
Abstract
Objective:
To determine whether emodin facilitates the mesenchymal–epithelial transition (MET) of endometrial stromal cells (ESCs) as well as to explore the mechanism through which emodin favored the MET of ESCs.
Methods:
Cell viability was tested by methyl thiazolyl tetrazolium assay. Cell migration and invasion abilities were detected by transwell assays. Levels of integrin-linked kinase (ILK) and epithelial–mesenchymal transition (EMT)-related proteins were detected by Western blot.
Results:
Upregulated ILK and increased abilities of migration and invasion were confirmed in the eutopic and ectopic ESCs (EuSCs and EcSCs), especially in the EcSCs. After treated with emodin, the expression of ILK was statistically downregulated in EcSCs, resulting in the MET and decreased migration and invasion abilities of EcSCs. Additionally, silencing of the ILK gene in EcSCs also achieved the above-mentioned effects, which were strengthened by emodin. Furthermore, exogenous expression of ILK in control ESCs (CSCs) resulted in the EMT and increased abilities of migration and invasion of CSCs, which can be abrogated by emodin. Besides, exogenous expression of ILK also abrogated the effects of emodin on CSCs.
Conclusion:
Emodin inhibits the migration and invasion abilities of human ESCs by facilitating the MET through targeting ILK.
Introduction
Endometriosis is characterized by the presence of functional endometrial stromal and epithelial cells at extrauterine locations. As a common gynecological disease, endometriosis affects about 10% of reproductive-age women and results in dysmenorrhea, chronic pelvic pain, and infertility. 1,2 Although laparoscopic surgery is now considered routine for diagnosis and removal of endometriosis, 3 the recurrence rate of endometriosis is more than 20% within 2 years after the surgery. 4 Up to date, the most widely accepted hypothesis of endometriosis etiology is still the retrograde menstruation and implantation theory, which suggests that endometrial tissues can retrograde into pelvic cavity during menstruation and then develop into endometriosis. 5 However, the exact mechanism through which the retrograded endometrial tissues survive and develop into endometriosis is still unknown.
Epithelial–mesenchymal transition (EMT) is a crucial event in tumor metastasis. During EMT, epithelial cells lose cell polarity and are converted into mesenchymal cells, endowing cells with invasive and metastatic properties. Although endometriosis is a benign disease, it behaves similar to malignant tumors by infiltrating and developing outside the uterus. Additionally, emerging evidences indicate that EMT plays a significant part in the initial formation of endometriosis. 6 –8 In contrast, mesenchymal–epithelial transition (MET) can decrease the migration, invasion, and metastatic properties of the cells.
Integrin-linked kinase (ILK), a serine–threonine kinase, plays an essential role in mediating the relationship between extracellular matrix (ECM) and intracellular processes. 9 Increased expression of ILK is discovered in various cancers, inducing cell proliferation, migration, invasion, EMT as well as vascular development and tumor angiogenesis. 10 –13 Forced expression of ILK enhanced the migration and invasion abilities of cancer cells, while silencing of ILK inhibited tumor growth and invasion. 13,14
Emodin (1,3,8-trihydroxy-6-methylanthraquinone), an active ingredient of various Chinese herbs, is a natural anthraquinone present in the roots and barks of numerous plants. 15 Previous studies demonstrate that emodin possesses various biological activities, including immunosuppressive, antibacterial, anti-inflammatory, and anticancer effects. 16 –19 Recent study demonstrated that emodin repressed the EMT of cells through the ILK pathway. 20 Having excellent cytotoxicity against cancer cells, emodin shows almost no toxic effect on normal cells. 21,22 Therefore, emodin has now been proposed as a potential agent in the treatment of cancers.
In our previous study, we observed significant upregulation of ILK and increased abilities of migration and invasion in endometrial stromal cells (ESCs) of endometriosis. 23 Therefore, the inhibition of ILK expression and activity can be considered as a new target for the prevention and treatment of endometriosis. Given that emodin can repress the EMT of cancers through ILK pathway, we herein hypothesize that emodin can inhibit the migration and invasion abilities of ESCs by facilitating the MET through targeting ILK. The current study was undertaken to determine whether emodin facilitates the MET of ESCs as well as to explore the mechanism through which emodin favored the MET of ESCs.
Materials and Methods
Sample Collection and Cell Culture
Eutopic and ectopic ESCs (EuSCs and EcSCs) were cultured from 14 women with ovarian endometriosis, and control ESCs (CSCs) were cultured from 12 women without endometriosis. All of the participants were at reproductive age, had regular menstruation, and received no hormonal therapy for at least 6 months before the study. Ovarian endometriosis were visually diagnosed during the laparoscopy for ovarian cysts and then ascertained by pathological examination. The control group consisted of women who had surgery for other benign ovarian cysts. All the participants were from the Department of Obstetrics and Gynecology, Qilu Hospital of Shandong University from July 2014 to May 2015. Informed consent was obtained from all participants prior to surgery. The Institutional Review Board of Shandong University approved the study.
Methods for isolation, culture, and evaluation of the purity of the ESCs were described in our previous study. 23 The ESCs with the purity over 95% were used for the following experiments.
Western Blot Analysis
Total protein was extracted from samples for Western blot, as described previously. 23 Protein samples (30 µg/lane) were equally loaded on 8% to 12% sodium dodecyl sulfate polyacrylamide gels for electrophoresis. Primary antibodies used for immunodetection were anti-ILK, anti-E-cadherin, anti-N-cadherin, anti-vimentin, anti-keratin as well as anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH; Cell Signaling Technology, Danvers, Massachusetts). Secondary antibodies were anti-rabbit and anti-mouse immunoglobulin G peroxidase conjugate (Millipore, Massachusetts). The concentration of primary and secondary antibodies was 1:1000 and 1:5000, respectively. GAPDH was used as a loading control. The results were quantified by densitometry, using Image J software (NIH, Bethesda, Maryland). Results were representative of 3 independent experiments of each sample and were expressed as a ratio of the protein of interest and GAPDH to correct for loading of each sample.
Transwell Migration and Invasion Assays
Upon reaching confluence of 80% to 90% with emodin (20 µM or 40 µM) treatment for 48 hours, cells were digested for transwell assays as described previously. 23 For cell migration or invasion assay, 4 × 104 cells were seeded in each well and incubated for 24 hours. Pictures of stained cells were taken by the Olympus IX51 inverted microscope (Olympus Corporation, Tokyo, Japan). Cells were counted in 5 random fields (200×) of each chamber. The average cell numbers of 3 duplicate wells for each specimen were used for statistical analysis.
Methyl Thiazolyl Tetrazolium Cell Viability Assay
Cell viability was measured using the methyl thiazolyl tetrazolium (MTT) assay as reported previously. 23 Briefly, ESCs (3 × 103 cells/well) were seeded into a 96-well, flat-bottomed microplate and incubated overnight. Then the cells were treated with increasing concentrations of emodin (Sigma, St Louis, Missouri) for 24, 48, and 72 hours, respectively. Thereafter, 10 μL MTT (5 g/L) solution was added to each well and then incubated for additional 4 hours before dimethyl sulfoxide (100 μL/well) treated. The absorbance at 490 nm was used to evaluate the ability of cell proliferation. Data were calculated as follows: (absorbance of test sample/absorbance of control).
Silencing of the ILK Gene in EcSCs
We silenced the ILK in EcSCs by transfecting EcSCs with small-interfering RNA sequences targeting human ILK (siRNA-ILK) because EcSCs showed the highest level of ILK. The siRNA-ILK sequences were designed by GenePharma Company (Shanghai, China). The EcSCs were seeded in 6-well plates without antibiotics treated overnight and then transfected with blank sequence or siRNA-ILK (50 nmol/L) using lipofectamine 2000 (Invitrogen Life Technologies, Carlsbad, California) when cell confluence gets 50% to 60%. After 48 hours of transfection, cells were treated with emodin (40 µmol/L) or digested for transwell assays or Western blot. The siRNA-ILK sequences are listed in Table 1.
Sequences of siRNA-ILK.
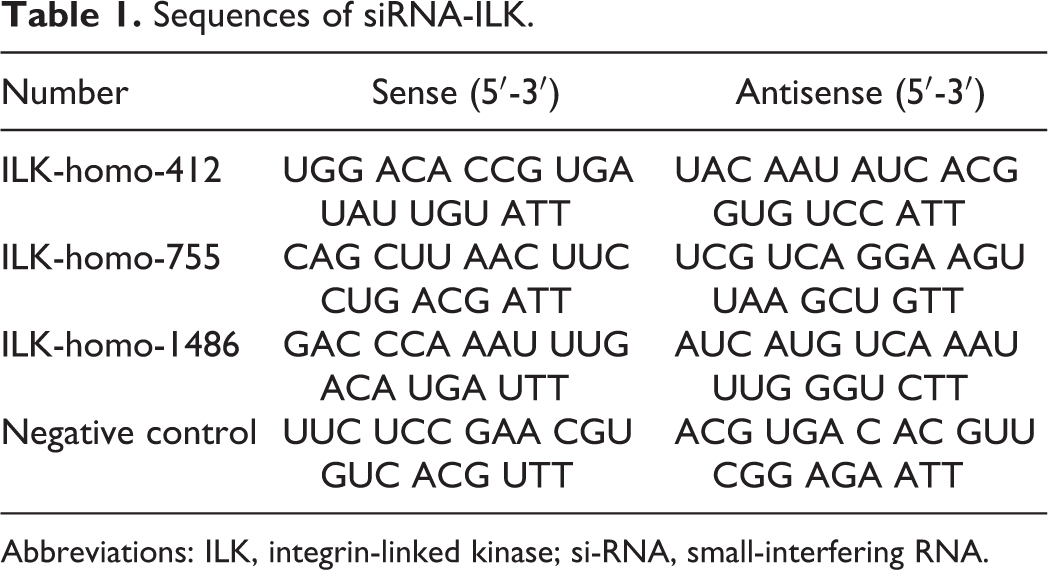
Abbreviations: ILK, integrin-linked kinase; si-RNA, small-interfering RNA.
Transient Transfection of the ILK Gene in CSCs
Integrin-linked kinase was exogenously expressed in CSCs by transfecting CSCs with ILK overexpression vector (pEGFP-C1-ILK) because CSCs showed the lowest level of ILK. The control vector (pEGFP-C1) and pEGFP-C1-ILK were also designed by GenePharma Company (Shanghai, China). The CSCs were seeded into 6-well plates without antibiotics treated overnight and then transfected with pEGFP-C1 or pEGFP-C1-ILK (3 μg/well) using lipofectamine 2000 (3 μg/well) when cell confluence gets 70% to 80%. After 48 hours of transfection, cells were treated with emodin (40 µmol/L) or digested for transwell assays or Western blot.
Statistical Analysis
GraphPad Prism Version 5.01 (GraphPad Software, San Diego, California) was used for statistical analysis. The data are shown as mean ± standard error of the mean. Student t test and 1-way analysis of variance were, respectively, conducted to analyze the differences between groups and among groups. P value < .05 was considered statistically significant.
Results
Expression of ILK and Abilities of Migration and Invasion in CSCs, EuSCs, and EcSCs
To verify the upregulated expression of ILK in ESCs of ovarian endometriosis, western blot was conducted to detect the expression of ILK in CSCs, EuSCs, and EcSCs. The expression of ILK was higher in EuSCs and EcSCs and highest in EcSCs compared to that in CSCs (Figure 1A and B). Apart from that, EuSCs and EcSCs were larger and more spindle like than CSCs in morphology (Figure 1C). Then, we detected the migration and invasion abilities of ESCs and discovered that EuSCs and EcSCs showed much stronger abilities of migration and invasion (Figure 1C-E).
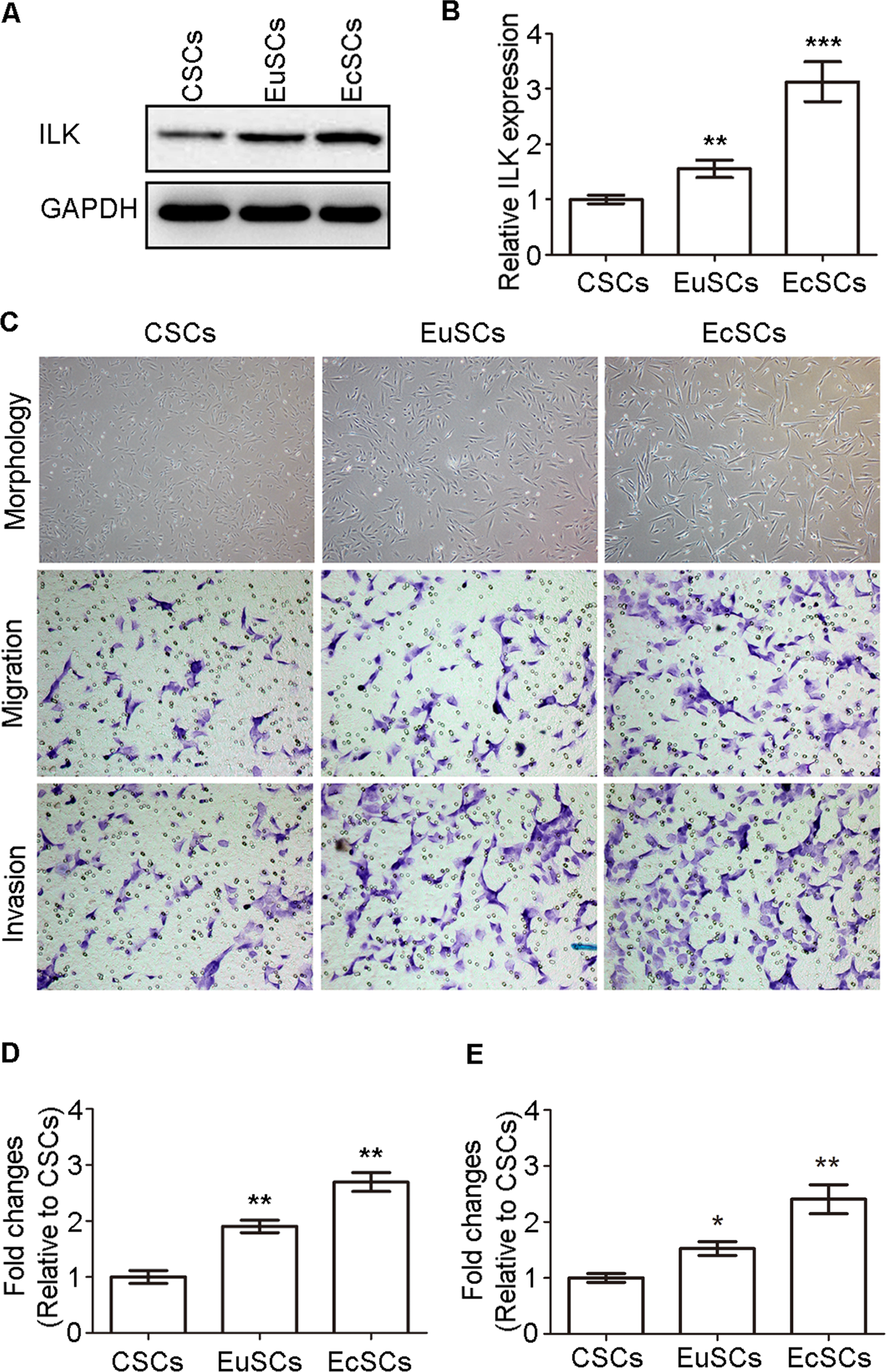
Expression of ILK and abilities of migration and invasion in CSCs, EuSCs, and EcSCs. A, Representative western blots showing the expression of ILK in CSCs, EuSCs, and EcSCs. B, Quantitative analysis of ILK expression (**P < .005 and ***P < .001). The y-axis represents the proportion of protein relative to CSCs. C, Representative morphology of CSCs, EuSCs, and EcSCs (original magnification, ×200); representative transwell migration and invasion assay of CSCs, EuSCs, and EcSCs (original magnification, 200×). D, Quantification of migration abilities of CSCs, EuSCs, and EcSCs (**P < .005). E, Quantification of invasion abilities of CSCs, EuSCs, and EcSCs (*P < .05 and **P < .005). ILK indicates integrin-linked kinase; CSCs, control stromal cells; EuSCs, eutopic stromal cells; EcSCs, ectopic stromal cells.
Emodin Inhibited the Proliferation, Migration, and Invasion of EuSCs and EcSCs
Emodin inhibited the proliferation of EuSCs in a dose- and time-dependent manner, with significant inhibition at 40 μmol/L (Figure 2A). Similar results were also found in EcSCs (Figure 2B). Given that EcSCs have the strongest abilities of migration and invasion, we tested the effect of emodin on the migration and invasion abilities of EcSCs. As expected, the migration and invasion abilities of EcSCs were significantly decreased after treatment with emodin for 48 hours (Figure 2C-E). Then, we detected the effect of emodin on the morphology of EcSCs and discovered that EcSCs were smaller and less spindle like after the treatment (Figure 2C).
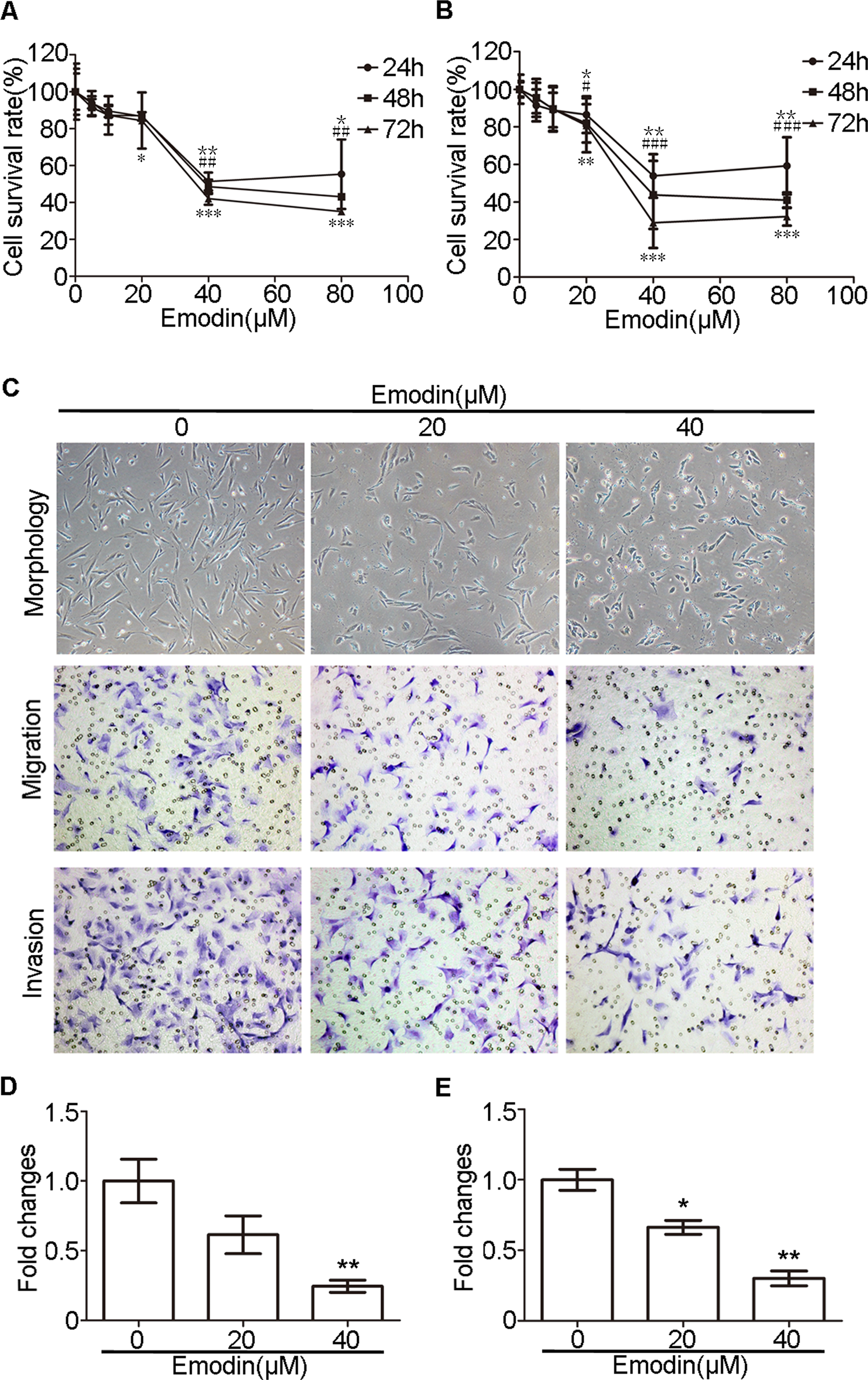
Emodin inhibited the proliferation, migration, and invasion of ESCs of endometriosis. A, Emodin repressed the proliferation of EuSCs in a dose- and time-dependent manner (24 h: *P < .05, **P < .005; 48 h: ## P < .005; 72 h: *P < .05, ***P < .001). B, Emodin repressed the proliferation of EcSCs in a dose- and time-dependent manner (24 h: *P < .05, **P < .005; 48 h: # P < .05, ### P < .001; 72 h: **P < .005, ***P < .001). C, Representative morphology and transwell migration and invasion assay of EcSCs after treated with emodin (original magnification, 200×). D, Quantification of migration abilities of EcSCs with and without the treatment with emodin (**P < .005). E, Quantification of invasion abilities of EcSCs with and without the treatment with emodin (*P < .05 and **P < .005). CSCs indicates control stromal cells; EuSCs, eutopic stromal cells; EcSCs, ectopic stromal cells; ESCs, endometrial stromal cells.
Emodin Decreased the Expression of ILK in EcSCs and Facilitated the MET of EcSCs
Dose-dependent decrease in ILK expression was observed in EcSCs after treatment with increasing concentrations emodin for 48 hours (Figure 3A and B). Moreover, emodin (40 μmol/L) treatment effectively decreased ILK expression in a time-dependent manner in EcSCs (Figure 3C and D). To examine whether emodin decreased the migration and invasion abilities by facilitating the MET of EcSCs, we tested the expression of EMT hallmarks, E-cadherin, keratin, N-cadherin, and vimentin by Western blot. As depicted in Figure 3, emodin upregulated the levels of E-cadherin and keratin while downregulated the levels of N-cadherin and vimentin in EcSCs in a dose- and time-dependent manner. Thus, emodin enhanced the MET of EcSCs in a dose- and time-dependent manner.
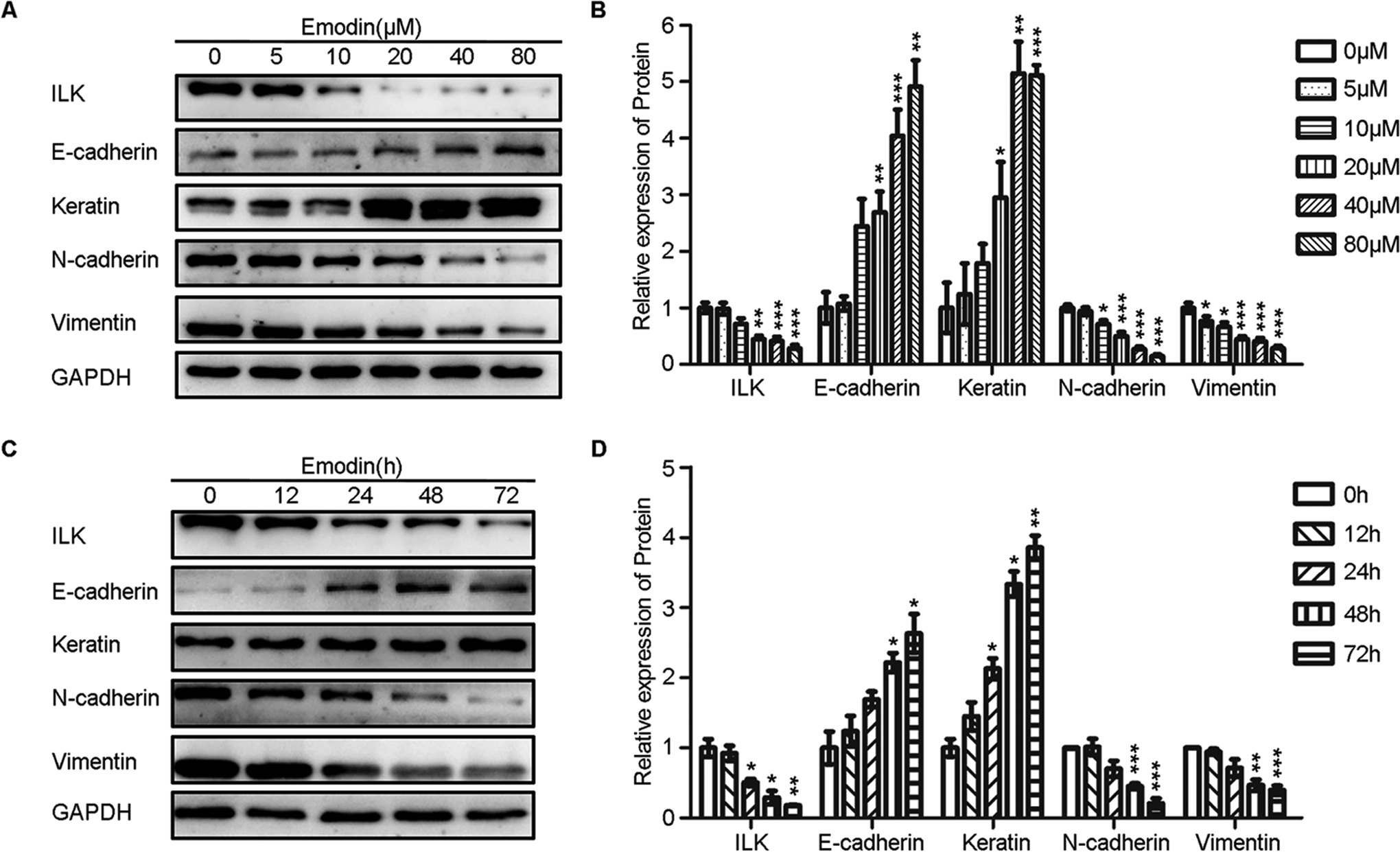
Emodin downregulated the ILK expression and facilitated the MET of EcSCs in a dose- and time-dependent manner. A, Representative western blots showing the expression of ILK and MET markers in EcSCs after treated with increasing concentrations of emodin. B, Quantitative analysis of ILK and MET markers in EcSCs after treated with increasing concentrations of emodin. (*P < .05, **P < .005, and ***P < .001). C, Representative western blots showing the expression of ILK and MET markers in EcSCs after treated with emodin for increasing time. D, Quantitative analysis of ILK and MET markers in EcSCs after treated with emodin for increasing time. (*P < .05, **P < .005, and ***P < .001). ILK indicates integrin-linked kinase; MET, mesenchymal–epithelial transition; EcSCs, ectopic stromal cells.
Emodin Enhanced the MET of ESCs by Targeting ILK
To verify whether emodin facilitated the MET of ESCs by targeting ILK, we silenced the ILK in EcSCs by transfecting EcSCs with siRNA-ILK and overexpressed the ILK in CSCs by transfecting CSCs with pEGFP-C1-ILK. The transfection of ILK-homo-755 had the strongest effect on the silencing of ILK (Supplementary Figure). So ILK-homo-755 was used in the remaining tests. After transfection of siRNA-ILK, the levels of E-cadherin and keratin were increased while the levels of N-cadherin and vimentin were decreased in EcSCs, which can be strengthened by emodin (40 μmol/L; Figure 4A and B). In addition, the migration and invasion abilities of EcSCs were decreased by the transfection of siRNA-ILK and the effect was also strengthened by emodin (40 μmol/L; Figure 4C and E). On the contrary, the levels of ILK, N-cadherin, and vimentin were increased, while the levels of E-cadherin and keratin were decreased after transfecting CSCs with pEGFP-C1-ILK, which can be abrogated by emodin (40 μmol/L; Figure 5A and B). Simultaneously, the migration and invasion abilities of CSCs were increased by the transfection of pEGFP-C1-ILK, which can also be abrogated by emodin (40 μmol/L; Figure 5C-E). Besides, exogenous expression of ILK also reversed the effects of emodin on CSCs (Figure 5A-E). Therefore, emodin decreased the migration and invasion abilities of ESCs by facilitating MET by targeting ILK.
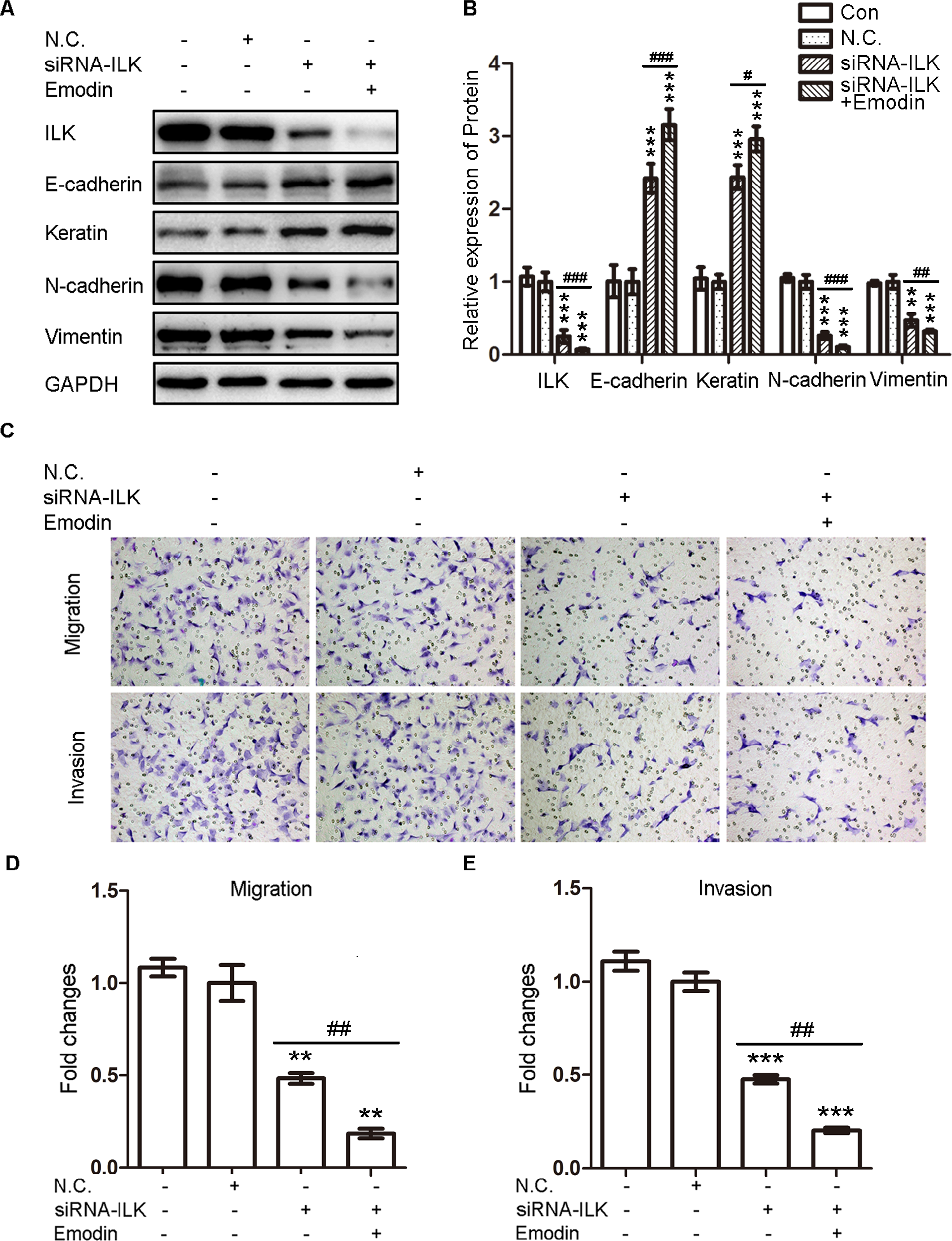
Silencing of ILK in EcSCs induced the MET and decreased migration and invasion abilities of EcSCs. A, Representative Western blots showing the expression of ILK and MET markers in EcSCs after transfection siRNA-ILK with or without the treatment with emodin. B, Quantitative analysis of ILK and MET markers in EcSCs after transfection siRNA-ILK (EcSCs transfected with negative control (N.C.) was used as a control: **P < .005 and ***P < .001) with or without the treatment with emodin (# P < .05, ## P < .005, and ### P < .001). C, Representative transwell migration and invasion assay of EcSCs after transfection siRNA-ILK with or without the treatment with emodin (original magnification, 200×). D, Quantification of migration abilities of EcSCs after transfection siRNA-ILK (EcSCs transfected with N.C. was used as a control: **P < .005) with or without the treatment with emodin (## P < .005). E, Quantification of invasion abilities of EcSCs after transfection siRNA-ILK (EcSCs transfected with N.C. was used as a control: ***P < .001) with or without the treatment with emodin (## P < .005). ILK indicates integrin-linked kinase; MET, mesenchymal–epithelial transition; EcSCs, ectopic stromal cells; siRNA-ILK, small-interfering RNA sequences targeting human ILK.
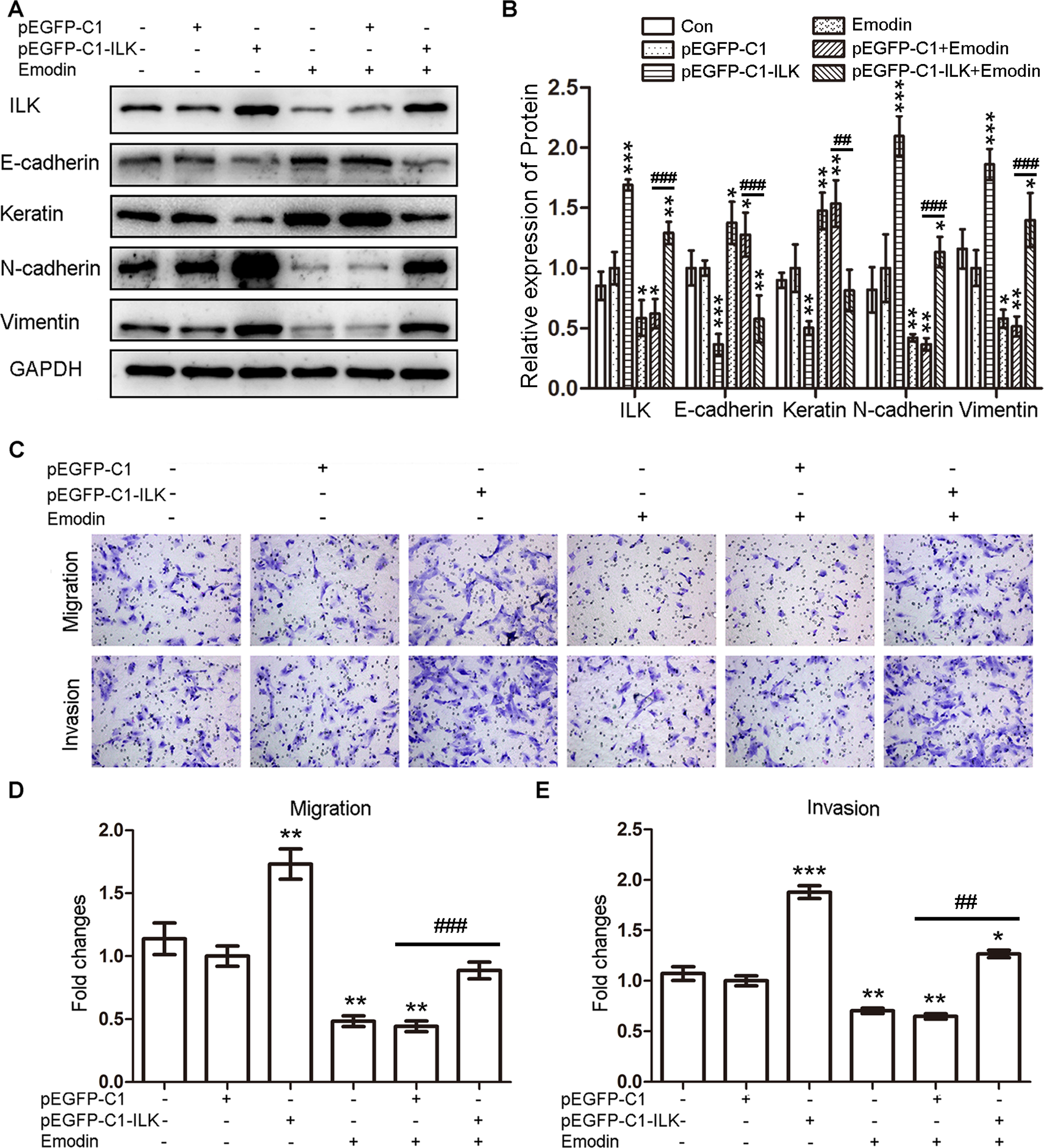
Exogenous expression of ILK in CSCs induced the EMT and increased migration and invasion abilities of CSCs. A, Representative Western blots showing the expression of ILK and MET markers in CSCs after transfection pEGFP-C1-ILK with or without the treatment with emodin. B, Quantitative analysis of ILK and MET markers in CSCs after transfection pEGFP-C1-ILK (CSCs transfected with pEGFP-C1 was used as a control: *P < .05, **P < .005, and ***P < .001) with or without the treatment with emodin (## P < .005 and ### P < .001). C, Representative transwell migration and invasion assay of CSCs after transfection pEGFP-C1-ILK with or without the treatment with emodin (original magnification, 200×). D, Quantification of migration abilities of CSCs after transfection pEGFP-C1-ILK (CSCs transfected with pEGFP-C1 was used as a control: **P < .005) with or without the treatment with emodin (### P < .001). E, Quantification of invasion abilities of CSCs after transfection pEGFP-C1-ILK (CSCs transfected with pEGFP-C1 was used as a control: *P < .05, **P < .005, and ***P < .001) with or without the treatment with emodin (## P < .005). ILK indicates integrin-linked kinase; CSCs, control stromal cells; EMT, epithelial–mesenchymal transition; MET, mesenchymal–epithelial transition.
Discussion
As an important kinase, ILK can directly phosphorylate its downstream targets to mediate cell–ECM and cell–cell interaction. Integrin-linked kinase was previously reported to facilitate the migration and invasion of many cancers. 24 –26 Then, subsequent publications demonstrated that ILK could enhance the migration and invasion abilities of cancers by promoting the EMT program. 13,27 Consistently, targeting ILK decreased the invasion and metastasis abilities of renal carcinoma cells via repressing the EMT process. 28 Consistent with our previous study, 23 the present study discovered upregulated expression of ILK as well as increased migration and invasion abilities in EuSCs and EcSCs. Differently, the expression of ILK was highest in EcSCs in this study, while no statistically significant difference existed between EuSCs and EcSCs in the previous study. The reason for the difference may be that all the paired EuSCs and EcSCs in this study were from the same patient while the previous paired EuSCs and EcSCs were not all from the same patient. Considering that ILK facilitated the migration and invasion abilities of ESCs, the current result may be more precise because EcSCs exhibited the highest abilities of migration and invasion. Given that EMT plays crucial roles in various cancers and endometriosis, targeting ILK may be a promising approach in treatment regimes for cancers and endometriosis.
Emodin possesses various biological activities and can inhibit the migration and invasion abilities of several kinds of cancer cells. 29 –31 Considering the essential role in the invasion–metastasis cascade, MET may account for the inhibition of migration and invasion abilities of cancer cells. In 2011, emodin was first reported to inhibit the EMT of colorectal cancer cells by increasing the expression of E-cadherin and decreasing the expression of vimentin. 32 Then, Way et al demonstrated that emodin repressed the Twist1-induced EMT in head and neck squamous cell carcinoma cells. 33 The present study was in accordance with these studies and found that emodin can inhibit the migration and invasion abilities of ESCs by facilitating the MET of ESCs. The concentration of emodin used here, with significantly inhibition of ESCs proliferation, was consistent or even lower than those previously reported in some cancers. 31,34,35 Although the repress EMT effect of emodin was discovered, the underlying molecular mechanism remains to be elucidated.
Possessing various biological activities, emodin was previously reported to function as AMPK activator, interleukin 6 inhibitor, casein kinase II inhibitor, phosphatase of regenerating liver 3 inhibitor, and 11β-hydroxysteroid dehydrogenase type 1 inhibitor. 36 –40 Besides, Tang et al discovered that emodin inhibited the expression of ILK. 34 Likewise, Chen et al demonstrated that emodin ameliorated the EMT of high glucose-induced podocyte by inhibiting the ILK expression. 20 Our current study also found that emodin decreased the expression of ILK, resulting in the repressed abilities of migration and invasion of ESCs via enhancing the MET of ESCs. The effect of emodin tended to be more effective in a dose-dependent manner than in a time-dependent manner. High-dose emodin was more effective than long time emodin treatment.
Taken together, emodin may be a potential new therapeutic option for ovarian endometriosis in the future. Although hormonal therapy is often effective and represents important therapeutic tool in the treatment of endometriosis, nonhormonal approaches are needed due to the numerous side effects of hormonal therapy. 41 The present study provided a new insight into the nonhormonal treatment of endometriosis. However, the current study is limited by the small sample size. In addition, the mechanisms of emodin targeting ILK and ILK facilitating EMT remain unclear. Studies with a larger sample size are necessary to confirm the results, and further investigations are required to explore the exact molecular mechanism. Additionally, in vivo studies are in needed to investigate the therapeutic effect of emodin on ovarian endometriosis.
In conclusion, our study verified that emodin could repress the proliferation, migration, and invasion abilities of ESCs, and the latter 2 were due to the enhancement of the MET of ESCs by emodin. Furthermore, emodin inhibited the migration and invasion abilities of ESCs by facilitating the MET of ESCs through targeting ILK. These findings might provide a potential new therapeutic option for ovarian endometriosis.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by grants from the National Natural Science Foundation of China (NSFC) (Grant number 81370696) and the Science and Technology Development Planning of Shandong (Grant number 2013GGE27031).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.