
Abstract
The molecular background of estrogen receptor (ER) expression is important to understand the pathophysiology of the high estrogen environment in endometriosis. However, the molecular details have not been fully understood. The objective of this study is to evaluate the molecular background of ERα and ERβ messenger RNA (mRNA) expression in endometriotic cells. The following summarizes our observations: (1) ERα mRNA expression in endometriotic cells was estimated to be approximately one-tenth of that in endometrial cells. (2) Three mRNAs, which include 3 different 5′-untranslated exons tagged to an open reading frame of wild-type ERα, were detected. (3) Expression of ERβ mRNA depends mostly on 0N promoter and includes 2 open reading frames: one for a wild-type ERβ1 and another for a splice variant ERβ2. (4) Expression of ERβ1 mRNA was approximately 40-fold higher than that in endometrial cells. (5) Expression of ERβ2 mRNA was almost at a comparable level of the ERβ1. 9 (6) ERα and ERβ mRNAs are equivalently expressed in endometriotic cells. These observations show the molecular background of ER mRNA expression in endometriotic cells and provide a clue to further understanding the estrogen-dependent pathophysiology leading to clinical application in endometriosis.
Introduction
Endometriosis is an estrogen-dependent, inflammatory disease characterized by the presence of endometrium-like tissue primarily on the pelvic peritoneum and ovaries. The role of estrogen is obvious because the symptoms associated with endometriosis often disappear after menopause, and the administration of GnRH agonists or progestin relieves the pelvic lesions and associated pain. A marked upregulation of aromatase gene associated with aberrant DNA demethylation 1,2 leads to a high estrogen environment in endometriotic tissues. Its inhibition by aromatase inhibitor is effective to reduce endometriosis-associated symptoms. 3 One of the important issues to be clarified is a further understanding of the role of estrogen-dependent growth mechanism in endometriotic tissues. To explain the pathophysiology leading to clinical applications, distinct profiles of estrogen receptor (ER) expression have been documented: a higher ERβ and a lower ERα expression in endometriotic tissues, which is in the inverse relationship in endometrium. 4 –7 Estrogen receptor α is the dominant receptor in the uterus and mediates a number of estrogen actions, including the stimulation of proliferation and the induction of progesterone receptor expression. In contrast, ERβ antagonizes the inflammatory and proliferative actions of ERα. 8,9 Therefore, the ER expression profile has been proposed as a major background of estrogen action in endometriosis. However, the molecular details of ER expression in endometriotic tissues have not been fully understood so far.
Recently, several splice variants of ERβ showing distinct transcriptional activities have been identified, and their roles along with the classical wild type have been extensively re-evaluated. 10 –13 These findings prompted us to re-evaluate the molecular background of ER expression in endometriosis. In this study, we focused on the mRNA expression in endometriotic cells.
We show the molecular background of ER mRNA expression in endometriotic cells. We observed the expression of a splice variant ERβ2 mRNA along with the wild-type ERβ1 in endometriotic cells. Importantly, these ERβ transcripts were expressed almost at a comparable level of the ERα in endometriotic cells.
Materials and Methods
Patients
The institutional review boards of Tottori University Faculty of Medicine approved this project. We obtained informed consent from all patients. Endometrial and endometriotic tissues were obtained from patients as described previously. 1,2,14 –16 Endometrial tissues were obtained from the uteri of (n = 19) cycling premenopausal women who underwent hysterectomy for uterine leiomyoma during the proliferative or secretory phase. The chocolate cyst lining of the ovaries of (n = 21) patients with endometriosis was the source of endometriotic tissue collected during the follicular or luteal phase. These patients had received no hormonal treatment before surgery. The menstrual cycle phase was determined by measuring serum estradiol and progesterone levels as well as by histological examination.
Endometrial and Endometriotic Stromal Cells
Stromal cells were collected from endometrial and endometriotic tissues as described. 1,2,14 –16 In brief, the endometrial and endometriotic tissues were minced and digested with 0.5% collagenase. The dispersed cells were filtered through a 70 μm nylon mesh to remove the undigested tissue pieces containing the glandular epithelium. The filtered fraction was separated further from epithelial cell clumps by differential sedimentation at unit gravity. The medium containing stromal cells was filtered through 40 μm nylon mesh. Final purification was achieved by allowing stromal cells, which attach rapidly to plates, to adhere selectively to culture dishes within 30 minutes at 37°C in 5% CO2 in air. To confirm the purity of the isolated stromal cells, immunocytochemical analysis was performed using cytokeratin (DAKO Corp, Kyoto, Japan) as a marker of epithelial cells, vimentin (DAKO Corp) as a marker of stromal cells, CD14 (Nichirei, Tokyo, Japan) as a marker of activated macrophages, and factor VIII F8/86 (DAKO Corp) as a marker of endothelial cells. The results showed that the purity of the stromal cells was more than 98%. Unless otherwise indicated, cells were cultured in DMEM/F12 supplemented with 10% fetal calf serum at 37°C in 5% CO2. At the first passage, endometrial and endometriotic stromal cells were plated at a density of 5 × 105 cells/60 mm dish and incubated to reach a confluence of 80% to 90% for RNA preparation.
Analysis of ER mRNA Expression
Total cellular RNA was prepared using the RNeasy Plus kit (QIAGEN, Tokyo, Japan) according to the manufacturer’s protocol. Using 2 μg of total RNA, single-stranded complementary DNAs (cDNAs) were prepared in 20 μL of reaction mixture containing 1 mmol/L each of deoxynucleotide, 20 units of PrimeScript RTase (Takara, Kyoto, Japan), 20 units of RNase inhibitor (Takara, Kyoto, Japan), and 20 μmol/L of oligo-dT16 primer at 42° C for 30 minutes. Then, 1 μL of the cDNA reaction mixture was subjected to polymerase chain reaction (PCR) amplification in a 25 μL Multiplex PCR Mixture (Takara, Kyoto, Japan) containing 50 pmol each of the respective forward and reverse primers. Using ERα-F (5′-CAGGGTGGCAGAGAAA-GATTGG-3′ in exon 2) and ERα-R (5′-GGTCTGGTAGGATCATACTCGG-3′’ in exon 4), a sequence of 532 bp in human ERα cDNA (Accession No: M12674) was amplified. Using ERβ-F (5′-CCTGCTGTGATGAATTACAG-3′ in exon 1) and ERβ-R (5′-TTCTCTGTCTCCGCACAAG-3′ in exon 4), a sequence of 550 bp in human ERβ cDNA (Accession No: NM001437) was amplified. The conditions for PCR were 30 seconds at 94°C, 15 seconds at 58°C (ERα) or 55°C (ERβ), and 30 seconds at 72°C for 38 cycles. At the end of the PCR cycles, 10 μL of the reaction mixture was removed and subjected to electrophoresis on a 1.8% agarose gel in Tris-Borate-EDTA buffer at a constant voltage of 100 V. Separated DNA sequence stained with ethidium bromide was visualized under UV light. As an internal control, β-tubulin mRNA was assayed in parallel.
Analysis of the Alternative 5′-Untranslated Exon
Using the cDNAs as described earlier, PCR was performed. Forward primers 17 in alternative 5′-untranslated exons of ERα are as follows: 1A-F: 5′-GGAGCTGGCGGGGGGCGTTG-3′; 1BF: 5′-CGCGTTTATTTTAAGCCCAG-3′; 1CF: 5′-CGGCCCTTGACTTCTACAAG-3′; 1DF: 5′-CTTCTTCACCTGAGAGAGCC-3′; 1EF: 5′-CAGAGAAATAATCGCAGAGC-3′; 1FF: 5′-CCAAAACTGAAAAT-GCAGGC-3′. The reverse primer, located in exon 2, was common to all reactions: 2R: 5′-CCTTGCAGCCCTCACAGGAC-3′. The primers 18 in alternative 5′-untranslated exons of ERβ are as follows: exon 0 N, 5′-AGATTTTTTAAATTTGAGATTGGGGTTG-3′ and 5′-CTTACCTTACAAATAAACACACC-3′; and exon 0 K, 5′-GTTGGGGTTATTTYGGGGTTGTT-3′ and 5′-CCTCCAACAAACACATTCA-3′. The PCR condition was 1 minute at 94°C, 1 minute at 62°C, and 1 minute at 72°C for 38 cycles. At the end of the PCR cycles, 10 μL of the reaction mixture was removed and subjected to electrophoresis on a 1.8% agarose gel. The separated DNA sequence was stained with ethidium bromide and visualized under UV light.
Analysis of ER Isoform Expression
The single-stranded cDNAs prepared from endometrial and endometriotic cells were used. For the analysis of ERα isoform expression, PCR with the forward primer H1 (5′-ACGGACCATGACCATGACCCT-3′ in exon 1) and the reverse primer HR5 (5′-GGTCTGGTAGGATCATACTCGG-3′ in exon 5) or HR8 (5′-TCAGACCGTGGCAGGGAAA-3′ in exon 8) was performed. For the analysis of ERβ isoform expression, PCR with primer sets of exon 7 and the variant-specific exon 8 was performed. 10 The forward primer, 5′-AGTATGTACCCTCTGGTCACAGCG-3′ in exon 7, is shared by all the ERβ isoforms. The isoform-specific reverse primers were as follows: ERβ1: 5′-CCAAATGAGGGACCACACAGCAG-3′ and ERβ3: 5′-GCAGTCAAGGTGTCGACAAAGGCTGC-3′. The reverse primer, 5′-CTTTAGGCCACCGAGTTGATTAGA-G-3′, was shared by ERβ2, ERβ4, and ERβ5.
Quantitative Real-Time PCR
Expression of ER mRNA was assessed using real-time PCR. As an internal control, β-tubulin mRNA expression was estimated in parallel. One μL of each cDNA samples prepared from endometrial and endometriotic stromal cells (∼50 ng) as indicated above was subjected to the TaqMan real-time PCR in triplicate. The real-time PCR was performed using the Universal Probe Library system (Roche Diagnostics, Tokyo, Japan). The specific probe (ERα #69, ERβ1 #3, and β-tubulin #43) was selected on the website of Universal Probe Library Assay Design Center (https://www.roche-appliedscience.com/sis/rtpcr/upl/adc.jsp). The FastStart TaqMan Probe Master (Roche) was used for the reaction solution. Relative expression of ERβ1, ERβ2, and ERβ5 mRNA was estimated using SYBR Premix Ex TaqII kit (Takara, Kyoto, Japan) using the respective primer sets of exon 7 and exon 8 as follows: ERβ1: 5′-CCGGCTAACCTCCTGATGC-3′ and 5′-CCAAATGAGGGACCACACAGCAG-3′, ERβ2: 5′-AGTATGTACCCTCTGGTCACAGCG-3′ and 5′-AGGCCTTTTCTGCCCCTCGATGC-3′, and ERβ5: 5′-ATGGGATTATGTGCTCCACGGA-3′ and 5′-CTTTAGGCCACCGAGTTGATTAGAG-3′. ABI PRISM 7900HT Sequence Detection System (Applied Biosystems, Tokyo, Japan) was used for the real-time PCR.
Statistical Analysis
Results were analyzed using one-way analysis of variance followed by Fisher protected least significant difference test. Values were expressed as means ± standard error of mean. P < .05 was considered statistically significant.
Results
Expression of ERα mRNA
Expression of ERα mRNA was observed in both endometriotic and endometrial cells (Figure 1A). We then searched for the splice variant mRNA using reverse transcription-PCR with primer sets to amplify the open reading frame (ORF) of wild-type ERα (from exon 1 to exon 8 in ESR1). As shown in Figure 1B, only a single cDNA sequence, which includes an ORF predicting a wild-type ERα, was always amplified from cDNAs prepared from endometriotic and endometrial cells. The 5′-untranslated exons, C, D, and F, were tagged to exon 1 in endometriotic cells, while in endometrial cells, the B, C, D, and F were tagged to exon 1 (Figure 1C). Among these ERα mRNAs, the exon C-tagged mRNA was the most abundant. Relative expression of ERα mRNA in endometriotic cells was estimated to be approximately one-tenth of that in endometrial cells (Figure 1D).
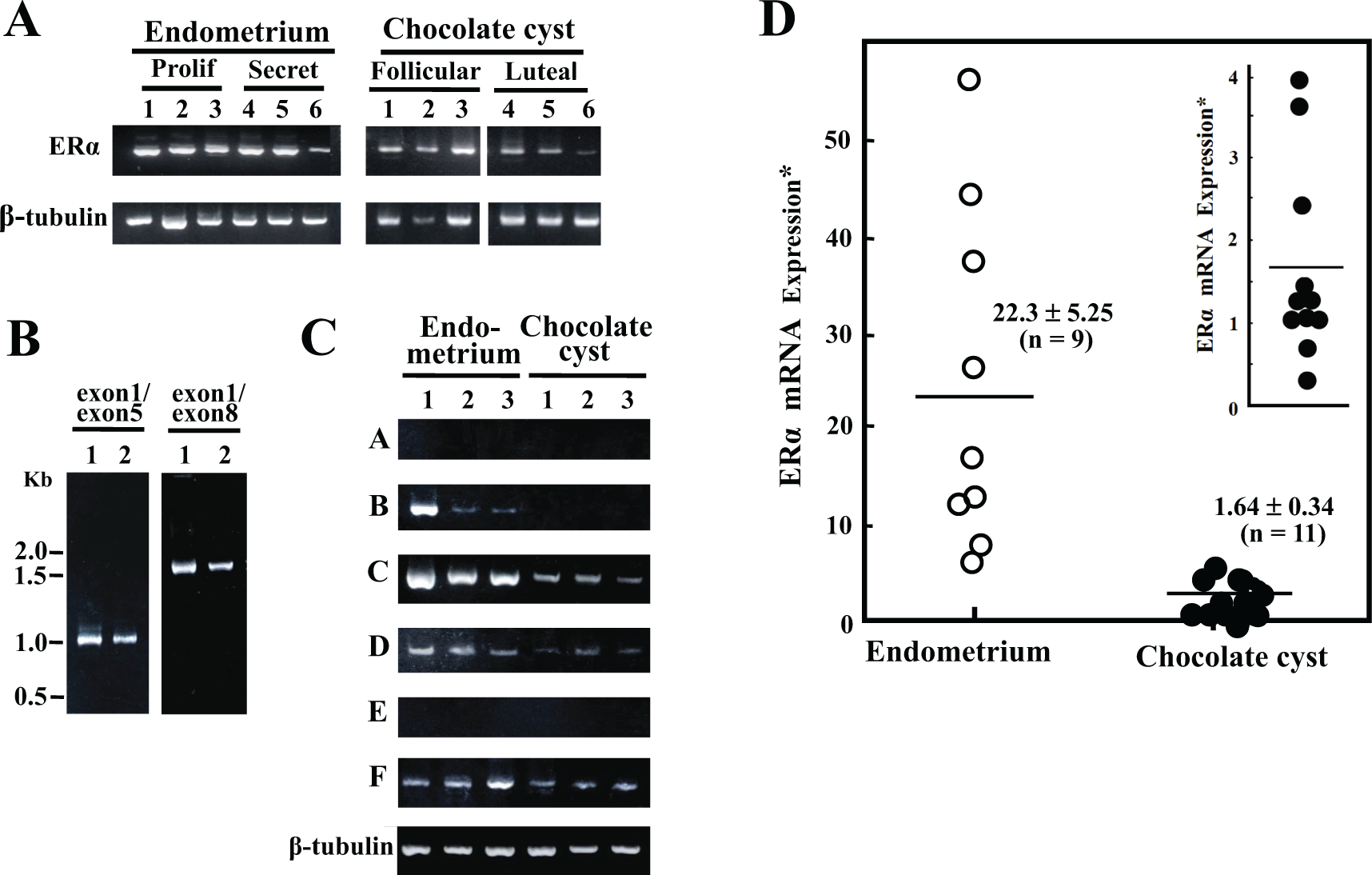
Estrogen receptor α (ERα) messenger RNA (mRNA) expression in endometrial and endometriotic cells. A, Analysis of ERα mRNA expression. Stromal cells were collected from 6 endometrial (3 from proliferative phase and 3 from secretory phase) and 6 endometriotic tissues (3 from follicular phase and 3 from luteal phase). Single-stranded complementary DNA (cDNA) was prepared from total cellular RNA and subjected to polymerase chain reaction (PCR). A sequence from exon 2 to exon 4 (532 bp in length) of ERα cDNA was amplified and subjected to a 1.8% agarose gel electrophoresis. As an internal control, β-tubulin mRNA was assayed in parallel. Amplified signals were visualized using ethidium bromide staining under UV light. B, Analysis of alternative splicing. Single-stranded cDNAs were prepared from endometrial (lane 1, proliferative phase) and endometriotic cells (lane 2, follicular phase). Following PCR using primer sets from exon 1 to exon 5 (1014 bp) and from exon 1 to exon 8 (1795 bp), amplified sequences were subjected to a 1.8% agarose gel electrophoresis and visualized. C, Analysis of untranslated first exons in ERα mRNA. Single-stranded cDNA was prepared from 3 endometrial (lanes 1 and 2 from proliferative phase and lane 3 from secretory phase) and 3 endometriotic cells (lanes 1 and 2 from follicular phase, and lane 3 from luteal phase). Following PCR using respective 6 forward primers in the untranslated exons, A, B, C, D, E, and F, and the reverse primer in the exon 2, amplified sequences were subjected to a 1.8% agarose gel electrophoresis and visualized. As an internal control, β-tubulin mRNA was assayed in parallel. D, Quantitative analysis of ERα mRNA expression. Single-stranded cDNAs from 9 endometrial (4 from proliferative phase and 5 from secretory phase) and 11 endometriotic cells (6 from follicular and 5 from luteal phase), which were randomly picked up from cells, were subjected to the TaqMan real-time PCR in triplicate. The specific probes (ERα mRNA, ESR1 #69, and β-tubulin #43) were selected on the website of the Universal Probe Library System. ABI PRISM 7900HT Sequence Detection System was used. Relative ERα mRNA expression was estimated using the β-tubulin mRNA expression as an internal control. *Values (mean ± SEM) are expressed as an arbitrary unit. Endometrium versus chocolate cyst, P < .001 inset, relative ERα mRNA expression in endometriotic cells is shown using a larger scale.
Expression of ERβ mRNA
Expression of ERβ mRNA was observed in both endometriotic and endometrial cells (Figure 2A). In endometriotic cells, the expression depends mostly on 0N promoter (Figure 2B) and includes 2 ORFs: one for a wild-type ERβ1 and another for a splice variant ERβ2 (Figure 2C). Expressions of ERβ2 and ERβ5 mRNA were detectable in endometrial cells (Figure 2C). The expression of ERβ1 mRNA in endometriotic cells was approximately 40-fold higher than that in endometrial cells (Figure 2D). The ERβ2 mRNA expression was almost at a comparable level of ERβ1 in endometriotic cells, while the ERβ5 mRNA expression was at a marginal level (Figure 2E).
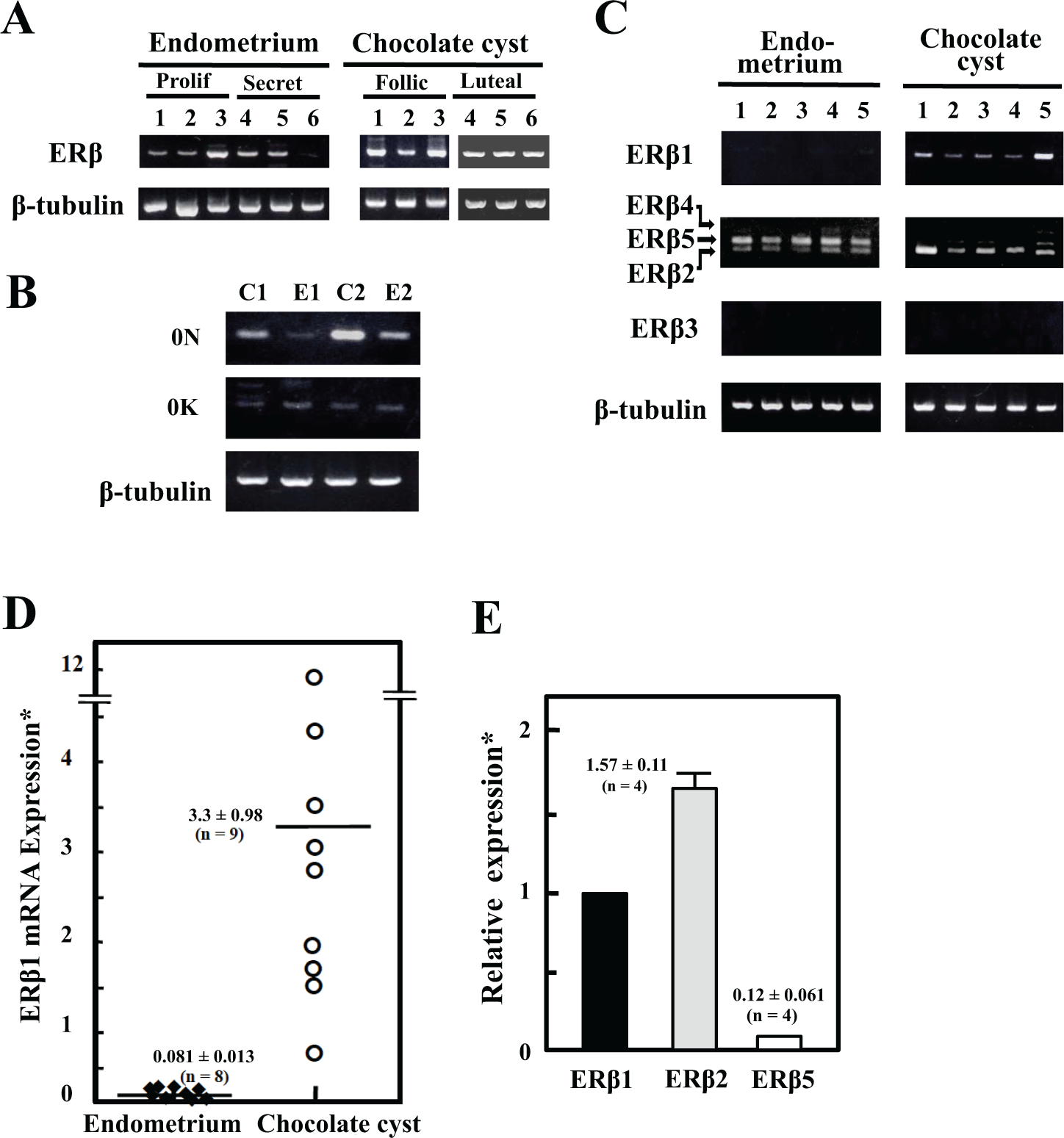
Estrogen receptor β (ERβ) messenger RNA (mRNA) expression in endometrial and endometriotic cells. A, ERβ mRNA expression. Stromal cells were collected from 6 endometrial (3 from proliferative phase and 3 from secretory phase) and 6 endometriotic tissues (3 from follicular phase and 3 from luteal phase). Single-stranded complementary DNA (cDNA) was prepared from total cellular RNA and subjected to polymerase chain reaction (PCR). An ERβ cDNA sequence from exon 1 to exon 4 (550 bp in length) was amplified and subjected to 1.8% agarose gel electrophoresis. As an internal control, β-tubulin mRNA was assayed in parallel. B, Analysis of the untranslated first exons. Single-stranded cDNA was prepared from endometrial (lanes E1 and E2) and endometriotic cells (lanes C1 and C2). PCR using 2 forward primers, 0 N and 0 K, and a reverse primer located in exon 2 was performed. As an internal control, β-tubulin mRNA was assayed in parallel. C, Analysis of alternative splicing. Single-stranded cDNAs were prepared from 5 endometrial and 5 endometriotic cells. Following PCR with primer sets of the variant-specific exon 7 and exon 8, amplified sequences were visualized on a 1.8% agarose gel electrophoresis. As an internal control, β-tubulin mRNA expression was assayed in parallel. Arrows indicate the location of amplified sequences corresponding to ERβ2, ERβ4 and ERβ5 mRNAs. D: Quantitative analysis of ERβ1 mRNA expression. Single-stranded cDNAs from 8 endometrial (4 from proliferative phase and 4 from secretory phase) and 9 endometriotic cells (6 from follicular phase and 3 from luteal phase) were prepared. The specific probes (ESR2 #3 for ERβ1 and β-tubulin #43) were selected using the Universal Probe Library System. ABI PRISM 7900HT System was used. Relative ERβ1 mRNA expression was estimated using the β-tubulin mRNA expression as an internal control. *Values are expressed as arbitrary unit (mean ± SEM). Endometrium versus chocolate cyst, P < .006. E, Relative expression of ERβ1, ERβ2, and ERβ5 mRNA in endometriotic cells. Using SYBR system with primer sets of variant-specific exon 7 and exon 8, the expression of ERβ1, ERβ2, and ERβ5 mRNAs was assayed. The expression was estimated using the β-tubulin mRNA expression as an internal control. Relative expression of ERβ2 and ERβ5 mRNA was normalized against ERβ1 mRNA expression. Values are the means ± SEM. ERβ1 versus ERβ2, P < .002, ERβ2 versus ERβ5, P < .006.
Discussion
The present study was undertaken to evaluate the molecular background of ER expression in endometriotic cells. So far, ER expression profile, a higher ERβ, and a lower ERα expression in endometriotic tissues, which is in the inverse relationship in endometrium, 4 –7 has been proposed as a major background of estrogen action in endometriosis. Although the ER mRNA expression is low in endometriotic cells (Figure 1D, 2D, and 2E), the expression of ERα and ERβ mRNAs is almost at a comparable level. It is interesting to note that ERβ2 mRNA 10,11,18 was found to be coexpressed along with ERα and ERβ1 mRNAs in endometriotic cells. These observations suggest a pathophysiological role of the ER expression profile in endometriosis.
Receptor dimerization in response to ligand binding is one of the crucial steps in ER-dependent transactivation. 19 Homodimerization of ERα accelerates cellular proliferation. 19 On the other hand, ERβ is thought to counteract the stimulatory effects of ERα through the formation of functional heterodimer with ERα. 20,21 Experimental evidence suggests that heterodimer-induced activation of target genes is markedly different from that induced by homodimers. 22,23 However, these studies have been conducted based on the ligand-dependent dimerization of wild-type ERs.
Recently, multiple ER isoforms have been identified in human tissues and cells. The ERα variants have been identified in a number of human cell types. More recently, ERβ variants with distinct biological functions have been identified. 11 –13 Markedly higher ERβ expression has been documented as one of the distinct gene expressions in endometriotic cells 4 –7 ; however, these observations were restricted to the expression of wild-type ERβ1, and the expression had been evaluated in comparison with that in endometrial cells.
We observed the expression of splice variant ERβ2 mRNA along with the wild-type ERβ1 in endometriotic cells (Figure 2C). More importantly, these ERβ transcripts were expressed almost at a comparable level of the ERα in endometriotic cells (Figure 1D, 2D and 2E). The observation suggests that the heterodimerization probably occurs in the cells. ERβ/ERβ and ERα/ERβ dimers are known to exhibit antiproliferative effects, 20,21 and ERα/ERβ is reported to be relatively more stable than the ERβ/ERβ homodimer. 13 Although ERβ2 exhibits no estrogen binding, ERβ1/ERβ2 heterodimer shows higher estrogen-induced transactivation than ERβ1/ERβ1 homodimer in a ligand-dependent manner. 11 To further understand the role of the ER expression profile may lead to the estrogen-dependent pathophysiology in endometriosis.
In conclusion, expression of ER isoforms, ERα and ERβ, is the major circumstance for estrogen action in endometriosis. Our results demonstrated that multiple ER isoforms, ERα, ERβ1, and ERβ2, are expressed simultaneously within endometriotic cells. However, little is known regarding the preference for formation of ER homodimer versus heterodimer and how this is affected by ER expression level and ligand occupancy. On therapeutic perspectives, promoting heterodimerization of ER isoforms using selective ER modulators may become an important strategy to design therapeutics for endometriosis. 24 To address these issues, clarifying the molecular basis of ER expression profile and the downstream ER-dependent gene expression may lead us to an avenue of further understanding the estrogen-dependent pathophysiology in endometriosis. It is interesting to note that ERβ and prostaglandin E2 integrate at Ras-like, estrogen-regulated, growth inhibitor, leading to the increase in endometriotic cell proliferation. 25
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported in part by a