
Abstract
Our electron microscopy observations demonstrate for the first time that the number of microvilli on the mice oocyte membrane decreases when meiosis progresses from prophase I to metaphase II (MII) stage, and the morphology of the microvilli also changes. Microvilli are significantly shorter and larger on the ovulated oocyte membrane than at the previous stages. Although clathrin vesicles clearly disappear during oocyte maturation, exosome-like vesicles begin to be secreted at the metaphase I stage, more strongly at the MII stage. Multivesicular bodies are visible only at the MII stage. Since several oocyte tetraspanins are involved in the gamete interaction, Cd9 being congregated on the MII oocyte microvilli, we analyzed the effect of tetraspanin deletion on oocyte membrane morphology. The Cd9−/− and Cd9−/− Cd81−/− deletions are associated with a decreased microvilli density on the MII oocyte surface. Microvilli thickness is significantly increased whatever the deleted tetraspanin gene be. Only Cd9 deletion clearly disturbs the vesicular traffic, increasing the number of clathrin and exosome vesicles. Additional investigations are necessary to elucidate how tetraspanins modulate the microvilli morphology, likely in relation with cytoskeleton. The role of oocyte exosomes in gamete adhesion/fusion remains to be further studied.
Keywords
Introduction
The Cd9 tetraspanin is essential for the oocyte fertilizing ability. Indeed Cd9 knockout (KO) mice present a severely reduced gestation number and reduced litter size. 1 –3 By contrast, Cd9−/− males have a normal fertility. A second tetraspanin Cd81 is also found on the oocyte membrane and presents an important homology with Cd9 4 suggesting similar properties. Actually, its overexpression partially recovers Cd9−/− oocyte fertilizing ability. 5 Interestingly, while Cd81-deficient oocytes present a reduced fusion ability and Cd9-deficient oocytes a severely impaired fusing ability, their double KO produces oocytes that are totally refractory to sperm oocyte fusion. 6 This suggests that both tetraspanins work synergistically and that the presence of either one is mandatory for the fusion to occur. This also suggests that their joint presence increases the oocyte fusogenic ability.
Interestingly, Runge et al have shown that Cd9 tetraspanin is located on the mouse oocyte microvillar membrane. In fact, to increase the exchange capacity of a cell while limiting volume variations (ie, increase the surface/volume ratio), epithelial cells, oocytes, lymphocytes, hepatocytes, and placental cells grow thin extensions termed microvilli. They extend the membrane surface to facilitate exchanges (absorption and secretion) and depending on the cell type can also serve mechanical functions, including physical support and mechanotransduction.
Microvilli are known to play a role in membrane fusion of somatic cells and gametes during fertilization since fusion doesn’t occur in the amicrovillar region 7,8 ; they could provide a scaffold for the presentation of adhesion/fusion proteins. 9 Microvilli of the oocyte are dynamic structures, 10 and it has been shown that after fusion with sperm 11 or resumption of cleavage, 12 they change their structure and/or distribution. Furthermore, Runge et al have also found that Cd9 molecules are preferentially congregated on the microvilli compared to the planar membrane regions in between the microvilli. On Cd9 KO oocytes, their density and curvature radius are increased and height reduced. 7 These authors suggest that microvilli could act as a platform, which concentrates adhesion/fusion proteins and/or provides a membrane protrusion with a low radius of curvature required for the sperm–oocyte interaction. 9 These data are in accordance with the fact that Cd9 generates adhesion sites responsible for the strongest gamete interaction previously reported. 13 Actually Cd9 generates adhesion sites that are strongly anchored to the microvilli actin core. 7 These adhesion sites mediated by Cd9 give rise to strong adhesion that imposes, during the whole interaction lifetime, a tight proximity of both gamete membranes, which is a requirement for the fusion to take place. 13 In the absence of the Cd9 tetraspanin, adhesion sites appear dispersed and weakly bound to the cytoskeleton, reducing the sperm–oocyte interaction efficiency. It is of interest to know whether these strong adhesion sites that we described are related to the microvilli morphology and what could be the effect of Cd9 and/or Cd81 deletion on their structure.
We have shown that oocyte membrane fragments containing at least Cd9 are transferred to sperm membrane by a trogocytosis process. 14 Furthermore, oocytes produce exosome-like vesicles that interact with sperm head upon their penetration into the perivitelline space (PVS). 15,16 We have also shown that oocyte membrane fragments containing at least Cd9 are transferred to sperm membrane by exosome-like vesicles. 15 These membrane fragment exchanges might be important in triggering gametes fusion. Exosomes and exosome-like vesicles consist of small vesicles (30-120 nm) formed in endosomal compartments containing internal vesicles (multivesicular bodies [MVBs]) that store membrane-bound structures. 17 Sets of specific surface or adhesion molecules allow exosomes to target specific recipient cells. 18,19 The protein family most commonly associated with exosomes is that of tetraspanin, specifically Cd9. 17 Even if Cd9 tetraspanin is nonfusogenic by itself, it could play a role in the formation of multimolecular complexes that will accomplish this function. 13 This is the reason why we also investigated the effect of tetraspanin deletions on vesicles secretion.
Materials and Methods
This work was approved and registered under the number CEEA34.BL.006.12 by the Comité d’Ethique pour l’Expérimentation Animale, Paris Descartes University.
Germinal Vesicle Oocytes Recovery and In Vitro Maturation
The C57Bl/6J female mice (5-8 weeks old; Janvier Labs, France) were stimulated with pregnant mare serum gonadotropin (PMSG, 5 IU; Intervet, Beaucouzé, France). Their ovaries were recovered 36 hours later in M2 medium (Sigma-Aldrich, Saint-Quentin Fallavier, France), and only the largest follicles were punctured to release their prophase I oocytes (containing a germinal vesicle [GV]). Some of these oocytes were immediately fixed, and the others were maintained in culture in FertiCult medium (FertiPro, Beernem, Belgium) at 37°C under 5% CO2 during 6 to 8 hours until metaphase I (MI) formation and were then fixed.
Metaphase II Oocytes Recovery
The Cd9−/− , Cd81−/− , double KO Cd9 and Cd81 female mice (a gift from C. Boucheix, Inserm, Villejuif, France), and wild-type (WT) C57Bl/6J female mice (5-8 weeks old; Janvier Labs) were stimulated with 5 IU PMSG and 5 IU human chorionic gonadotropin (hCG; Intervet) 48 hours later. Twelve to 14 hours after the hCG injection, female mice were killed and their cumulus-enclosed metaphase II (MII) oocytes were collected by tearing the ampulla wall of the oviduct and placed in FertiCult medium supplemented with 3% bovine serum albumin (BSA; Sigma-Aldrich) at 37°C under 5% CO2 in air under mineral oil (Sigma-Aldrich). The recovered ovulated oocytes were in the MII stage. Surrounding cumulus cells were removed by a brief exposure to 0.01% hyaluronidase (Sigma-Aldrich) and kept at 37°C before fixation.
Oocytes Preparation for Electronic Microscopy Observation
After 5 washings, oocytes were fixed in Sorensen medium supplemented with 2.5% glutaraldehyde for 30 minutes at room temperature and 1 hour at 4°C. After rinsing, they were fixed again, put in Sorensen medium containing osmic acid 0.1 M, and dehydrated by successive 15 minutes bathes in ethanol 70%, 90%, and 100%, 30 minutes in Epon/ethanol 100% (50:50), and finally 24 hours in Epon at 60°C for polymerization. Sections (90 nm) were performed using a Reichert S microtome (leica microsysteme, Paris, France) and observed with a Jeol 1011 (Croissy sur Seine, France) microscope. Images were visualized through a Gatan camera (Evry, France). Microvilli, MVBs, clathrin, and exosome-like vesicles were counted and their numbers were related to the oocyte membrane length, the section thickness being constant (90 nm). A membrane structure was considered as a microvillus and measured when its height was higher than its width. For each microvillus, its length was measured and its thickness in the middle were measured using a digital micrograph software.
In Vitro Insemination and Analysis of Sperm Exosomes and/or Oocyte Membrane Fragments Incorporation
Before insemination, oocyte zona pellucida (ZP) was dissolved with acidic Tyrode solution (pH 2.5; Sigma-Aldrich) under visual monitoring at 4°C. Then, ZP-free oocytes were rapidly washed 5 times and kept at 37°C under 5% CO2 in air for 2 hours, before insemination.
A Cd9−/− C57Bl/6J 14-week-old male mouse was sacrified by cervical dislocation. Cauda epididymis was recovered in 500 µL FertiCult supplemented with 3% BSA and gently squeezed to release spermatozoa, which remained in the medium covered by oil during 1.5 hours at 37°C under 5% CO2 for capacitation. The concentration of motile spermatozoa used for insemination was 1 × 105 cells/mL.
Zona pellucida-free oocytes were placed in the sperm suspension for 8 minutes and then fixed in Sorensen medium containing 4% paraformaldehyde for 30 minutes. After rinsing, they were incubated with an anti-Cd9 antibody (10 µg/mL in FertiCult, KMC8; BD PharMingen, San Diego, MD, USA) for 3 hours on a moving platform. Then they were washed 5 times for 2 minutes in FertiCult and incubated for 1.5 hours with a gold-conjugated secondary antirat antibody (1/30, EM GAR 5 nm; British Biocell International, Cardiff, United Kingdom).
Statistical Analysis
Student t test and variance analysis were performed. The number of clathrin vesicles was related to the total circumference of the oocytes section. Differences were considered significant at P < .05.
Results
Evolution of WT Oocyte Microvilli Morphology and Vesicles During Meiosis
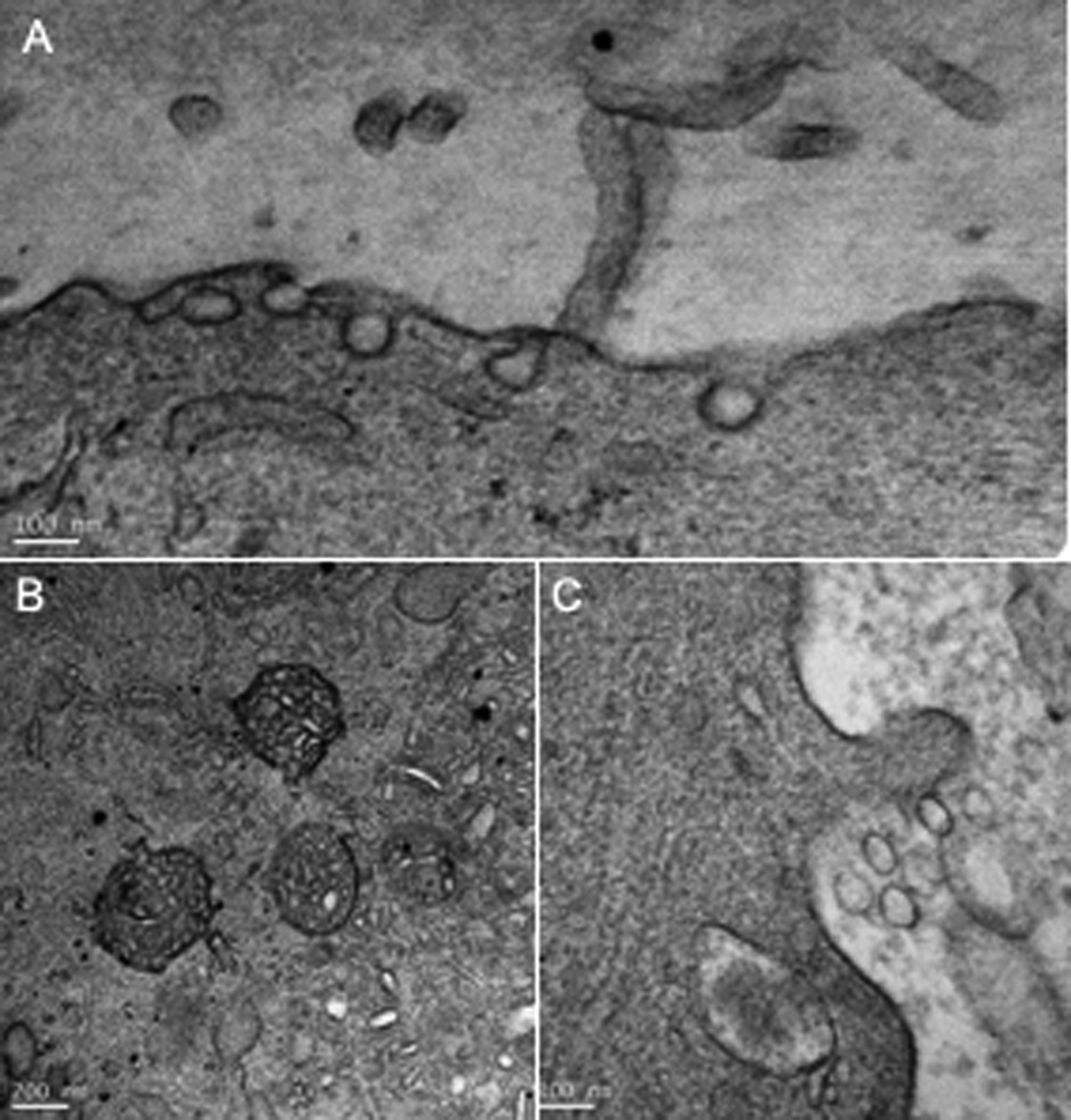
As shown in Figure 1 and Table 1, at the GV stage (number of oocytes, n = 2), the oocyte microvilli density was 4.8 ± 0.9 per µm. They appear almost parallel and included in the ZP thickness; this rendered their measure somewhat approximate, however, with a similar approximation for each one. Microvilli were 386 ± 132 nm long and 75 ± 20 nm wide. There was a virtual PVS at that stage of oocyte maturation.

Oocyte microvilli morphology related to meiosis stages. Germinal vesicle (GV): regular and parallel microvilli, homogeneously distributed on the membrane, well set in the zona pellucida (ZP). Metaphase I (MI): longer microvilli, often gathered together. Metaphase II (MII): less, larger, and shorter microvilli, nonhomogeneously distributed on the membrane, exhibiting different curvatures and orientations (magnification, ×20 000).
Wild-Type Oocyte Morphological Characteristics According to Meiosis Stage.
Abbreviations: GV, germinal vesicle; MI, metaphase I; MII, metaphase II.
aSignificantly different from GV and MI, P < .001.
bSignificantly different from GV and MII, P < .001.
When meiosis resumed after the luteinizing hormone (LH) surge, rupture of the tight junctions between oocyte microvilli and cumulus cells and microvilli morphology changes occurred. The PVS increased (529 ± 192 nm) and microvilli appeared free in it. At the MI stage (n = 3), their number was unchanged (4.3 ± 1.1 per µm). They presented the same thickness, but they were longer (P < .001). At the MII stage (n = 3), the microvilli density decreased significantly (P < .001). Their length was reduced compared to GV and MI oocytes (P < .001) and their thickness increased (P < .001).
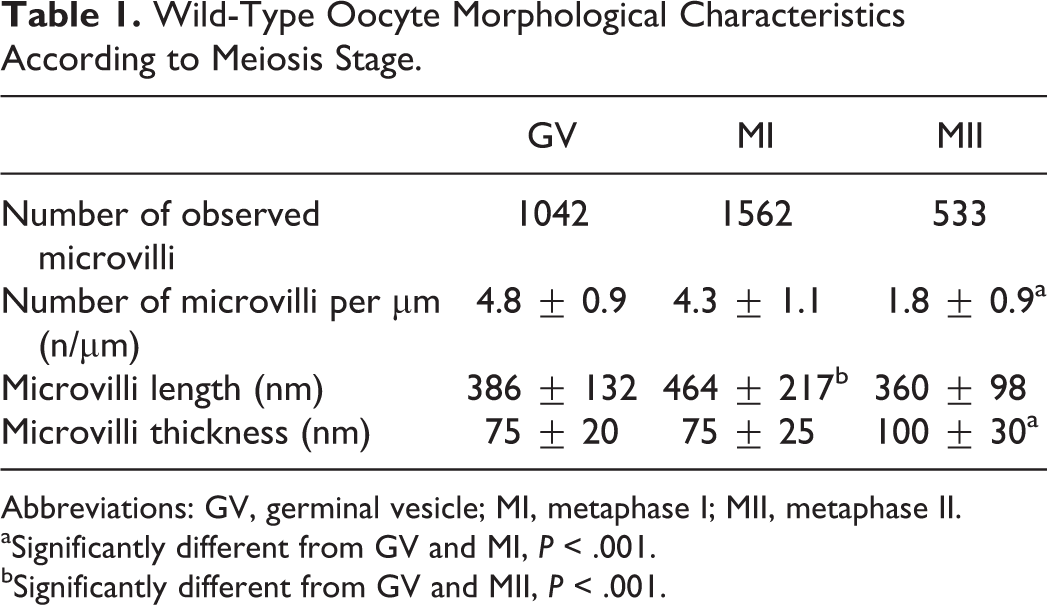
Interestingly, the number of clathrin vesicles that were numerous at the GV stage (Figure 2A) decreased at the MI stage to disappear at the MII stage, meanwhile, MVBs were seen only at the MII stage (Figure 2B) and few exosome-like vesicles appeared at the MI stage to be numerous at the MII stage (Figure 2C; Table 2).
Oocyte exosomes cycle. A, Clathrin vesicles: juxtaposition of opened or closed clathrin vesicles mostly observed in a germinal vesicle (GV) membrane oocyte (magnification, ×100 000). B, Multivesicular bodies (MVBs) observed in a wild-type (WT) metaphase II (MII) oocyte (magnification, ×50 000). C, Exosome-like vesicles observed in the perivitelline space (PVS) of a WT MII oocyte (magnification, × 100 000).
Wild-Type Oocyte Vesicle Secretion Cycle.
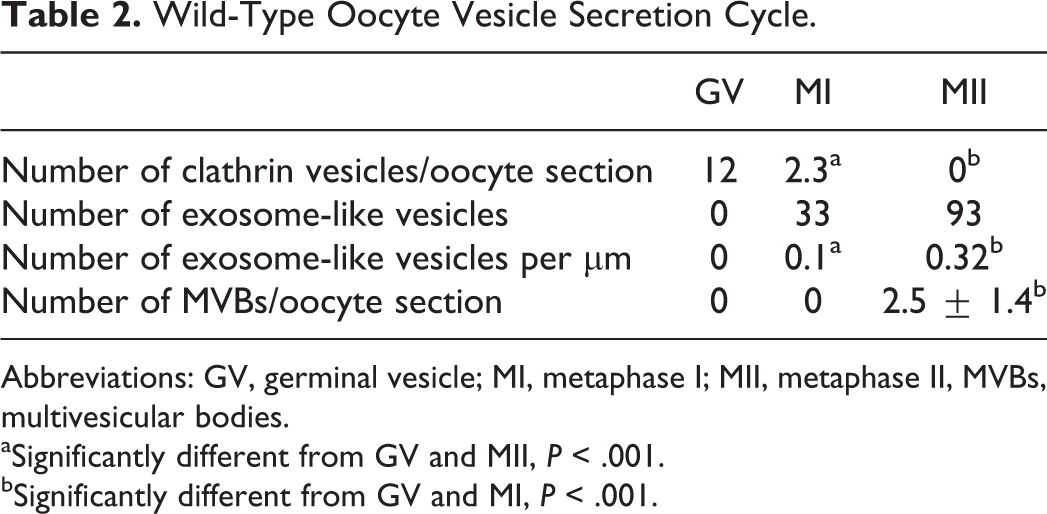
Abbreviations: GV, germinal vesicle; MI, metaphase I; MII, metaphase II, MVBs, multivesicular bodies.
aSignificantly different from GV and MII, P < .001.
bSignificantly different from GV and MI, P < .001.
Metaphase II Oocyte Membrane Microvilli Morphology and Exosome-Like Vesicles Associated With Tetraspanin Gene Deletion
As shown in Figure 3 and Table 3, the different studied parameters of the microvilli were modified after gene deletion.

Metaphase II (MII) oocyte microvilli morphology related to tetraspanin gene deletion. Wild-type (WT) oocyte: numerous entangled microvilli. Cd9−/− oocyte: clearly less microvilli. Cd81−/− oocyte: a little bit less microvilli. Cd81−/− Cd9−/− oocyte: almost no microvilli on a flat membrane.
Effect of Gene Deletion on MII Oocyte Morphological Characteristics.a
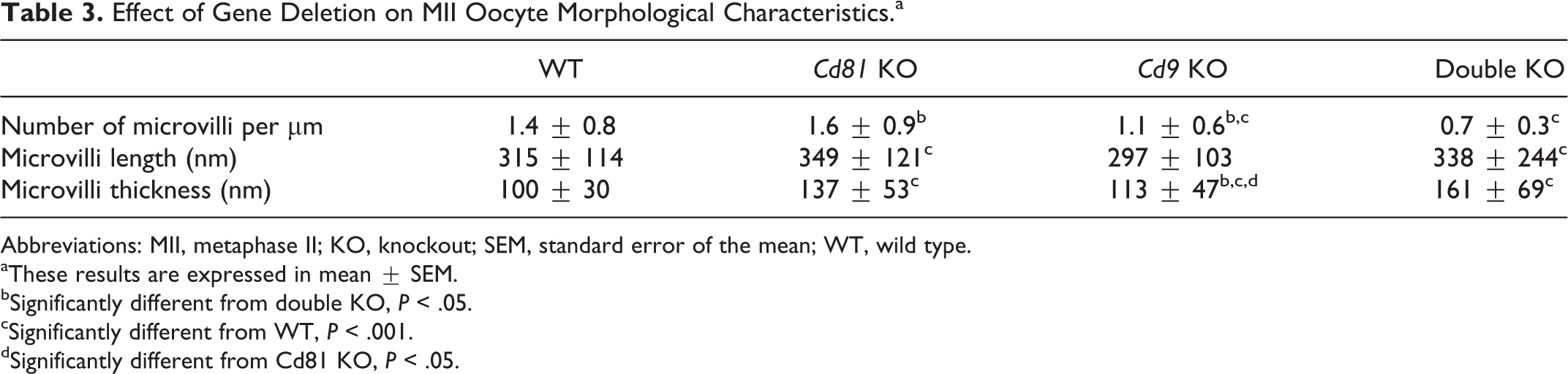
Abbreviations: MII, metaphase II; KO, knockout; SEM, standard error of the mean; WT, wild type.
aThese results are expressed in mean ± SEM.
bSignificantly different from double KO, P < .05.
cSignificantly different from WT, P < .001.
dSignificantly different from Cd81 KO, P < .05.
Microvilli density
Although the microvilli density was similar between Cd81−/− and WT oocytes (n = 3 and 3, respectively), it was significantly lower (1.1 ± 0.6 vs 1.4 ± 0.8, P < .001) in Cd9-deleted oocytes (n = 2) and (0.7 ± 0.3 vs 1.4 ± 0.8, P < .001) in double Cd9 Cd81–deleted oocytes (n = 3).
Microvilli length
If their lengths were similar to WT oocytes in Cd9−/− oocytes, they were significantly longer in Cd81−/− (349 ± 121 vs 315 ± 114) and in Cd81−/− Cd9−/− oocytes (338 ± 244 vs 315 ± 114; P < .001, respectively).
Microvilli width
Their thickness was significantly larger whatever the tetraspanin gene deleted when compared to WT oocytes (P < .001). After many comparisons, we considered that the radius of curvature at the tip of the microvilli, determined to be the radius of a circle, which fits the curve at the tip of the microvillus, 7 was finally not very different from half of the thickness of a microvillus. Thus, compared to WT oocytes, the microvilli radius of curvature was significantly increased whatever the tetraspanin gene deleted be (WT: 49.8 ± 15.2 nm: Cd81−/− : 68.6 ± 32; Cd9−/− : 56.6 ± 23.5; Cd9−/− Cd81−/− : 80.6 ± 34.5; P < .001).
Exosome-like vesicles
Surprisingly, although their number was strongly increased in Cd9−/− oocytes (0.9 vs 0.32, P < .001), it was significantly smaller in Cd81−/− and in double KO oocytes (0.1 per µm, in both cases, P < .001; Figure 2 and Table 4). The number of MVBs was smaller in Cd9−/− and in double KO oocytes (2.5 ± 1.4 vs 1.5 ± 1.4, P < .001) and similar to WT in Cd81−/− oocytes. Although no clathrin vesicles were visible in WT and in double KO oocytes, very few were observed in Cd81−/− oocytes (0.14, P < .001) and many in Cd9−/− oocytes (2, P < .001).
Effect of Gene Deletion on MII Oocyte Vesicle Secretion Cycle.
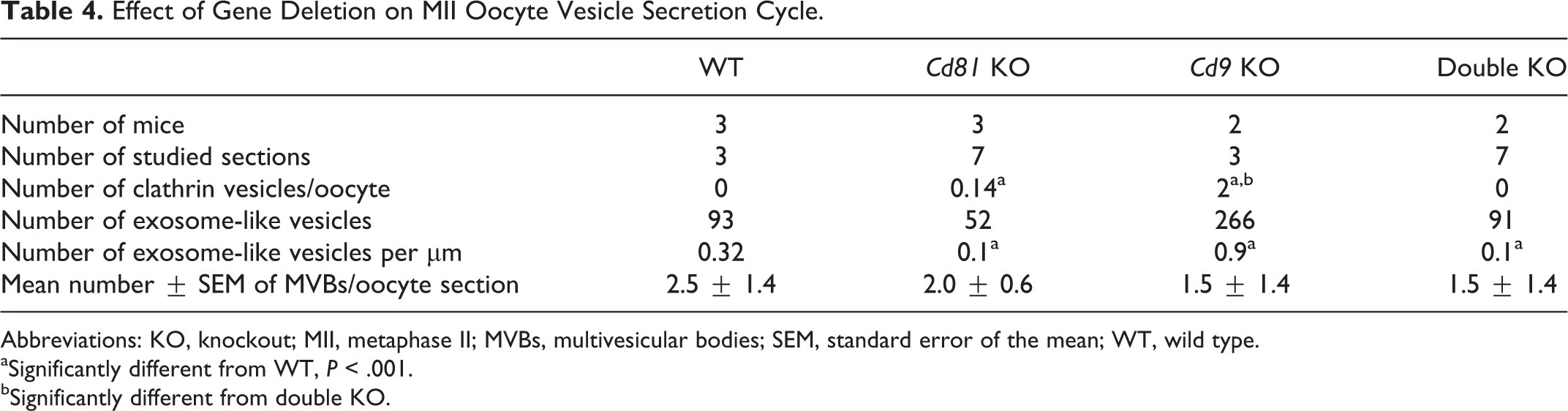
Abbreviations: KO, knockout; MII, metaphase II; MVBs, multivesicular bodies; SEM, standard error of the mean; WT, wild type.
aSignificantly different from WT, P < .001.
bSignificantly different from double KO.
Oocyte Membrane Fragments Incorporated by Spermatozoa
To verify the incorporation of Cd9-containing oocyte membrane fragments in sperm, we inseminated WT ZP-free MII oocytes with Cd9−/− sperm in order to be sure that the observed Cd9 was really belonging to the female gamete. Oocytes were fixed after contact with sperm and stained with a gold-conjugated anti-Cd9 antibody as described above. Eight minutes after insemination, all the observed sperm exhibited a gold staining on their membrane, confirming the transfer of membrane fragments from the oocyte to the sperm (Figure 4).
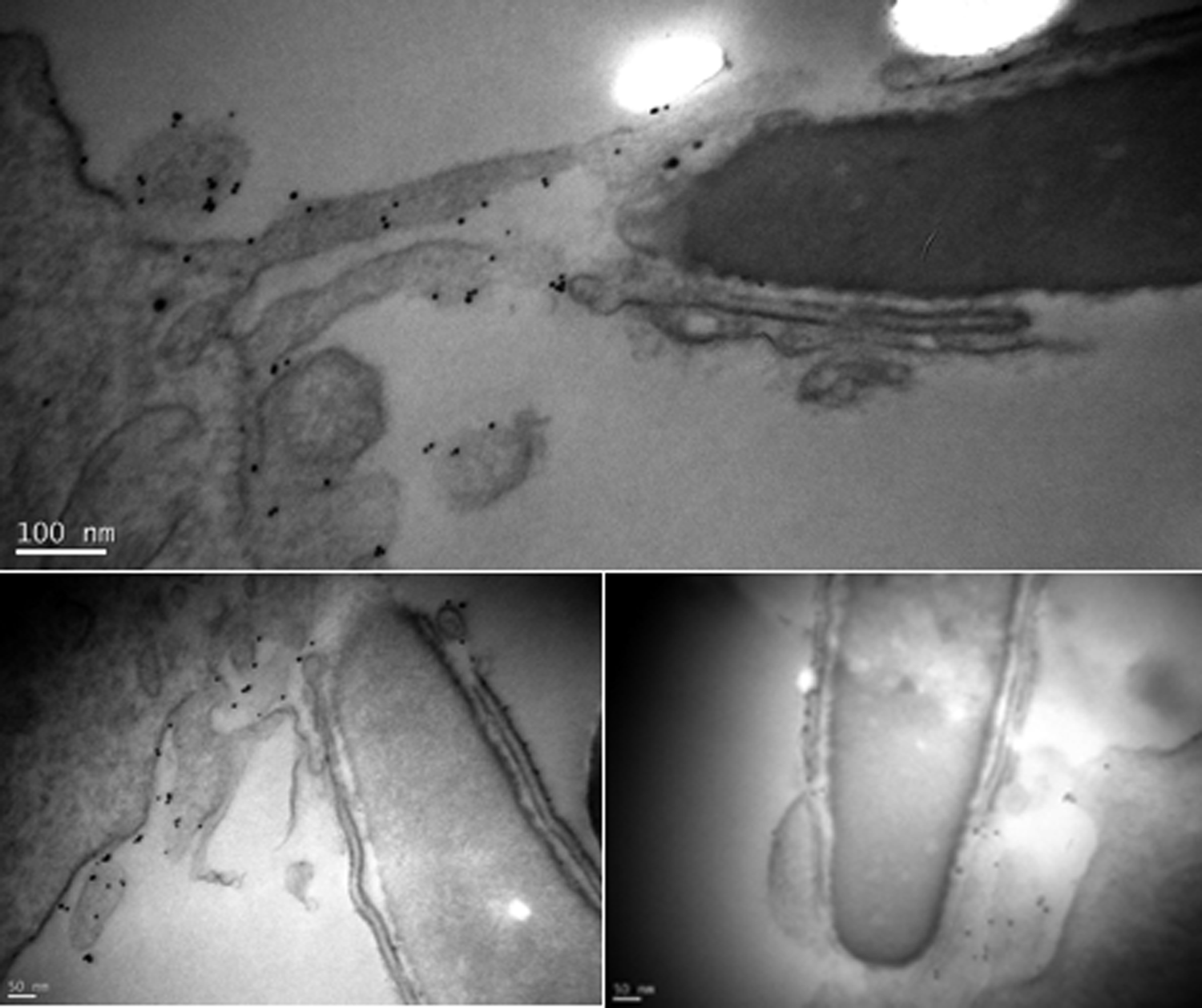
Oocyte membrane fragments incorporated by a spermatozoon. Gold spots were revealed on the Cd9−/− spermatozoa membrane 8 minutes after fertilization, demonstrating fragments transfer from the oocyte to the spermatozoon.
Discussion
Our study describes for the first time the morphological evolution of the mouse oocyte membrane analyzed by electron microscopy during meiosis. At the GV stage, microvilli are numerous, thin, and very close to the other, settled into the ZP. Due to their connections with the cytoplasmic extensions of the corona radiata cells and to the fact that the PVS is almost nonexistent, the microvilli are held as wheel spokes. In fact, gap junctions have been observed between these 2 structures 20 that are disrupted at the LH surge, the disruption being followed by meiosis resumption. At the MI stage, the PVS appears, probably because of gap junctions rupture. The microvilli become free in this space and longer than at the previous stage. By contrast, at the MII stage, the number of microvilli drops by half, their length decreases, while their thickness increases. During oocyte maturation, just before GV breakdown, the links between the oocyte and its surrounding cells are broken; thus, it can be speculated that the microvilli are no more stretched, explaining mechanically that they are at the shorter and larger end. A similar decrease in the number and length of microvilli has been observed during the in vitro maturation of human GV oocytes. 21 An approximately 4-fold decrease in microvillar length is also described in 1-methyladenine-induced starfish oocytes maturation. 22 It is also interesting to notice that when the Xenopus stage VI oocyte matures, a significant portion of the membrane area in microvilli is internalized, leading to a loss of microvilli and flattening of the cell membrane. 23 Very recently, Courjaret et al have demonstrated that Ano1 regulates the length and diameter of microvilli in Xenopus oocytes through its interaction with ezrin–radixin–moesin (ERM) proteins and the actin cytoskeleton. 24 Accordingly, we have observed that the inhibition of the 3 ERM proteins disrupted the morphology of mouse oocyte microvilli, increasing their radius of curvature (unpublished data). The decrease in the number and length of oocyte microvilli could facilitate sperm movements inside the PVS and better permit contact and then fusion between sperm and oocyte membranes. Another important reason for this decrease in microvilli number seems related to the fact that although GV oocytes have many necessary interconnections with their surrounding granulosa than cumulus cells through these microvilli and gap junctions, MI oocytes have fewer contacts, whereas MII oocytes that will be ovulated have no more contact with their surroundings cells.
Our data also highlight the disruption of microvilli morphology associated with Cd9 and/or Cd81 tetraspanin gene deletion. According to the results already published by us and by others on the effects of tetraspanin gene deletion on fertilization, we tried to establish a correlation between these effects and microvilli structure modifications. The comparison between MII oocytes from mice deficient for one or the other or both tetraspanins and MII oocytes from WT mice reveals a progressive decrease in the number of microvilli related to the deleted gene. Considering that Cd81−/− mice are slightly hypofertile, when Cd9−/− mice are very hypofertile and Cd81−/−Cd9−/− mice are sterile, 6 it is interesting to notice that the number of microvilli is similar in Cd81−/− and WT oocytes, it decreases in Cd9−/− oocytes, and even more in Cd81−/−Cd9−/− oocytes. Thus, microvilli density seems to be a good marker of fertility. Moreover, tetraspanin deletion, whatever the gene, induces an increase in microvilli thickness and thus of their radius of curvature. It is probably this aspect that explains the complementary and partially redundant nature of the role of these 2 tetraspanins. This has already been shown, but only for the Cd9−/− oocyte. 7 By contrast, microvilli length, which is not modified in Cd9−/− oocytes and increased in Cd81−/− and double KO oocytes compared to WT oocytes, does not appear as a marker of fertility.
We summarized the importance of the microvilli in the fertilization process in Table 5: the more numerous (while remaining below the values found at the GV stage) and thin the microvilli are, the more the oocytes can be fertilized.
Effect of the Absence of Tetraspanin Cd9 and/or Cd81 on Microvilli Morphology Correlated With Fertility Degree.a
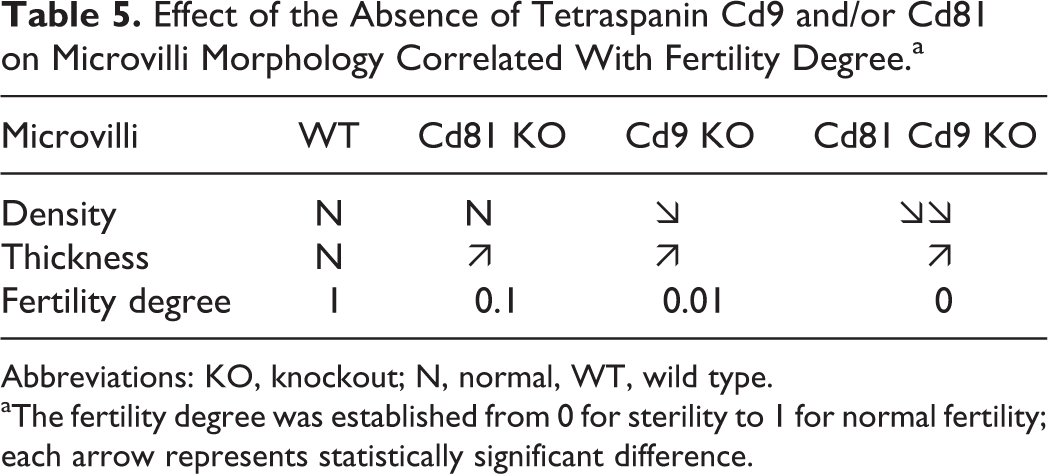
Abbreviations: KO, knockout; N, normal, WT, wild type.
aThe fertility degree was established from 0 for sterility to 1 for normal fertility; each arrow represents statistically significant difference.
Another important point demonstrated for the first time in the oocyte is the presence of the different elements of the exosomes cycle production; although clathrin vesicles are numerous at the GV stage, they disappear when meiosis resumes and MVBs and exosome-like structures appear. Exosomes are a particular type of membrane vesicles secreted by pluricellular organisms. They are delimited by a membrane and located near the plasma membrane of the cell. 25 They are formed in MVBs of cells. 26,27 Our observation of few MVBs inside the ooplasm of MII oocytes is in accordance with a previous report on human egg, in which no clear picture is shown. 28 Hence, among the vesicles observed in the PVS, and called exosome-like vesicles, some are real exosomes. Actually, as observed in eukaryote cells, 29 the sequence clathrin vesicles, MVBs, extracellular vesicles of 50 to 100 nm in diameter seen in the oocytes strengthen the existence of the exosomes formation circuit. A vesicular traffic exists in the mouse oocyte, which seems to occur sequentially according to oocyte maturity. Clathrin vesicles disappear when MVBs and exosomes appear in the PVS. Even if oocyte exosome functions are not yet known, during its journey, the sperm acquires many exosomes from different tissues. This is the case of epididymosomes in the epididymis and prostatosomes in the ejaculate, 30 –32 then within the female reproductive tract in the sperm reservoir. 33 At each step, the spermatozoa acquire a new function or a modification of an already acquired function. We can therefore hypothesize that these oocyte exosomes are important for the sperm oocyte membrane binding and fusion process. In fact, only Cd9 deletion clearly disturbs the vesicular traffic, increasing the number of clathrin and exosomes vesicles. This vesicular traffic disturbance could contribute to explain how oocyte Cd9 deletion prevents membrane fusion at fertilization, preventing the reorganization of some molecules at the surface of the oocyte, necessary for the sperm fusion.
Finally, our data confirm for the first time by electronic microscopy that oocyte membrane fragments containing Cd9 can be observed on sperm heads after in vitro fertilization assays as already demonstrated by immunofluorescence microscopy. 14 –16 However, oocytes from Cd9−/− mice are also able to transfer membrane fragments to sperm, 15 meaning that Cd9 is not the necessary element from the transferred fragment, if this transfer is really necessary. In fact, whereas 1 team has demonstrated that this transfer is necessary for the fertilization process, 16 2 other teams have been unable to confirm this result. 15,34
Conclusion
Microvilli number and morphology are modified during oocyte meiosis in WT mice. Associated with tetraspanin gene deletion, a decrease in the number and an increase in the thickness of the microvilli are observed. In parallel, the exosomes cycle production gets ready during meiosis and is also disturbed when tetraspanin genes are deleted.
Footnotes
Authors’ Note
Brigitte Lefèvre and Jean-Philippe Wolf contributed equally to this work.
Acknowledgments
The authors thank C. Boucheix and E. Rubinstein for their gift of Cd9−/− , Cd81−/− , and Cd9−/− Cd81−/− mice (Inserm, UMR-S1004, Villejuif, France). They also thank A. Schmitt from the Plateforme Microscopie Electronique Cochin Imagerie for achieving electronic microscopy experiments.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by ANR-13-BSV5-0004-02 (DYNAGAME).