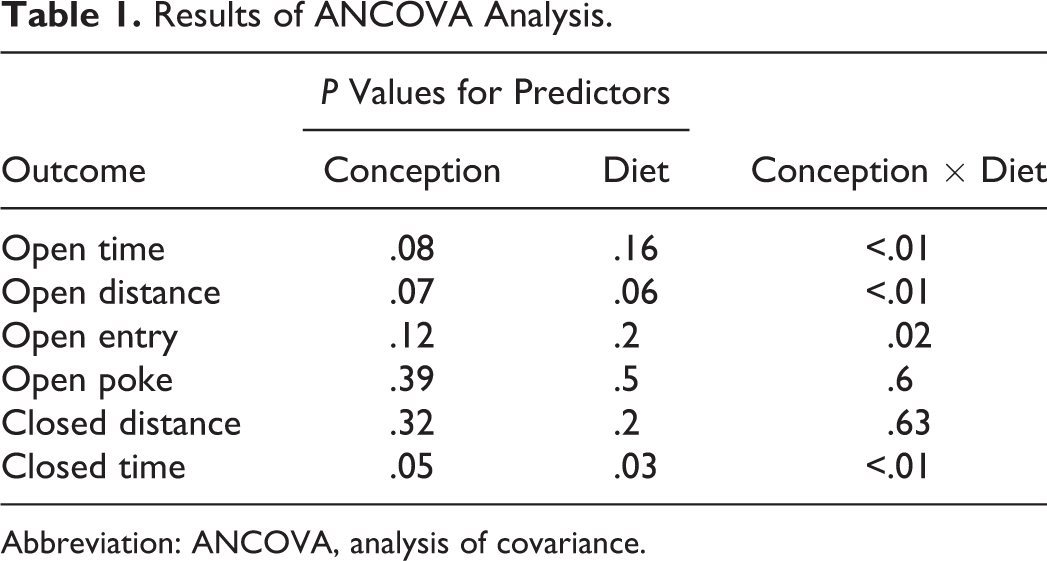
Abbreviation: ANCOVA, analysis of covariance.
Molecular Substrates for Diet-Induced Changes in Anxiety in IVF Mice
Quantitative real-time polymerase chain reaction was employed to assess whether altered EPM behavior was associated with changes in gene expression of selected neurotransmitters and related enzymes in selected brain regions. The genes and brain regions that were chosen have been shown to be involved in the stress response to anxiety and fear. 30 Given that males and females showed the same behavioral response to diet and conception and given the large number of potential samples, a subgroup of 8 male mice per group were chosen for analysis by q-PCR.
Comparisons were performed between NC and IVF mice from the same diet group. Although there were no significant behavioral differences between NC and IVF mice fed the WD, IVF-WD mouse brains showed increased expression of the Crf1 (1.16×; P < 0.05), Ucn gene in the caudate (6.17×; P < 0.05), and reduced levels of GABA markers genes in several structures (Figure 3A; Table S3).
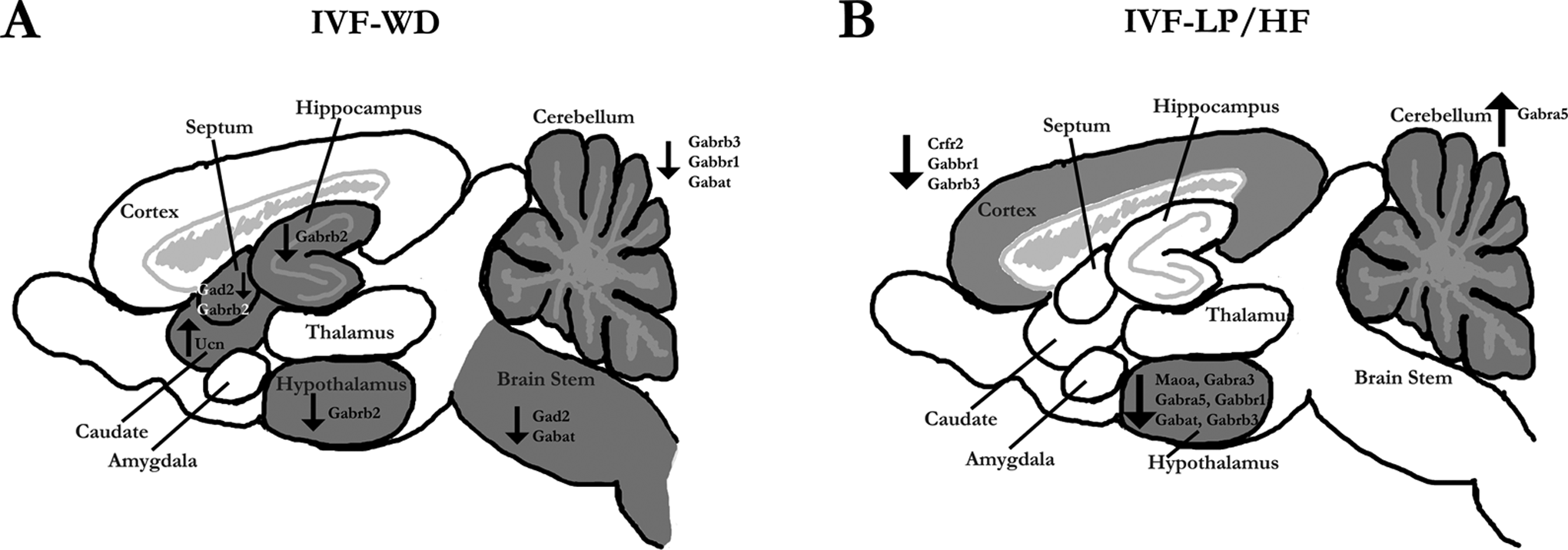
Gene expression in brain regions. Differential gene expression between male IVF and NC mice exposed to different dietary protocols. Areas shaded in gray are brain regions where statistically significant changes in gene expression, indicated by the gene name, were detected. A, Mice fed the Western diet (WD). B, Mice fed the low-protein diet prenatally and a high-fat diet postnatally (LP/HF) diet. Upward arrows mark an increase and downward arrows a decrease.
For the LP/HF diet groups, IVF mice exhibited significantly reduced expression of several genes in the hypothalamus and cortex compared to NC mice. In the hypothalamus Mao-A and Gabra3, Gabra5, Gabrb3, Gabbr1, and Gabat were reduced. In the cortex there were reductions in Crfr2 and Gabrb3 and Gabbr1 (Figure 3B; Table S3). The only increased gene expression was observed in the cerebellum where Gabra5 was increased 1.99× (Figure 3B; Table S3).
Discussion
The present study examined differences in anxiety-like behavior on the EPM and brain gene expression in mice that were NC or IVF and fed either a mouse version of our WD or a diet that was designed to exert additional stress both prenatally and postnatally (LP/HF). Both NC and IVF mice showed similar behaviors when they were fed the WD. By contrast, when exposed to the more stressful LP/HF diet, NC and IVF mice responded in opposite manners: NC mice by reducing EPM open arm exploratory activity and IVF by increasing it. The most relevant findings of this study is that neither conception nor stressful diet alone was sufficient to alter the behavior of the mice but that the combination of these factors acted together to increase mice activity on open space. In fact, analysis of covariance showed that the interaction of the method of conception and diet had a statistically significant effect on adult mice behavior as tested by the EPM, while controlling for age, sex and weight at testing. These behavioral response differences were accompanied by modest changes in gene expression in selected areas of the brain. In summary, when IVF mice were fed the LP/HF diet, they exhibited a decreased anxiety-like behavior compared to the NC controls.
In line with previous studies, body growth of mice was related to litter size. 31 Indeed IVF litters, particularly those from dams fed a low protein diet during gestation, were smaller in number than NC litters fed either diet, and IVF pups were significantly heavier than NC pups. Because of this significant difference, weight was controlled in the statistical analysis of behavior.
The EPM is broadly used and accepted as a measure of anxiety-like behavior in laboratory testing, as it uses 2 features of rodents’ nature to avoid open spaces and height. 29,32 –35 Anxious rodents tend to spend more time in protected enclosed arms, while those with lower anxiety spend more time exploring open alleys. Changes in EPM activity have been reported after variations in diets’ macronutrient composition. Rats fed a high-fat diet, 36 an isocaloric low-protein diet 37,38 or the offspring of mothers fed a low-protein diet during gestation 39 showed reduced levels of anxiety-like behavior on the EPM. In the present study, mothers were fed a low-protein diet during gestation and then the offspring were exposed to a high-fat diet after weaning. Both dietary protocols would be predicted to elicit an anxiolytic effect by increasing open arms exploratory activity. Interestingly, increased levels of open arm activity were observed only in the IVF mice.
Naturally conceived and IVF mice in the WD group showed no difference in their stress/anxiety-like response on the EPM (Figure 2). This is in contrast to studies reporting that after preimplantation culture, IVF mice showed higher levels of exploratory activity on open arms of the EPM 22 or open areas of the zero maze. 21 One explanation for this dissimilarity might be the different diets used or the different strains of mice. 40 Variation in the timing and conditions of culture may also account for differences in behavior. In the present study gametes were fertilized in vitro, while other groups have looked at normally fertilized embryos subjected to in vitro culture (IVC) 22 or cultured embryos only at the 2-cell stage. 21 We have previously shown that the gene expression profile of IVF and IVC embryos is significantly different. 18,20
Anxiety is a physiological response to stress characterized by neuroendocrine and behavioral components. The neuroendocrine response is commonly associated with the activation of the HPA axis but also rooted in an intermingled array of molecular mechanisms and brain structures. 41,42 MAO-A, corticotropin, and GABA are the most well-known modulators of anxiety and fear, 41 and changes in gene expression for these and related receptors and transporters have been correlated with altered levels of anxiety. 43 Brain structures best known for their role in the modulation of fear and anxiety are the amygdala-prefrontal cortex circuit, the cingulate cortex, brain stem, caudate nucleus, cerebellum, medial hypothalamus, and the septo-hippocampal system. 30
Although there were no differences in EPM open arm exploratory activity between IVF and NC mice fed the WD, a substantial increase in Ucn associated with a minor increase in Crfr1 messenger RNA (mRNA) levels was detected in the caudate nucleus of IVF male brains (Figure 3B and Table S3). The role of Ucn and Crfr1 receptors in the caudate nucleus is not known, however, a recent report has suggested an anxiolytic role for CRFR1 in the globus pallidus, also part of the basal ganglia. Increased expression of Ucn and Crfr1 in the caudate nucleus of IVF-WD mice was associated with reduced expression of GABA markers in various areas of the brain, including the cerebellum. The GABA receptors have long been known to play a key role in the regulation of emotional states such as anxiety and fear. 44,45
Naturally conceived and IVF mice displayed opposite responses to the LP/HF diet. The NC mice tended to have less exploratory activity in the EPM open arm with the stressful diet, while the IVF mice had significantly more activity in the open arms of the EPM with the stressful diet. The lower anxiety-like phenotype IVF-LP/HF males showed changes in gene expression only in the hypothalamus and cortex. Crfr2 gene expression was reduced in the cortex, Mao-a in the hypothalamus, and several GABA markers were reduced in both hypothalamus and cortex.
In the present study, Crfr2 mRNA was strongly reduced in the cortex of IVF males fed the LP/HF diet but unexpectedly was associated with increased levels of open arm activity. The observed discrepancy between Crfr2 expression levels and behavior on open arm may be due to a compensatory adaptive stress response of IVF mice to a second hit represented by the LP/HF diet or could be related to the multifaceted role played by the CRF in the stress response. 46,47 In fact, CRF activation is a common response to a variety of stressors. 48 Administration of CRF in the brain of rats is correlated with reduced exploratory activity and enhanced anxiety 49,50 mediated by CRFR1. In contrast, CRFR2 are upregulated by corticosterone 51 and exert a tonic inhibitory influence over anxiety-like states. 52 –54
The IVF-LP/HF males also showed a substantial reduction in Maoa gene expression in the hypothalamus, which may provide an alternative explanation for the increased open arm activity observed in these mice. The Maoa gene codes for the MAO-A enzyme, critical for the degradation of serotonin (5-HT) and norepinephrine (NE) in the brain. It plays a pivotal role in mood, social behavior and aggression 55 –57 but also in feeding behavior. 58 Reduced MAO-A activity in the hypothalamus is associated with reduced turnover of monoamines and increased levels of NE and corticosterone. 59,60
The expression of the Maoa gene was reduced in the hypothalamus of IVF mice fed the LP/HF diet. This is in contrast to a study where a high-fat diet in mice induced a 2-fold increase in Maoa mRNA in the hypothalamus. 58 Since MAO-A knockout mice show a significant reduction in risk assessment as well as unconditioned avoidance and escape, 61 we can postulate that this decrease in Maoa might explain a more disinhibited exploratory behavior in the IVF-LP/HF mice.
The hypothesis of a disinhibited phenotype is also supported by the reduced expression of GABA receptors and the GABA transporter genes found in both the hypothalamus and the cortex of IVF-LP/HF animals. The GABA is the main inhibitory neurotransmitter in the central nervous system and it has long been known to play a critical role in the regulation of emotional states. 44 Reduced levels of GABA mRNA have been found in the brain stem of rats fed a complex high-fat/high-carbohydrate diet. 62 Interestingly, Poulter et al. 63 observed that GABA receptor subunits were upregulated in stressor-reactive mice but downregulated in stress resilient mice. The reduced GABA markers expression detected in IVF-LP/HF males may reflect similarities between the stress resilient mice and the disinhibited phenotype suggested.
Of note, although changes in mRNA levels in the different brain regions selected for q-PCR analysis were modest, changes in affected neurons may be far greater due to dilution by unaffected neurons in the same area of the brain. Another limitation of this study includes the use of an in vivo group as a control as opposed to a flushed blastocyst group, that is, blastocysts conceived in vivo, flushed out of the uterus, and transferred to a recipient dam. 64 This method controls for the embryo transfer procedure and the in utero litter size. Therefore, we cannot rule out that some of the effects described in IVF mice are secondary to the embryo transfer procedure.
In conclusion, the present study shows that the stress/anxiety-like response on the EPM in mice can be significantly altered by the interaction of conception method with diet. This suggests that IVF functions as a first “hit,” or physiological insult, predisposing offspring to increased susceptibility to a postnatal second “hit.” This has also been shown in a recent article, where IVF mice exposed to a high-fat diet had reduced life span, as opposed to those fed a normal diet. 65 These data are particularly relevant considering the increasing use of ART to treat human infertility, and it underlines the relevance to optimize dietary protocols for women undergoing IVF and for IVF offspring in order to achieve the best health benefits. Future studies should explore whether the observed effects of anxiety-like behavior are present in mice of different strains and sexes 66 or if different interventions, like housing mice alone or in-group further affect their behavioral response. 67
Footnotes
Acknowledgments
The authors wish to thank Dr. Richard Weiner for his invaluable help in dissecting mice brain and Dr. Ivilin P. Stoianov for his help with the statistical analysis and comments on an earlier version of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: FS was partially supported by the Italian Ministry of University and Research, PRIN 2008.