
Abstract
Almost every part of our body has a coevolved microbial community. The expressed microbial genes comprise the various microbiomes that play important roles in normal physiology and development. The various microbiomes are separate, yet often connected, with the species composition of one affecting others. The female reproductive system microbiomes (eg, vaginal, placental, and mammary/milk) remain less well explored than the gut microbiome although they comprise a large proportion of the female microbial network. This review examines the evidence for interconnectivity between the female reproductive microbiomes, other maternal microbiomes, and developing infant microbiomes and the potential roles of each in health and disease. Disruptions in maternal microbiomes may be linked to pregnancy complications and maternal, fetal, and neonatal health. The diversity of the vaginal microbiome’s makeup, which appears to vary across ethnicity, has led researchers to reconsider the idea of a “healthy” or “normal” vaginal microbial community. Less is known about the possible placental microbiome, although an association between the placenta’s bacterial makeup and preterm labor and other pregnancy complications is being investigated. The mammary/milk microbiome appears to be influenced by maternal characteristics and may play a role in inoculating the infant but may also be affected by the infant’s oral microbiome. Probiotic therapies such as “vaginal seeding” offer potential health benefits but require more rigorous testing. Exploring the reproductive microbiomes in detail and pairing this information with an individual’s detailed medical history will provide a more complete picture of the status and importance of the microbial network to health.
Introduction
Few if any tissues in our body are truly sterile. Just about every part of our body appears to have a coevolved microbial community living on it. The total mass of microbes on and in our bodies is about 3% of our total weight, and it is comprised of approximately 10 times as many cells as our own body. Microbial DNA represents several orders of magnitude more expressed genes than is in our own genome. 1 The human microbiota comprises multiple microbial ecologies in different organs that have coevolved to coexist with the human host. It is becoming increasingly clear that the various microbial ecologies on and in our bodies have important function for our health and well-being. They represent complex microbial communities in which the members produce bioactive molecules both necessary for and inimical to other microbes and also interact with our own expressed genome to regulate and influence our metabolism, physiology, and immune function and are important for our health and resistance to disease.
Many microbiotas of the human body have been studied recently due to the advent of advanced sequencing techniques and the production of libraries for the prokaryote 16S ribosomal RNA gene (16S rRNA) that enable detection of microbial taxa that cannot be cultured. Body fluids previously considered mostly sterile (eg, amniotic fluid and milk) have been shown to contain microbial DNA, suggesting they may support microbial communities or be in contact with tissue that does support a microbiota (eg, placenta and mammary tissue, respectively). 2,3 As knowledge regarding the various microbiotas grows, it is becoming evident that many are interconnected to varying extents and that characteristics of one can affect the characteristics of others. 4 For example, the newly discovered placental microbiota shares many microbial taxa with the maternal oral microbiota, 5 and in mice, it has been demonstrated that members of the oral microbiota can reach the placenta through hematogenous transmission. 6 The milk microbiota contains species from the maternal gut microbiota. 3,7
The term microbiome technically refers to the totality of the expressed genetics of the microbiota 1 although it is sometimes loosely used to refer to the structure of the microbial community as well. Thus, the vaginal microbiome properly refers to the suite of expressed genes from the vaginal microbiota, but in common parlance the distinction is often not made and the vaginal microbiome is used to refer to the vaginal microbial community as well as to the expressed microbial genome. The distinction can be important, however. For example, individuals have been shown to share suites of microbial genes that produced functionally similar metabolic pathways although they differed in the actual taxonomic units of microbes harbored in their gut microbiota. 8 Thus, their microbiotas can be more different than their microbiomes. 1 Conversely, different strains of bacteria that cannot be distinguished by their 16S ribosomal genes can have different gene expression patterns, or the same strain may express genes differently depending on environmental conditions, 9 potentially making the microbiomes more different than the microbiotas. In essence, microbiotas are distinguished by their microbial taxa based on the 16S rRNA gene and microbiomes by their gene expression potential, usually estimated by whole genome shotgun sequencing or even possibly by sequencing the transcriptome (the messenger RNA from expressed DNA). In practice, the term microbiome tends to be used to refer to both.
Major research efforts are beginning to produce an understanding of the diversity and extent of the microbial communities that have coevolved with us (eg, MetaHIT; NIH Human Microbiome Project). Although the gut microbiome is currently the best studied, for providers of health care for women, there are other microbiomes perhaps as or even more relevant to the reproductive health of their patients, such as the vaginal, placental, and milk microbiomes. This review focuses on the microbiomes most relevant to obstetrician-gynecologists, the roles of these microbiota in health and disease, and the potential interconnectivity between microbiomes.
Microbiome Benefits
It has been long known that portions of the digestive tract can harbor large microbial communities. The stomachs of cows, the ceca of rabbits, and the human colon house substantial microbial ecologies that assist the host in digestion. Microbes play a substantial role in the digestion of food, especially fibrous foods, for many animals. It has recently become abundantly clear that our gut microbiota does far more than digest fiber. The gut microbiome produces a large array of biologically active products that can affect other gut residents, gut epithelium, and even be absorbed into circulation. An aberrant gut microbiome is associated with increased disease risk in infants. 10 Obesity in adults is associated with an altered gut microbial population (as well as an altered milk microbiome in lactating women; see Maternal Milk Microbiome section). Recent studies have shown the importance of the gut microbiome in human physiology and metabolism. For example, malnutrition in twins studied in Malawi has been linked to differences in the gut microbiome which affected digestive and absorptive efficiency and even affected the Krebs cycle. 11 The range of benefits conferred to the host by the various microbiomes include defense against pathogens, microbial gene products for use by the host, and products of microbial metabolism (eg, fatty acids and vitamins).
Providing resistance to pathogenic microbial colonization is a prime benefit of a healthy microbiome. Although antibiotics have saved millions of lives, one side effect noticed early on was a greater susceptibility to colonization by pathogens after antibiotic treatment. 12 A healthy microbiome resists colonization by noncommensal microbes. Infection by the pathogen Clostridium difficile is often associated with recent antibiotic treatment. A normal fecal microbiota has been shown to be associated with enhanced resistance to C difficile colonization. In a randomized trial, duodenal infusion of fecal microbiota from healthy donors was more successful at treating patients with recurring C difficile infection compared to treatment with vancomysin. 13 Researchers are currently in the early discovery phase for developing probiotic treatments for protection against infectious diseases, especially for the new antibiotic-resistant strains that are becoming uncomfortably common. 12
In addition, there are extensive interactions between microbes, their products and the host immune system that are often symbiotic, not antagonistic. Gnotobiotic rodent models display anatomical, physiological, metabolic, and immunological deficiencies. 14 The lack of a microbiome produces disruptive changes in the developmental pattern. The maturation of the intestinal epithelia, especially its associated immune system, appears to be strongly influenced by commensal colonizing microbes. 15,16 The development of the neonatal immune system appears highly influenced by microbiome effects. 14 There appear to be critical time windows in which the developing immune system “learns” to be tolerant of its commensal microbes.
Many Microbiomes
The Human Microbiome Project sampled 300 healthy, 18- to 40-year-old adult men and women over multiple body sites (9 oral cavity sites, 4 skin sites, the nares, and a stool sample, with an additional 3 vaginal sites for women) to create a reference data set for exploring and understanding the structure and function of the coevolved human microbiome. 17 –19 This foundational work has established the extent of the remarkable diversity of microbial communities that exist on us and the variation both between body sites and between individuals. The oral and the stool (representing lower gut) microbiotas were the most diverse communities, with the oral cavity microbiome rivaling the gut microbiome in complexity, diversity, and numbers of taxa. 17 Different tissues within the oral cavity support different microbial communities, with different microbiotas found on the tongue, teeth, below the gum line, palate, tonsils, and so forth. In contrast, the vaginal microbiota appears relatively simple, although with significant diversity between individuals and within individuals over time. 17
Race/ethnicity was among the stronger metadata factors associated with microbiome composition. 17 Given the likely extensive coevolution between host and microbiome, it is hardly surprising that there would be strong associations between the structure and function of the microbiome and the host ancestral genome. Several host mitochondrial DNA (mtDNA) haplotypes exhibit significant associations with the host stool and vaginal microbiomes, 20 and evidence from twin studies suggest host genetic effects on gut microbiome structure. 21,22 A recent study that mined the Human Microbiome Project whole-genome shotgun (HMP WGS) data for host DNA reads found significant associations between host DNA and microbiomes in many body sites. 23
The bodily microbiomes are interconnected in many ways. Microbes in the mouth can easily transfer into the bloodstream and enter systemic circulation. The oral cavity microbiome shows a strong association with what appears to be a placental microbiome. 5 Periodontal disease has long been considered to be a risk factor for adverse pregnancy outcomes such as low birth weight and preterm birth. 24 Recent meta-analyses support a moderate association between periodontal disease and preterm birth, although somewhat complicated by differing definitions of periodontal disease and other factors. 25,26 The connection between the oral and the placental microbiotas provides a potential mechanism. Microbes associated with the gut and vagina are also found in the placenta. Microbes of one microbiota appear to have the possibility of spreading to others, sometimes with no ill effects, but other times spreading pathogens or even commensals of one microbiota that can be pathogenic in another. Although in this article we will focus on the microbiomes of the female reproductive system, ultimately, each microbiome cannot truly be understood outside the interconnected microbial web within the body.
Vaginal Microbiome
The vaginal microbiome has been investigated for over 100 years, beginning with Albert Döderlein’s 1892 monograph Das Scheidensekret, 27 in which he reported the existence of gram-positive, rod-shaped, nonsporing bacteria in vaginal secretions of healthy women. Döderlein correctly hypothesized that these Lactobacillus species were the source of the lactic acid present in the vagina. The presence of Lactobacillus and an acidic vaginal environment has long been considered important for vaginal health. 28
Bacterial vaginosis (BV) has been called a disease of microbiota dysbiosis, a disruption of the normal vaginal microbial community leading to overgrowth of pathogenic or at least noncommensal microorganisms. 29 Bacterial vaginosis is a common problem among women of child-bearing age, in some high-risk populations affecting as much as half the population. In studies of US women, BV prevalence ranged from 6.1% to 51.4%. 30 Prevalence rates tended to be lower in Asian-Pacific Islanders and white women and highest in non-Hispanic African American women. It is characterized by vaginal discharge which releases a “fishy” odor after adding 10% potassium hydroxide to the discharge sample.
Bacterial vaginosis increases a woman’s vulnerable to sexually transmitted infections, pelvic inflammatory disease (PID), and during pregnancy, premature birth, and fetal loss. 31 Screening for and treating BV is an important aspect of women’s health relevant to the obstetrician-gynecologists. Often used characteristics as part of BV diagnosis are a vaginal pH greater than 4.5 (one aspect of the Amsel criteria 32 and a low prevalence of Lactobacillus species among the vaginal microbiota, Nugent criteria). 33 However, both of these characteristics have been shown to exist in asymptomatic healthy women, especially among women of African descent and of Hispanic origin, 34 contributing to the confusion and uncertainty regarding BV in different populations. 29
The vaginal microbial community serves many purposes, including maintaining a healthy vaginal pH and resisting invasion by noncommensal microbes. In general, an asymptomatic vaginal microbiota is thought to be comprised of many species of Lactobacillus and other microbes that contribute to a diverse ecosystem. But the makeup of the vaginal microbiota is not necessarily consistent, either across populations of healthy women or within a woman over time; it varies between races/ethnicities, 17,28,34 differs between pregnant and nonpregnant women, 35,36 and can change depending on the menstrual cycle, sexual activity, and environmental change. 37 There are healthy women with vaginal microbiotas that contain few, if any, Lactobacillus species. About one-fourth of women who are of reproductive age and display no symptoms of BV have low levels of Lactobacillus (the typical biomarker of BV) and a vaginal pH higher than 4.5. 34 A study of asymptomatic South African women found that only 37% had a cervicovaginal microbiota dominated by Lactobacillus. 38
Interestingly, the vaginal communities with few, if any, Lactobacillus species contained high proportions of other lactic acid-producing species (eg, Atopobium and Megasphaera), suggesting that the production of lactic acid is an important conserved element of the human vaginal microbiome, 39,40 a possible example of microbiota differences that are greater than the microbiome differences. However, the community with low levels of Lactobacillus species generally produces a slightly higher vaginal pH (Table 1), leading to asymptomatic women with that vaginal microbiota type being at risk of a false-positive diagnosis of BV by Amsel and Nugent criteria.
Suggested Clustering of Vaginal Microbiota into Five Types Based on the Dominant Species.

aAdapted from Ravel et al. 34
Stability of the vaginal microbiota also varies among healthy women. In some healthy women, the microbial makeup of the vaginal microbiota can change rapidly and dramatically, without leading to disease. 37 Although the microbial structure changed over time in terms of taxa, the authors suggested that the inherent functions of the microbiome (eg, production of lactic acid) may have varied less, an example of the microbiota being more variable than the microbiome. These data suggest that typical markers of BV might not be as accurate as once considered, given that asymptomatic patients exhibit such varied microbial profiles. 39
Some researchers have attempted to cluster vaginal microbial communities into community state types (CSTs). For example, Ravel and colleagues identified 5 different types of vaginal CSTs; 4 (types I, II, II, and V in Table 1) dominated by lactic acid-producing species in the genus Lactobacillus, and one community (type IV) with low proportions of Lactobacillus and high proportions of anaerobic species from a number of different genera (eg, Atopobium, Megasphaera, Prevotella, and Sneathia; Table 1). 34 Women of European and Asian ancestry are more likely to have a vaginal microbiota dominated by Lactobacillus (eg, types I, II, III, and V), while Hispanic and African American women are more likely to have a type IV vaginal microbiota. 34 A more recent study with a larger sample size found the same general pattern between African American and white women but also detected 2 other vaginal microbiota types: one dominated by Gardnerella vaginalis and the other by bacterial vaginosis-associated bacterium 1 (BVAB1), both of which were more common among African American women. 41
Other researchers have cautioned that there is a lack of consensus regarding defining CSTs and in their interpretation. For example, Koren and colleagues showed that factors such as clustering methodology, distance metric, sequencing depth, whether the sequence data come from WGS or 16s rRNA, and the variable 16s rRNA region used can affect the number of CSTs a researcher will find in a data set. 42 Different methodologies can derive different results from the same data set. For many body sites, the variation in microbial taxa appears to be better explained by gradients rather than discrete clusters, although variation among women in their vaginal microbiome appears more likely to be multimodal than microbiomes of other body sites. 42 And defining a CST by the lack of dominance by Lactobacillus may be questionable.
For many women, their vaginal microbiota is not stable and will change over relatively short time periods and can change due to life events. For example, an increase in Lactobacillus species appears to be a common finding during pregnancy. 43 Although nonpregnant Hispanic women in the United States were found to be less likely to have a Lactobacillus-dominated vaginal microbiota, 98% of pregnant Mexican women had Lactobacillus in their vaginal microbiota. 44 Although the vaginal microbiota appears to be generally more stable during pregnancy, in some women transitions between CSTs appear common. 43
There also appears to be a dramatic and lasting change in the vaginal microbiota after parturition, with a substantial decrease in Lactobacillus and an increase in the overall diversity of the vaginal microbiota. 43,45 This change in microbial community occurred regardless of the pregnancy microbiota composition 45 and was sustained for up to 1 year postpartum. 43
There is a rapid decrease in circulating estrogen in women immediately postpartum, which will be sustained during lactation. If a woman breast-feeds her baby, she will likely remain in a low-estrogen state. High-circulating estrogen, such as during pregnancy, is thought to create a vaginal environment hospitable to Lactobacillus species by increasing the glycogen content of the vaginal epithelium. Although Lactobacillus species generally cannot ferment glycogen, the digestive enzyme α-amylase is capable of breaking down glycogen into products Lactobacillus species can ferment. 46 Human vaginal fluids have α-amylase activity, 46 although whether the amylase is from the host genome or from other microbes is uncertain. 47 Thus, a combination of high glycogen content in vaginal epithelium and the presence of an enzyme with α-amylase activity in the vaginal fluid will be conducive to a vaginal microbiota dominated by Lactobacillus. Conversely, a low-estrogen state (eg, lactation or post menopause) will produce a vaginal environment less conducive to dominance by Lactobacillus. Postmenopausal women generally have lower abundance and diversity of Lactobacillus in their vaginal microbiota. 48 In one study, reduced vaginal colonization by Lactobacillus was associated with more severe symptoms of vaginal dryness 49 ; however, low abundance of Lactobacillus was seen in studies of asymptomatic postmenopausal women as well. 48 A lack of Lactobacillus does not necessarily imply a disease state.
Vaginal Microbiota and the Infant
Infants are born relatively sterile; not as sterile as was previously thought, but the biomass and diversity of microbes in the various infant microbiotas are certainly low compared to adults or even to their microbiotas a few days after birth. Maternal inoculation of her infant is considered a critical component of the development of infant microbiotas, with inoculation by the vaginal microbiota thought to be a critical factor for infants born vaginally. Interestingly, the vaginal microbiota appears to undergo consistent changes during pregnancy, with a decrease in both microbial diversity and richness, 35,36,43,50 suggesting the possibility of regulated changes to the vaginal microbiota prior to parturition. Thus, a neonate born vaginally potentially will be exposed to a regulated, probably coevolved microbiota that serves as an early microbial inoculation.
Babies delivered by cesarean section will not naturally receive this vaginal inoculation of microbes. An early study relying on culture methods found that infants delivered by cesarean section start life with a different gut microbial community than vaginally delivered babies. 51 Studies using 16S rRNA and WGS sequencing methods have generally found similar results but with some qualifications. A small study found that the various microbiotas (eg, oral, skin, and stool) of vaginally delivered babies were enriched in maternal vaginal microbes, while those of cesarean-delivered babies were enriched in maternal skin microbiota. 52 Many studies have found babies born by cesarean section to have low or even absent levels of Bacteriodes species in their stool microbiota, at least over the first months of life. 53–56 However, in the study by Yassour and colleagues, in addition to all 4 infants born by cesarean section, 7 of 35 vaginally born infants also lacked Bacteriodes species in their stool over the first 6 months. Infants with this early-life low fecal Bacteriodes characteristic had low fecal bacterial species diversity, regardless of delivery mode, even at 3 years of age. 56
The establishment of the gut microbiome in infants is affected by many inputs beyond mode of delivery. Other significant factors generally found to be associated with the structure of the infant gut microbiota are feeding method, age, and antibiotic exposure. 53,56 –58 Factors such as maternal adiposity appear to play a role, as well. A small study found that the intestinal microbiota of vaginally born infants (but not those born by cesarean section) differed between those born to obese mothers and normal weight mothers. 59 Nonetheless, even after accounting for these other inputs, many studies have found mode of delivery to be a significant factor at least in early life. 54,55,58 Caution is warranted, however, given that many of these studies have had small sample sizes, especially for babies born by cesarean section and that the effect of other maternal factors (eg, pregnancy complications) on the inoculation of neonates are not completely understood.
A recent pilot study attempted to restore the microbiota of cesarean section-delivered infants to the vaginal delivery state by swabbing them with maternal vaginal fluids after birth. 60 Thirty days after birth, swabbed cesarean section-delivered infant microbiotas were similar to those of vaginally delivered infants, suggesting the promising use of vaginal swabs in inoculating infants with the appropriate microbiota. 60 In this study, mothers with sexually transmitted diseases (including group B Streptococcus infection), BV, or signs of viral disease were excluded. The results of this pilot study suggest that normalization of cesarean-delivered infant microbiotas by transfer of maternal vaginal fluids is possible, but the study was small and the results far from definitive.
These findings have contributed to the practice termed “vaginal seeding” for babies born by cesarean section, in which a gauze swab is used to transfer maternal vaginal fluid (and thus vaginal microbes) to the infant. 61 A clinical trial to investigate vaginal seeding is underway, however, the main outcome being investigated is whether vaginal seeding can affect the neonatal microbiome and not clinical outcomes. 62 At present, a benefit to the infant from vaginal seeding is not demonstrated, and the possibility that pathogens asymptomatic in the mother may have the potential for serious harm to the infant has led the American College of Obstetricians and Gynecologists to release a Practice Advisory recommending against the practice. 63 Vaginal seeding of babies born by cesarean section deserves rigorous study. It represents an inexpensive, low-tech possibility to restore an important component of health to these babies. However, at present, the evidence suggests only the potential for benefit, with significant potential risks, as well. 61
Pelvic Inflammatory Disease
Pelvic inflammatory disease (PID) is an inflammation of the upper reproductive tract caused by infection by pathogens such as Chlamydia trachomatis, Neisseria gonorrhoeae, and Mycoplasma genitalium. It also can be associated with pathogens linked to BV. 64 Pelvic inflammatory disease appears to be a polymicrobial disease. 65 Clinical diagnosis of PID is based on pelvic organ tenderness in conjunction with lower genital tract inflammation. Verification of the clinical diagnosis can involve laparoscopy, transcervical aspiration for histopathology, transvaginal ultrasonography, or magnetic resonance imaging, with laparoscopy considered the standard. 64 Similar to BV, the clinical diagnosis of PID can be imprecise. In one study, as many as 25% of women with a clinical diagnosis were not confirmed to have PID by laparoscopy. 66
Disruptions of the vaginal microbiota associated with BV are risk factors for PID. 65 A healthy vaginal microbiota appears to be protective against PID, possibly due to resistance to pathogens associated with PID. What is not clearly understood is whether there is a microbiota in the upper genital tract that may also serve as protection. 65 The upper genital tract has been assumed to be mostly sterile with the presence of bacteria indicating a probable disease state. Indeed, upper reproductive tract infections are associated with many pregnancy complications, including miscarriage, preterm birth, and neonatal sepsis. 67 However, both culture and culture-independent methods have shown the presence of bacteria in the upper reproductive tract of asymptomatic women, 65,67 although with both lower abundance and diversity than in the vagina. 68 The colonization of the female upper reproductive tract by microbes generally did not induce a significant inflammatory response, 68 indicating that the microbial community was benign. Nonpathogenic microbes routinely can be found in amniotic fluid 43 and in meconium of newborns 69 in the absence of apparent disease. The womb naturally may not be completely sterile.
Recent studies have detected what appears to be microbial communities in the upper reproductive tract including ovaries and the fallopian tubes of women who were undergoing surgery for removal of these organs. 70 –72 Women who had ovarian cancer displayed a different microbial community structure than women without cancer. 72 The results of these studies have been presented at scientific meetings but have not yet gone through the rigorous peer review process and thus must be considered preliminary.
Placental Microbiome
Starting with the discovery of bacteria in the amniotic fluid of cesarean section pregnancies in 1927, 73 the idea that the womb is a sterile place has been challenged. Recent research has shown that a unique, although relatively low biomass microbiota may exist within the human placenta. Intracellular bacteria have been found within the placental basal plate using both culture and light microscopy methods 74 ; the authors suggested that these bacteria may have come from the endometrial epithelium after implantation. Another study using culture methods found bacteria in placentas from extremely preterm births (23-27 weeks gestation). Organisms were more likely to be cultured from placentas exhibiting inflammation (31% versus 68%). 75 Species associated with BV were more likely found in placentas from women with self-reported cervical/vaginal infection 76 and were associated with an inflammatory response by the neonate, while the presence of Lactobacillus was associated with a reduced risk of an inflammatory response. 77
Several studies using sequencing technologies have also demonstrated the existence of microbes in the placenta, 5,78 –83 although the study by Amarasekara and colleagues using polymerase chain reaction on 16S rRNA only found evidence for bacteria in placentas from pregnancies complicated by preeclampsia and even then only in 12.7% of cases. 76 The weight of evidence from these studies, especially from those using WGS, suggests the placenta has a low-biomass microbiota, 5,84 with potential associations between the microbiota composition and pregnancy complications, which are briefly reviewed subsequently.
Studying women who give birth to preterm infants offers an opportunity to explore the role placental bacteria may play in parturition. Based on 16S rRNA sequence data, a study found that term and preterm placenta differ in their microbial communities. 78 Another study found Burkholderia is enhanced in preterm placenta. 5 A study using WGS found inflammation to be a significant factor for the microbiotas of both term and preterm placentas and that both preterm birth and chorioamnionitis were associated with a less diverse placental microbiota. 83 It is possible that the bacterial infiltration of the placenta induces labor through inflammation. 78,84 The bodily response to bacteria, both maternal and fetal, may well be important factors in the timing of labor and parturition. 69
The microbes detected in placenta appear more similar to bacteria sampled from maternal oral microbiotas, rather than other maternal microbiomes (ie, stool or vaginal microbiomes). 5 Hematogenous spread of bacteria from the maternal oral cavity has been suggested as a route of bacterial movement to the placenta. 5,84 This process may be facilitated by bacteria like Fusobacterium nucleatum. 5 Accessibility to the bloodstream in the mouth provides a potential mechanism that links the oral microbiome to the placental microbiome and a potential proximate mechanism for why periodontal disease may be related to an increased risk of preterm birth. In one study, 8 of 26 women who gave birth preterm had the periodontal disease-associated bacterium Porphyromonas gingivalis in both maternal oral and amniotic fluid samples. 24
Other maternal factors may influence the placental microbiota. For example, a 16S rRNA sequence study considered the effect of preexisting obesity and gestational weight gain on the makeup of the placental microbiota and showed that excessive maternal weight gain (but not preexisting obesity) alters the placental microbiota in cases of preterm (but not term) parturition. 85
Researchers believe that the placental microbiome may play a role in colonizing the infant gut microbiome if the placental bacteria are able to enter amniotic fluid and be swallowed by the infant. 67,82 Variation in the gut microbiome of infants of differing gestation lengths suggests that in utero colonization via placental bacteria may be an ongoing process throughout gestation. 82 In a study of 2 births, 1 vaginal and 1 cesarean section, the microbiota of the first feces from the neonates more closely resembled the placental microbiota compared to the maternal vaginal microbiota. 80
One caution regarding the evidence for a placenta microbiome: the fact that bacteria can travel to the placenta and take up residence is not proof that there is a naturally occurring, stable placental microbial community. Although the womb is not a sterile environment, it is more protected from microbial colonization than the other microbiomes as evidenced by the lower biomass of microbes in the placental microbiota. Future research will decide the issue; at present, in our opinion, the evidence is highly suggestive but not yet definitive for the existence of a stable placental microbial community with coevolved function for the host. The evidence appears strong, however, in support of a role for bacterial–placental interactions in the risk of early parturition, likely mediated through inflammatory responses.
Maternal Milk Microbiome
Milk was also previously considered to be a relatively sterile body fluid, which was one of the putative advantages to breast-feeding. Microbes found in milk were thought to represent contamination from skin or the environment or a sign of infection in the mammary gland. However, 16S rRNA sequencing has shown that milk contains a consistent microbiota, which includes species that are maternal symbiotic gut microbes. Breast milk is known to influence the infant’s gut microbiome through milk oligosaccharides; it may also be another mechanism for the inoculation of the infant gut with maternal gut microbes. The gut microbial populations of breast-fed infants differ from that of bottle-fed infants 86 and generally reflect their mother’s gut biome. Evidence from human and rodent studies suggests that maternal breast milk might provide a crucial source of microbes that initially colonize the infant gut. 3,7,87 Breast milk contains Staphylococci, Streptococci, bifidobacteria, and lactic acid-producing bacteria, and the same strains that are found in a mother’s milk are usually found in her infant’s feces. 88 The feces of infants born by cesarean section to women given oral Lactobacilli during gestation contain that strain of Lactobacilli, although they were not exposed to the vaginal environment. 89 Breast milk is a possible colonizing source, implying that live Lactobacilli in the maternal gut can reach the mammary glands, enter the milk, and potentially colonize the neonate.
Thus, it appears possible that mothers can inoculate their infants with maternal gut microbes via milk, with potential physiological consequences for the infants. Maternal characteristics affect the milk microbiome as well. 90,91 Obese women produce milk with a less diverse set of microbes, and women who gain excessive weight produce milk with different microbes from those in milk of women who had normal weight gain during pregnancy. 90 This suggests that obese mothers may pass on their obesogenic gut microbiome to their breast-fed infants at least in part through their milk.
Breast milk is certainly not the only source of colonizing microbes for infants. Indeed, it is virtually impossible for babies not to be inoculated with their mothers’ microbiota. Molecules in milk, especially milk oligosaccharides, may play a larger role in shaping the infant gut microbiome than direct inoculation of microbes from milk. Interestingly, the microbial biomass of the milk microbiome is comparable to that of the placenta, indicating both placenta and mammary are relatively protected tissues. A similar caution to the one for the placental microbiota is warranted. The existence of microbes in breast milk samples does not definitively prove there is a natural microbiome in the lactating mammary gland. Microbes certainly get into breast milk, and the evidence is suggestive that maternal mechanisms exist to transport microbes into milk from other maternal microbiota, but the milk microbiota may not be a true microbial community, analogous to the vaginal or gut microbiotas.
One possibility is that the microbial content of milk serves as a signal to the mammary gland providing information regarding the maternal microbiomes, the infant microbiomes, and even the microbial content of the external environment. Imaging studies of the infant’s oral cavity and the breast during suckling have shown that there is retrograde flow in the ducts 92 which might carry fluid from the baby (mixture of saliva and milk) to the mammary. 93,94 Thus, the baby’s mouth may be a source of microbes that will be found in milk and vice versa. There is a functioning immune system in milk 95 ; milk contains living neutrophils, macrophages, and lymphocytes. 96 The bacteria and other microbes that get into milk within the ducts of the mammary gland, from maternal sources, the infant, or the environment, may “inform” immune cells of milk and the mammary gland. Breast milk has considerable immune function, from immune cells, and from a plethora of immune function molecules that are secreted into milk. Might the microbial content of milk be part of the regulatory system that shapes milk immune function?
A final comment regarding the milk/mammary microbiota concerns using probiotics to treat/prevent mastitis. There have been several studies of the effectiveness of probiotics to treat and prevent mastitis in dairy cattle that have generally shown positive results 97 –100 although not in all cases. 101 Mastitis is a major economic concern in the dairy industry, and its prevention without the use of antibiotics in this era of increasing antibiotic-resistant pathogens would be a significant advance. There are 2 published studies in women, one for treatment 102 and one for prevention 103 of mastitis that have shown some promise; however, both papers have been questioned due to concerns over methodology and the appropriateness of the outcomes measured. 104 There are several other trials currently ongoing that will hopefully shed more light on the issue. Despite the lack of definitive evidence, probiotic treatments for prevention and treatment of mastitis in women are being marketed to health professionals in Australia. 104
Concluding Remarks
Within every individual exists a unique and dynamic series of interconnected microbiomes important for health and resistance to disease. These microbial communities have coevolved with us and provide a multitude of benefits. Dysbiosis, the dysregulation of these microbial communities, can have negative health consequences. Indeed, the general prediction is that disrupting the coevolved microbial communities in our bodies will increase the risk of disease and poor health.
The vaginal microbiome is a critical component of a woman’s general and reproductive health and thus an area of biology relevant to the practice of obstetrics and gynecology. The study of the vaginal microbiome should inform and guide practice in areas such as bacterial vaginosis and other infections of the lower reproductive tract. The interconnected microbiomes of pregnant women (the maternal microbiomes; Figure 1) have implications for fetal and neonatal health.
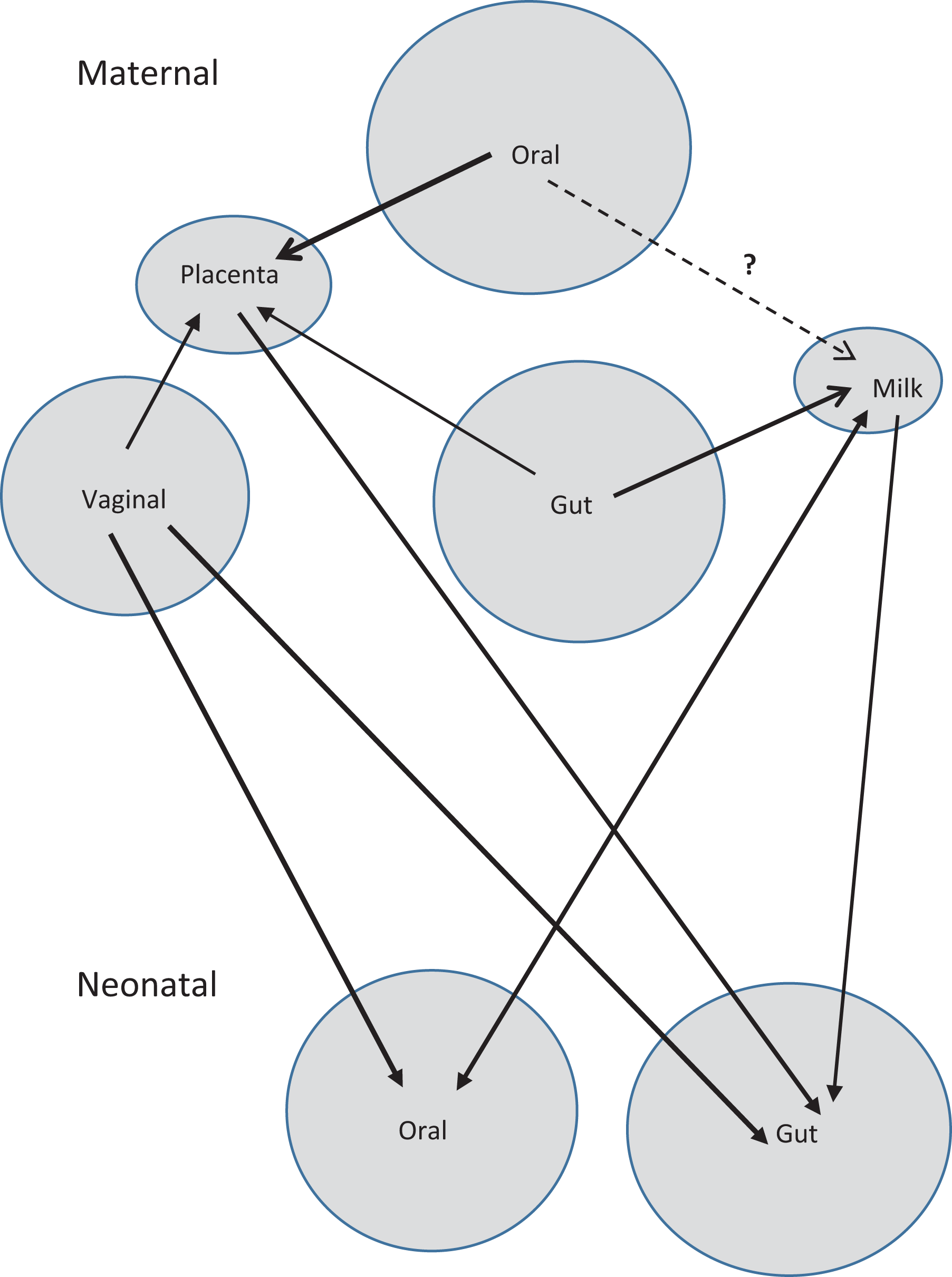
A representation of the interconnectedness of the maternal microbiomes both within the mother and with her neonate born vaginally. The microbiomes of a neonate born by cesarean section would show more connection with the maternal skin microbiome.
The discovery of potential microbiota in the upper reproductive tract, within regions of the body previously considered well protected against microbial invasion, needs to be better understood. Are these microbial communities stable, coevolved elements of female reproductive health? Or do they represent the ubiquity of microbes such that microbial invasions will always occur at a low level, but that the normal condition is for the upper reproductive tract, including placenta, to be relatively sterile?
With our increased understanding of the importance of the various microbiomes to human health and well-being has come well-intentioned therapies to “adjust” supposed dysbiosis back to a healthy microbiota. An important aspect to consider is the extent to which the structural dynamics of microbiomes are affected by universal factors (eg, environmental factors and initial colonization), group factors (eg, race/ethnicity, delivery mode, breast-fed or formula fed, and so on), or individual factors (eg, specific haplotypes, small nucleotide polymorphisms, and other unmodifiable characteristics of an individual). 105 The stronger the effects universal factors have on the structural dynamics of the microbiome, the more likely that manipulations will have consistent, predictable, and stable results. If group factors have a predominant effect, then manipulations that do not take group differences into account are likely to result in variable, unpredictable, and unstable results. If individual characteristics have strong effects, then generic probiotic treatments and other attempts at manipulating the microbiome will run the risk of not just being ineffective but potentially detrimental. 105 Therapies designed to improve health through manipulations of microbiomes (eg, treating/preventing BV or mastitis) deserve careful study. They have great potential for improving health and well-being. But our understanding of the structural dynamics of any microbiome is currently lacking to be able to confidently predict the outcomes of such manipulations for all people.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received the following financial support for the research, authorship, and/or publication of this article: Funding was provided in part by grant no. UA6MC19010 from the Maternal Child Health Bureau of the Health Resources and Services Administration (HRSA).