
Abstract
Aim:
Although previous studies found that 1-time acute stress applied during follicle maturation impaired oocyte competence, it is unknown whether repeated chronic stress, which is known to cause animal behavioral adaptation, would damage oocytes when applied during follicle growth.
Methods and Results:
In this study, female mice were exposed to repeated restraint stress (RRS) or unpredictable stress (UPS) for different days before equine chorionic gonadotropin injection to initiate oocyte prematuration development and to observe effects of different stressors on oocytes in the growing follicles. The results showed that although oocyte pre- and postimplantation development was unaffected when mice were exposed to RRS or UPS once a day for 4 days, development was impaired when mice were exposed to RRS for 8 or more days or to UPS twice a day for 4 days (4 × 2). The 4 × 2 UPS caused more oxidative stress in oocytes and severer apoptosis in antral follicles than did the 4-day RRS. The RRS mice were stressed consistently from days 1 to 23 of restraint, and the stress that a mouse had 4 × 2 UPS was severer than that from 4-day RRS.
Conclusion:
The results suggest that (1) the degree that a stress damages oocytes is the product of duration × severity of the stress; (2) RRS impaired oocyte developmental potential through cumulative effects on growing follicles; and (3) preantral follicles were not as sensitive to stress as antral follicles were.
Introduction
Humans are often exposed to psychosocial stress, either acute or chronic. Studies suggest that psychological stress can exert detrimental effects on reproduction in women. 1 –6 Adverse effects of psychological stress on reproduction have also been reported in sows 7,8 and ewes. 9 Restraint of small animals is a mature experimental procedure developed and commonly used for studying psychogenic stress. 10,11 Mice and rats exposed to restraint stress during pregnancy exhibit impaired function of corpora lutea and reduced pregnancy rates and litter size. 12 –14
However, evidences for the direct effect of psychological stress on the oocyte are limited. Although our recent studies demonstrated that a 24-hour acute restraint stress applied to female mice during follicle maturation impaired the developmental potential of oocytes, 15,16 the underlying mechanisms are largely unknown. Humans are challenged not only by acute psychological stressors but also by chronic ones during their daily life. However, the effect of chronic stresses on oocyte competence has rarely been studied. Although a recent study indicated that chronic unpredictable stresses (UPSs) decreased oocyte developmental potential, 17 whether chronic stress applied repeatedly over multiple days during follicle growth will impair oocyte developmental potential has yet to be studied, because behavioral adaptation could develop when rodents were repeatedly exposed to a predictable stressor. 18,19 In addition, the use of ultrasonography has demonstrated that exposing dairy cows to elevated temperatures alters follicular growth and function. 20 However, although those studies suggested that heat stress affects not only the antral follicles emerging in the follicular wave but also the ovarian pool of small antral follicles, resulting in carryover effects on follicular function, 21 whether the preantral follicles are sensitive to stress has not been reported.
The objective of the present study was to determine (a) whether the chronic repeated restraint stresses (RRSs) on females, which occurred repeatedly over multiple days during follicle growth, would impair oocyte developmental potential via cumulative effects. To this end, female mice were exposed to RRS or UPS for different days during follicular growth. At the end of each stress episode, mice were either injected with equine chorionic gonadotropin (eCG) to obtain oocytes for observation of their developmental potential or tested for anxiety behavior or killed to collect blood and ovaries for hormone or cell apoptosis assays. Results indicted that (1) the degree that a stress damages oocytes is the product of duration × severity of the stress; (2) RRS impaired oocyte developmental potential through cumulative effects on growing follicles; and (3) preantral follicles were not as sensitive to stress as antral follicles were.
Methods
Unless otherwise specified, all chemicals and reagents used in the present study were purchased from Sigma Chemical Co (St Louis, Missouri).
Mice and Treatments
Mice of the Kunming strain were used at the age of 6 to 8 weeks. The mice were kept in a room with 14-hour/10-hour light–dark cycles, the dark starting form 8
For RRS treatment, an individual mouse was put in a microcage constructed by the authors, which was placed in an ordinary home cage. The microcage offered the same photoperiod and controlled temperature as in the home cage for the unstressed animals. While in the microcage, mice could move back and forth to some extent, but they could not turn around. Each restraint session lasted for 8 hours (from 8:00
For UPS treatment, mice were exposed to different stressors for 4 days either once daily (4 × 1) at 3:00
Oocyte Recovery
At the end of each stress session, mice were injected with eCG (10 IU intraperitoneal [ip]). Both stressed and unstressed control mice were killed at 48 hours after eCG injection by decollation to collect ovaries for the recovery of immature oocytes at the germinal vesicle stage. The large follicles on the ovary were ruptured in M2 medium to release cumulus–oocyte complexes (COCs). Only COCs with more than 3 layers of unexpanded cumulus cells and containing oocytes larger than 70 µm in diameter and with a homogenous cytoplasm were selected for in vitro maturation. To obtain in vivo matured oocytes, female mice were injected with human chorionic gonadotropin (hCG, 10 IU ip) at 48 hours after eCG injection. At 13 hours after hCG injection, the superovulated mice were killed, and the oviductal ampullae were broken in M2 medium to release freshly ovulated oocytes for in vitro fertilization (IVF).
Oocyte Maturation In Vitro
After being washed 3 times in M2 and once in the maturation medium, the COCs recovered for in vitro maturation were cultured in groups of around 30 in 100 μL drops of maturation medium at 37.5°C in a humidified atmosphere of 5% CO2 in air. The media used for oocyte maturation were TCM-199 (Gibco, Grand Island, New York) supplemented with 10% (v/v) fetal bovine serum (Gibco), 1 µg/mL 17β-estradiol, 24.2 mg/mL sodium pyruvate, 0.05 IU/mL follicle-stimulating hormone (FSH), 0.05 IU/mL luteinizing hormone, and 10 ng/mL epidermal growth factor.
Oocyte Activation
At 24 hours of maturation culture, oocytes were stripped of their cumulus cells by pipetting with a thin pipette in M2 containing 0.1% hyaluronidase. Oocytes were then incubated in activating medium (Ca2+-free CZB medium supplemented with 10 mmol/L SrCl2 and 5µg/mL cytochalasin B) for 6 hours at 37.5°C in a humidified atmosphere with 5% CO2 in air. At the end of the activation treatment, oocytes were examined with a microscope for activation. Oocytes were considered activated when each contained 2 well-developed pronuclei.
In Vitro Fertilization (IVF)
Masses of dense sperm were collected from the cauda epididymis of fertile male mice and were placed at the bottom of a test tube containing T6 medium supplemented with 10 mg/mL bovine serum albumin (BSA). After 3 to 5 minutes, the supernatant containing highly motile spermatozoa was removed and capacitated in the same medium under mineral oil at 37.5°C for 1.5 hours. At 13 hours of maturation culture, in vitro matured oocytes were washed in the fertilization medium (T6 containing 20 mg/mL BSA) and were placed in fertilization drops (around 30 oocytes per 100 µL drop). Capacitated sperm were added to the fertilization drops to give a final sperm concentration of about 1 × 106/mL. After 6 hours of incubation, oocytes were observed under a microscope for fertilization. Oocytes with 2 pronuclei and the second polar body were considered fertilized.
Embryo Culture
Activated oocytes and IVF zygotes were washed in M2 and regular CZB without glucose before culture in regular CZB without glucose. Glucose (5.5 mmol/L) was added to CZB when embryos were beyond 3- or 4-cell stages. Embryo development was examined at the end (day 4) of embryo culture. Although some of the blastocysts obtained were stained with Hoechst 33342 for cell number counting, others were used for embryo transfer.
Embryo Transfer
Female mice aged 8 to 10 weeks (28-35 g bodyweight) were used as pseudopregnant recipients. The IVF blastocysts were transferred to the uterine horns of the 2.5-day post coitus (pc) pseudopregnant recipients. Fifteen embryos were transferred to each recipient, 8 or 7 embryos per uterine horn. After the embryo transfer, the recipients were housed singly in cages until parturition.
Immunohistochemical Detection of Active Caspase 3 in Ovarian Sections
Ovaries were embedded in paraffin and sectioned serially at a thickness of 5 µm. Immunoperoxidase staining was performed on deparaffinized sections. Sections were incubated with 3% peroxide for 15 minutes to exhaust endogenous peroxidase activity and with 0.1% trypsin for 15 minutes at 37°C for antigen retrieval. After being washed in phosphate-buffered saline (PBS), the sections were incubated with rabbit antiactive caspase 3 (1:50, ab52302; Abcam Co, Ltd, Shanghai, China) or PBS (for negative control) overnight at 4°C, biotin-labeled goat antirabbit immunoglobulin (Ig) G (1:200, SP-9001; Zhongshan Golden Bridge Biotechnology Co, Ltd, Beijing, China) for 2 hours at 37°C, and then horseradish peroxidase-labeled streptavidin (1:200, SP-9001; Zhongshan Golden Bridge Biotechnology Co, Ltd) for 1 hour at 37°C. Sections were then immersed in diaminobenzidine solution for 2 minutes to develop color and counterstained with hematoxylin. Positive staining was revealed as brown spots and nonspecific staining of all cell nuclei as light blue. Follicles containing more than 10 caspase-3-positive cells were considered atretic. Each treatment was repeated 3 times with each replicate containing 4 ovaries from 2 mice. About 10 antral follicles and 5 to 7 pre-antral follicles were randomly observed on each section each ovary to calculate the percentages of atretic follicles.
Blood Serum Preparation and Ovarian Homogenization
Mice were killed by decollation, and trunk blood (about 1 mL) was collected into ice-cooled centrifugal tubes and centrifuged (1700×g, 10 minutes, 4°C) to separate serum. The serum collected was stored at −80°C until hormone assay. For ovarian homogenization, the ovaries were weighed and transferred to a homogenizer with 800 µL PBS per 100 mg ovarian tissue. Homogenization was carried out manually at room temperature. Homogenates were centrifuged (15 000×g, 10 min, 4°C), and the supernatant was collected for immediate use or stored at −80°C until use.
Hormone Assays
Radioimmunoassay for cortisol was conducted by the Central Hospital of Tai-An City using commercial kits from Jiuding Biomedical Techniques Co Ltd, Tianjin, China. The minimum levels of detection for assays of cortisol were 10 ng/mL. The intra- and interassay coefficients of variation were <10% and <15%.
Enzyme-linked immunosorbent assay (ELISA) for corticotrophin-releasing hormone (CRH) was conducted using a Mouse CRH ELISA kit purchased from Shanghai Lanji Biological Technology Company. Briefly, 100 µL of standards or samples were added in duplicate to wells of the microtiter plate, 50 µL of enzyme conjugate was added to each well, mixed thoroughly, and incubated for 1 hour at 37°C. Then, after the microtiter plate was washed and blot dried, 50 µL of substrates A and B were added to each well and incubated for 10 to 15 minutes at 37°C.
Enzyme-linked immunosorbent assay for corticosterone was performed using a ELISA kit purchased from Arbor Assays Company (Catalog Number K014-H1). Briefly, 50 µL of standards or samples were added in duplicate to wells of the microtiter plate. Assay buffer of 75 µL was added to the nonspecific binding (NSB) wells, and 50 µL of assay buffer was added to wells to act as maximum binding wells. Then, 25 µL of the DetectX Corticosterone Conjugate and 25 µL of the DetectX Corticosterone Antibody (except the NSB wells) were added to each well, and the titer plate was shaken for 1 hour at room temperature. After the plate was washed using the wash solution and blot dried by hitting plate onto paper towels, 100 µL of the 3,3′,5,5′-tetramethylbenzidine (TMB) substrate were added to each well and incubated for 30 minutes at room temperature.
The optical density of both CRH and corticosterone was read at 450 nm wavelength using a plate reader within 15 minutes after the reaction was terminated by adding 50 µL of the stop solution. The concentrations of CRH and corticosterone were calculated according to their respective standard curves.
Elevated Plus-Maze Test
The plus maze consisted of 2 open arms (30 × 6 cm), alternating in right angles with 2 closed arms (30 × 6 × 15 cm). The central platform delimited by the 4 arms was 36 cm2. The whole maze was elevated 50 cm above the floor. The animal’s behavior on the maze was recorded via a video camera mounted on the ceiling above the center of the maze. The camera was connected to an Any-maze video tracking motion analysis system (Stoelting, Wood Dale, Illinois) running on a personal computer. The temperature of the test room was maintained at 22°C to 25°C with 50 to 55 dB white noise continuously provided. Before the start of the test, naïve mice were individually placed in a rectangular plastic arena for 30 minutes in order to habituate them to the test environment. To start the test, a mouse was placed on the central platform, facing an open arm, and was allowed to explore the maze for 5 minutes. Following a 4-paw criterion, number of entries and time spent on each arm over the total exploration of both open and closed arms were calculated using the Any-maze software (Stoelting Co., Wood Dale, IL, USA). The device was cleaned with 10% ethanol after each trial to effectively remove the scent of the previously tested animal. Although the mice exposed to restraint stress were elevated plus-maze (EPM) tested immediately at the end of each stress session, mice exposed to UPS were given some time for recovery before the EPM test. This recovery time was 30 to 60 minutes after 8 to 10°C and 31°C swimming stress and 5 minutes after hot stress and shaker stress.
Assay for Intracellular Glutathione
Samples (each containing 35-40 cumulus-free oocytes) were frozen and thawed 3 times. The concentrations of total glutathione (GSX) in oocytes were determined by the 5,5′-dithiobis (2-nitrobenzoic acid)-oxidized glutathione (GSSG) reductase recycling assay. Standards (0.01, 0.02, 0.1, 0.2, and 1.0 mmol/L) of GSX and a sample blank without GSX were also assayed. The amount of GSX in each sample was divided by the number of oocytes to determine the intracellular GSX concentration per oocyte (pmol/oocyte). The reduced glutathione (GSH) values were calculated from the difference between GSX and GSSG values for each oocyte.
Data Analysis
There were at least 3 replicates for each treatment. Each replicate contained either different numbers of oocytes or different numbers of animals as specified in different sections. Percentage of data was arc sine transformed and analyzed with analysis of variance; a Duncan multiple comparison test was used to locate differences. The software used was SPSS (SPSS 11.5; SPSS Inc, Chicago, Illinois). Data were expressed as mean ± standard error of the mean, and P < .05 was considered significant.
Results
Effects of Stress Duration (Days) and Severity on Oocyte In Vitro Development After Sr2+ Activation
Female mice were injected with eCG after being exposed to RRS or UPS for different days. Age-matched unstressed mice were also injected with eCG to serve as controls. At 48 hours after eCG injection, mice were killed to recover oocytes for in vitro maturation, SrCl2 activation, and embryo development. Rates for oocyte maturation and activation (ranging from 96% to 100%) did not differ (P < .05) between treatments (data not shown). Although percentages of 4-cell embryos and blastocysts and cell counts per blastocyst did not differ between mice restrained for 4 days and the unstressed control mice, all these developmental parameters decreased significantly in mice restrained for 8 or more days (Table 1). Although the developmental parameters did not differ between control mice and mice subjected to UPS once a day for 4 days (4 × 1), they decreased significantly in mice exposed to UPS twice a day for 4 days (4 × 2). The results suggested that the extent that a stress damages oocytes was the product of duration × severity of the stress. Thus, at the same severity, stress damage to oocytes increased with duration (sessions or days), and at the same duration, the damage increased with severity.
Effects of Duration (Days) and Severity of Maternal Stress on Oocyte In Vitro Development After Sr2+ Activation.
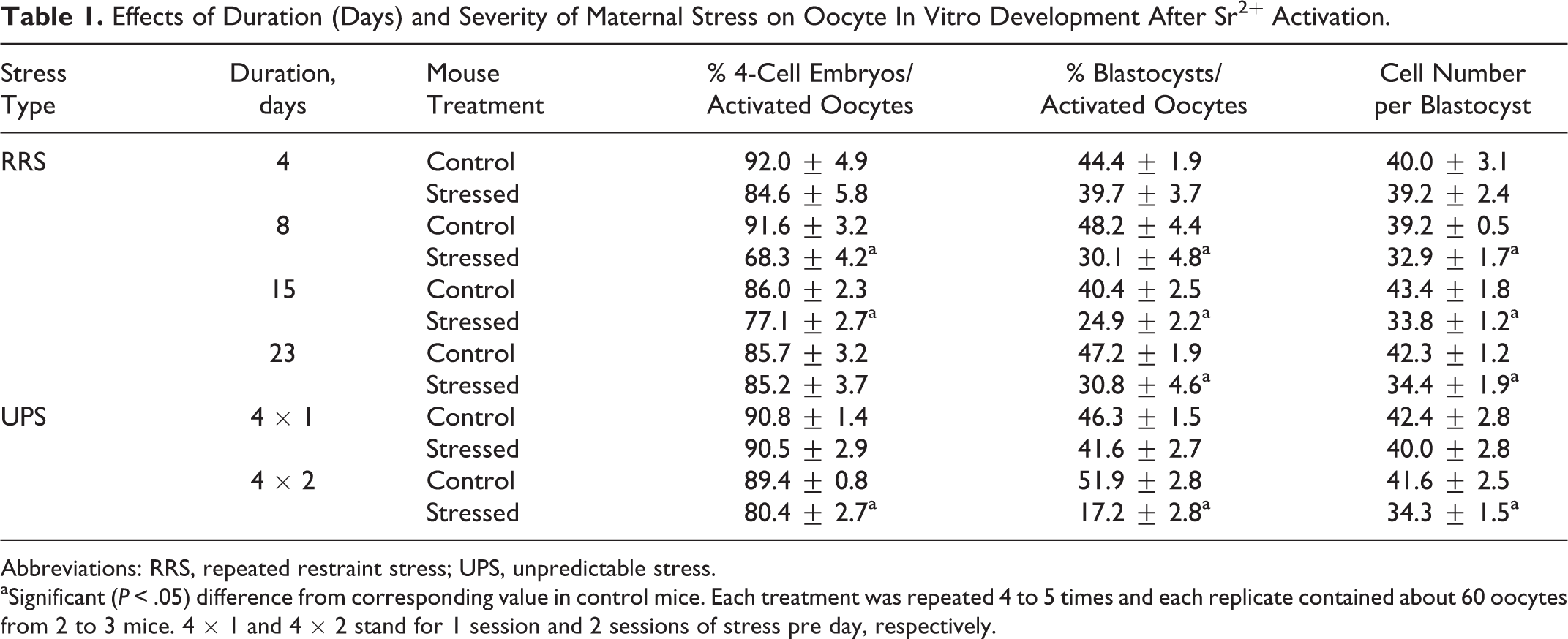
Abbreviations: RRS, repeated restraint stress; UPS, unpredictable stress.
aSignificant (P < .05) difference from corresponding value in control mice. Each treatment was repeated 4 to 5 times and each replicate contained about 60 oocytes from 2 to 3 mice. 4 × 1 and 4 × 2 stand for 1 session and 2 sessions of stress pre day, respectively.
Effects of Stress Severity on Oocyte Fertilization and Postimplantation Development
To further verify effects of stress severity on oocyte fertilization and postimplantation development, female mice were exposed to RRS or 4 × 2 UPS for 4 days before being injected with eCG. At 48 hours after eCG injection, some mice were killed to recover oocytes for in vitro maturation, others were injected with hCG to recover in vivo matured oocytes. The proportion of mice that ovulated after hCG injection did not differ significantly (P > .05) between control mice (87.5%, 7 of 8) and mice exposed to RRS (75.0%, 6 of 8) or 4 × 2 UPS (77.8%, 7 of 9) for 4 days. The average number of oocytes obtained from ovulating mice did not differ (P > .05) between control mice (29.6 ± 3.7) and mice exposed to RRS (32.8 ± 2.0) or 4 × 2 UPS (28.3 ± 1.6) for 4 days. The results suggested that these stress procedures had a mild effect on mouse estrus cyclicity, and they affect mouse reproduction mainly by impairing oocyte competence.
Results from both in vitro and in vivo matured oocytes showed that percentages of fertilized oocytes and blastocysts or cell counts per blastocyst did not differ between oocytes from RRS mice and oocytes from control mice, but all the parameters decreased significantly in oocytes from 4 × 2 UPS mice (Table 2). Embryo transfer showed that while percentage of term pregnancy and birth weight of pups did not differ among different treatments, live pups per pregnant recipient and live pups/embryos transferred decreased significantly in 4 × 2 UPS oocytes compared to control and RRS oocytes (Table 3).
Fertilization and Embryo Development in Vitro After Maturation in Vitro or in Vivo of Oocytes Recovered From Unstressed Control Mice or Mice Exposed to 4-Day RRS or 4 × 2 UPS.
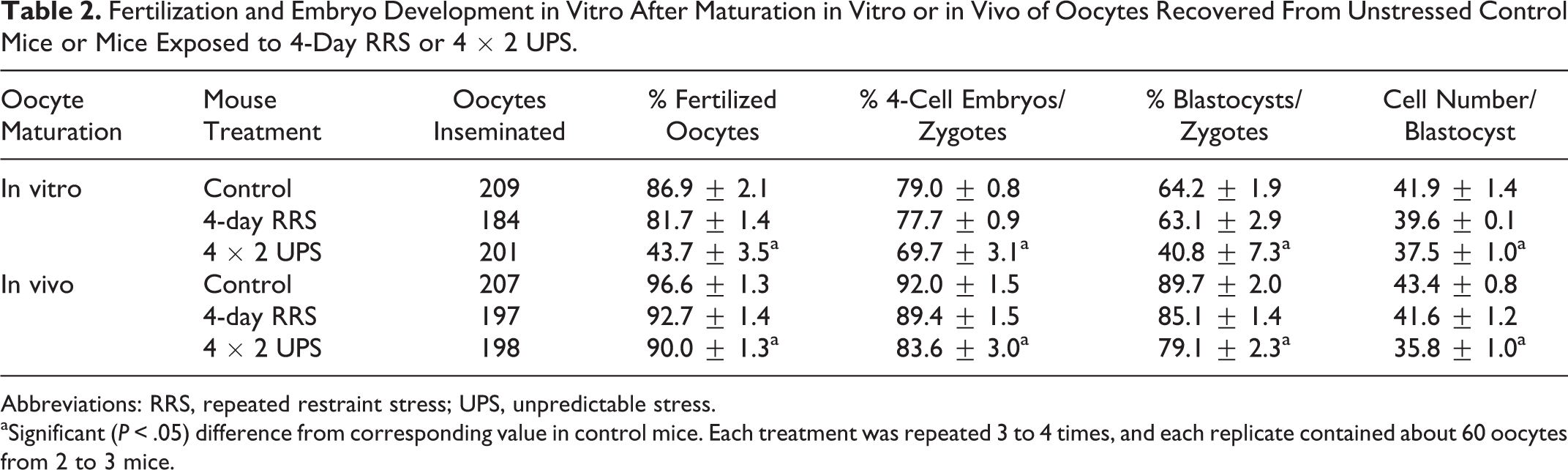
Abbreviations: RRS, repeated restraint stress; UPS, unpredictable stress.
aSignificant (P < .05) difference from corresponding value in control mice. Each treatment was repeated 3 to 4 times, and each replicate contained about 60 oocytes from 2 to 3 mice.
Pregnancy and Birth of Live Young After Embryo Transfer Following In Vitro Maturation of Oocytes Recovered From Unstressed Control Mice or Mice Exposed to 4-Day RRS or 4 × 2 UPS.

Abbreviations: ANOVA, analysis of variance; RRS, repeated restraint stress; UPS, unpredictable stress.
aSignificant (P < .05) difference from corresponding value in control mice. Each treatment was repeated 12 to 13 times with each replicate containing 1 mouse. While data for live pups per pregnant recipient and for birth weight of pups were analyzed with ANOVA as described in the Materials and Methods section, percentage data for term pregnancy and for live pups/embryos transferred were analyzed with chi-square test.
Effects of 4-Day RRS and 4 × 2 UPS on Apoptosis of Preantral and Antral Follicles
Female mice were exposed to RRS or 4 × 2 UPS for 4 days before being killed to recover ovaries for immunohistochemical analysis of active caspase 3 expression. While preantral and antral follicles that contained no or less than 10 apoptotic cells were judged as nonatretic (Figure 1A and B), follicles showing more than 10 caspase 3-positive cells were considered atretic (Figure 1C-E). Although about 17% of the antral follicles from unstressed control mice underwent atresia (Figure 1F), none of the preantral follicles observed in control (0 of 69), RRS (0 of 80), or UPS mice (0 of 63) could be judged as atretic. Percentages of atretic antral follicles increased significantly following UPS (Figure 1F). The results suggested that (a) 4 × UPS was more harmful than the 4-day RRS in causing follicular atresia and (b) preantral follicles were not as sensitive to stress as antral follicles were.
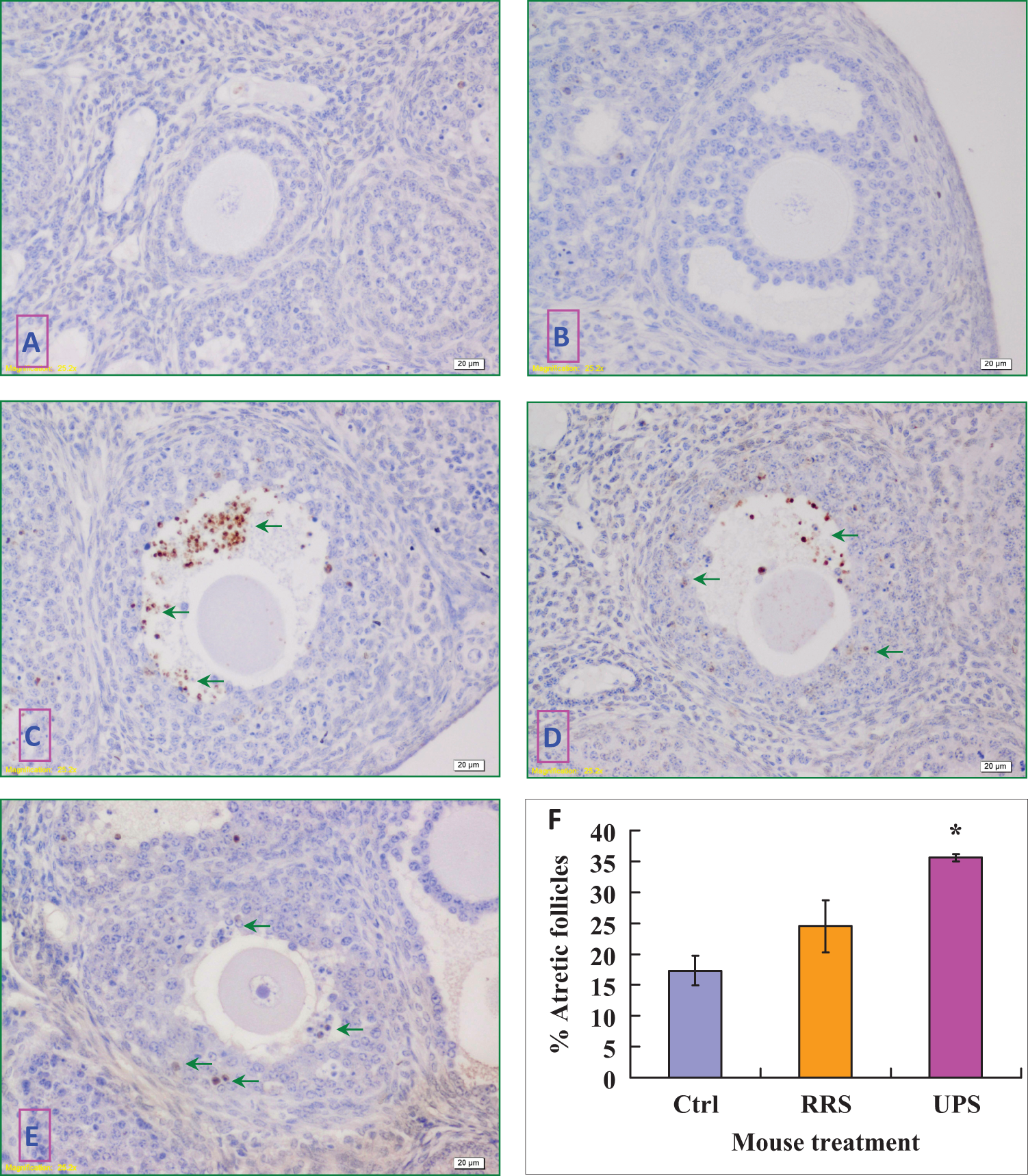
Immunohistochemical localization of active caspase 3 in the mouse ovary and percentages of atretic follicles with apoptotic cells. In the images, positive caspase 3 staining appears as brown spots and nonspecific staining of nuclei as light blue. The bar is 20 µm. While follicles with no apoptotic cells were considered nonatretic, those showing numerous apoptotic cells (arrows) were classified as atretic follicles. A, Nonatretic preantral follicles from 4 × 2 UPS mice. B, Nonatretic antral follicles from unstressed control mice. C, Atretic follicles from 4 × 2 UPS mice. D, Atretic follicles from 4-day RRS mice. E, Atretic follicles from unstressed control mice. F, A graph showing the percentages of atretic antral follicles in unstressed control (Ctrl), 4-day RRS and 4 × 2 UPS mice. Each treatment was repeated 3 times with each replicate containing 4 ovaries from 2 mice. *Significant (P < .05) difference from corresponding value in control mice. UPS indicates unpredictable stress. (The color version of this figure is available in the online version at http://rs.sagepub.com/.)
Effects of Restraint Duration on Bodyweight, Levels of Serum Glucocorticoid, and Anxiety-Like Behavior of Mice
To understand the mechanisms for the cumulative effect of RRS on oocyte competence, effects of restraint duration on bodyweight, levels of serum glucocorticoids, and anxiety-like behavior were observed. Corticosterone levels were measured along with cortisol levels to confirm that cortisol levels were as representative as corticosterone levels in mice in response to our stress systems. The results showed that RRS decreased mouse bodyweight significantly, and the difference between stressed and control mice was more obvious with increasing stress durations (Table 4). On all the restraint days, while the levels of cortisol or corticosterone were higher, the time that mice spent in the open arm was shorter significantly in stressed than in control mice (Figure 2). The results suggested that our restraint system stressed the animals consistently and that the accumulation of hypothalamic–pituitary–adrenal (HPA) products over a long stress period may account for the cumulative effect of RRS on oocyte developmental potential.
Effects of Different Stressors on Bodyweight of Mice.
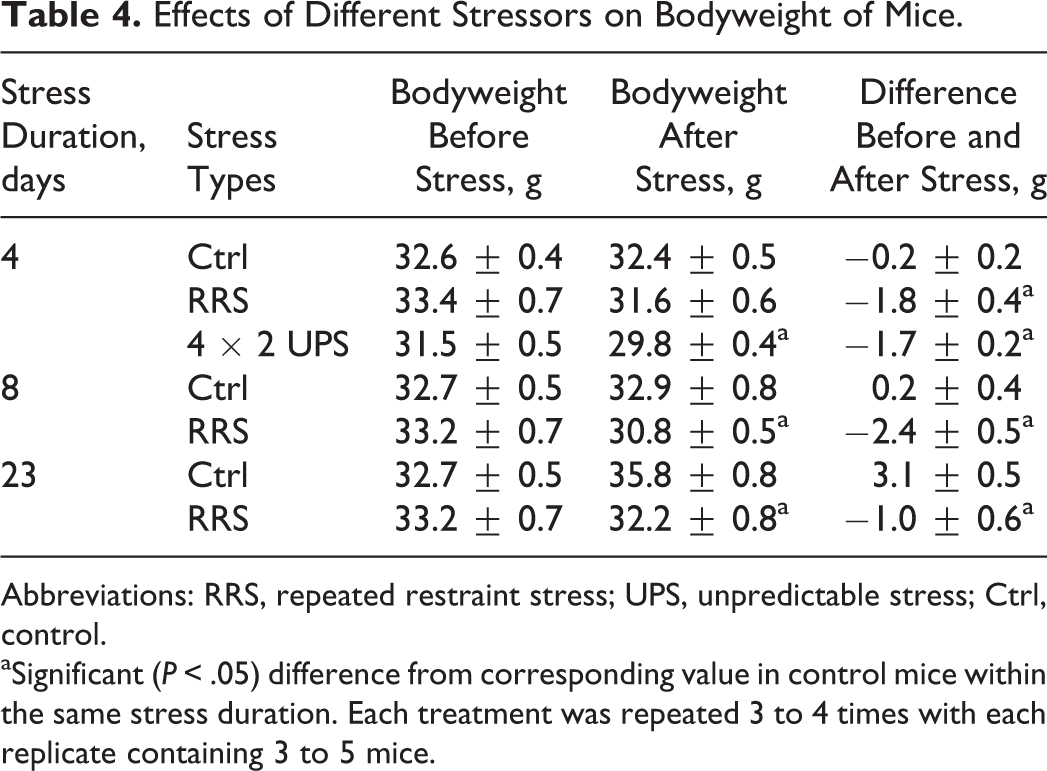
Abbreviations: RRS, repeated restraint stress; UPS, unpredictable stress; Ctrl, control.
aSignificant (P < .05) difference from corresponding value in control mice within the same stress duration. Each treatment was repeated 3 to 4 times with each replicate containing 3 to 5 mice.
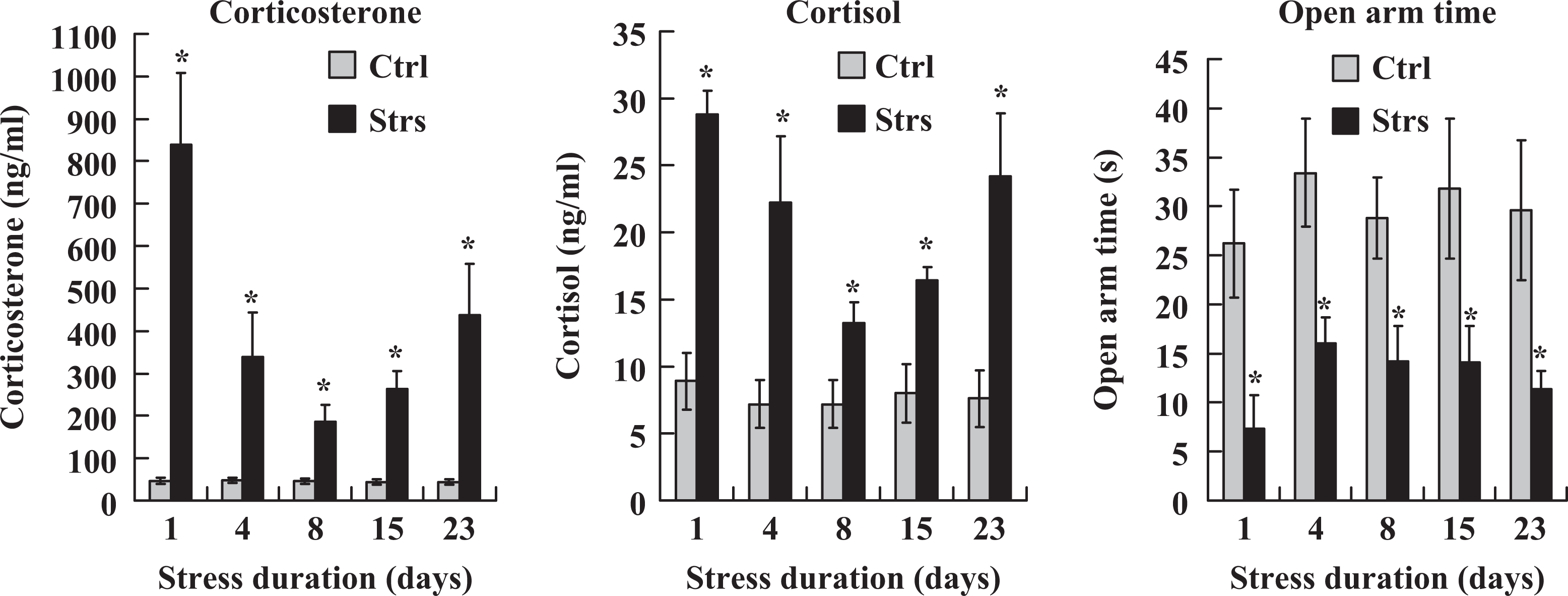
Serum levels of corticosterone and cortisol and the time (s) spent in the open arm during the Elevated Plus-Maze (EPM) test in unstressed control (Ctrl) mice and mice restraint-stressed (Strs) for different days. For hormone assays, each treatment was repeated 4 to 6 times with each replicate containing 1 mouse. For EPM test, each treatment was repeated 3 to 4 times with each replicate containing 3 to 4 mice. *Significant (P < .05) difference from corresponding value in control mice.
Bodyweight, Serum Cortisol, and EPM Performance of Mice Following RRS or UPS
To verify the severity of RRS and UPS, their effects on bodyweight, serum cortisol, and EPM performance were observed in mice exposed to 4-day RRS or 4 × 2 UPS. Bodyweight loss was significantly pronounced in mice exposed to 4-day RRS or 4 × 2 UPS relative to that in control mice (Table 4). On each day of stress, while the serum cortisol was higher, the time spent in the open arm during the EPM test was lower significantly in stressed animals than in controls in both stress systems (Figure 3). To further compare the severity between RRS and UPS, the control and stress (C–S) difference values in serum cortisol level and open arm time were calculated between control and stressed animals. The C–S difference value for serum cortisol level was calculated by subtracting the value of control mice from that of stressed mice, while the C–S difference for the open arm time was calculated by subtracting the value of stressed mice from that of control mice. The results indicated that the total C–S difference values for both serum cortisol and open arm time were significantly higher following UPS than following RRS (Figure 4). Thus, while the total C–S difference value of serum cortisol following RRS was (31.1 + 20.6 + 24.0 + 29.7) − (9.1 + 8.6 + 8.5 + 13.8) = 65.4, that following UPS was (31.7 + 35.9 + 29.9 + 38.0) − (9.0 + 9.7 + 10.2 + 11.4) = 95.2. Likewise, the total C–S difference value of open arm time following RRS was (30.9 + 30.7 + 30.3 + 24.3) − (18.1 + 16.2 + 19.0 + 17.8) = 45.1, whereas that following UPS was (32.2 + 31.8 + 34.1 + 32.3) − (15.5 + 13.5 + 9.0 + 9.1) = 83.3. The results confirmed that the stress that animals suffered from UPS was severer than that they suffered from RRS.
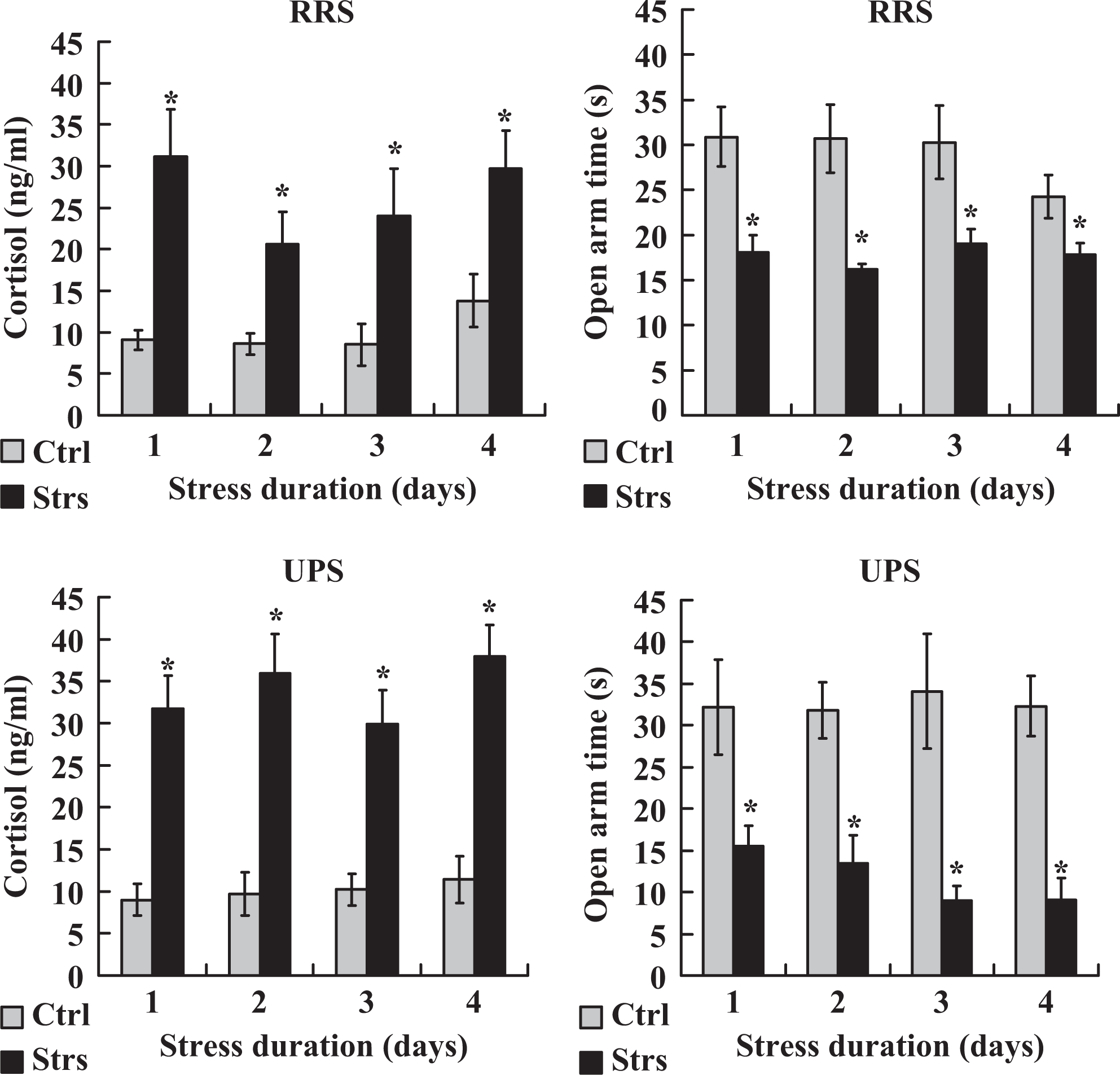
Serum levels of cortisol and the time (s) spent in the open arm during the EPM test in unstressed control (Ctrl) mice and mice stressed (Strs) for different days by RRS or 4 × 2 UPS procedures. For hormone assay, each treatment was repeated 5 to 7 times with each replicate containing 1 mouse. For EPM test, each treatment was repeated 3 times with each replicate containing 3 to 4 mice. *Significant (P < .05) difference from corresponding value in control mice. EPM indicates elevated plus-maze; UPS, unpredictable stress.
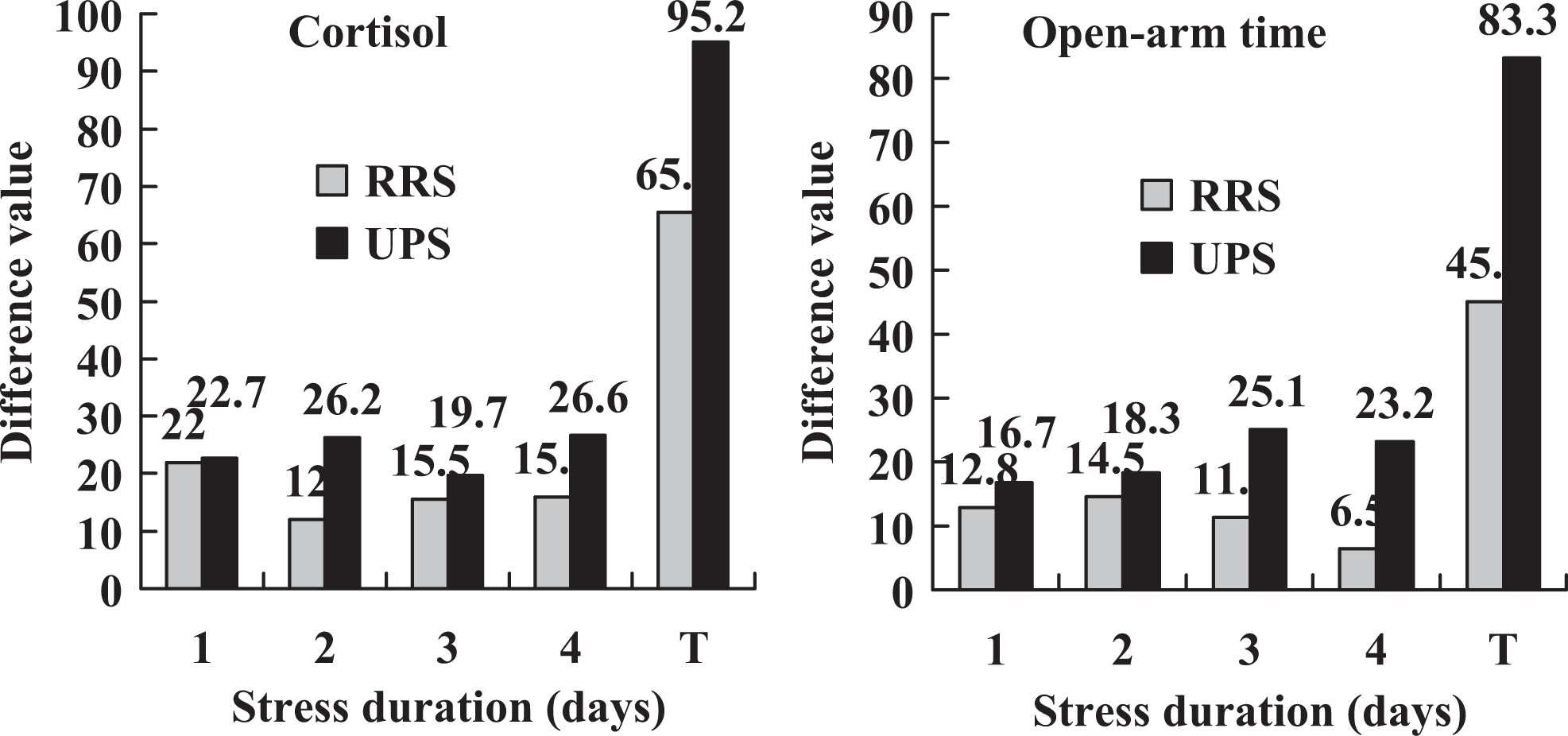
A comparison of the C–S difference values in serum cortisol level and open arm time between RRS and 4 × 2 UPS stresses. The C–S difference values were calculated using data from Figure 3. Although the C–S difference value for serum cortisol was calculated by subtracting the value of control mice from that of stressed mice on each day of stress, that for open arm time was calculated by subtracting the value of stressed mice from that of control mice. T indicates total C–S difference value that represents a sum of the difference values calculated from 4 days. C–S indicates control and stress; UPS, unpredictable stress.
Serum and Ovarian CRH Levels of Mice Following RRS or UPS
To compare the severity of RRS and UPS, concentrations of CRH in serum and ovarian homogenates were assayed by ELISA. Female mice exposed to 4-day RRS or 4 × 2 UPS were killed on each day of the stress to collect blood and ovaries for hormone assays. On each day of stress, the CRH levels in both serum and ovary were higher in UPS mice than in RRS mice (Figure 5). Again, the results confirmed that the stress that mice suffered from UPS was severer than that from RRS.
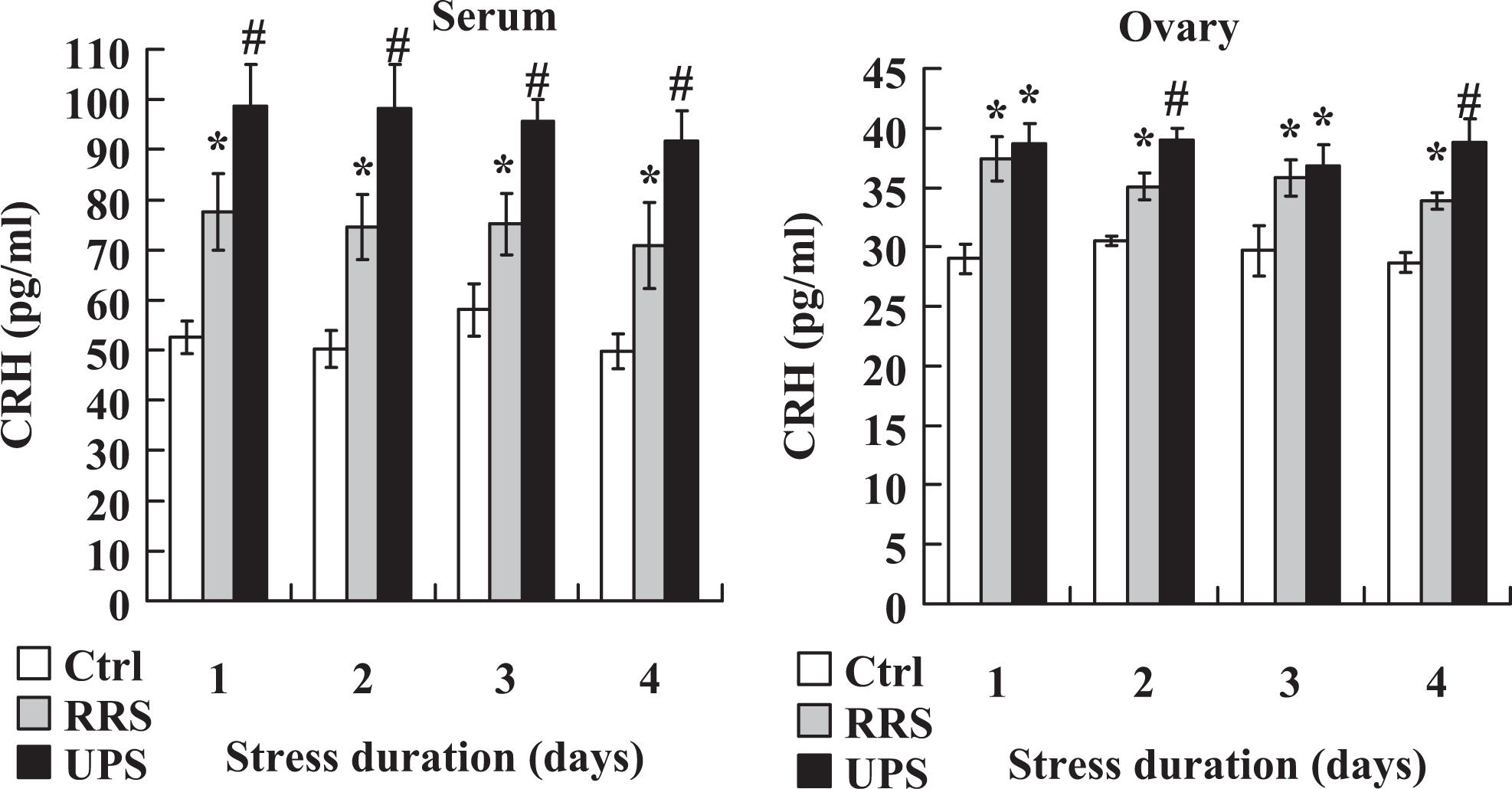
Concentrations of CRH in serum and ovarian homogenates of unstressed control mice (Ctrl) and mice stressed for different days by the RRS or 4 × 2 UPS procedures. Each treatment was repeated 5 to 6 times with each replicate containing 3 mice. *Significant (P < .05) difference from corresponding value in control mice. #Significant (P < .05) difference from corresponding value in both control mice and RRS mice. CRH indicates corticotrophin-releasing hormone; RRS, repeated restraint stress; UPS, unpredictable stress.
Effects of Stress Severity on Levels of Intraoocyte GSH
To gain more evidence for the effect of stress severity on ooplasmic maturation, intraoocyte GSX contents and GSH–GSSG ratio were assayed following in vitro maturation of oocytes recovered from different mouse groups. Although the GSX content did not differ, the GSH/GSSG ratio decreased significantly in stressed mice as compared to that in the unstressed control mice (Figure 6). Furthermore, the GSH–GSSG ratio was significantly lower in mice exposed to 4 × 2 UPS than in mice exposed to 4-day RRS. The results suggested that 4 × 2 UPS caused more oxidative stress in oocytes than did the 4-day RRS.
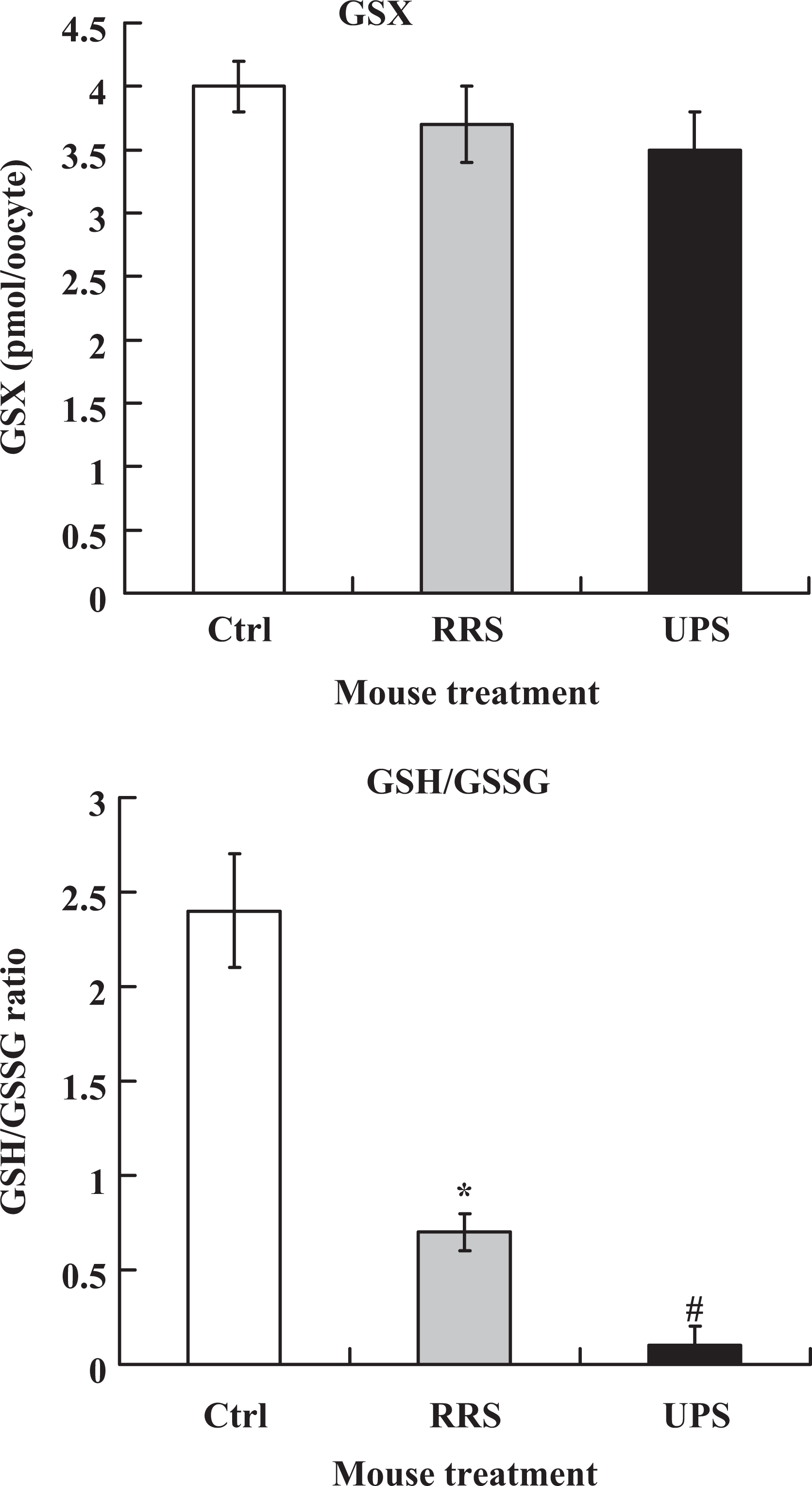
Intraoocyte GSX contents and GSH–GSSG ratio after maturation of oocytes recovered from unstressed control (Ctrl) mice or mice exposed to 4-day RRS or 4 × 2 UPS. Each treatment was repeated 3 times and each replicate contained 35 to 40 oocytes from 2 mice. *Significant (P < .05) difference from corresponding value in control mice. #Significant (P < .05) difference from corresponding value in both control mice and RRS mice. GSX, total glutathione; GSSG, oxidized glutathione; RRS, repeated restraint stress; UPS, unpredictable stress.
Discussion
The present results showed that although oocytes obtained from mice exposed to RRS for 4 days or to 4 × 1 UPS for 4 days did not differ from oocytes recovered from unstressed control mice in both pre- and postimplantation development, the developmental potential of oocytes decreased significantly when mice were exposed to RRS for 8 or more days or to 4 × 2 UPS for 4 days. Further observations indicated that the RRS mice showed increased levels of glucocorticoids and anxiety on each day of stress from days 1 to 23 and that levels of cortisol, CRH, and anxiety-like behavior in mice exposed to 4 × 2 UPS were significantly higher than that observed in mice exposed to 4-day RRS. Assays for intraoocyte glutathione revealed more oxidative stress in oocytes following 4 × 2 UPS than following 4-day RRS. Furthermore, using the same eCG priming protocol as that used in this study, another study in this laboratory showed that one1time restraint of female mice for 48 hours during follicle growth impaired blastocyst development of oocytes significantly although restraint for 24 hours showed no effect. 22 Taken together, the results suggest that the degree that a stress damages oocytes is the product of duration × severity of the stress, and that RRS applied repeatedly over multiple days impairs oocyte developmental potential by cumulative effects.
Studies on the relationship between psychological stress and reproductive outcome in women undergoing IVF treatment showed that anxiety state scores were negatively correlated with live birth rate but positively associated with serum norepinephrine and cortisol values. 2,23 Our recent studies indicated that the impairment of oocyte competence was always associated with elevated levels of plasma glucocorticoids following psychological (restraint) stress of mice. 15,24 Injection of cortisol also decreased oocyte developmental potential in both eCG-stimulated and unstimulated mice. 15 Another study of this laboratory demonstrated that restraint stress impaired oocyte competence by increasing CRH, and the increased CRH initiated a latent apoptotic program in cumulus cells and oocytes during their intraovarian development, which was executed later during in vitro maturation (IVM) to impair oocyte competence. 25 Furthermore, in this study, control mice remained undisturbed in their home cages during the time when treated mice were stressed, suggesting that animals that received the most stress were also handled the most relative to controls. We therefore proposed that repeated homotypic stressors (RRS in this case) might have impaired oocyte developmental potential by a cumulative effect, which was produced by accumulation of the HPA products, glucocorticoids, and CRH, for example. Thus, oocytes from mice exposed to RRS for more than 8 days would be damaged greatly by more HPA products than oocytes from mice exposed to the stress for 4 days, so would be oocytes from mice exposed to 4 × 2 UPS than oocytes from mice exposed to RRS for 4 days because mice subjected to 4 × 2 UPS secreted more HPA products daily than did the mice exposed to RRS for 4 days as shown in the present study.
In the current study, oocyte developmental potential was examined after female mice that had been exposed to RRS or UPS for different days were injected with eCG to initiate the ovulatory follicle development of oocytes. Our immunohistochemistry showed that while many antral follicles underwent atresia, few preantral follicles did following 4 × 2 UPS or 4-day RRS. Furthermore, it has been reported that only the growing follicles that reach the gonadotropin -responsive and/or -dependent (antral) stage can be stimulated by gonadotrophins to become ovulatory follicles. 26,27 Thus, the present results demonstrated that chronic stress applied during follicle growth impaired the developmental potential of oocytes mainly in antral follicles. Although our previous studies showed that one-time acute restraint stress applied at the ovulatory stage of follicles impaired oocyte developmental potential, 15,16 effects of stress applied at other stages of follicular development on oocyte competence have rarely been studied. 17
In this study, levels of both glucocorticoids and CRH increased significantly following RRS or UPS. There have been reports that glucocorticoids and CRH increased simultaneously following stress. 25,28 For example, an air puff startle test elicited 10-fold increases in plasma adrenocorticotropic hormone and corticosterone in adult rats that were maternally separated during the neonatal period compared with control animals. Maternal deprivation in these animals also resulted in increased CRH concentrations in the hypothalamus, locus ceruleus, amygdale, the median eminence, portal blood, and cerebrospinal fluid compared with controls. 29 –31 Normally, if the level of glucocorticoids is increased, the level of CRH should be decreased due to a negative feedback by the glucocorticoids. Thus, the mechanism for the simultaneous elevation of glucocorticoids and CRH following stress requires explanation. Clinical studies have demonstrated hyperactivity of the HPA axis and increased levels of cortisol in patients with major depression, which has been attributed to an impairment of glucocorticoid receptor-mediated negative feedback. 32
In summary, although our previous work focused on the effects of 1-time acute restraint stress applied during the eCG priming period on the developmental potential of mouse oocytes to mimic the human IVF-associated distress taking place sometime during the FSH-stimulation period in the follicular phase, 15,16,24,33 research on the effect of chronic psychological stress during daily life on oocytes in growing follicles are limited. However, since only a small percentage of women chose to conceive by IVF, studies on the effect of chronic psychological stress during daily life on oocytes are of greater importance. Furthermore, since the UPS procedure used in this study included several stressors (hot, cold, and swimming) that cannot be considered exclusively psychological but potentially alter physiological parameters, the impact of these stressors on oocyte competence suggested that some physical stressors that a woman may undergo under certain conditions may also affect her reproductive status. In conclusion, the present results showed that (1) the degree that a stress damages oocytes is the product of duration × severity of the stress; (2) RRS impaired oocyte developmental potential through cumulative effects on growing follicles; and (3) preantral follicles were not as sensitive to stress as antral follicles were. Data are important for the healthy reproduction of both humans and animals.
Footnotes
Authors’ Note
Yan Gao and Fei Chen contributed equally to this work. Jing-He Tan and Ming-Jiu Luo are both corresponding authors.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Supported by grants from the