
Abstract
Rationale:
Development of cervical squamous carcinoma (CXCA) is accompanied by changes in estrogen receptors (ERs, ERα and ERβ) and ezrin expression; however, reports have been conflicting. Using histologically documented staging of cervical biopsies, we determined ezrin and ER relationships during CXCA development.
Methods:
Immunoreactive (ir) ezrin, ir-ERα, and ir-ERβ were studied in normal epithelium, carcinoma in situ/cervical intraepithelial neoplasia (CIN) 1 to 3, and local invasion or metastatic CXCA. Results were compared using H scoring. Cultures of Caski metastatic CXCA cells were treated with estradiol and/or tamoxifen and studied for ER-driven ir-ezrin and the morphologic response.
Results:
Koilocytosis was present and indicated viral presence. The ezrin H score increased from CIN1 to CIN3, reaching significant differences from normal by CIN3 (P = .004) and 2× normal in metastatic CXCA. Estrogen receptor α and ERβ H scores fell, reaching significance by CIN3 (ERα, P = .0001; ERβ, P = .024). During estradiol treatment, ezrin in Caski cells increased and localized to the periphery, in ruffles and processes. The selective ER modulator tamoxifen blocked the estradiol-induced changes.
Conclusions:
During cervical carcinogenesis, the usual relationship between estrogen and ezrin induction is abridged. This is consistent with the effects of human papilloma virus viral proteins such as E6 and E7 that upregulate SIX1, a protein that induces ezrin. Cervical carcinogenesis is progressive but arrests at the preinvasive stage for varying lengths of time. These studies suggest that changes in ezrin may be associated with the development of the invasive phenotype and penetration of the basement membrane. They also raise the possibility that inhibiting ezrin expression could be a target for the prevention of invasive CXCA.
Introduction
An estimated 13 000 cases of invasive epidermoid carcinoma of the cervix, cervical squamous carcinoma (CXCA), were diagnosed, and an estimated 41 000 deaths from cervical cancer were expected in 2015. 1,2 Human papilloma virus (HPV), especially HPV16, is the main cause of this lesion. 3 Despite the availability of an anti-HPV vaccine, the likelihood of eradication of this disease remains a distant hope; therefore, study of the mechanisms underlying the pathogenesis of CXCA remains a high priority.
Ezrin is a member of the ezrin–radixin–moesin protein family. It drives cellular specialization and functions as a signal transducer. Ezrin is involved in the regulation of phenotypic changes in cellular architecture. Moreover, it facilitated cellular movement and motility, recognizes internal and external signals, and controls cellular growth. 4 -6 These functions are essential to maintain homeostasis in normal cells and are highly involved in the process of tumorigenesis, invasion, and distant metastasis. 6 -8 Abnormal ezrin forms have been found in cancers and nonmalignant conditions 9,10 (A. Fadiel and F. Naftolin, personal communication). We and others have shown high expression of ezrin in CXCA and other cancers, especially metastatic lesions. 11
Although estrogen receptors (ERs) have been shown to be present in the normal cervical epithelium and CXCA, reports have indicated low expression of ERs in developing and invasive CXCA. However, preclinical trials using the selective estrogen receptor modulator (SERM) raloxifene showed inhibition of mouse CXCA supporting retention of ER activity. 12 -14
Since estrogen is a well-known regulator of maturation (keratinization) of the cervical epithelium and ezrin is important in normal and malignant cell polarization, adhesion, and motility, we sought to determine the changes in these key proteins during the progress of CXCA. The present report correlates the expression of ezrin and ERs with the progression of cervical carcinogenesis and supports a causative role for ezrin and ER expression in the development of CXCA.
Materials and Methods
Under an institutional review board–approved protocol, archival cervical tissue and sections were obtained from Soonchunhyang Medical Center Hospital. A total of 57 specimens containing the squamocolumnar junction (SCJ; with the exception of the metastatic lesions were studied. Of these, 10 specimens were classified into each of the following 5 groups: normal, cervix intraepithelial neoplasia (CIN) I to III, and metastatic CXCA. Another 7 specimens were classified as invasive CXCA. The stage of cancer was determined by a single pathologist according to the International Federation of Gynecology and Obstetrics system. Specimens with CIN were labeled according to the disposition of normal versus abnormal cells (CIN1 = cells in lower one-third of epithelial height abnormal, CIN2 = cells in lower two-thirds of epithelial height abnormal, CIN3 = cells in more than two-thirds of epithelial height abnormal). All specimens were obtained by cold conization or hysterectomy, fixed in 10% formalin and cut as 5 microsections. Caski cells were purchased from American Type Culture Collection (Rockville, Maryland).
Tissue Processing and Immunohistochemistry for Ezrin
Immunohistochemical staining was conducted on paraffin sections (10 specimens in each of the normal, CIN I-III, and metastatic CXCA groups and 7 invasive CXCA specimens). Each section contained a well-conserved SCJ and a full thickness layer of epithelial plus stromal tissue. Sections were collected on poly-
Immunofluorescent Staining of Ezrin in the Caski Cell Line
A modification was used in our estrogen-free method for tissue culture. 15 Prior to study, the cells were maintained in minimal essential medium (MEM) supplemented with penicillin, streptomycin, and 10% fetal calf serum (GIBCO BRL, Gaithersburg, Maryland). Cells were cultured in 5% CO2 in humidified air, and the medium was changed twice a week. After flask incubation to subconfluence (60%-70%) in MEM containing 10% fetal bovine serum, the medium was changed to phenol red-free, serum-free Opti-MEM 1× for more than 72 hours during which time the cells grew to 100% confluence without evidence of cell death (apoptotic bodies, floating cells, etc). Following 24 hours of preincubation in Opti-MEM, 1×, the cells were exposed to 0.05% trypsin (supplemented with 5.3 nM EDTA–4Na, mycoplasma-free; GIBCO BRL, Gaithersburg, Maryland) for 5 minutes and lightly shaken to loosen them from the flask bottom. After cells were detached, they were washed with fresh Opti-MEM 1× to stop the action of trypsin. The cells were transferred to plastic Falcon tubes (Fisher Scientific, Chino, California) in Opti-MEM 1× and gently centrifuged. The cell pellet was resuspended in culture medium and transferred to 4-chambered plates for the estrogen and tamoxifen treatment to establish an estrogen-controlled environment, and the cells were then incubated for an additional 24 hours at 37°C with serum-free Opti-MEM 1× with added 0.001% ethanol (timed-vehicle control), 7 estrogen (10−8 M), or estrogen plus tamoxifen (10−6 M) or tamoxifen (10−6 M). The cells were fixed with Cytofix solution, washed with 1× PBS 3 times, and then blocked with 1% bovine serum albumin. Subsequently, the cells were incubated with anti-ezrin mouse antibodies (Neomarkers, lot 661 × 503) and diluted (1:200) overnight at 4°C. Next, the cells were incubated with secondary antibodies (Alexa Fluor antimouse IgG; 1:600) for 30 minutes at room temperature and observed by fluorescence microscopy.
Scoring and Statistical Analysis
Modified “H testing” was used for semiquantitative evaluation of proteins in the stained sections. Two blinded observers using ×40 objectives (final magnification ×400) evaluated the intensity of staining—0 = no staining, 1+ = weak but detectable staining, 2+ = moderate or distinct staining, and 3+ = strong staining. H scores were calculated using the following equation: H = ΣPi (i + 1), where i is the intensity of staining with a value from 0 to 3 and Pi is the percentage of stained cells, varying from 0% to 100%. In each slide, 5 different random areas and 100 cells in each area were evaluated and the average score was calculated. Comparisons of the means were performed by 1-way analysis of variance. Statistical significance was defined as P < .05.
Results
Microscopical examination confirmed that all tissues were staged correctly and that the sections were satisfactory for immunostaining and H scoring (see Figure 1 and Discussion section). Granular immunostaining in normal endocervical glands at the SCJ was used as a positive control, and ezrin, ERα, and ERβ were expressed in all stages. Koilocytosis was present in all sections of the SCJ, that is, all tissues had viral signatures (data not shown).
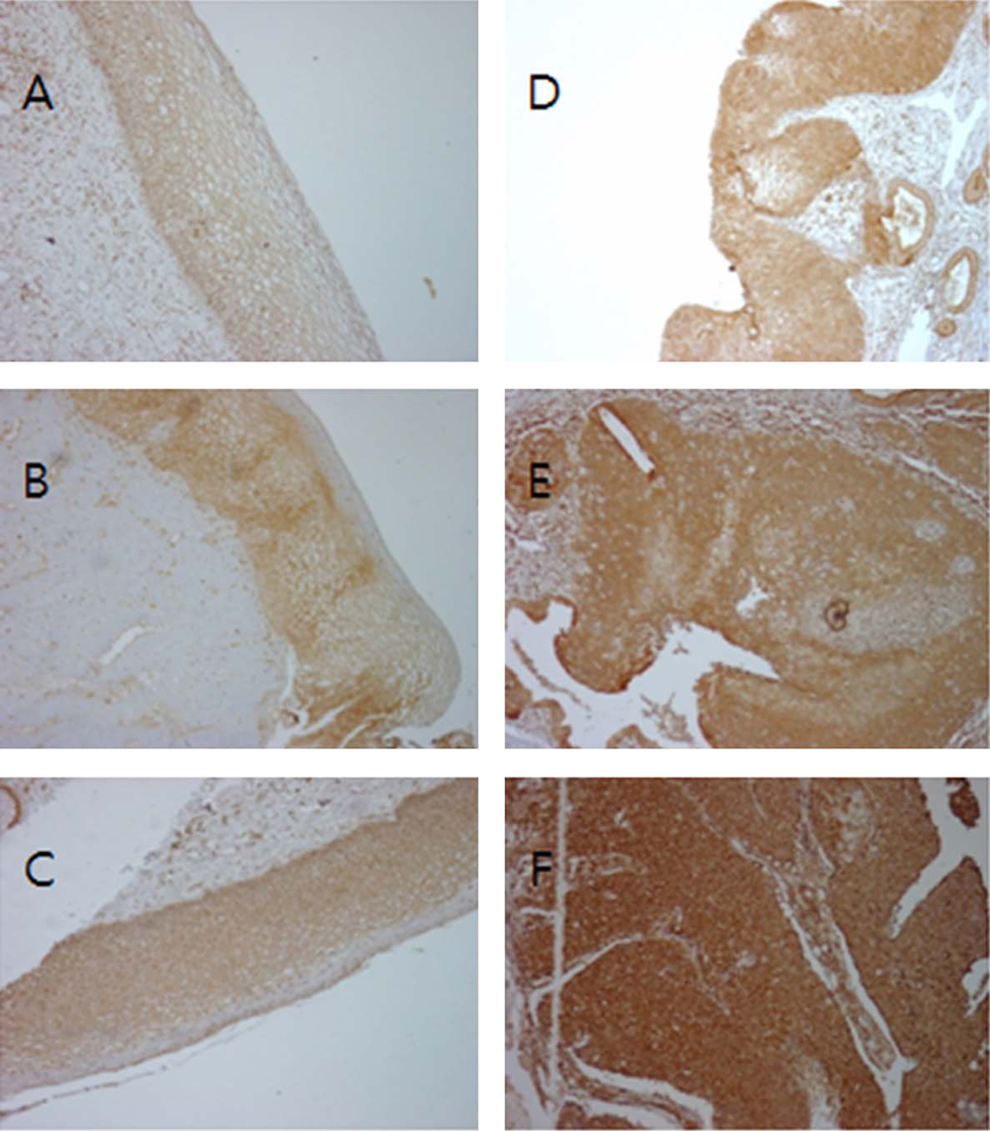
Immunoreactive-ezrin staining (brown) in the cervix (×20): (A) normal cervix; (B) cervix intraepithelial neoplasia (CIN) I; (C) CIN II; (D) CIN III; (E) invasive cancer; (F) metastatic cancer. The density of ezrin staining increased with disease progression.
Ezrin Expression in Cervical Tissue and Caski Cells
Ezrin immunoreactivity was positive and observed to increase in the following order of stages of transformation: normal < cervix < squamous epithelium < CIN 1 < 2 < 3 < invasive < metastatic squamous cell carcinoma (Figure 1).
Normal epithelial cells showed a light staining pattern at the basal or parabasal cell borders. In CIN1, the ezrin scores were slightly higher and staining appeared more intense around the luminal boarder of the cells. Compared to normal tissues, ezrin expression showed statistically significant increases by the attainment of CIN3. The difference in H score between ezrin staining in CIN2 and CIN3 nearly doubled between the stages before CIN2 or after CIN3 but this difference did not reach statistical significance (P = .223; Figure 1). The mean H scores from CIN3 invasion was 2 or greater with an up-going trend. High levels of ezrin expression were noted in cancer tissue compared with normal and low-grade CIN samples (P < .05).
Estrogen Receptors α and β Expression in Cervical Tissue
In normal cervical epithelium, ER was mainly observed in basal cells of the squamous epithelium and endocervical glands (data not shown). The strongest staining for ERα and β was present in the normal epithelium; this continuously decreased as cancer progressed. Overall, immunostaining for both ERs showed a sustained and almost complete decline during the transformation to invasion and metastases (P < .05; Figure 2B and C).
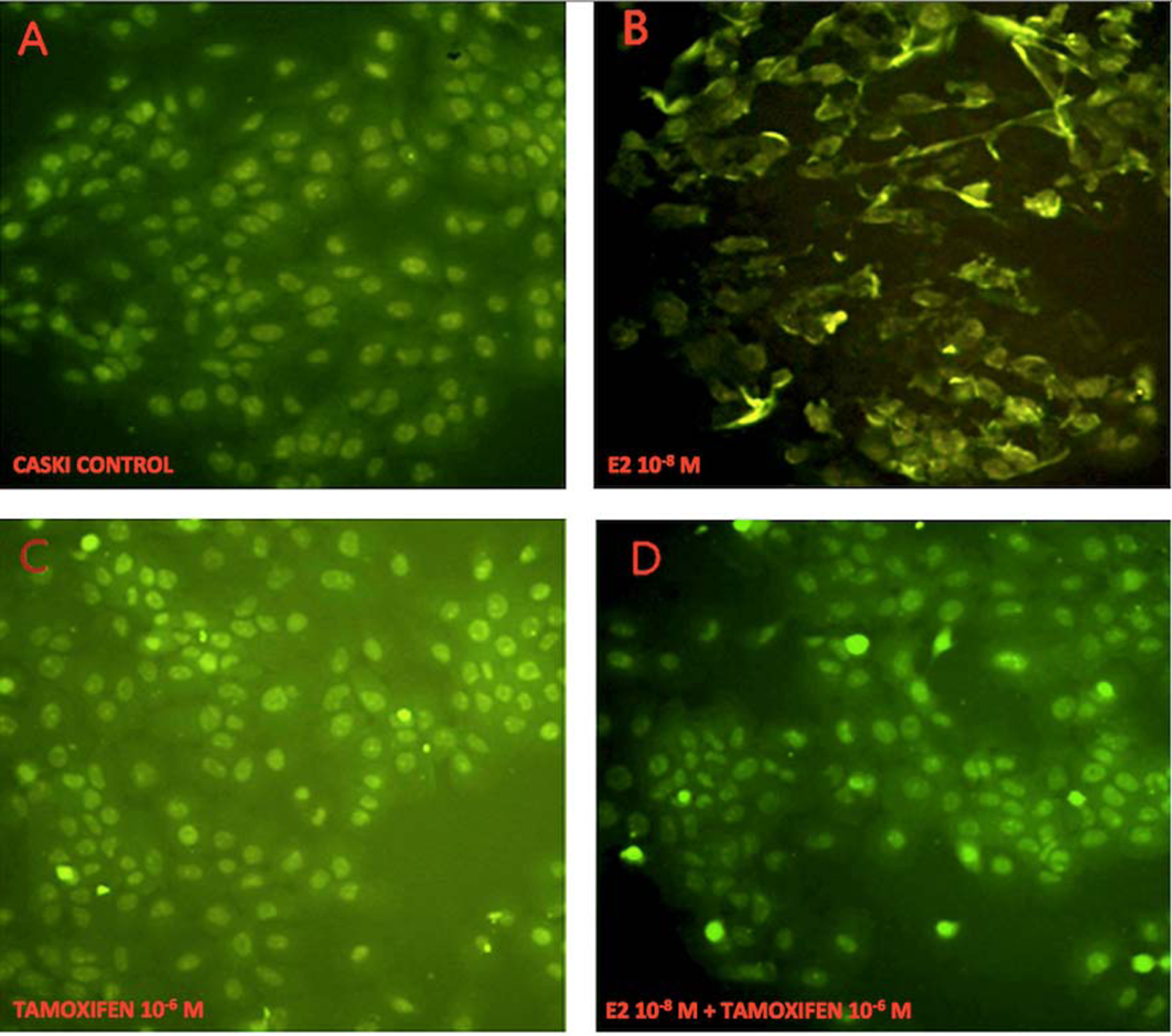
Ezrin expression in Caski cells with immunohistochemistry (×20). A, Control (cultured Caski cells): ezrin was observed in the perinuclear and cytoplasmic areas, and cells were round or ovoid without spindles. B, Following 24-hour treatment with estrogen (E2, 10−8 M), cells formed pseudopodia and ruffled cell membranes, and ezrin translocated to the cell membrane. C, Following 24-hour treatment with tamoxifen (10−6 M), cells stained more heavily than controls but did not form processes. D, Following 24-hour treatment with tamoxifen (10−6 M) and E2 (10−8 M), cells were round without spindles, and morphological changes appeared with pseudopodia.
Estrogen Treatment of Caski Cells
In estrogen-free cultures, immunoreactive (ir) ezrin was weakly detected in the cytoplasmic and perinuclear areas of the flat, ovoid-shaped Caski CXCA cells. Following 24 hours of estradiol treatment, ir-ezrin appeared to be more dense and peripheral. Significant phenotypic changes were observed such as spindles, protruding processes, nuclear pleomorphism, and irregular cellular shapes. Tamoxifen treatment had little apparent effect, with some translocation to the cell periphery. The cells appeared smaller and more rounded than those that were untreated. Estradiol plus tamoxifen-treated Caski cells were not different from untreated cells (Figure 3).
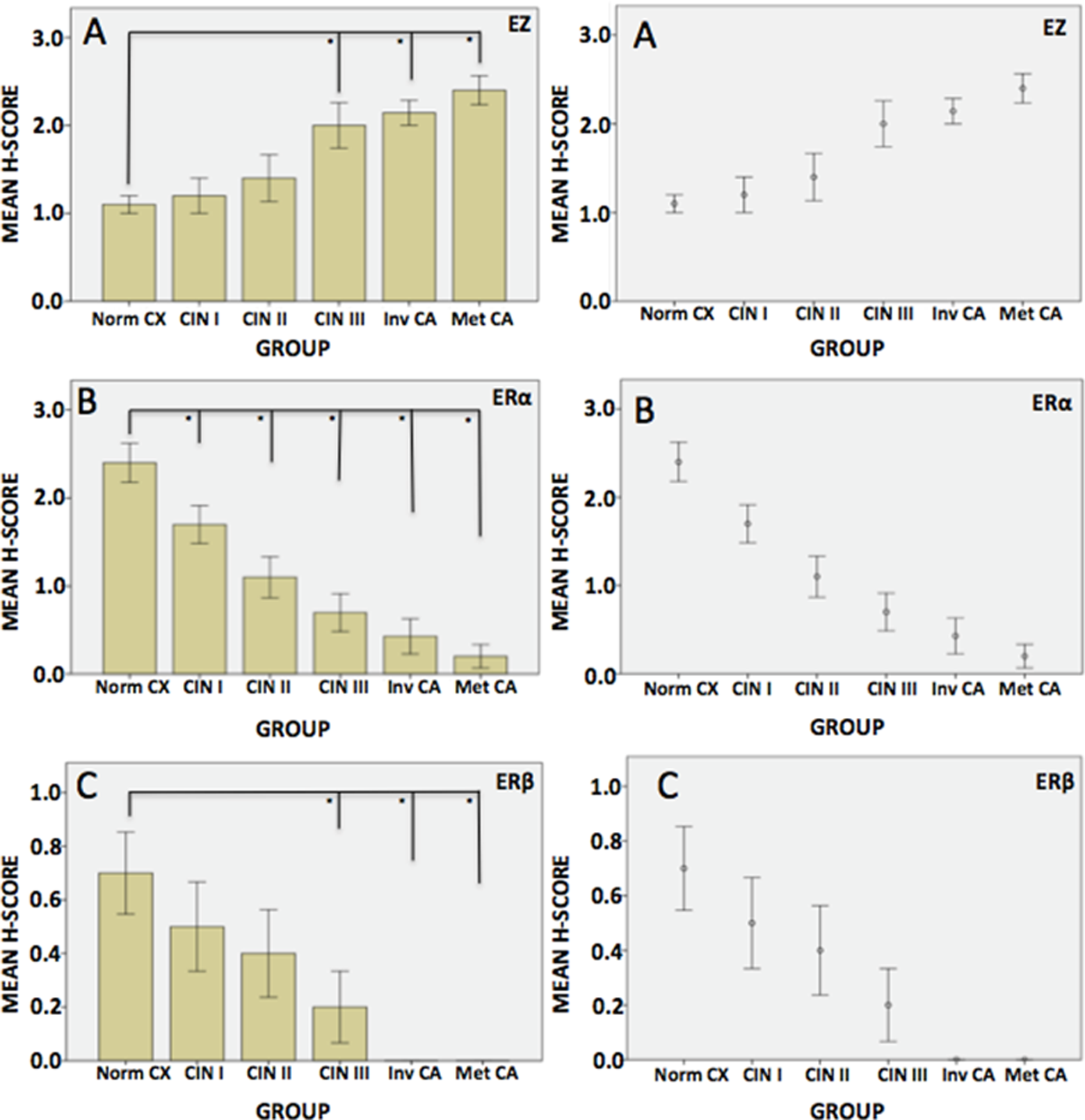
A, Ezrin mean H score values in cervical tissue. *P < .05 versus control. B, Estrogen receptor α (ERα) mean H score values in cervical tissue. *P < .05 versus control. C, Estrogen receptor β (ERβ) mean H score values in cervical tissue. *P < .05 versus control.
Discussion
In this project, we delineated the expression of ERs and ezrin during the development of cervical cancer. To categorize the study sections, we followed the pathological diagnosis of normal, CIN 1 to 3, and invasive CXCA. Although this classification has been replaced for the purpose of interpretation of Papanicolaou smears, 16 it served quite well for the present project in which we directly visualized the morphohistology being tested. The milestones chosen for this analysis included CIN1 (malignant cells limited to the basal one-third of the section), CIN2 (malignant cells limited to the basal two-thirds of the section), and CIN3 (malignant cells occupying entire thickness of the section). There was no evidence of penetration of the basement membrane in any of the “CIN” sections. We carefully reviewed each tissue section to ascertain which stage of preinvasive cancer was being examined.
Expressions of ezrin and ER were studied in this project because of our previous demonstration of increased ezrin in many cancers, including estrogen-sensitive female genital cancers and because estrogen/ER induces ezrin. 15,17 Animal studies have shown the requirement for ER in HPV E6/7-induced CXCA. 18 In support of the role of ER in CXCA, Spurgeon and others have shown that the SERM raloxifene retards the growth of viral E6/E7-induced CXCA in mice. 13,14 The present project documented the decline in ER expression during the development of CXCA. To confirm the functional activity of ER in CXCA, we showed that treating Caski cells with estrogen-induced ezrin expression and development of the invasive phenotype, that is, ER in CXCA, are functional. However, the decline in ER may explain the disappearance of ectocervical glycogen, that is the hallmark of cervical epithelial transformation.
Despite the decrease in ER expression, the expression of ezrin increased during carcinogenesis; in CXCA, the relationship between ezrin and estrogen is dichotomous. SIX1 induction by viral E6/7 may explain this apparent contradiction of the relationship of ER with ezrin. 19
Both ERα and β showed a sustained decrease in ir-ERα and β during the progression from normal epithelium to invasive CXCA. There have been conflicting reports regarding the expression of both ERα and β in CXCA. These may have been related to antibody-targeted epitopes. 20 However, ER-null animal studies have confirmed the necessity for ER in the development and growth of CXCA. 18 There are also several reports showing that SERM cause decreased growth and apoptosis of CXCA. 20 Moreover, estradiol resulted in clear estrogen actions in CXCA cells in culture, and this was reversed by the SERM tamoxifen. 13,14,21 -24
Sex steroids 25 and growth factors 22 regulate the expression of ERs. Since ERα and β are products of separate genes on separate chromosomes, a single mutational event that affects both genes is not likely. As well, there is no evidence of a systemic loss of ER or ER function in CXCA. Epigenetic regulation of ERs by cancers has been most widely studied in breast cancer. The 2 prominent regulators are methylation of the promoter site and dysregulation of microRNAs (miRNAs). 26 -28 It has been shown that in prostate cancer, methylation of 2 sites on the ERβ promoter caused the progressive loss of ERβ observed during carcinogenesis. 29 Although there are dysregulations of miRNA that affect ezrin, see below, thus far, none that affect ERs in CXCA have been reported. Although G protein-related ER (GPER) has been linked to carcinogenesis, 22 no direct effect of HPV E proteins on GPER has been reported. Taken together, these considerations indicate that the change in ER expression must reflect local conditions, probably related to the effects of HPV E6/7. 18,19,30 -33
Conclusions
We have elucidated the progressive fall of ir-ER α and β during the development of CXCA. This was accompanied by a progressive rise in ir-ezrin expression. This dichotomy is contradictory to previous findings that show estrogen to induce ezrin expression. This result implies that another agent is in play. SIX1 induces ezrin and has, itself, been shown to be induced by HPV E6/7.
Ezrin regulates the cell phenotype, including normal characteristics such as cell polarization, junctional specializations, and cell shape; but, excess ezrin could contribute to the development of the metastatic phenotype including lack of cell polarization, tenuous cell adhesion specializations, and the metastatic phenotype. These considerations raise the possibility that the rise and/or mutations of ezrin play a role in the transition from preinvasive to invasive cancer. We have recently submitted evidence of the latter possibility.
Human papilloma virus has been confirmed to be the causative agent in numerous cancers not generally associated with sex steroid action. The present findings is reasonable to test for the presence and action of sex steroid receptors and of ezrin in these cancers.
SERMs have been shown to inhibit the growth and progression of CXCA in animal models. While this is a transient response, the same is true in the treatment of other estrogen-sensitive lesions such as breast cancer. It is reasonable to suggest epidemiologic studies into the incidence of cervical cancer in women undergoing long-term antiestrogen treatment and the initiation of clinical antiestrogen trials in cases of cervical neoplasia.
Footnotes
Acknowledgments
Seung Do Choi and Ahmed Fadiel contributed equally to this project.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Supported by Varadi Ovarian Initiative for Cancer Education