
Abstract
Objective:
The nuclear receptor estrogen receptor α (ERα) is one of the key players in energy balance, insulin resistance, and trophoblast differentiation. We tested the hypothesis that gestational diabetes mellitus (GDM) alters expression of placental ERα in a cell type-specific manner and that this regulation may involve epigenetic changes.
Study Design:
Expression of ERα was analyzed by immunohistochemistry using the semiquantitative immunoreactive score in 80 placentas (40 GDM/40 controls). Quantitative real-time polymerase chain reaction (PCR) measured ERα messenger RNA (mRNA) in decidual tissue. Methylation-specific PCR was performed to analyze cytosine-phosphatidyl-guanine-island methylation of the ERα promoter.
Results:
Expression of ERα protein is upregulated (P = .011) in GDM in extravillous trophoblasts but not in syncytiotrophoblast. Gestational diabetes mellitus downregulated ERα in decidual vessels only in pregnancies with male but not female fetuses. Furthermore, mRNA of the ERα encoding gene estrogen receptor gene 1 (ESR1) was increased (+1.77 fold) in GDM decidua when compared to controls (P = .024). In parallel, the promoter of ESR1 was methylated only in decidua of healthy control individuals but not in GDM.
Conclusion:
Gestational diabetes mellitus affects expression of placental ERα in a cell type-dependent way, on epigenetic level. These data link GDM with epigenetic deregulations of ERα expression and open new insights into the intrauterine programming hypothesis of GDM.
Introduction
Estrogens exert a great variety of actions in almost every cell type and through diverse cellular and molecular pathways. Activation of intracellular estrogen receptors (ERs) induces their translocation into the nucleus where they act as transcription factors by binding to the estrogen response element, thus regulating gene transcription. 1
The ERs, ERα and ERβ, encoded by the ESR1 and estrogen receptor gene 2 (ESR2) gene, have been both involved in energy balance, 2 although evidence indicates ERα as the main mediator: Knockout of ERα as well as of the estrogen-synthesizing enzyme aromatase in mice results in obesity and insulin resistance. 3,4 Especially, ERα is related to glucose uptake and metabolism in different tissues including skeletal muscle, adipose tissue, liver, brain, and endocrine pancreas. 5 Besides these classical insulin target tissues, ERα is expressed also in maternal and fetal parts of the placenta. However, the exact function of ERα in this organ is yet to be identified. Transcriptional regulation of the leptin gene by ERα in the placental syncytiotrophoblast (SCT) 6 suggests a role of ERα in hormonal regulation of metabolism.
Gestational diabetes mellitus (GDM) is defined as glucose intolerance first diagnosed during pregnancy. It is characterized by insulin resistance with insulin levels insufficient to compensate the increased demand. 7 Between 5% and 15% of all pregnancies are currently affected by GDM. 8 As a consequence of the worldwide obesity epidemic, GDM prevalence is further rising. 9 Gestational diabetes mellitus is associated with short- and long-term complications for the offspring: Short-term consequences include perinatal complications due to high birth weight and fetal hyperinsulinemia. 10,11 Long-term consequences include a higher risk to develop obesity, metabolic syndrome, and type 2 diabetes in later life. 12 Epidemiologic studies and animal experiments link the susceptibility for these metabolic diseases in the offspring to epigenetic changes by the diabetic environment in utero. 13,14 Chromatin modifications due to DNA methylation are one of the underlying mechanisms for intrauterine programming by causing persistent changes in gene expression. 15 Methylation of cytosine-phosphatidyl-guanine (CpG) sites within the promoter region of certain genes can reduce gene expression 16 –19 and, vice versa, hypomethylation of promoter regions allows increased gene expression. 20
The placenta is a fetal organ and exposed to the same intrauterine environment as the fetus. In a pregnancy complicated by GDM, the placenta is affected by maternal metabolic derangements. Due to its critical role in fetal growth and nutrient exchange, the placenta is a great model to study epigenetic changes in metabolic programming in fetal life. 15 Gestational diabetes mellitus has epigenetic effects on genes preferentially involved in metabolic disease pathways, supporting the notion that DNA methylation is involved in fetal metabolic programming. 9,21 –24 The placenta is one of the tissues in which GDM alters methylation. 25,26
The known regulation of the leptin gene by ERα in the placental SCT in parallel with increased leptin levels in GDM prompted us to hypothesize changes in placental expression of ERα in GDM and that these changes may result from methylation changes of the ESR1 promoter region.
Materials and Methods
The study was approved by the ethics committee of the Ludwig-Maximilian-University (LMU) Munich, Germany, and written informed consent was obtained from the patients. Placental tissue was obtained from 80 women. All women underwent an oral glucose tolerance test 27 at weeks 24 to 28 of gestation. According to the criteria of the German Diabetes Society (capillary whole blood; fasting glucose >90 mg/dL, 1 hour > 180 mg/dL, 2 hours > 155 mg/dL; GDM was defined with 2 values above this limits), 40 women were diagnosed with GDM. Immediately after delivery, the samples were taken from the central part of the placenta. In addition, a cotyledon structure showing sufficient blood supply was chosen. Calcified, necrotic, and visually ischemic areas were excluded from collection. Two pieces (2 × 2 × 2 centimeter) were dissected, containing decidua, extravillous, and villous trophoblasts. Each sample was divided in 2 parts. One part was further separated into decidual and villous tissue, frozen separately, and stored at −80°C. A second part of placental tissue was fixed in 4% buffered formalin for 24 hours and thereafter embedded in paraffin. Thus, immunohistochemistry, gene expression, and DNA methylation analysis were performed with tissues from the same sampling site.
All patients in the GDM group were treated with insulin. The patients were monitored at least once a week at the Diabetes Center of the Department of Internal Medicine, LMU; 75% of patients were under good glycemic control according to their mean blood glucose. Demographic and clinical data of the study population are summarized in Table 1. Perinatal and clinical data of the study group were published recently. 28
Clinical Details of the Patients With Gestational Diabetes Mellitus (GDM) and of the Normal Control Group (Mean ± SD).a
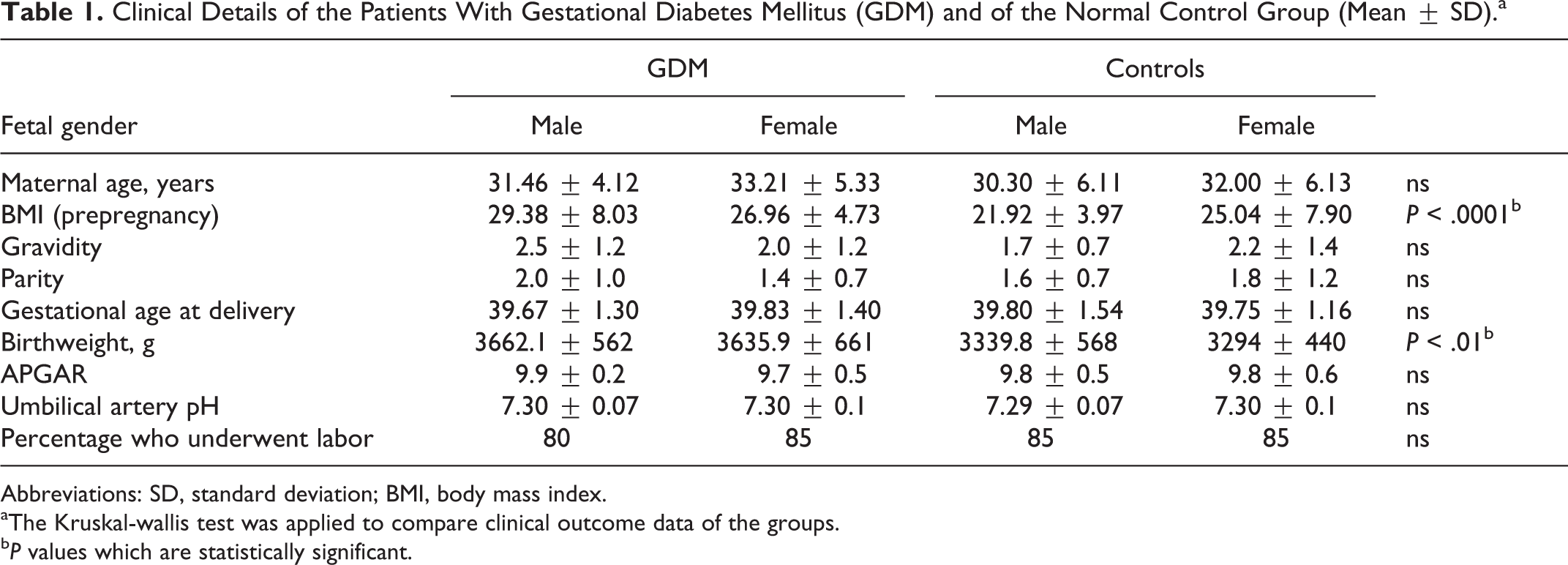
Abbreviations: SD, standard deviation; BMI, body mass index.
aThe Kruskal-wallis test was applied to compare clinical outcome data of the groups.
b P values which are statistically significant.
Immunohistochemistry
Sections were dewaxed in xylol (20 minutes), and endogenous peroxidase was quenched with 3% hydrogen peroxide (Merck, Darmstadt, Germany) in methanol (20 minutes). Following rehydration in a descending series of alcohol, epitope retrieval was performed in a pressure cooker (5 minutes) using sodium citrate buffer (0.1 mol/L citric acid/0.1 mol/L sodium citrate pH 6.0). Slides were then cooled to room temperature and washed in distilled water followed by phosphate-buffered saline. Then, specimens were blocked (3 minutes at room temperature) with Power Block Universal Blocking Reagent (BioGenex, Fremont, California). Slides were incubated with the primary antibody (anti-ERα antibody, Immunotech, Marseille, France) for 1 hour at room temperature. The Vectastain Elite Mouse IgG ABC-Kit (Vector Laboratories, Burlingame, California) was used for visualization. Sections were stained with 3, 3′-diaminobenzidine (Dako, Glostrup, Denmark) and counterstained with hemalaun. Sections were dehydrated and cover-slipped with Shandon Consul Mount Medium (Thermo Fisher Scientific, Waltham). For the negative control, the primary antibody was replaced by species-specific isotype control antibody. All placentas (40 GDM and 40 controls) were examined. Five representative images per placenta were taken with a digital camera system (JVC, Victor Company of Japan, Yokohama, Japan), and cells were analyzed with a Leitz Diaplan light microscope (Leitz, Wetzlar, Germany). Staining intensity was investigated semiquantitatively by an immune-reactivity score (IRS). This was calculated by multiplying the percentage of stained cells (0 = no staining, 1 = ≤ 10% of the cells, 2 = 11% to 50% of the cells, 3 = 51%-80% of the cells, and 4 = ≥ 81% of the cells) and the different staining intensities (graded as no staining = 0 [negative], weak staining = 1, moderate staining = 2, and strong staining = 3). The intensity and distribution pattern of the immunochemical staining reaction were evaluated by 2 independent blinded observers. 29,30 In 2 cases (n = 2.5%), the evaluation of the 2 observers differed. These cases was reevaluated by both observers together. After the reevaluation, both observers came to the same result. The concordance before the reevaluation was 97.5%.
RNA Isolation, Processing, and Real-Time Polymerase Chain Reaction
Total RNA was extracted from the decidual tissue using the RNeasy Lipid Tissue Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s protocol. Quantification and evaluation of the purity of the isolated RNA were carried out with a NanoPhotometer (Implen, Munich, Germany). A ratio of absorbance at 260 nm/280 nm ˜ 2.0 was accepted as pure. The reverse transcription was performed with the High Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Weiterstadt, Germany) according to the manufacturer’s protocol in a mastercycler gradient (Eppendorf, Hamburg, Germany) for 10 minutes at 25°C, 2 hours at 37°C, 5 seconds at 85°C, and 4°C on hold. The real-time polymerase chain reactions (PCRs) were accomplished in optical 96-well reaction microtiter plates covered with optical caps in a volume of 20 µL, containing 1 µL TaqMan Gene Expression Assay 20× (Hs01045840_m1 for VDR, Applied Biosystems), 10 µL TaqMan Univeral PCR Master Mix 2x (Applied Biosystems), 8 µL H2O (DEPC treated DI water, Sigma, Taufkirchen, Germany), and 1 µL template. For thermal cycling, an ABI PRISM 7500 Fast (Applied Biosystems) was used for 20 seconds at 95°C, followed by 40 cycles of amplification for 3 seconds at 95°C, and 30 seconds at 60°C. Quantification was performed using the 2−ΔΔCT method with β-actin (Hs99999903_m1) as housekeeping gene. β-Actin was chosen as it revealed a stable expression in placenta and decidua in normal versus GDM pregnancies.
DNA Extraction and Bisulfite Conversion
For the extraction of the genomic DNA, 25 mg placental tissues were cut into small pieces and incubated with lysis buffer and proteinase K (both Macherey-Nagel, Düren, Germany) at 56°C overnight. Further steps of DNA isolation were carried out using the NucleoSpin Tissue Kit (Macherey-Nagel) according to the manufacture’s protocol. DNA concentrations were measured (Nanodrop; PeqLab, Erlangen, Germany) and 2 µg of each DNA sample were treated with the EpiTect Bisulfite Kit (Qiagen, Hilden, Germany) according to the manufacture’s protocol. Bisulfite treatment of DNA leads to a conversion of unmethylated cytosine to uracil while leaving methylated cytosine intact.
Estrogen Receptor Promoter Analysis With Real-Time Methylation-Specific PCR
Promoter analysis was performed on a promoter region according to former investigation by our group. 31,32 In brief, Lapidus et al 33 divided the CpG island of the ER promoter into 6 regions (ER1-ER6) and tested them for methylation and functional relevance in gene expression. The loci ER3, ER4, and ER5 (−310 and −375 bp upstream of the transcription start site) showed the highest negative correlation between methylation status and ER expression. Comparison of the different primer sets showed the region ER4 of the CpG island to be most specific for gene expression. 31,33 The primers and probe used here were published recently 34 and given in Table 2.
Sequences and Concentrations of Primer and Probes Used for rt-MSP.

Abbreviations: rt-MSP, real-time methylation specific polymerase chain reaction; FAM, 6-carboxyfluorescein; TAMRA, carboxy-tetramethyl-rhodamine.
Real-time methylation specific PCR (rt-MSP) was performed to distinguish between methylated and unmethylated promoters: In optical 96-well plates covered with optical caps, the rt-MSP was accomplished in a volume of 25 µL, containing 2.5 µL forward primer (Applied Biosystems), 2.5 µL reverse primer (Applied Biosystems), 3 µL TaqMan probe (Applied Biosystems), 12.5 µL TaqMan Univeral PCR Master Mix 2x (Applied Biosystems), 3 µL H2O (DEPC treated DI water, Sigma, Taufkirchen, Germany), and 2 µL bisulphite-treated DNA. Thermal cycling conditions were 20 seconds at 95°C, followed by 40 cycles of amplification for 3 seconds at 95°C, and 30 seconds at 60°C. Each plate contained patient samples and water blanks as well as positive (MDA-MB-231) and negative controls (Michigan Cancer Foundation-7 [MCF-7] cell line). The ABI PRISM 7500 Fast (Applied Biosystems) was used for the PCR.
Statistical Analysis
Statistical analysis was performed using the nonparametrical Mann-Whitney U signed rank tests and the t-test for comparison of the means. Univariate and multiple linear regression models were used to analyze the associations of clinical outcome data with IRS. The models were adjusted for potential and known confounders (maternal age, parity, maternal body mass index (BMI), fetal sex, gestational age, and umbilical artery pH). 35 Statistical significance was assumed at P values <.05.
Results
Immunoreactivity of ERα in Normal and GDM Placenta
Immunohistochemistry identified ERα in maternal decidual endothelium, in the SCT, and in the extravillous trophoblasts (EVTs) that invaded into the decidua. In ERα expressing cells, ERα was located in the nucleus and in the cytoplasm. Neither villous macrophage nor feto–placental endothelium was stained for ERα.
When immunoreactivies were compared semiquantitatively between normal and GDM placentas using the IRS, SCT (mean IRS 1 vs 1, P = .66, Figure 1A-C) showed no expression differences between both the groups. However, in the maternal endothelium, GDM downregulated ERα in decidual vessels, but this GDM effect was only found in pregnancies with male (control IRS 3.5 vs GDM IRS 1, P = .01) but not female fetuses (Figure 1D-F). The ERα protein signal was lower in the EVT (mean control IRS 2 vs GDM IRS 4, P = .011) in GDM when compared to normal pregnancy (Figure 2A-C). In both EVT and SCT, ERα immunoreactivity was not different between fetal sexes.
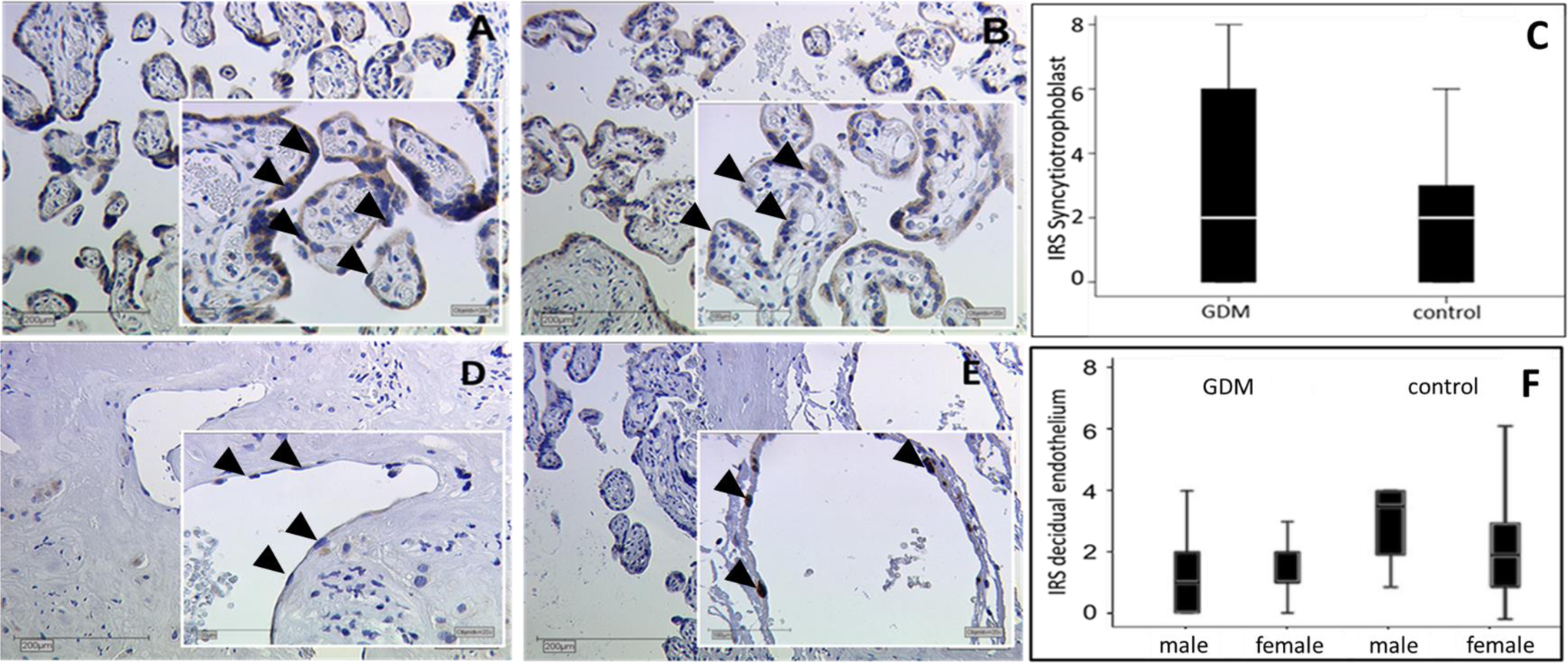
Representative slides of immunhistochemical staining for ERα expression in the SCT of GDM positive placentas (A) and controls (B), endothelium of GDM positive decidua (D) and controls (E), both with male neonates. Pictures were taken at a magnification of 10× and 25×. Arrowheads indicate staining of the ERα in the SCT of GDM (A) and control tissue (B). Arrowheads indicate different staining of ER in cell nuclei of endothelial tissue in the decidua (GDM [D] and control [E]). The immune-reactivity score (IRS) in GDM and control placentas for SCT is shown in the boxplot (C), for endothelium gender specific in boxplot (F). The boxes represent the range between the 25th and 75th percentiles with a horizontal line at the median. The bars delineate the 5th and the 95th percentiles. Staining was performed in 40 GDM and 40 control placenta. ERα indicates estrogen receptor α; SCT, syncytiotrophoblast; GDM, gestational diabetes mellitus. (The color version of this figure is available in the online version at http://rs.sagepub.com/.)
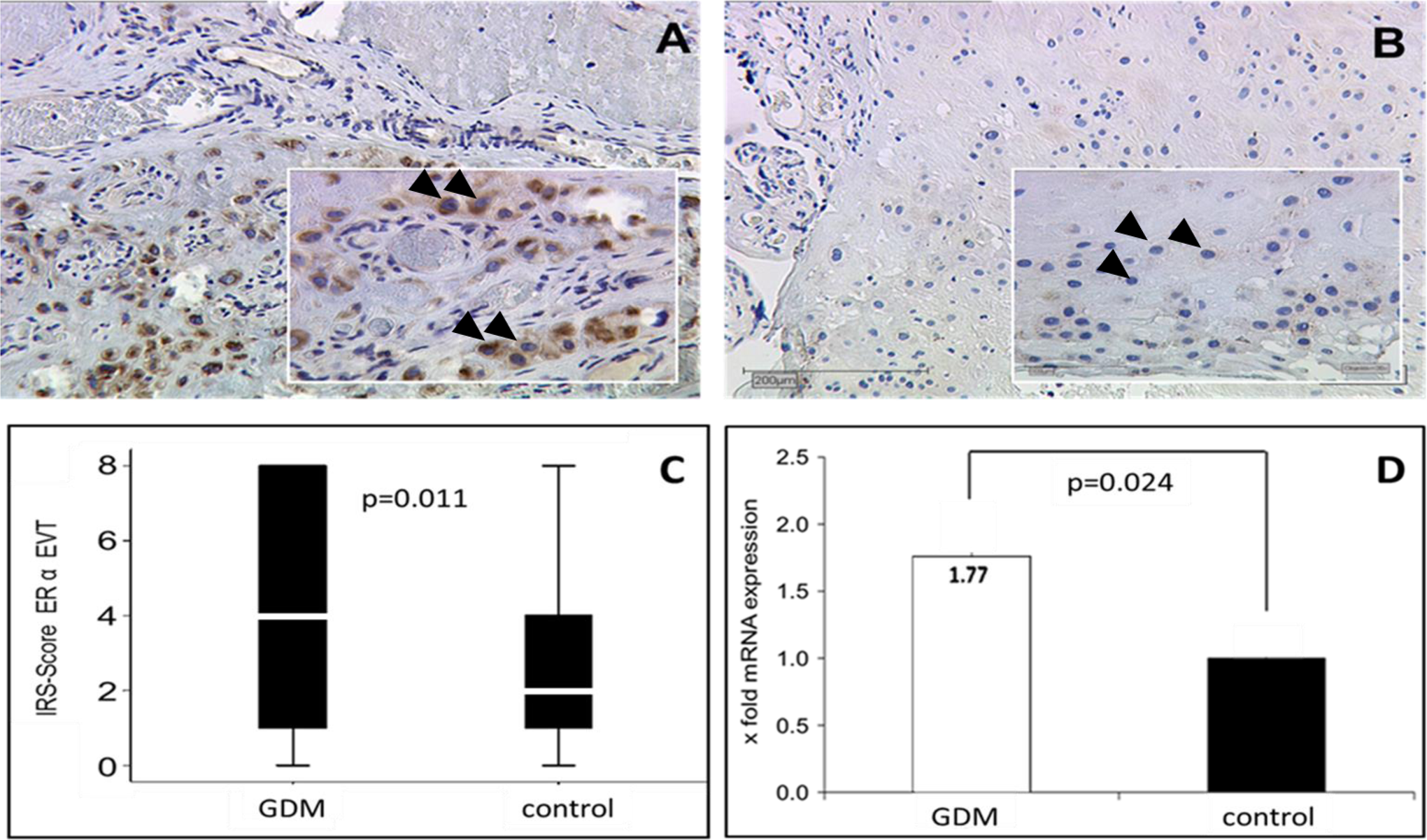
Representative slides of immunhistochemical staining for ERα expression in the EVT of GDM-positive placentas (A) and controls (B). Arrowheads indicate staining of the ERα in the EVT of GDM (A) and control tissue (B). Pictures were taken at a magnification of 10× and 25×. The immune-reactivity score (IRS) in GDM and control placentas for EVT is shown in the boxplot (C). The boxes represent the range between the 25th and 75th percentiles with a horizontal line at the median. The bars delineate the 5th and the 95th percentiles quantitative RT-PCR: significant (P = .024) upregulation (1.77-fold) of ERα mRNA expression in GDM positive placentas compared to normal controls (D). Staining and quantitative RT-PCR were performed in 40 GDM and 40 control placenta. ERα indicates estrogen receptor α; EVT, extravillous trophoblast; GDM, gestational diabetes mellitus; RT-PCR, real-time quantitative polymerase chain reaction; mRNA, messenger RNA. (The color version of this figure is available in the online version at http://rs.sagepub.com/.)
Expression of ERα Messenger RNA in Normal and GDM Decidua
In order to determine whether increased ERα protein in GDM EVT was a result of increased gene expression, real-time quantitative RT-PCR was performed. Because only EVT but no other placental or decidual cell type showed altered ERα protein levels, decidual tissue containing EVT and decidual stromal cells was separated from villous placental tissue and qPCR was performed. Indeed, expression of ERα messenger RNA (mRNA) was increased about 1.8-fold in decidua of GDM women when compared to normal controls (P = .024; Figure 2D. In parallel to the low abundance of maternal endothelial cells and the high abundance of EVT in decidual tissue, no differences in ERα transcripts related to fetal sex were found in the total tissue.
Methylation of ESR1 Promoter in Normal and GDM Placenta
Altered gene expression may result from promoter methylation changes. To determine whether higher levels of ESR1 mRNA in GDM may result from promoter hypomethylation, we performed rt-MSP for highly methylation sensitive CpG sites in the ESR1 promoter. Decidual tissue of controls revealed ESR1 promoter methylation at this site, whereas in all GDM samples these CpGs were unmethylated. Methylation was not different between tissue from male versus female pregnancies.
Statistical Confounder Analysis
As the study groups differed in BMI and birth weight (see Table 1), we analyzed the statistical associations of these potential confounders with ERα expression. Neither birth weight nor BMI was associated with ERα IRS univariately and in addition not after adjusting for other confounders (maternal age, parity, BMI, fetal sex, gestational age, and umbilical artery pH).
Discussion
In the present study, we identified elevated levels of ERα protein and mRNA in the EVT of women with GDM. Furthermore, this increased expression of ERα in GDM was paralleled by a demethylation of a CpG in the ESR1 promoter, which is essential for transcriptional activity of ESR1.
This is the first study to demonstrate a link between increased ERα levels and ESR1 promoter hypomethylation in placenta in GDM. In line with our results, a previous study showed an upregulation in ERα in decidual GDM trophoblastic tissue. 36 A later study found no differences in the ESR1 expression in placentas of women with GDM. 37 However, the absence of difference in ESR1 expression in the latter study may be due to tissue sampling only from the fetal side of the placenta, as we found expression changes only at the maternal side of the placenta. Fetal endothelium was negative for ERα immunostaining, which parallels findings on absent ERα in human umbilical vein endothelial cells. 38
Previous studies already demonstrated that the proinflammatory environment of GDM causes promoter demethylation including genes linked to fetal growth and nutrition. 18,23,26,39 Furthermore, the link between ESR1 promoter demethylation and upregulation of ESR1 expression was established in rat trophoblast cells using 5-aza-2-deoxycytidine treatment. 40
The function of ERα in the placenta and in particular in EVT is unknown. In the SCT, the ERα receptor mediates the estrogen-stimulated expression of leptin 41 and aromatase, 42 suggesting a role in hormone secretion. In addition, a function in glucose metabolism may be hypothesized since ERα is a key regulator of glucose metabolism in adipose tissue and muscle. 1,38,39 Extravillous trophoblast is the placental cell type that invades the maternal uterus, anchors the placenta, and remodels uterine vessels to increase blood flow. Thus, the EVT is known as the site of maternal–fetal immune interactions. 43,44 The role of ERα in EVT function remains to be established, but based on its immunomodulatory function outside pregnancy, 45,46 it is tempting to hypothesize a similar role also in pregnancy. Insulin resistance is associated with a chronic inflammatory reaction in GDM placenta. 47 In the light of this proinflammatory environment of GDM and, thus, increased immune modulation, ERα upregulation may be necessary to maintain immune tolerance.
We do not know why only the EVT, but not the SCT, responds to maternal GDM with increased ERα expression. We hypothesize that local decidual factors rather than circulating maternal factors are the underlying reason. Interestingly, our earlier studies showed that in GDM the EVT is the site of strongest change in nuclear receptor expression: While VDR is upregulated in EVT of GDM placentas, 48 peroxisome proliferator-activated receptor (PPAR) γ is downregulated. 28 Estrogen receptor α physically binds to PPARγ and inhibits its transcriptional activity, 49 which may further attenuate PPARγ in GDM.
The placenta is regarded as a model to study the effect of the intrauterine environment on epigenetic changes. We found demethylation of a CpG island in the ESR1 promoter. However, we have to keep in mind that methylation changes in the placenta may not necessarily reflect events in other target tissues of metabolic programming 23 as epigenomes can differ between cells and tissues. 50
Estrogen is a crucial regulator in fat metabolism: Knockout of ERα in mice results in obesity, 4 and ESR1 expression in adipose tissue is inversely correlated with BMI. 51 Since BMI is the most significantly different confounder between the GDM and the control group in our study, careful attention was paid in the statistical analysis to identify a potential BMI effect on placental ERα protein expression. This analysis clearly showed that BMI was not associated with placental ERα IRS in univariate analysis (P = .96) nor after adjusting for other possible confounders (P > .05).
A striking finding was the sex-specific effect of GDM on ERα protein expression changes in maternal decidual endothelial cells: While in controls deciduas from pregnancies with male fetuses had higher levels of endothelial ERα expression than deciduas from female pregnancies, this sex difference disappeared in the GDM group. Estrogen was shown to downregulate ESR1 expression in uterine endothelium. 52 This may explain lower ERα expression in the female decidua. Interestingly, gender differences were found to decline in patients with type 2 diabetes. 53 Our data reflect this clinical observation: Metabolic deteriorations curb the gender-related differences, since the sex-specific differences in decidual endothelium disappeared in GDM.
In summary, the present study characterizes changes in expression of ERα in the EVT in GDM. In contrast, the SCT showed no changes. Maternal endothelial ERα is subject to sex-specific regulation, which disappears in GDM. Estrogen receptor α promoter hypomethylation in the GDM group may contribute to the reduced ERα levels.
Footnotes
Acknowledgments
The authors thank Simone Hofmann and Christina Kuhn for excellent technical support.
Authors’ Note
Julia Knabl and Ursula Hiden contributed equally to this work.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.