
Abstract
Mammalian embryonic development is an intricate succession of physiological and morphological events. Many studies have focused on optimizing in vitro culture systems for improvement in embryonic development. In this study, we established a novel coculture method with outgrowth embryos and investigated how this coculture system improves the preimplantation and peri-implantation embryonic development both in vitro and in utero. We found that outgrowth embryos secrete vesicles, as observed by time-lapse monitoring and scanning electron microscopy. Coculture with outgrowth embryos also significantly increased the percentages of morula, blastocyst, hatching, and outgrowth (P < .01). The total number of cells and inner cell mass were increased, and apoptotic index was decreased (P < .05) by upregulating Survivin and Lif messenger RNA expression levels (P < .05) in the coculture compared to the control group. Furthermore, implantation rates in utero after embryo transfer were significantly higher for cocultured embryos than those for the control group (P < .05). We conclude that embryotrophic factors secreted from outgrowth embryos could improve the developmental competence of in vitro cultured mouse preimplantation embryos. Findings of specific embryotrophic factors from outgrowth embryos might be valuable for advancing reproductive technologies in the future.
Introduction
The early stage of mammalian embryonic development, from fertilization to implantation, is a series of dynamic events including both physiological and morphological changes driven by a complex succession of genome activation. 1 –5 Successful development occurs under highly regulated physiological conditions synchronized by the reproductive organs. 6 –8 Especially, trophoblast invasion by peri-implantation embryos in utero is a critical event required for the completion of implantation and pregnancy and is related to cyclic changes in hormones such as estrogen and progesterone. 9 –12 Optimizing the culture system that enhances survival and development in vitro is desirable to improve human in vitro fertilization–embryo transfer (IVF-ET) program; early studies have compared various culture systems and trophic factors, including embryo culture media, gas phase, optimizing endometrial receptivity, and several growth factors. 13 –18 However, there is still considerable interest in improving current human IVF-ET procedures by determining the optimal embryonic culture conditions. Live birth rate per transfer from assisted reproductive technologies remains as low as 30%. 19
Fertilized zygotes are bathed in maternal tissue-derived secretions from the time they first enter the reproductive tract and throughout the peri-implantation period. 20 –24 To mimic this environmental factor in vitro, many groups have attempted to develop coculture systems or use conditioned media to include various embryotrophic factors derived from reproductive cells, including cumulus, oviductal, and endometrial cells. 25 –27 Additionally, coculture of embryos appears to enhance in vitro growth independently of reproductive tissues; preimplantation embryos cultured in groups develop better than those in individual culture. 28 –30 Recently, immunoregulatory and physiological advantages of both coculture and conditioned media methods were shown using trophoblast cell lines or embryonic stem cells. 31 –33 Thus, coculture systems allow extended in vitro culture of embryos to the blastocyst stage, suggesting that postimplantation embryonic development would be enhanced. 34,35
Outgrowth of blastocysts is an in vitro model for implantation of developing embryos in utero that enables the study of implantation mechanisms, which are difficult to decipher in vivo. 36,37 Aspects of trophoblastic outgrowth, such as activation and spreading, have been correlated with in vivo implantation processes, such as proliferation and invasion, supporting this model for determining optimal conditions for embryonic culture. 11,38 –40 Outgrowth assays have also shown that the morphokinetics and metabolism of embryos are related to viability and implantation potential. 19 Additionally, differential secretion profiles have been described between implanted and nonimplanted embryos at the outgrowth stage in vitro. 41 Overall, outgrowth assays are an important tool for understanding the factors that influence embryonic viability and implantation.
In this study, we established a novel coculture system with outgrowth embryos and investigated whether the coculture with outgrowth embryos improves the preimplantation embryonic development in vitro and implantation in utero.
Materials and Methods
The overall scheme is presented in Figure 1.
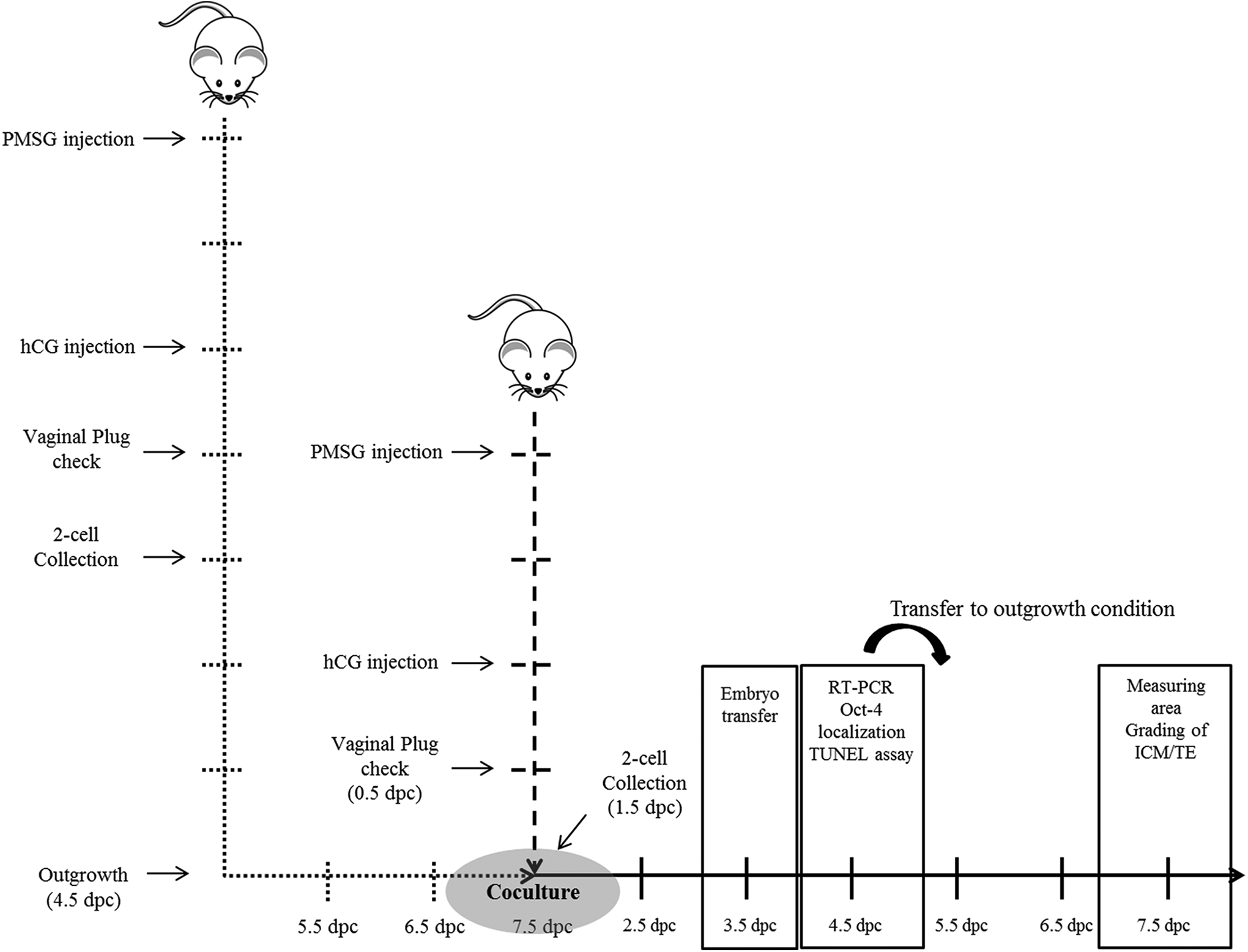
Schematic representation of the coculture method and timing of assays to investigate the effects of coculture system with outgrowth embryos on the developmental competence. Mouse 2-cell embryos were cultured to blastocyst and outgrowth stage in the control and coculture groups, respectively. The developmental competence of preimplantation embryo was assessed in terms of developmental rates, cell numbers, apoptotic cells, and messenger RNA (mRNA) expression. The developmental competence of peri-implantation embryonic development was assessed by outgrowth rate, trophoblastic spreading area, grading score, and implantation rate in utero by embryo transfer.
Animals and Hormonal Stimulation
This study was approved by the Eulji University Institutional Animal Care and Use Committee (EUIACUC 12-19). Six- to eight-week old ICR mice were superovulated with intraperitoneal injections of 5 IU pregnant mare’s serum gonadotropin (Sigma, USA), and 48 hours later, the mice were injected with 5 IU human chorionic gonadotropin (hCG; Sigma). Superovulated mice were then individually mated with a fertile ICR male. 42 The day of vaginal plugging was designated as 0.5 days postcoitum (dpc).
Embryo Collection and Culture to Outgrowth Stage
Mouse 2-cell embryos were collected at 46 hours post-hCG injection (1.5 dpc), and they were cultured together in 10 μL drops of Quinn’s Advantage Blastocyst Media (QABM; SAGE, USA) with serum protein substitute (SPS; SAGE) in 5% CO2 at 37°C. Embryos were cultured for 3 days without changing the medium, and the blastocysts were randomly selected at 120 hours post-hCG injection (4.5 dpc) and assigned to each assay. Each blastocyst was individually transferred to a fibronectin (FN)-coated dish (Sigma) and cultured to generate outgrowth embryos in Dulbecco modified Eagle media (DMEM; Gibco, USA) for 3 days (7.5 dpc). 19,20,38,43 Outgrowth embryos were washed with Dulbecco phosphate-buffered saline (Welgene, Korea) and prepared for coculture in serum-free QABM. The embryo collection was repeated more than 5 times.
Observation of Outgrowth Embryo by Time-Lapse Monitoring System and Scanning Electron Microscopy
Mouse blastocysts were transferred to a well-of-well embryo culture dish of a Primo time-lapse system (Vitrolife, Sweden) and cultured to outgrowth for 3 days. Images of embryos were captured every 30 minutes, and morphological development of outgrowth embryos was analyzed by the recorded images. For detailed observation of outgrowth embryos, blastocysts were cultured on a FN-coated 10-mm rounded coverslip in a dish and prefixed in 2.5% glutaraldehyde, followed by postfixation with 1% osmium tetroxide for 1 hour. The fixed samples were transferred from ethanol to amyl acetate and critical point dried with liquid CO2. Dried samples were sputter coated with platinum–palladium (HITACHI, E1030, Japan) and observed with a HITACHI S4700 scanning electron microscope.
Coculture of Preimplantation Embryos With Outgrowth Embryos
Each 2-cell embryo on 1.5 dpc was cocultured with a prepared outgrowth embryo for 3 days in a 5 μL drop of QABM under mineral oil (SAGE) in 5% CO2 at 37°C for 3 days. Control embryos were cultured alone. The percentages of morula on 3.5 dpc, blastocysts on 4.5 dpc, and hatching blastocysts on 4.5 dpc were determined based on morphological criteria.
Immunofluorescence and Terminal Deoxynucleotidyl Transferase-Mediated dUTP Nick End Labeling Assay
The percentage of blastocysts on 4.5 dpc was determined following final morphological assessment, and the blastocysts were assigned to immunocytochemistry and terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) assay. Localization of octamer-binding transcription factor 4 (Oct-4), which is the essential transcription factor for the formation of inner cell mass (ICM), 33,44 was determined by immunocytochemistry. Apoptotic cells were detected by fluorescein isothiocyanate-conjugated in situ cell death detection kit (TUNEL; Promega, USA). Blastocysts from the control and coculture groups were fixed using 4% paraformaldehyde (Sigma) and permeabilized with 0.5% Triton X-100 solution for 10 minutes. The blastocysts were incubated in blocking solution of 3% bovine serum albumin (Sigma), reacted with 1:200 diluted anti–Oct-4 antibody (Abcam, United Kingdom), and then stained with a secondary Alexa 594 antibody (Life technology, Norway). Apoptotic cells were stained by TUNEL kit according to the manufacturer’s instructions. Nuclei were stained in a solution of 10 µg/mL bisbenzimide (Hoechst 33342; Sigma) for 10 minutes before observation with a fluorescence microscope (AX-70; Olympus, Japan). Additionally, the number of cells with Oct-4 and TUNEL-positive nuclei was determined using an image capture system (IMT i-Solution, British Columbia, Canada). The apoptotic index was calculated as the percentage of TUNEL-positive nuclei divided by the total number of nuclei in a single blastocyst.
Outgrowth Assay
Blastocysts from the control and coculture groups were transferred to a FN-coated dish and outgrown to examine the effects of coculture on peri-implantation embryonic development. Morphologies during peri-implantation development were observed every 24 hours, and captured images were analyzed. The area of trophoblastic outgrowth was measured on 7.5 dpc using Image J software (National Institutes of Health, USA). Inner cell mass and trophectoderm (TE) of outgrowth embryos were graded and scored in accordance with morphological criteria as described previously. 45 Grading score of ICM (from 0 to 3) was as follows: (1) for grade score 0, no ICM development, (2) for grade score 1, a small area of ICM development, (3) for grade score 2, a larger ICM without development of depth, and (4) for grade score 3, a group of ICM completely developed with depth. Grading score of TE (from 0 to 3) was as follows: (1) for grade score 0, no trophoblast cells spread out, (2) for grade score 1, a small area of spreading trophoblast cells, (3) for grade score 2, a larger area with small amount of trophoblast cells, and (4) for grade score 3, completely spread out with a large amount of trophoblast cells.
Embryo Transfer
Implantation potential of embryos from the control and coculture groups was examined via embryo transfer as described previously. 46,47 Embryos were transferred on 3.5 dpc to pseudopregnant recipients provided by mating with vasectomized male. Two days after transfer (5.5 dpc), implantation sites in uterine horns of pregnancy were detected by intravenous injection of Chicago blue dye. Clear blue bands in utero were considered implantation sites.
Quantitative Reverse Transcription-Polymerase Chain Reaction (qRT-PCR)
Total messenger RNA (mRNA; 10 μL) from 5 blastocysts was isolated using mRNA Direct kit (Dynal, Oslo, Norway), and 80 μL of complementary DNAs (cDNAs) was synthesized using 2 μL of total mRNA (10 μL) with PrimeScript first strand cDNA synthesis kit (Takara Bio, Japan). A portion of the reverse transcription product (2 μL) was used directly for real-time quantitative polymerase chain reaction (qPCR). The qPCR was performed in a final reaction volume of 20 μL with SYBR Green using primer sets for apoptosis and survival-related genes (Table 1), according to the manufacturer’s instructions (Applied Biosystems, United Kingdom). The internal standard for normalization and comparison was 16 S ribosomal RNA. The quantitative analysis was presented using a modification of the 2−ΔΔct method. 48,49 This experiment was performed 3 times.
Primer Sequences and Product Sizes Used for qRT-PCR.

Abbreviation: qRT-PCR, quantitative reverse transcription-polymerase chain reaction.
Statistical Analysis
All experiments were performed at least in triplicates. Statistical significance of multiple comparisons was determined by 1-way analysis of variance or χ2 test. Significances between 2 experimental groups were analyzed by Student t test. A P value of less than .05 was considered statistically significant.
Results
Observation of Outgrowth Embryos by Time-Lapse Monitoring System and Scanning Electron Microscopy
To characterize outgrowth embryos, mouse embryonic development (from 4.5 dpc of blastocyst stage) was observed every 30 minutes for 3 days with time-lapse monitoring system during trophoblastic outgrowth in vitro. Transferred blastocysts began to hatch out from the zona pellucida and attached to the dish on 5.5 dpc. The trophoblast of the attached blastocysts started to spread out on 5.5 dpc and continued to spread during culture. Motility was observed in blastocysts, and some particles and vesicles were shed and secreted from them during spreading (Figure 2A). To provide a more detailed observation, on 7.5 dpc, morphologies of outgrowth embryos and secreted vesicles were observed by scanning electron microscopy (SEM) in Figure 2B. The observed size of secreted vesicles was 50 to 200 nm.

Morphological changes in a hatched blastocyst to outgrowth stage for 3 days. Serial images were captured by time-lapse monitoring system for 3 days. Scale bar, 50 μm. B1 to 4, An outgrowth embryo and membrane budding microvesicles from an outgrowth embryo on 7.5 days postcoitum (dpc) by scanning electron microscopy. B1, A whole outgrowth embryo. B2 and B3, A part of inner cell mass and trophectoderm in outgrowth embryos, respectively. B4, An enlarged feature of B3.
Establishment of a Novel Coculture System With Outgrowth Embryos
To improve embryonic viability and implantation, we developed a novel coculture system using outgrowth embryos to support the development of the preimplantation embryos. Mouse 2-cell embryos were cocultured with outgrowth embryos (described previously) to blastocyst stage for 3 days, and the developed blastocysts were transferred to FN-coated dish in DMEM for 3 days to model implantation in vitro via the embryo outgrowth assay (Figure 1). Transferred blastocysts began to hatch out from zone pellucida and attached to the dish on 5.5 dpc. Attached blastocysts were spread out on 7.5 dpc, mimicking the process of embryonic invasion into maternal tissue during implantation (Figure 3).
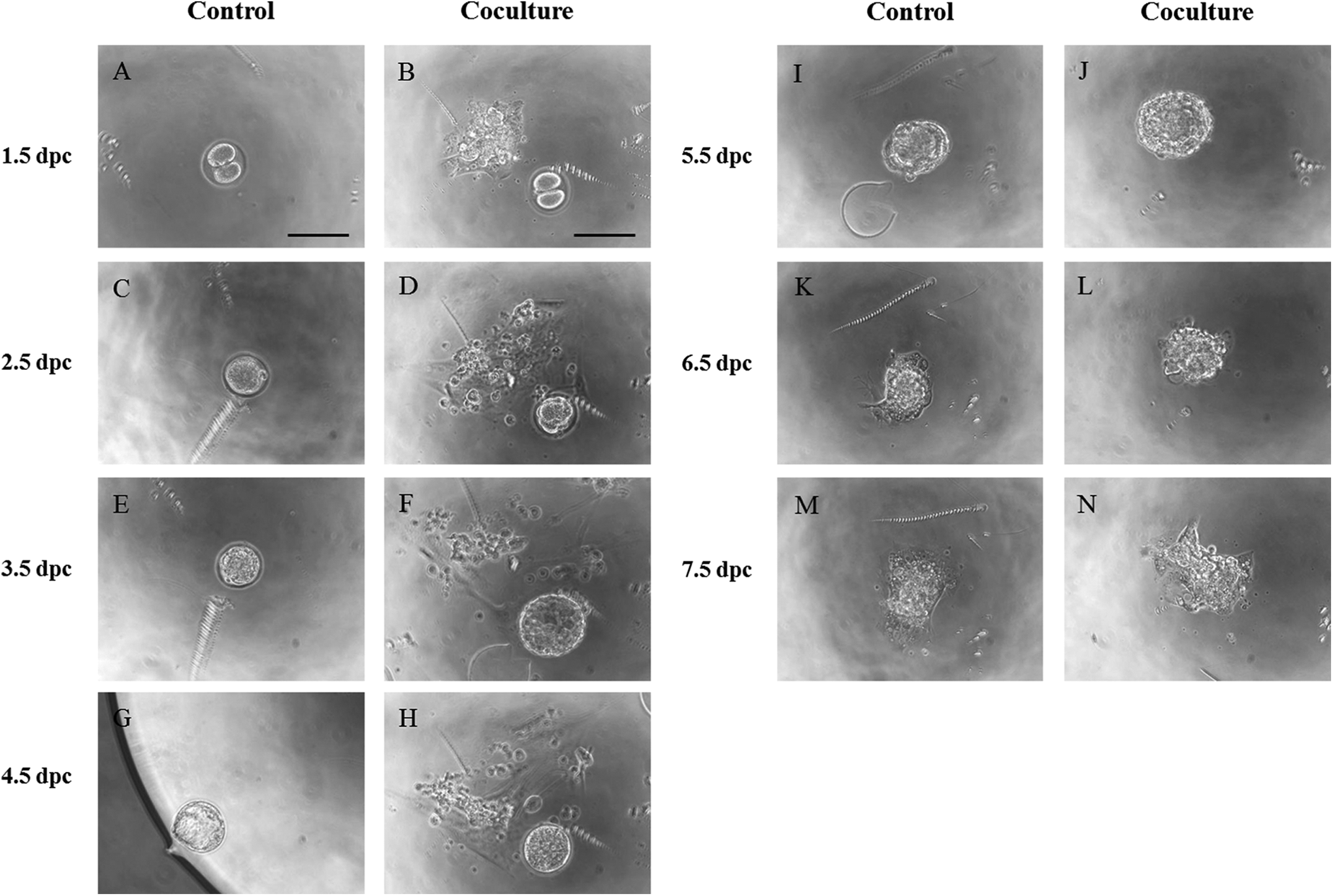
Preimplantation embryos were cultured from 1.5 days postcoitum (dpc) to 4.5 dpc without and with outgrowth embryos as the control and coculture groups, respectively. Outgrowth embryos were prepared for coculture by transfer of 2-cell embryos in fibronectin-coated dish on 1.5 dpc. For evaluation of peri-implantation developmental competence, cultured blastocysts (4.5 dpc) of the 2 groups were transferred to another dish and cultured to outgrowth up to 7.5 dpc. A, C, E, G, I, K, and M, Cultured embryo in the control group. B, D, F, H, J, L, and N, Cultured embryo in the coculture group. Scale bars = 100 μm.
Development of Preimplantation Embryos in Coculture System
The effects of coculture with outgrowth embryos on preimplantation embryonic development were evaluated by the percentages of morula, blastocyst, and hatching, respectively. The morulation, blastulation, and hatching rates were significantly improved in the coculture group (Figure 4; P < .05). In terms of cell proliferation, mean number of total cells and ICM were significantly higher in the coculture conditions than those in the control group (Table 2; P < .05). Correspondingly, the cellular apoptosis was lower in the coculture than in the control group (Table 2; P < .01).
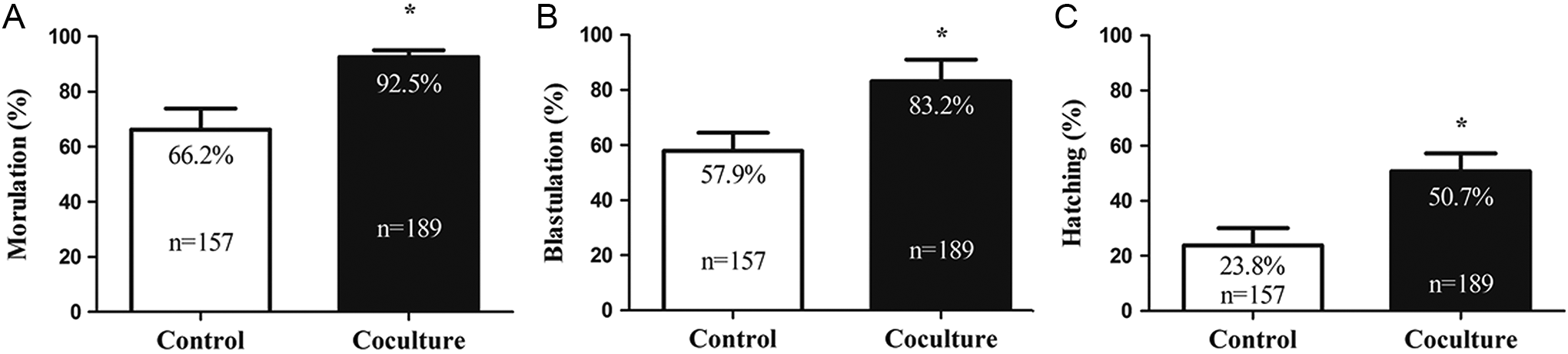
Development rates of preimplantation embryos in the 2 groups. Rates of morulation on 3.5 days postcoitum (dpc; A), blastulation on 4.5 dpc (B), and hatching on 4.5 dpc (C) are compared between the control and coculture groups, respectively. *P < .05, significant difference from the control group.
Effects of Coculture With Outgrowth Embryos on Developmental Competence of Preimplantation Embryos.a,b
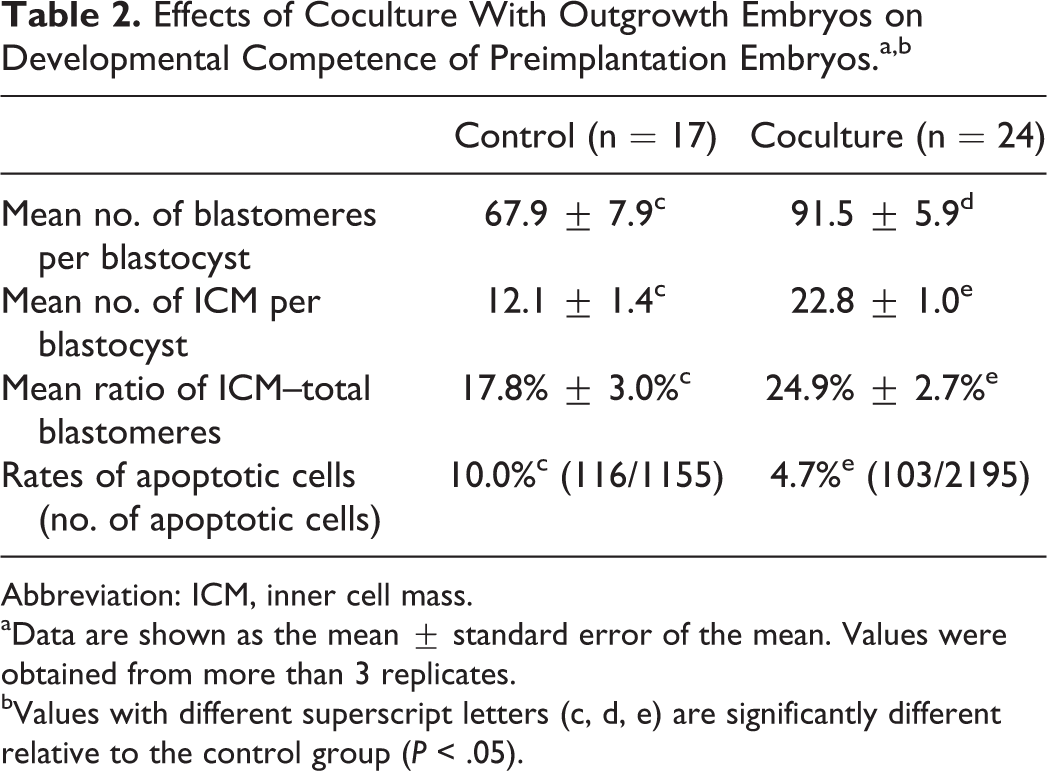
Abbreviation: ICM, inner cell mass.
aData are shown as the mean ± standard error of the mean. Values were obtained from more than 3 replicates.
bValues with different superscript letters (c, d, e) are significantly different relative to the control group (P < .05).
Development of Peri-Implantation Embryos From Coculture System
In addition to growth and viability, the ability of cocultured embryos to implant was assessed in vitro. Mouse 2-cell embryos were cocultured with outgrowth embryos for 3 days, and cultured blastocysts were cultured in FN-coated dishes until 7.5 dpc to evaluate implantation potential in vitro such as outgrowth rate, measurement of trophoblastic spreading area, and morphological scoring of ICM and TE by outgrowth assay (Figure 5). On 7.5 dpc, the outgrowth rate in the coculture group was significantly higher than that in the control group (Table 3; P < .01). However, there was no significant difference in trophoblastic spreading area and morphological grading of ICM and TE (Table 3) between the coculture and control groups.
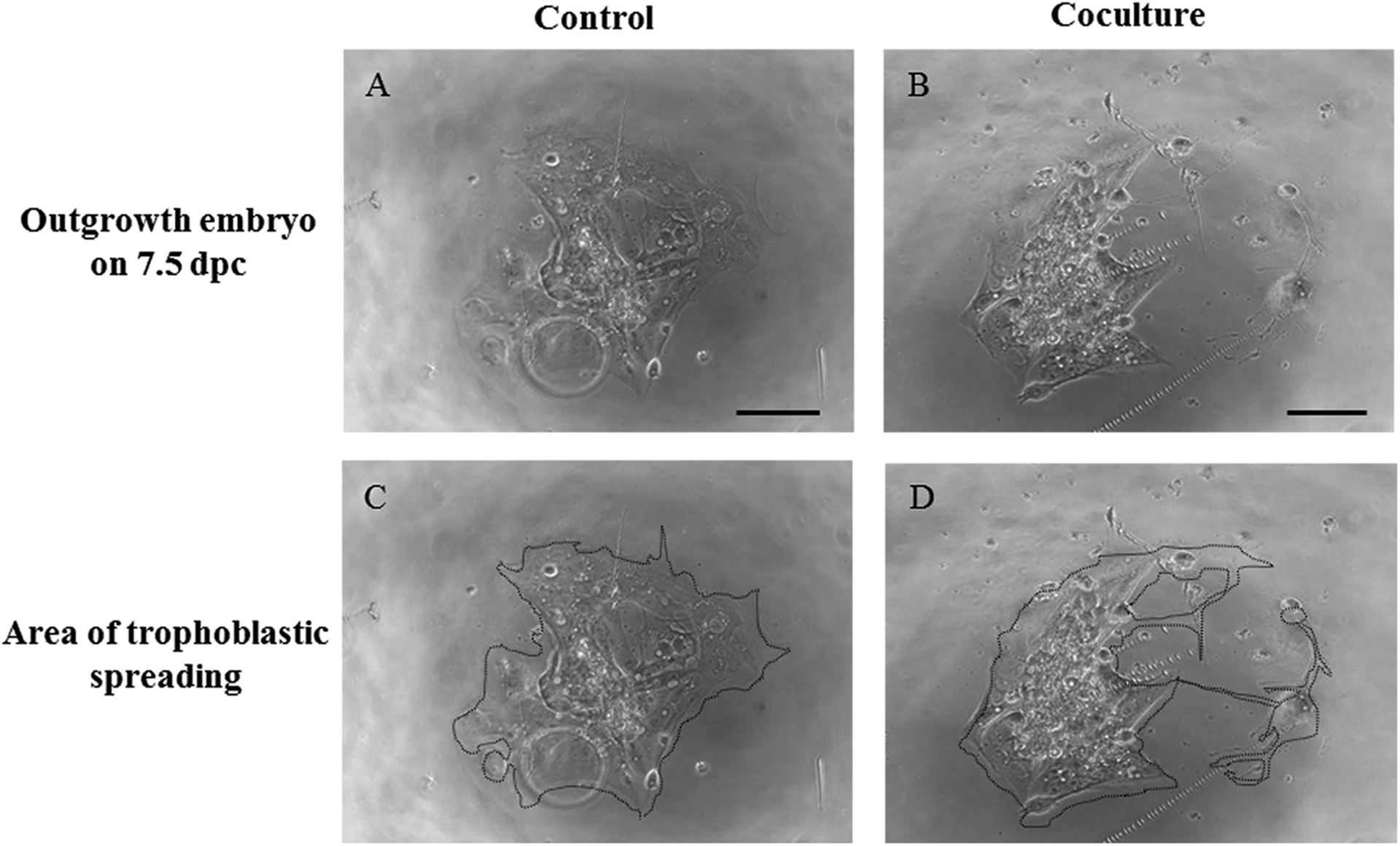
Assessment of outgrowth embryos in the control and coculture groups. A and B, Morphologies of outgrowth embryos. C and D, Area of trophoblastic spreading. Scale bars = 100 μm.
Effects of Coculture With Outgrowth Embryos on Development of Peri-Implantation Embryos.a,b
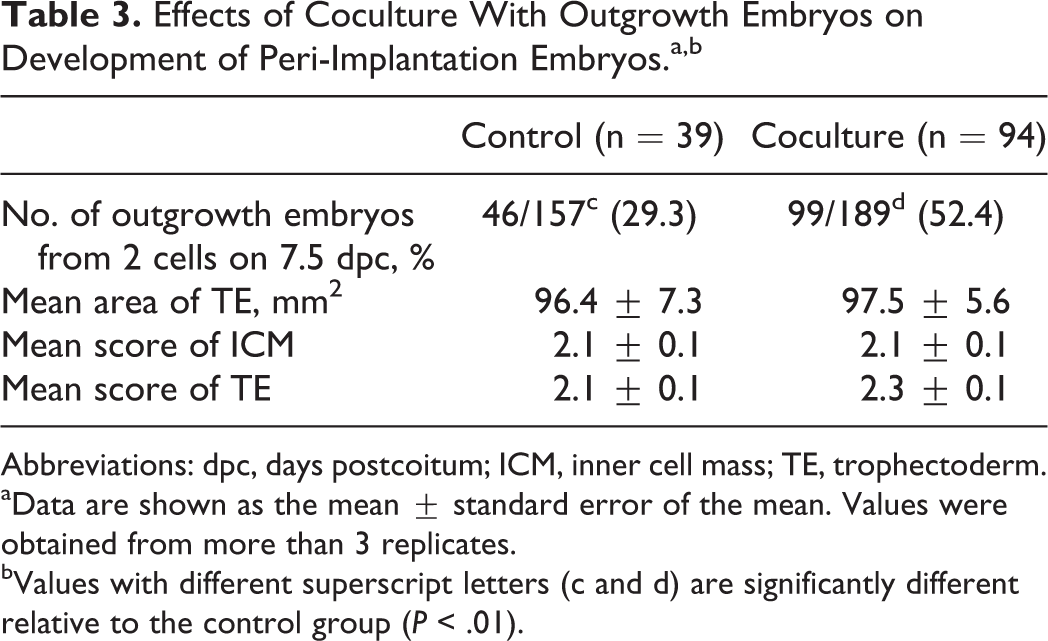
Abbreviations: dpc, days postcoitum; ICM, inner cell mass; TE, trophectoderm.
aData are shown as the mean ± standard error of the mean. Values were obtained from more than 3 replicates.
bValues with different superscript letters (c and d) are significantly different relative to the control group (P < .01).
Alteration of mRNA Expression in Cocultured Embryos
To further assess the viability of embryos that had been cocultured, RT-qPCR was performed to examine the alteration of gene expression in the coculture with outgrowth embryos. Expression levels of antiapoptotic genes such as Survivin, Bcl-2, and Mcl-1 mRNA increased in blastocysts of the coculture group compared to those in the control group (Figure 6), supporting the decrease in apoptotic index observed in Table 2. In contrast, Bax expression, which regulates proapoptotic signals, was similar between the 2 groups.
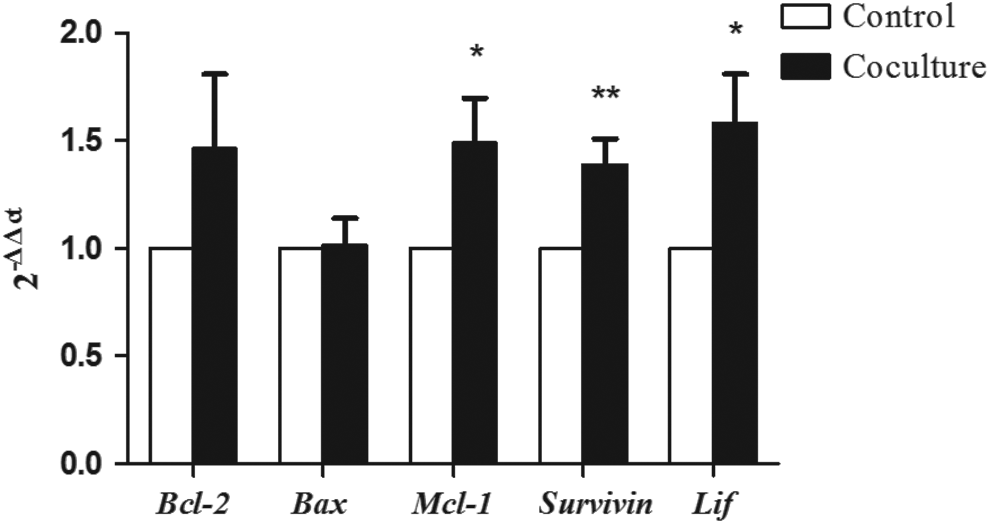
Expression levels of genes associated with apoptosis pathway (Bcl-2, Bax, Mcl-1, Survivin, and LIF) in the 2 groups. Expression was compared between the control and those cocultured with outgrowth embryos. Data are shown as mean ± standard error of the mean. *P < .05 and **P < .01.
Implantation Rate of Embryos From Coculture System
In order to assess whether the positive effects of coculture observed in vitro improve implantation, cocultured embryos were transferred to pseudopregnant females. Implantation rate in utero was significantly higher in the coculture group compared to the control group (Table 4; P < .05), confirming that the increased implantation competence observed in the coculture embryos is relevant in vivo and supporting the use of our coculture system.
Implantation Rates After Embryo Transfer.a,b

aData are shown as the mean ± standard error of the mean. Values were obtained from 3 replicates.
bValues with different superscript letters (c and d) are significantly different relative to the control group (P < .05).
Discussion
Coculture is an alternative method to improve embryo development during in vitro culture by providing a microenvironment that mimics physiological conditions. 27 Here, we describe a novel coculture system using outgrowth embryos and its effects on preimplantation and peri-implantation embryonic development. We have found that coculture with outgrowth embryos significantly increases the development and implantation competence of embryos and increases their rate of implantation in utero. Together, this suggests that outgrowth embryos provide a microenvironment that favors the development and implantation of embryos for transfer.
Bongso and Fong suggested that in vitro culture of embryos may be improved by coculture due to both positive and negative conditioning provided by the cocultured helper cells. 50 The positive conditioning is related to the putative embryotrophic factors such as antioxidants, glycoproteins, and growth factors, and the negative conditioning is related to the removal of hypoxanthine and reactive oxygen species reduction. 26,51 Our results support a role for cocultured cells in enhancing the development of cultured embryos, as we were able to grow 2-cell embryos with increased viability, which implanted in vitro and in vivo at greater rates than embryos cultured alone. Our results showed that the control group had a low blastulation rate, which was normally more than 80% of the blastulation rate achieved using commercial medium with serum components such as SPS. In this study, we cultured embryos of the control and coculture groups in medium without any protein supplementation to determine the exact coculture effect, which could be masked by serum components. Other groups have attempted to improve embryonic development in vitro using coculture systems with reproductive cells such as cumulus cells, oviductal, and uterine cells. 25,27,52,53 Our study has introduced outgrown embryos as a novel source of cells for coculture. Outgrowth of embryos is usually used as an in vitro model for the study of the peri-implantation embryonic development and the process of trophoblastic invasion during implantation. But, we have shown that these cells can support the in vitro growth of the preimplantation embryos during coculture to increase the likelihood of implantation. 33,54,55
Interestingly, we observed that outgrowth embryos secreted some vesicles as observed by time-lapse monitoring system and SEM. Membrane-derived vesicles and their contents may be contributing to the beneficial effects on preimplantation embryonic development. Because coculture with outgrowth embryos improved both development and proliferation of embryos, these vesicles could contain positive conditioning factors. Indeed, O’Neill and coauthors reported that embryos have abilities to secrete various factors such as cytokines, growth factors, and membrane vesicles. 56 Among these, membrane vesicles, also called extracellular vesicles or exosomes, have been investigated recently and found to have important roles in immune modulation. 57 Recent studies also reported that exosomes include proteins, DNAs, RNAs, and microRNAs to regulate gene expression and are implicated in endometrial receptivity, embryo development, and implantation failure. 58 –62 Thus, our finding that outgrowth embryos also secrete membrane vesicles is an important observation that may indicate how these cells are providing support for embryos in coculture.
An interesting question remains as to the content of these vesicles in our system. Human trophoblast cell lines are known to secrete development-promoting molecules such as hCG and vascular endothelial growth factor. 20,31,38,63 –69 The importance of hCG in successful pregnancy is recognized, and this hormone is used both as an indicator of successful pregnancy and to promote embryonic development. 70,71 Notably, hCG is stored in the zona pellucida of embryos, 72,73 and these reserves may play a role in attachment, invasion, and implantation during pregnancy. 74 Given the improvement in both embryonic health and implantation, one factor secreted by outgrowth embryos may be murine hCG. However, multiple factors are differentially expressed between embryos that either progress to pregnancy or fail to implant, including metabolic, proteomic, and secretomic. 41 Importantly, implantation was associated with secretomes containing granulocyte macrophage–colony-stimulating factor, insulin-like growth factor 1, and CXC motif chemokine 13. These factors may also be contributing to implantation competence via positive conditioning. 41,75 Whether the vesicles we observed on outgrowth embryos contain any of these factors is an important area for further study.
In addition to the secretomes of embryos, endometrial secretions have been recognized as complex and important regulators of uterine physiology that interact with embryos to promote survival and successful pregnancy. 76,77 Unsurprisingly, these various factors are known to regulate the expression of genes critical for survival and death. Apoptosis is the process of programmed cell death that may occur in multicellular organisms. 78 –80 During mouse embryonic development, multiple antiapoptotic and proapoptotic genes are expressed and regulated by maternal factors in early stages and by embryonic factors in later stages. 81 –84 Our data support the observation that embryonic factors modulate apoptosis; indeed, apoptosis was decreased in cocultured embryos, likely due to the significant upregulation of Survivin and Lif mRNA. Because the cocultured embryos also had a greater implantation rate, we conclude that upregulation of survival genes is a beneficial effect of coculture that can increase the rate of implantation. These embryotrophic effects of coculture with an outgrowth embryo should be substantiated by comparing the developmental competence of in vivo embryos in the further studies.
Although we clearly showed that coculture with outgrowth embryos improved preimplantation embryonic development in vitro and implantation competence in utero, there were no differences in the peri-implantation embryonic developmental competence investigated by trophoblastic outgrowth area and morphological grading of outgrowth embryos in vitro. There were 2 explanations for the inconsistency between the preimplantation development and the outgrowth assay. First, the in vitro culture condition for outgrowth stage was a similar environment to the previous culture conditions, which the cells had already adapted to. Thus, the effect of the coculture could be masked until additional environmental signals test the fitness of the cells, as are encountered during implantation in utero. Second, we only used well-developed blastocysts in our outgrowth assays, thus, we may have preselected cells with similar developmental competence from both the coculture and the control groups.
Mouse models are routinely used to improve in vitro embryo culture techniques to increase the success of human IVF-ET program. 14,17,20 However, there are limitations to in vitro culture system, as the potent development and implantation have less than those from in vivo environment. 13,51,85 Our study provides a novel coculture system using outgrowth embryos to enhance mouse preimplantation and peri-implantation blastocyst development, thus enabling the study of mouse embryonic development in more detail. Additionally, our findings raise an interesting question as to whether the addition of certain secretomes from trophoblasts to human embryo culture could improve in vitro fertilization outcomes.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by a grant of the