
Abstract
Background:
Cytochrome P450 aromatase (encoded by the CYP19A1/aromatase gene) plays a critical physiologic role in endometriosis. Metformin is known to suppress prostaglandin E2 (PGE2)-induced CYP19A1 messenger RNA (mRNA) expression in human endometriotic stromal cells (ESCs). However, the possible mechanism behind this suppression remains to be determined.
Methods:
In this study, ESCs were cultured with metformin, PGE2, and adenosine monophosphate (AMP)-activated protein kinase (AMPK) inhibitors. Expression of CYP19A1 mRNA and aromatase activity were measured by quantitative polymerase chain reaction and aromatase activity assay, respectively. The binding of the cyclic AMP response element-binding (CREB) protein to CYP19A1 promoter II (PII) was assessed by chromatin immunoprecipitation assay.
Results:
We demonstrated that metformin downregulated the expression of aromatase mRNA (32%) and activity (25%) stimulated by PGE2 (4.18-fold and 2.14-fold) in ESCs via stimulation of AMPK. Following PGE2 treatment, there was a marked increase in CREB binding to aromatase PII, while metformin attenuated the above-mentioned stimulation by 67%.
Conclusion:
Metformin could inhibit PGE2-induced CYP19A1 mRNA expression and aromatase activity via AMPK activation and inhibition of CREB to CYP19A1 PII in human ESCs. The results of the present study suggest that metformin may have unique therapeutic potential as an antiendometriotic drug in the future.
Keywords
Background
Endometriosis is an estrogen-dependent disease characterized by the presence of endometrium-like tissue outside the uterine cavity, primarily on the ovaries and pelvic peritoneum. It is a systemic disorder affecting 1 in 10 women of reproductive age and is one of the most common causes of chronic pelvic pain, dysmenorrhea, and infertility. 1,2 Only half of all women with a previous diagnosis of endometriosis achieve pain relief upon receiving hormone therapy or conservative surgery. 1 Therefore, it is necessary to understand the underlying mechanisms in order to develop novel and effective methods for the treatment of endometriosis.
The significance of estrogen biosynthesis in endometriosis is exemplified by the clinical observations that local estrogen production in endometriotic tissue is essential for the growth of endometriosis. 3 –6 Cytochrome P450 aromatase (P450arom encoded by the CYP19A1/aromatase gene), which catalyzes the final step of estrogen production by converting C19 steroids to estrogens, 7 is the key enzyme for estrogen biosynthesis. 8 CYP19A1 is expressed in the endometriotic stromal cells (ESCs), whereas they are undetectable in the eutopic endometrial stromal cells of disease-free women. 9,10 Moreover, prostaglandin E2 (PGE2) has been shown to induce higher aromatase activity and CYP19A1 messenger RNA (mRNA) levels in cultured ESCs, compared to eutopic endometrial stromal cells where significant CYP19A1 mRNA level and aromatase activity could not be detected before or after PGE2 treatment. 9 The expression of CYP19A1 is partly regulated by the alternative use of tissue-specific promoters in the placenta (distally located promoter I.1), ovary (classically located proximal promoter II [PII]), and adipose tissue (promoters I.4, I.3, and II). 11 Expression of CYP19A1 in PGE2-stimulated ESCs is primarily regulated by the classically located promoter II (PII). 9 Furthermore, PGE2 has been shown to activate the PGE2 receptor 2 (EP2)-coupled protein kinase A (PKA) pathway and the EP2-induced phosphorylation of the cyclic AMP (cAMP) response element-binding protein (CREB), 12 which when phosphorylated binds to a cAMP response element (CRE) in the gonad-specific PII of CYP19A1 gene. 13,14
Metformin is a widely used antidiabetic agent that improves insulin sensitivity and is used for the treatment of polycystic ovary syndrome (PCOS). 15 Metformin may also reduce the obesity-associated inflammatory status and other inflammatory responses 16 –18 and affect the steroidogenesis in ovarian granulosa and thecal cells. 19,20 In a recent study in a rat model, metformin was shown to cause a statistically significant regression of endometriotic implants, and the effects of metformin on endometriotic tissue were at least comparable to those of letrozole. 21 Metformin is believed to act primarily via stimulation of AMP-activated protein kinase (AMPK), 22 which is a key regulatory enzyme in cellular energy homeostasis. 23 The AMPK is a serine/threonine protein kinase consisting of a catalytic α subunit and regulatory β and γ subunits, each of which is encoded by 2 or 3 distinct genes (α1, α2; β1, β2; γ1, γ2, and γ3). The 2 isoforms of α (α1 and α2) and β (β1 and β2) subunits have very similar structures. The activity of AMPK results from phosphorylation of the Thr172 residue of the α subunit. 24 In addition, metformin has been shown to induce mitogen-activated protein kinase kinase (MEK)/extracellular-signal-regulated kinases (ERK) phosphorylation. 25,26
This study aimed to determine whether metformin could inhibit expression of CYP19A1 mRNA and aromatase activity in human ESCs and to determine the possible molecular mechanism responsible for this effect.
Methods
Participants and Primary Cell Culture
An ovarian endometrioma is a cystic ovarian lesion composed of endometrium-like tissue in the cyst wall with bloody fluid in the cyst. Ectopic endometrial tissues from the cyst walls of ovarian endometriomas and eutopic endometrial tissues were obtained from 12 women with endometriosis immediately after they underwent surgery, composing 12 self-control pairs. Normal endometrial tissues (n = 8) were from endometriosis-free women. The age range of the participants was 23 to 40 years. The experimental procedures were approved by the Institutional Review Board of the First Hospital of Peking University (No.2014[789] and No.2014[790]), and signed informed consents for use of the samples were obtained from each patient. All the patients had regular menstrual cycles, and none received any preoperative hormonal therapy. All the samples were histologically confirmed, and the phase of the menstrual cycle was determined by preoperative history and histological examination. Half of the tissue samples were in the proliferative phase and the other half in the secretory phase. Human ESCs and endometrial cells (EMs) were isolated from tissue samples using the protocol described previously by Ryan et al, 27 with minor modifications. 9 Briefly, the endometriotic tissues were rinsed with sterile phosphate-buffered saline (PBS) to remove blood cells, minced finely, and digested with collagenase (Sigma, St Louis, Missouri) and DNase (Sigma) at 37°C for 60 minutes. Epithelial cells were removed by filtration of stromal cells through 70- and 20-μm sieves, after which the cells were suspended in Dulbecco Modified Eagle medium (DMEM)/F12 (1:1; HyClone, Logan, Utah) with 10% fetal bovine serum (GIBCO/BRL, Grand Island, New York), 100 U/mL penicillin (Lonza, Basel, Switzerland), 100 U/mL streptomycin (Lonza), and 250 ng/mL amphotericin B (Lonza) at 37°C in a humidified atmosphere containing 5% CO2. The culture medium with the size of 100 mm in diameter was changed every 3 days.
Drug Treatments
When the ESCs or EMs were approaching confluence, the cells were washed thrice in sterile PBS and then serum starved for 24 hours in phenol red-free medium containing penicillin (100 U/mL) with streptomycin (100 U/mL). The cells were then incubated in serum-free DMEM/F-12 medium containing 100 μmol/L metformin (Calbiochem, San Diego, California) or vehicle for 24 hours, and then stimulated with 1 μmol/L PGE2 (Sigma) or vehicle for 24 hours. After starvation overnight, the ESCs were preincubated with an AMPK inhibitor Compound C (CC, 5 μmol/L; Sigma) or a MEK/ERK inhibitor PD98059 (PD, 25μmol/L; Sigma) for 1 hour and then treated with or without metformin for 24 hours, after which they were cultured with or without PGE2 for another 24 hours.
RNA Extraction and Quantitative Analysis by Real-Time Polymerase Chain Reaction
Total RNA was isolated from stromal cells with TRIzol (Invitrogen, Carlsbad, CA), according to the manufacturer’s protocol. In brief, 2 µg of total RNA was subjected to reverse transcription using an ABI High Capacity cDNA Archive kit (Applied Biosystems, Foster City, California). Real-time (RT) quantitative polymerase chain reaction (qPCR) was performed using an ABI 7500 Sequence Detection system and an ABI Power Sybr Green gene expression system (Applied Biosystems) to quantify CYP19A1 (GenBank Accession No. NM_007982.1) mRNA. Human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA was used as an internal control. Relative quantification for all transcripts was analyzed by the comparative threshold cycle method described previously. 28 The forward and reverse primers for CYP19A1 mRNA (coding region-specific) for RT-PCR were 5′-CACATCCTCAATACCAGGTCC-3′ and 5′-CAGAGATCCAGACTCGCATG-3′. The forward and reverse primers for GAPDH mRNA were 5′-GAAGGTGAAGGTCGGAGTC-3′ and 5′-GAAGATGGTGATGGGATTTC-3′.
Knockdown of Small-Interfering RNA
Endometrial stromal cells were cultured in growth medium as described previously to achieve approximately 50% to 60% confluence at the time of transfection. Transfections were performed using a nontargeting negative control small-interfering RNA (siRNA; Invitrogen) or siRNAs against human AMPKα1 (Invitrogen) at a final concentration of 100 nmol/L using Lipofectamine RNAiMAX (Invitrogen) in Opti-MEM reduced serum medium (Invitrogen). Thirty-six hours after transfection, the cells were serum starved for 12 hours, treated with 100 μmol/L of metformin for 24 hours, and stimulated with 1 μmol/L PGE2 for 24 hours. After this, the cells were processed for RT-PCR and aromatase activity assay.
Western Blot Analysis
The ESCs were washed in ice-cold PBS and lysed in mammalian protein extraction reagent (KeyGen Biotech, Nanjing, China) supplemented with protease inhibitor (Amresco, Solon, Ohio). Protein concentrations were determined using a micro-BCA protein assay kit (KeyGen). Equal amounts of protein (at least 30 μg) were resolved on 10% polyacrylamide gel using sodium dodecyl sulfate-polyacrylamide gel electrophoresis, transferred onto nitrocellulose membranes, and incubated with antihuman AMPKα antibody (1:1000 dilution; Cell Signaling, Boston, Massachusetts). Anti-GAPDH antibody was used as the loading control. The results were visualized on a digital imaging system (KODAK, Jacksonville, Florida).
Aromatase Activity Assay
The aromatase activity of ESCs was measured by a [3H]H2O release assay as described previously. 29 In each well, 60 pmol of [3H]androstenedione (PerkinElmer Life Sciences, Waltham, Massachusetts) and 240 pmol of cold androstenedione (Dr Ehrenstorfer, Augsburg, Germany) were added to 3 mL of serum-free DMEF/F-12 that covered cells in culture dishes. Experiments were conducted when cells reached 80% confluency. After starvation overnight, the cells were incubated with metformin for 24 hours and then stimulated with PGE2 for another 24 hours. Each treatment was performed in triplicate. After 42 hours of incubation after the drug therapy, a mixture of labeled and cold androstenedione was added to each well, and the cells were incubated for another 6 hours. [3H]Androstenedione conversion to [3H]estrogen was stopped by adding 10% trichloroacetic acid. Steroidal compounds containing unconverted [3H]androstenedione were removed from the mixture by first mixing with 4 mL of chloroform followed by centrifugation at 3000 rpm. The upper aqueous layer was removed and mixed with dextran-coated charcoal. Charcoal was precipitated by centrifugation. From each tube, 2 mL of clear solution was taken into 10 mL of scintillation vial and counted in a scintillation counter (Beckman Coulter LS 6500; Beckman Coulter Inc, Brea, California).
Chromatin Immunoprecipitation Assay
For the chromatin immunoprecipitation (ChIP) assay, the ESCs were cultured in 15-cm dishes to confluence, serum starved for 16 hours, incubated in the presence or absence of 100 μmol/L metformin for 11 hours, and then stimulated with 1 μmol/L PGE2 for 1 hour. The ChIP assay was carried out using a ChIP assay kit (Pierce, Rockford, Illinois) according to the manufacturer’s protocol. Briefly, the cells were harvested and subjected to ChIP with either anti-CREB antibody (Cell Signaling) or control antibody (immunoglobulin G; Santa Cruz Biotechnology, Santa Cruz, California) at 4°C overnight with rotation. The purified DNA was then analyzed by semiquantitative PCR and RT-qPCR using primers flanking the CREs of CYP19A1 PII (F: 5′-GCCTAAACAAAACCTGCTGATGA-3′ and R: 5′-CTTATCATCTTGCCCTTGAGTGG-3′). The obtained PCR products were 139 bp.
Statistical Analyses
All experiments were carried out at least 3 times using samples from different women. Statistical analyses were performed by 1-way analysis of variance (ANOVA) followed by Tukey multiple comparison test. A P value of <.05 was considered statistically significant. All the values are given as the mean, with the bars showing the standard error of mean.
Results
Metformin Reduces the PGE2-Stimulated CYP19A1 mRNA Levels and Aromatase Activity in ESCs
Real-time qPCR was used to quantify CYP19A1 mRNA levels in cultured ESCs and EMs from 3 types of samples. The CYP19A1 mRNA (CT > 40, n = 8) was not detectable in EMs from healthy women, while CYP19A1 mRNA was expressed in ESCs and EMs from patients with endometriosis. Level of CYP19A1 mRNA in ESCs was markedly higher (20 720.70-fold) than that in paired EMs (Figure 1A). As endometriosis is an estrogen-dependent disease, and PGE2 is known to stimulate the expression of CYP19A1 mRNA, 9 which is critical for the local production of estrogens driving the development of the disease, 7 we aimed to investigate the effect of metformin on PGE2-induced CYP19A1 mRNA expression in ESCs (Figure 1B). Results of the RT-PCR revealed a 4- to 5-fold increase in the CYP19A1 mRNA level following treatment with PGE2. Addition of metformin markedly reduced this induction by 30%. Furthermore, metformin could reduce basal CYP19A1 mRNA expression by 19%, but the difference was not statistically significant. As depicted in Figure 1C, similar findings were obtained with the aromatase activity assay. In addition, we selected EMs from women with endometriosis or without endometriosis as control group. After PGE2 and metformin treatment, CYP19A1 mRNA (CT > 40, n = 3) was not detectable in EMs from healthy women, while there was no effect on CYP19A1 mRNA in EMs from patients with endometriosis (Supplemental Figure 1).
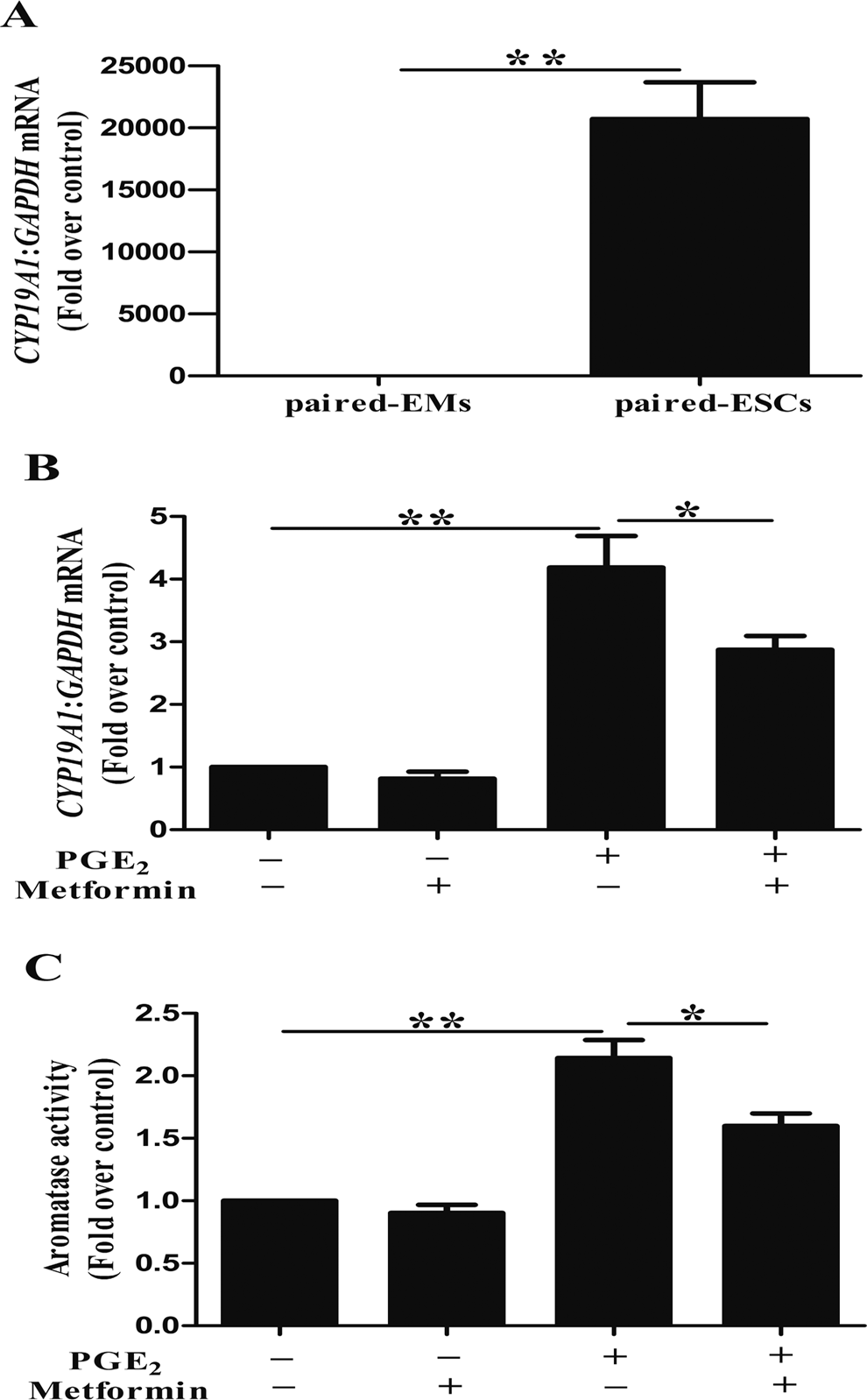
Metformin reduces prostaglandin E2 (PGE2)-stimulated CYP19A1 messenger (mRNA) levels and aromatase activity in human endometriotic stromal cells (ESCs). (A) CYP19A1 mRNA levels in primary ESCs were markedly higher (20 720.70-fold) than those in paired endometrial cells (EMs; n = 12; **, P < .01, t test). (B) ESCs were incubated with or without metformin (100 μmol/L) for 24 hours and then stimulated with or without PGE2 (1 μmol/L) for another 24 hours. Total RNA was extracted from untreated (control) or treated ESCs. The CYP19A1 mRNA levels were evaluated using real-time reverse transcription polymerase chain reaction (PCR; n = 3; *, P < .05, **, P < .01, analysis of variance [ANOVA]). (C) Aromatase activity was assayed. The basal aromatase activity in the control group was normalized to 1 (n = 3, *, P < .05, **, P < .01, ANOVA).
Inhibition of CYP19A1 mRNA Expression by Metformin is Independent of MEK/ERK
As treatment of ESCs with metformin induced MEK/ERK and AMPK phosphorylation, 25 we examined the effect of ERK phosphorylation on CYP19A1 mRNA expression. ESCs were preincubated with a MEK/ERK inhibitor PD for 1 hour and then cultured with metformin for 24 hours, after which they were treated with PGE2 for another 24 hours. Compared with PGE2 + metformin, treatment with PGE2 + metformin + PD stimulated a 1.72-fold increase in the CYP19A1 mRNA levels. Although PD reversed the metformin-induced decrease in CYP19A1 mRNA expression, the difference was not statistically significant (Figure 2A). In aromatase activity assay, compared with PGE2 + metformin, treatment with PGE2 + metformin + PD stimulated only a 1.31-fold increase and this difference did not achieve statistical significance (Figure 2B).
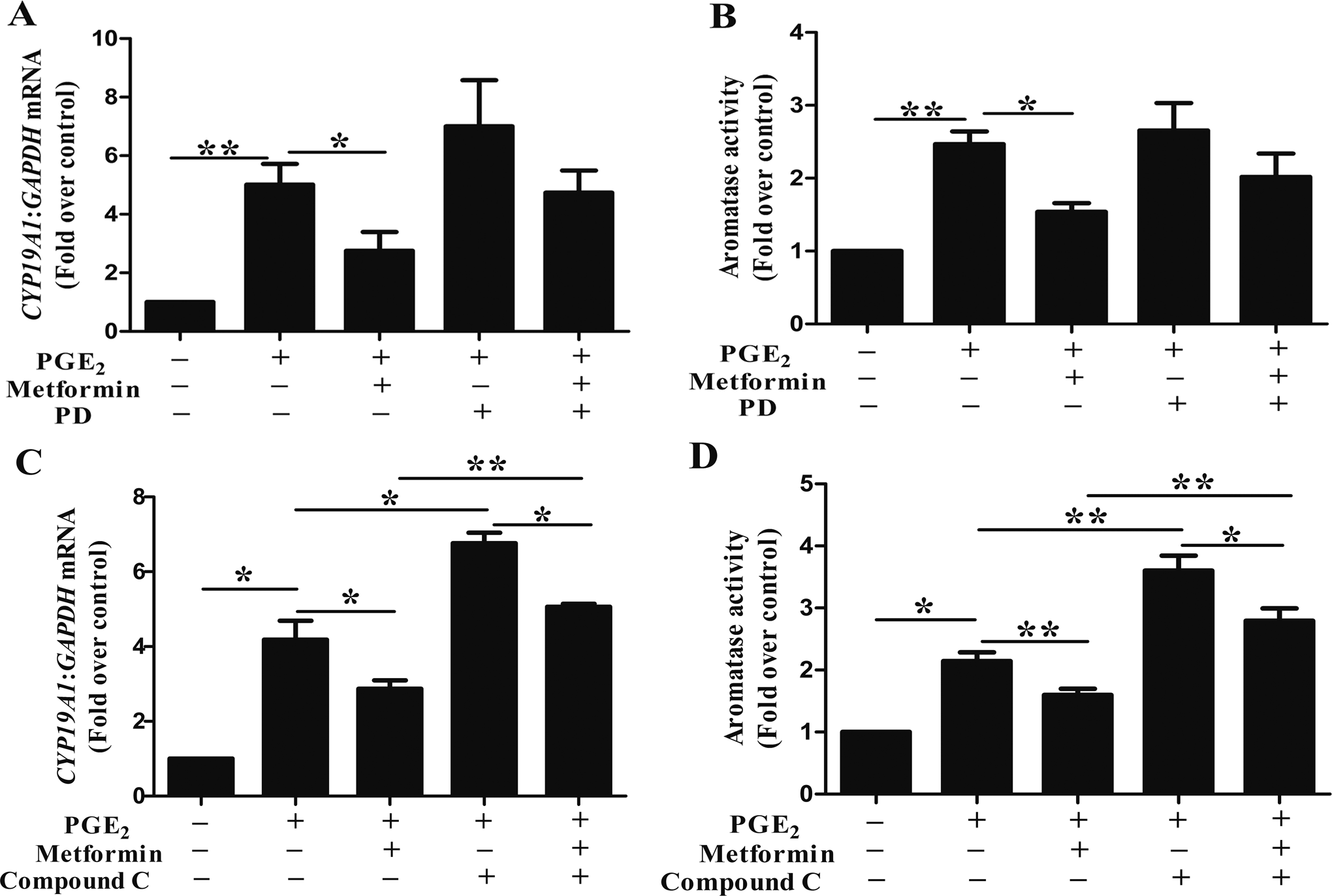
Effects of inhibitor of mitogen-activated protein kinase kinase (MEK)/ extracellular-signal-regulated kinases (ERK) or adenosine monophosphate-activated protein kinase (AMPK) on (metformin + prostaglandin E2 [PGE2])-induced CYP19A1 messenger RNA (mRNA) expression and aromatase activity in human endometriotic stromal cells (ESCs). (A and B) ESCs were preincubated with or without PD (25 μmol/L) for 1 hour, then incubated with metformin (100 μmol/L) for 24 hours, and then stimulated with or without PGE2 (1 μmol/L) for 24 hours. The ESCs with no treatment were used as control. Quantification of aromatase mRNA levels (A) and aromatase activity assays (B) was done (n = 6; *, P < .05, **, P < .01, analysis of variance [ANOVA]). (C and D) ESCs were preincubated with or without CC (5 μmol/L) for 1 hour, then incubated with metformin (100 μmol/L) for 24 hours, and then stimulated with or without PGE2 (1 μmol/L) for another 24 hours. The ESCs with no treatment were used as control. Quantification of CYP19A1 mRNA levels (C) and aromatase activity assays (D) was done (n = 6; *, P < .05, **, P < .01, ANOVA).
Effects of Inhibitor or siRNA-Mediated Knockdown of AMPK on CYP19A1 mRNA Expression and Aromatase Activity in ESCs
We measured the CYP19A1 mRNA levels and aromatase activity in the absence or presence of the AMPK inhibitor (CC) following metformin and PGE2 treatment for 48 hours. Compared to PGE2 + metformin, treatment with PGE2 + metformin + CC stimulated a nearly 2-fold increase in the CYP19A1 mRNA levels (Figure 2C). To provide additional evidence to illustrate the role of AMPK in the regulation of aromatase by metformin and PGE2, we used siAMPKα1 to knock down the endogenous expression of AMPKα in ESCs. There was a dose–response relationship between concentration of siRNA and functional AMPK knockdown with the maximum efficiency at 100 nmol/L (Figure 3A). The PGE2 stimulated a marked increase in the CYP19A1 mRNA levels in the control siRNA-transfected ESCs, while metformin treatment attenuated this effect. And more importantly, compared to PGE2 + metformin + siControl, the CYP19A1 mRNA level showed an almost 1.8-fold increase in the ESCs treated with metformin + PGE2 + siAMPKα1 (Figure 3B). Similar results were obtained in the aromatase activity assay (Figure 2D and 3C). Interestingly, we also found that CC or siAMPK could increase the PGE2-induced CYP19A1 mRNA expression level (Figure 2C and 3B) and aromatase activity (Figure 2D and 3C). But when compared to PGE2 + CC, treatment with PGE2 + metformin + CC reduced the CYP19A1 mRNA level by 25% (Figure 2C) and the aromatase activity by 22% (Figure 2D). Similarly, compared to PGE2 + siAMPKα1, treatment with PGE2 + metformin + siAMPKα1 reduced the aromatase activity by 30% but not CYP19A1 mRNA level (Figure 3C).
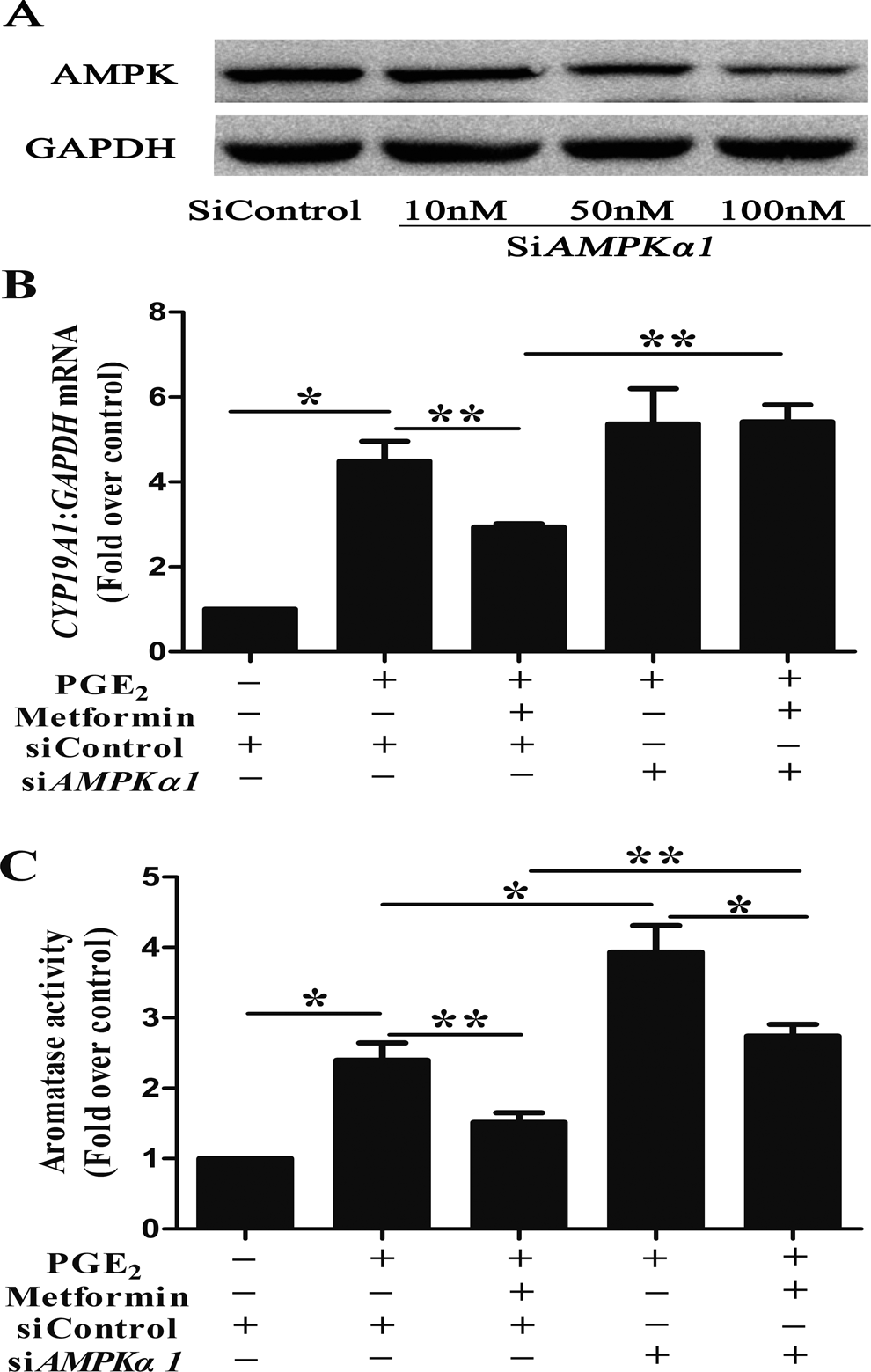
Effects of small-interfering RNA (siRNA)-mediated knockdown of adenosine monophosphate kinase (AMPK) on (metformin + prostaglandin E2 [PGE2])-induced messenger RNA (mRNA) expression and aromatase activity in human endometriotic stromal cells (ESCs). (A to C) ESCs transfected with the indicated siRNAs, serum starved, and treated with or without PGE2 and metformin. Cells were then harvested for Western blot analysis with anti-AMPKα, and anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibodies (A), CYP19A1 coding region-specific real-time PCR (B), and aromatase activity assays (C). Cells transfected with the siControl were incubated with no hormonal treatment as the control (n = 3; *, P < .05, **, P < .01, analysis of variance [ANOVA]).
Metformin Attenuates PGE2-Stimulated CREB Binding to CYP19A1 PII Region
As CREB has reported to be phosphorylated by metformin, 30 it is suggested that metformin might regulate expression of CYP19A1 gene through CREB by interacting with CYP19A1 PII. To determine whether CREB binds to CYP19A1 PII, we treated ESCs with either PGE2 for 1 hour or metformin + PGE2 for 12 hours and used ChIP with anti-CREB antibody followed by CYP19A1 PII-specific PCR (Figure 4A and B). There was a marked increase in CREB binding after PGE2 treatment, while treatment with metformin attenuated the above-mentioned stimulation by 67%.
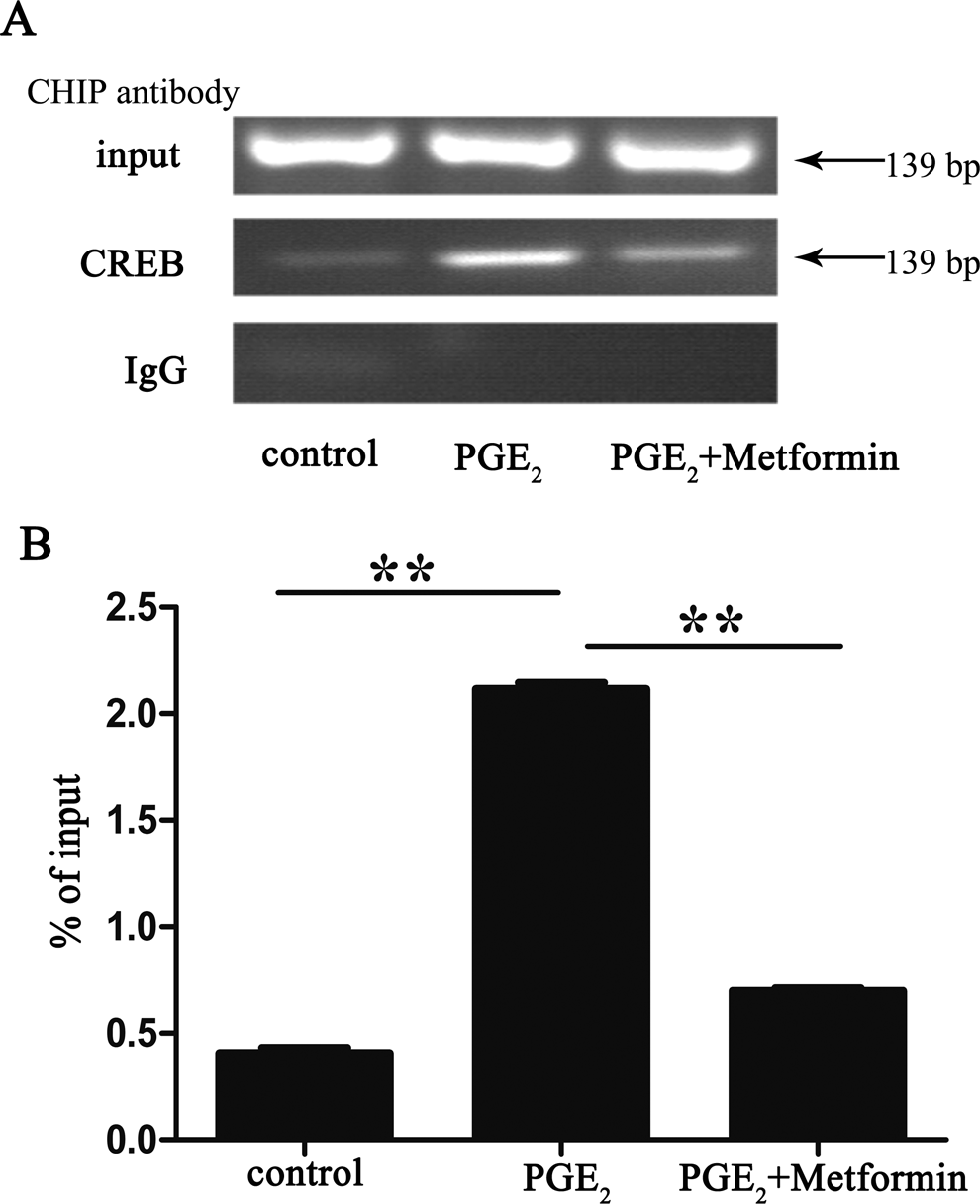
Metformin attenuates the prostaglandin E2 (PGE2)-stimulated binding of cyclic adenosine monophosphate response element-binding (CREB) protein to the CYP19A1 promoter II region. After a 16-hour serum starvation, human endometriotic stromal cells (ESCs) were incubated in the presence or absence of 100 μmol/L metformin for 12 hours and then stimulated with 1 μmol/L PGE2 for another 1 hour. The treated and untreated cells were harvested and subjected to chromatin immunoprecipitation (ChIP) with anti-CREB antibody or control immunoglobulin G (IgG) for the control. Semiquantitative polymerase chain reaction (PCR; A) and real-time quantitative PCR (B; n = 3; **, P < .01, analysis of variance [ANOVA]).
Discussion
Numerous reports have shown abundant CYP19A1 mRNA expression and elevated local estrogen production in endometriotic tissues, 7 suggesting that P450arom is responsible for the local production of estrogen. A review about the use of aromatase inhibitors (AIs) for the treatment of endometriosis have suggested that in reproductive-aged women, the combination of an AI with conventional therapy does alleviate endometriosis-related pain. But it also points that more studies need to be done examining pregnancy rates and outcomes following AI treatment for endometriosis and larger multicenter randomized clinical trials using AI for the treatment of endometriosis-related chronic pelvic pain need to be done. 31 In addition, the equivalency of metformin and letrozole is shown in a rat model but not a clinical trial. 21 In this study, we demonstrated that metformin inhibits PGE2-induced CYP19A1 mRNA expression and aromatase activity in human ESCs by inhibiting the binding of CREB to the PII promoter. This involved the stimulation of AMPK. This is especially relevant for our understanding of the mechanism of action of metformin in women with endometriosis.
Most studies using PCR and immunohistochemistry show that the aromatase is expressed in both eutopic and ectopic endometrium of patients with endometriosis, while this enzyme is not detectable in eutopic endometrium obtained from healthy women. 5,9,10,32 –36 Although a few of studies failed to detect aromatase immunoreactivity in ectopic lesions or eutopic endometrium from patients with endometriosis, 37,38 the results obtained in our study were similar to the conclusion of most of the previous studies. These findings suggest that aromatase still plays an important role in local estrogen synthesis in endometriosis. 39
It is well established that PGE2 enhances CYP19A1 mRNA expression in human ESCs. 4,9,33,40 We have shown that addition of metformin to the PGE2-treated ESCs notably attenuated the PGE2-stimulated upregulation of CYP19A1 mRNA expression, as expected from previous observations on the metformin-induced suppression of cAMP or PGE2-induced CYP19A1 mRNA expression. 25,41 More importantly, the changes in CYP19A1 mRNA expression could translate into significant reduction in the aromatase activity. In addition, metformin has been shown to inhibit the mRNA expression of follicle-stimulating hormone (FSH) and insulin-stimulated CYP19A1 in granulosa cells 20,26,30 as well as significantly reduce the FSK/PMA-dependent upregulation of CYP19A1 mRNA expression in primary human breast adipose stromal cells. 42
What is the intracellular mechanism underlying these diverse effects of metformin in ESCs? The AMPK is a known target of metformin action in various cells. 22,42 –47 When comparing PGE2 + metformin with PGE2 + metformin + CC/siAMPKα1, addition of an AMPK inhibitor (CC) or siAMPKα1 could reverse the reduction by metformin. That is to say, the AMPK signaling pathway plays a role in regulation of aromatase expression by metformin. But when compared to PGE2 + CC/siAMPKα1, treatment with PGE2 + metformin + CC/siAMPKα1 still reduced the CYP19A1 mRNA level or the aromatase activity. The interpretation of this result is that a component of the reduction in CYP19A1 mRNA expression and aromatase activity by metformin is independent of AMPK. The possible reason of the discrepancy between the above-mentioned results is that metformin may also regulate aromatase expression through other signaling pathways and other mechanisms that are still unknown. In addition, we have proven that the AMPK pathway was phosphorylated gradually after metformin stimulation in ESCs. 25 In summary, inhibition of CYP19A1 mRNA expression and aromatase activity by metformin is partly AMPK dependent. Other mechanisms are for further study. Furthermore, the possible reason of the discrepancy between the mRNA expression results and the aromatase activity results in Figure 3 is that siAMPK may be involved in the complex regulation of translation and posttranslation modification. Interestingly, in the absence of metformin, when we used the AMPK inhibitor CC or siAMPKα1 to block AMPK phosphorylation, we found that PGE2-induced CYP19A1 mRNA levels were increased. In other words, the AMPK inhibitor and PGE2 might have a synergistic effect on the gene expression of CYP19A1 in human ESCs. Moreover, adiponectin, whose levels in the serum and peritoneal fluid in women with endometriosis were decreased, stimulated AMPK and inhibited inflammatory cytokine production in EMs. 48 –50 In addition, AMPK might be involved in the anti-inflammatory effects of metformin demonstrated in ESCs. 26 These findings indicated that AMPK activation in endometriosis was likely to be a protective factor. The MEK/ERK signaling cascade has been demonstrated to regulate CYP19A1 gene expression and steroidogenesis; however, conflicting findings have been reported with respect to the mechanism in different steroidogenic cells. For example, inhibition of MEK activity with PD or U0126 (U0) has been reported to be associated with stimulation, 26,51 inhibition, 52,53 or no effect 25,54 on the steroidogenic response. Thus, our data demonstrated that inhibition of CYP19A1 mRNA expression by metformin was independent of the MEK/ERK pathway. Activation of the MEK/ERK pathway may play a role in other aspects of metformin’s function. Together, metformin inhibits CYP19A1 mRNA expression and aromatase activity in human ESCs partly via the stimulation of AMPK.
Our findings indicated that metformin could reduce the PGE2-stimulated binding of CREB to CYP19A1 PII in human ESCs. It is noteworthy that metformin reduced the expression of CYP19A1 at the transcriptional level. In addition, metformin interacted with the FSH-stimulated cAMP/PKA/CREB pathway, which is the primary signaling pathway for the regulation of CYP19A1 gene expression in the ovary. 55 These findings suggest that therapeutic effects of metformin might also be mediated by the suppression of ovarian estrogen production through the same mechanisms as shown in the ESCs. In 2003, a new family of CREB coactivators known as CREB-regulated transcription coactivators (CRTCs) or transducers of regulated CREB activity was identified. 56,57 In human breast adipose stromal cells, metformin inhibited the nuclear translocation of CRTC2, which is known to increase aromatase expression by binding to CYP19A1 PII, which is also a direct downstream target of AMPK. 42,47 In human granulosa cells, metformin reduced the FSH-induced phosphorylation of CREB thereby reducing the CRE activity, which could result in disruption of the CREB-CBP-CRTC2 coactivator complex that binds to CRE in PII of the CYP19A1 gene. 30 In our previous article, we have showed metformin’s ability to disrupt CREB–CRTC2 complex in human ESCs. 25 Together, these findings suggest that metformin attenuates the PGE2-stimulated binding of CREB to CYP19A1 PII probably via disrupting the CREB–CRTC2 complex.
Conclusion
In summary, metformin successfully reduced the PGE2-stimulated CYP19A1 mRNA expression and aromatase activity partly by stimulating AMPK and decreased the binding of CREB to CYP19A1 PII. These findings suggest the unique therapeutic potential of metformin as an antiendometriotic drug.
Footnotes
Acknowledgement
We appreciate Prof Ding-Fang Bu for his generous advice for the study.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by grants from the National Natural Science Foundation of China (Grant No. 81270674) and the Natural Science Foundation of Beijing, China (Grant No.7132204).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.