
Abstract
Objectives:
To estimate the effects and mechanisms of choline, an essential nutrient and a selective α7 nicotinic acetylcholine receptor (α7nAChR) agonist, on the prevention of symptoms and the effects on the cholinergic anti-inflammatory pathways (CAP) in a lipopolysaccharide (LPS)-induced inflammatory response in a rat model.
Methods:
Inflammation was induced by LPS treatment (1.0 μg LPS/kg body weight) on gestational day (GD) 14. Nonpregnant and pregnant Sprague Dawley rats were placed on a normal choline diet (1.1 g/kg) or supplemented choline diet (5.0 g/kg) from GDs 1 to 20. Systolic blood pressure (SBP), urinary albumin, and pregnancy outcomes were recorded. On GD 20, serum and placentas were assayed for cytokines. Western blots were used to determine the expression of placenta α7nAChR and components of the α7nAChR-CAP, including nuclear factor-κB (NF-κB) and protein kinase B (AKT). Immunohistochemistry was used to localize placental sites for the p65 subunit of NF-κB.
Results:
Lipopolysaccharide significantly increased SBP and urinary albumin and decreased pregnancy outcomes, and these effects were partially reversed by higher choline treatment. Choline supplementation also significantly attenuated the LPS-induced increase in serum and placental inflammatory cytokines, decreased the expression of placental α7nAChR, lowered the activation of NF-κB signaling in placenta mononuclear cells, and inhibited placental AKT phosphorylation.
Conclusion:
This study confirms that LPS induces inflammatory conditions in pregnant rats and shows that choline supplementation protects against the inflammatory symptoms through its action on α7nAChR and CAP. These observations have important implications for the prevention and treatment of inflammatory responses associated with pregnancy.
Keywords
Introduction
Inflammation during pregnancy is known to produce many adverse reactions and outcomes, including abortion, preterm birth, intrauterine growth restriction, and preeclampsia (PE). 1 The precise mechanisms leading to the development of inflammatory disorders of pregnancy remain elusive. Many reports suggest that these adverse conditions may be associated with an abnormal maternal inflammatory response, both systemically and locally in the placenta. 2 –4 Lipopolysaccharide (LPS), an endotoxin of gram-negative bacterial cell walls, has been used to demonstrate inflammatory processes associated with female reproduction in animals, and treatment with LPS leads to adverse outcomes including spontaneous abortion, preterm labor, PE-like conditions, and other short- and long-term fetal morbidities. 1,5 –10 We and others found that LPS injection in rodents caused preterm birth 9,11,12 and PE-like symptoms such as hypertension, proteinuria, adverse pregnant outcomes, and maternal inflammatory responses, both systemically and locally at the placenta. 12 –17 Our group has found that the LPS-induced inflammatory model exhibits cytokine profiles similar to those seen in women with PE. 12,14,17 These studies also demonstrate that treatment with nicotine, which is a selective agonist of α7nAChR, protects rodents from LPS-induced symptoms and inflammatory responses in the placenta by action on the cholinergic anti-inflammatory pathway (CAP) or nicotinic anti-inflammatory pathway.
Choline is also an agonist of α7 nicotinic acetylcholine receptor (α7nAChR) and is an essential nutrient that functions in phospholipid metabolism and neurotransmission and as a methyl donor. 18 Several earlier studies show that biological processes including inflammation, apoptosis, and angiogenesis are modified by choline supplementation. 19 –21 During pregnancy, the demand for choline increases substantially to support the growth and development of the placenta and fetus. 22 –24 Adults with diets very low in choline develop fatty liver, liver cell death, and muscle damage. 25 –29 In addition, higher dietary intake of choline improves signaling mechanisms responsible for placental angiogenesis, which may mitigate some of the pathological antecedents of inflammation. 30 Notably, the epigenome of the human placenta is especially responsive to maternal choline intake, for example, altered gene methylation and the expression of placental corticotropin-releasing hormone, which is a key regulator in stress responses. 31
Increasing evidence demonstrates that choline has an anti-inflammatory effect. An earlier report showed that choline exhibited anti-inflammatory activity in an animal model of arthritis. 32 Another study indicates that choline administration reduces oxidative stress and inhibits inflammatory responses in a rodent model. 33 Based on our previous and other studies of the effects of the CAP in inflammation and the anti-inflammatory effects of choline, in this study, we investigated whether gestational choline supplementation has a protective effect on LPS-induced inflammation and its effects on CAP.
Materials and Methods
Animals
The Sprague Dawley rats (male rat: 250-300 g; female rat: 180-220 g) were purchased from the Medical Experimental Animal Center (Guangzhou, Guangdong, China). Rats were housed in a standard laboratory condition (temperature 23°C-26°C, relative humidity 50%-60%, illumination between 06:00
Diets
Pregnant (P) and nonpregnant (NP) female rats were randomly assigned to either a choline-controlled, standard rodent chow diet (AIN-76A with 1.1 g choline chloride/kg; Dyets Inc, Bethlehem, Pennsylvania) or a rodent chow diet with choline supplementation (AIN-76A with 5.0 g choline chloride/kg; Dyets Inc), as reported previously. 34,35 These 2 levels of maternal choline intake were selected for these studies based on numerous prior investigations demonstrating lasting cognitive benefits of increased maternal choline intake in normal rodents. 35 The choline content of the control diet is considered to provide adequate choline intake during pregnancy. The choline-supplemented diet provides approximately 4.5 times the concentration of choline in the normal diet, within the range of dietary variation observed in the human population. Rats were provided ad libitum access to their assigned diets.
Rat Model
Experimental inflammation in rats was induced by infusion of LPS (1 µg/kg body weight) in 2 mL of sterile saline through an infusion pump into the tail vein (infusion rate, 2 mL/h) on day 14 of pregnancy. The NP and P control rats were infused with 2 mL of saline alone. Before tail vein injection, the rats were anesthetized by intraperitoneal injection of 10% chloral hydrate. The P and NP rats were placed on a normal choline diet (1.1 g/kg) or a supplemented choline diet (5.0 g/kg) from GD 1 through GD 20. The rats were randomly assigned to 5 groups: (1) NP (nonpregnant group placed on 1.1 g/kg choline diet, n = 6); (2) P (pregnant group placed on 1.1 g/kg choline diet, n = 6); (3) P + C (pregnant rats on 5.0 g/kg choline diet, n = 6); (4) LPS treated (inflammatory model group on LPS + 1.1 g/kg choline diet, n = 9); (5) LPS treated + C (inflammation-induced rats on LPS + 5.0 g/kg choline diet, n = 9). We did not consider it necessary to have NP groups with LPS and high choline because this study focuses on pregnancy.
Reagents and Antibodies
Lipopolysaccharide (Sigma-Aldrich, St Louis, Missouri) was dissolved in sterile pyrogen-free 0.9% saline. Stock solutions of 1.0 mg/mL LPS were stored in aliquots in a freezer and thawed and used whenever required. Antirat antibodies to the components of the CAP pathways were as follows: anti-α7nAChR and anti–nuclear factor-κB (NF-κB) p65 were purchased from Abcam (Grand Island, New York). An endogenous inhibitor of NF-κB (IκBα), phosphorylated IκBα (pIκBα), protein kinase B (AKT), and phosphorylated AKT (pAKT) antibodies were obtained from Cell Signaling Technology (Beverley, Massachusetts).
Measurement of Systolic Blood Pressure
On GDs 6, 11, 14 (before LPS infusion), 16, and 18 (between 9
Measurements of Urinary Albumin, Serum Parameters, Fetal Loss, and Fetal, Placenta, and Maternal Weights
On days 7, 12, 17, and 19 of pregnancy, rats were placed in individually metabolic cages for 24 hours (from the previous day at 6
On day 20 of pregnancy, maternal weights were measured and the rats were then anaesthetized by intraperitoneal injection with 10% chloral hydrate (30 mg/kg). Maternal serum was obtained from blood of the inferior cava vena. Fetal loss (%) was estimated by dividing the empty fetal attachment sites, due to both absorption and early fetal expulsion, by total number of placentas with fetuses + empty placental attachment sites. Fetal pups and placentas were removed and weighed. All placental samples were stored at −80°C for further assessment. The blood samples were centrifuged at 1400g for 10 minutes at room temperature, and serum creatinine (Cr), blood urea nitrogen (BUN), alanine aminotransferase (ALT), and aspartate aminotransferase (AST) were measured using an automatic analyzer (HITACHI 7600-020).
Measurement of Serum and Placental Pro-Inflammatory and Anti-Inflammatory Cytokines
Total lysates from 2 placentas per rat from each group except for the NP group were prepared by homogenizing 60 mg placental tissue in 600 µL of radio immunoprecipitation assay lysis buffer (50 mmol/L Tris–HCl, pH 7.4; 150 mmol/L NaCl; 1% NP-40; 0.5% sodium deoxycholate; and 1 mmol/L phenylmethanesulfonyl fluoride supplemented with a mixture of protease inhibitors; Roche Diagnostics GmbH, Mannheim, German) and then kept on ice for 30 minutes and centrifuged for 10 minutes at 14 000g. Protein concentrations were determined with bicinchoninic acid protein assay reagents (Pierce Biotechnology, Rockford, Illinois), according to the manufacturer’s instructions. Serum and placenta protein samples were thawed and a rat cytokine panel (ProcartaPlex Rat Th Complete Panel, 14 plex. Cat. No. EPX140-30120-901, eBiosciences, San Diego, California) was used to quantify various cytokines. The panel included pro-inflammatory cytokines (interleukin [IL]-1α, IL-1β, IL-2, IL-5, IL-6, IL-12P70, IL-13, interferon γ [IFN-γ], tumor necrosis factor α [TNF-α], granulocyte-macrophage colony-stimulating factor [GM-CSF], granulocyte colony-stimulating factor [G-CSF], and IL-17A) and anti-inflammatory cytokines (IL-4 and IL-10). Then multiplex magnetic bead-based antibody detection kits were used according to the manufacturer’s instructions using the Bio-Plex 200 system (Bio-Rad Laboratories, Inc, Hercules, California).
Immunohistochemistry of NF-κB
After fixation in neutral formalin, placental tissues (6 tissues/group) were embedded in paraffin using standard procedures. Sections (approximately 3-µm thick) were made with a microtome and placed on glass slides, deparaffinized in xylene, rinsed in ethanol, and brought to water through a series of decreasing concentrations of ethanol. Sections were then microwaved for antigen retrieval and pretreated with 0.3% H2O2. Subsequently, the sections were incubated in a humidified chamber at 4°C overnight, with goat serum, and in a primary antibody solution containing rabbit anti-NF-κB p65 antibody (1:1000 dilution). After washing, the samples were incubated with a suitable secondary antibody solution for 1 hour at room temperature. Finally, the sections were incubated in horseradish peroxidase–streptavidin for 1 hour at room temperature, and the color reaction was developed with diaminobenzidine. Sections were counterstained with hematoxylin, dehydrated through a series of increasing ethanol concentrations to xylene, and placed under coverslips and then photographed under a light microscope.
Western Blotting
Preparation of total lysate from the placentas has been described earlier. For nuclear protein extraction, total lysate from placenta was suspended in hypotonic buffer and then kept on ice for 15 minutes. The suspension was mixed with detergent and centrifuged for 30 seconds at 14 000 rpm. The nuclear pellet obtained was resuspended in complete lysis buffer in the presence of a protease inhibitor mixture, incubated for 30 minutes on ice, and centrifuged for 10 minutes at 14 000 rpm. Protein concentrations were determined with the bicinchoninic acid protein assay reagents (Pierce Biotechnology), according to the manufacturer’s instructions. For immunoblots, the same amount of protein (40-80 mg) was separated electrophoretically by sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane. The membranes were incubated for 2 hours with the following antirat monoclonal rabbit antibodies at 1:500 (α7nAChR) to 1:1000 (all others) dilution: α7nAChR, AKT, pAKT, IκBα, pIκBα, and NF-κB p65. For total proteins in the immunoblots, β-actin and glyceraldehyde 3-phosphate dehydrogenase were used as loading controls. For nuclear protein, anti-histone H3 antibody was used as a loading control. After washing in Dulbecco phosphate-buffered saline (PBS) containing 0.05% Tween 20 four times for 10 minutes each, the membranes were incubated with goat antirabbit immunoglobulin G or goat antirat antibodies for 2 hours. The membranes were then washed in Dulbecco PBS containing 0.05% Tween 20 four times for 10 minutes each, followed by signal development using enhanced chemiluminescence system (Thermo Scientific, Rockford, Illinois).
Statistical Analysis
All data are expressed as mean ± SEM (standard error of mean). The differences between groups were analyzed by 2-way analysis of variance (ANOVA) for studies measured at various times, followed by the least significant difference post hoc test or Dunnett test as appropriate. Measurements done at one specific time were analyzed by one-way ANOVA. Ratios were compared using the χ2 test or Fisher exact test. Before executing the ANOVA, data were tested for normality and equal variance. Data were analyzed for statistical significance using the Statistical Package for Social Sciences software for Windows, version 13.0 (SPSS Inc, Chicago, Illinois). The level of significance was defined as P < .05.
Results
Effects of LPS and Choline Supplementation on Manifestation of Inflammation
The SBP of the various pregnant groups, on GDs 6, 11, 14, 16, and 18, and corresponding days for nonpregnant rats, is shown in Figure 1. There were no significant differences (P > .05) among the 5 groups before LPS treatment on GDs 6 to 14. In the NP, P, and P + C groups, SBP showed no significant differences (P > .05) during later pregnancy (GDs 16 and 18). A higher SBP (P < .01) was produced by LPS treatment in the LPS-treated group compared to other groups on GD 16, which persisted to GD 18. The effect of choline supplementation on SBP in LPS-induced model (LPS + C group) showed a significantly lower (P < .01) SBP compared to the LPS-treated rats. However, the SBP of the LPS + C group was higher (P < .05) than the P control group.
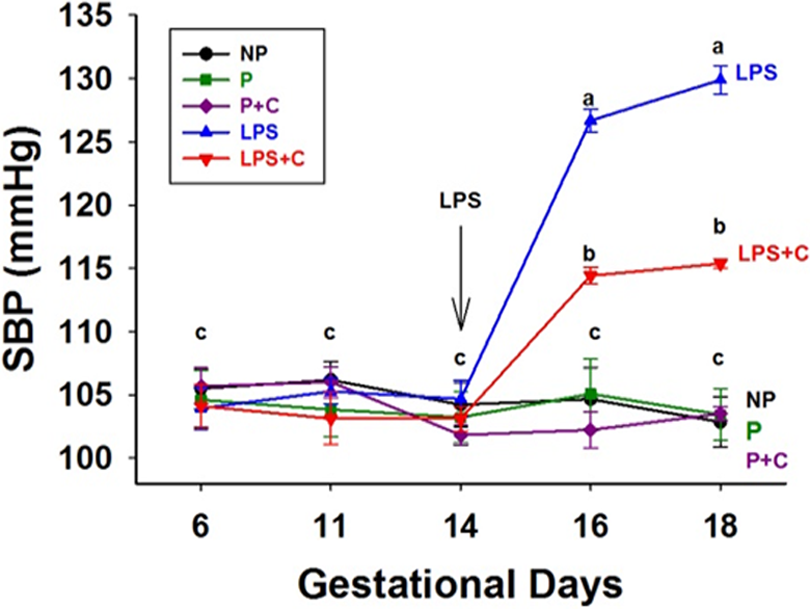
Lipopolysaccharide (LPS) increases and choline supplementation decreases systolic blood pressure (SBP) in LPS-treated rats. Data are expressed as mean (standard error of the mean [SEM]) of the SBP (mm Hg; n = 6-9/group, 2-way analysis of variance). The SBP was measured on gestational days (GDs) 6, 11, 14, 16, and 18 in pregnant (P) group and on corresponding days in nonpregnant (NP) group. Gestational choline supplementation decreased SBP compared to the LPS group on GDs 16 and 18 (P < .01), although it was still higher than other control groups, including P and NP groups. a P < .01, LPS groups on GDs 16 and 18 compared to other groups on the corresponding GDs and prior to the infusion of LPS on GD 14; b P < .05, LPS + choline (C) group compared to the P group.
Figure 2 shows the urinary albumin levels in the various groups. The 24-hour urinary albumin level in NP, P, and P + C groups did not show significant differences on GDs 7 to 19. In the LPS-treated group, LPS administration caused a significant increase (P < .01) in 24-hour urinary albumin levels on GDs 17 and 19. Added gestational choline (LPS + C group) dramatically attenuated the increase in 24-hour urinary albumin level in comparison with the LPS group (P < .01). The levels of serum ALT, AST, BUN, and Cr did not change significantly among groups (data not shown).
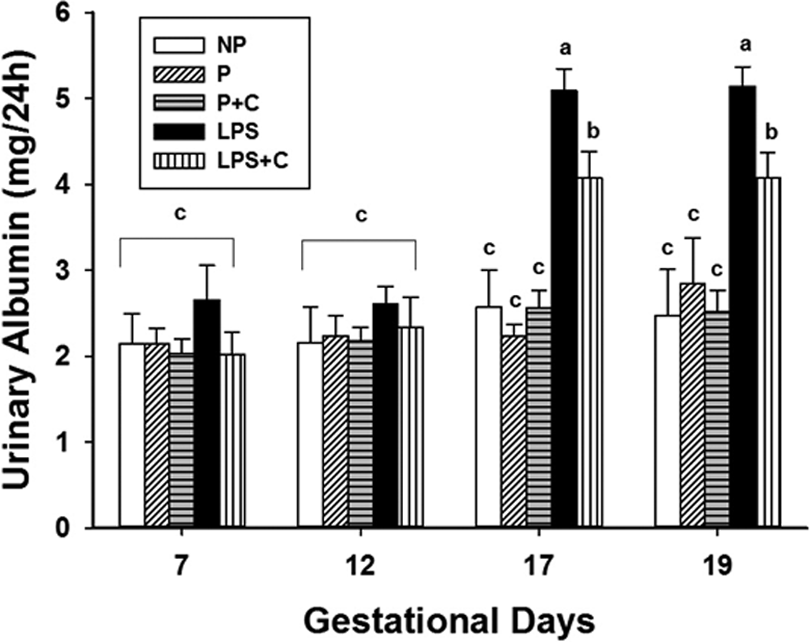
Choline supplementation decreases 24-hour urinary albumin induced by lipopolysaccharide (LPS). Twenty-four-hour urinary protein concentrations on gestational days (GDs) 7, 12, 17, and 19 of pregnancy for the groups are demonstrated in Figure 2. The bar values are mean (standard error of the mean [SEM]) of the 24-hour urinary albumin in each group (n = 6-9/group, 2-way analysis of variance). There were no significant differences among groups on GDs 7 and 12 (P > .05). On GDs 17 and 19, LPS increased urinary protein excretion in the LPS group and gestational choline supplementation significantly decreased urinary protein compared to the LPS group (P < .05). a P < .01, LPS group versus nonpregnant (NP), pregnant (P), and pregnant rats on 5.0 g/kg choline diet (P + C) groups on GDs 17 and 19. b P < .05, LPS + choline (C) group versus LPS group on GDs 17 and 19.
The effects of LPS and choline supplementation on pregnancy outcomes in the various groups are shown in Table 1. Maternal exposure to LPS significantly reduced placenta weights, P = 0.49 g (0.004 g) versus LPS = 0.45 g (0.005 g), P < .05, and fetal weights, P = 4.08 g (0.025 g) versus LPS = 3.76 g (0.042 g), P < .05, and increased the loss of fetuses, P = 2.5% versus LPS = 14.4%, P < .05, compared to the control group. Choline supplementation significantly alleviated LPS-induced reduction in placenta weights, LPS = 0.45 g (0.005 g) versus LPS + C = 0.48 g (0.005 g), P < .05, and fetal weights, LPS = 3.76 g (0.042 g) versus LPS + C = 4.02 g (0.026 g), P < .05, and decreased the loss of fetuses, LPS = 14.4% versus LPS + C = 6.4%, P < .05. Interestingly, choline supplementation during normal pregnancy significantly increased both placental, P = 0.49 g (0.004 g) versus P + C = 0.52 g (0.006 g), P < .05, and fetal, P = 4.08 g (0.025 g) versus P + C = 4.35 g (0.042 g), P < .05, weights. Maternal weights among the 4 groups showed no significant differences (P > .05).
Fetal Loss and Fetal, Placental, and Maternal Weights in Various Groups.a
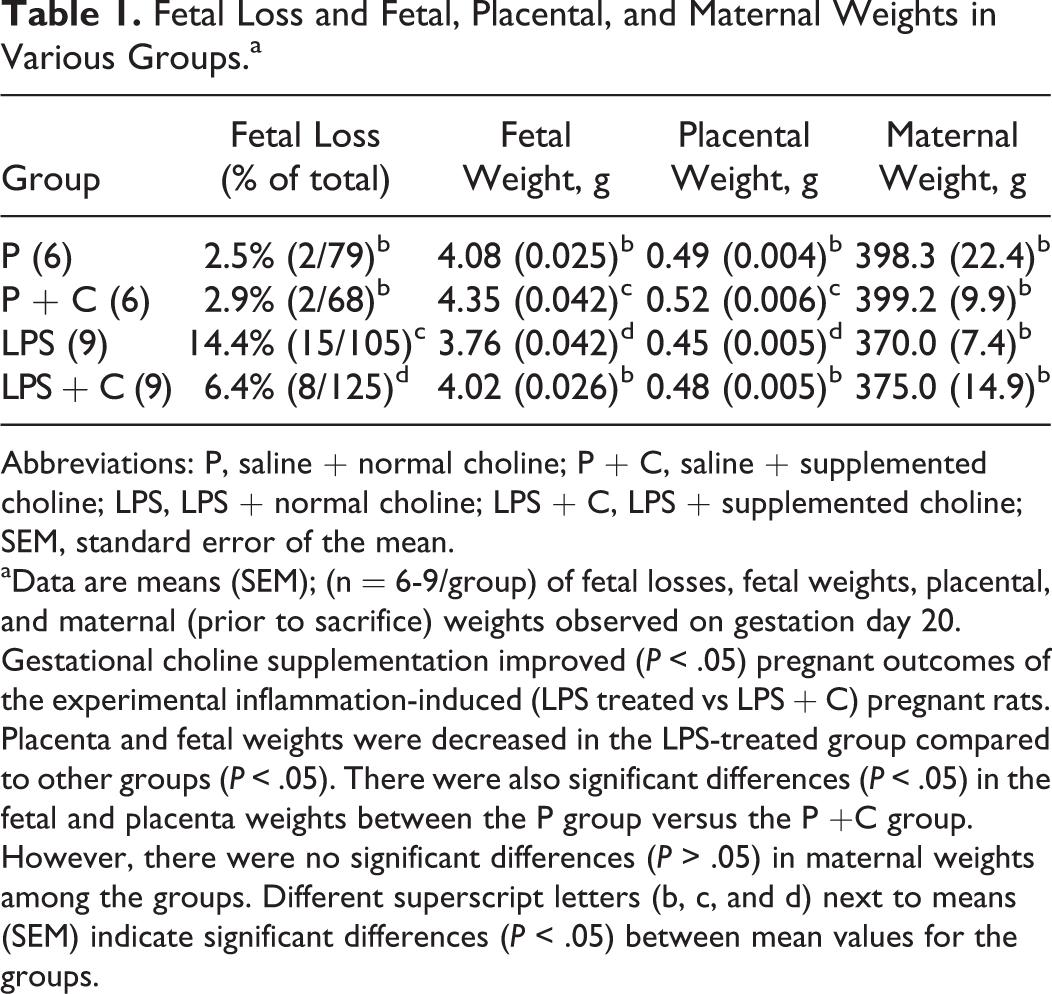
Abbreviations: P, saline + normal choline; P + C, saline + supplemented choline; LPS, LPS + normal choline; LPS + C, LPS + supplemented choline; SEM, standard error of the mean.
aData are means (SEM); (n = 6-9/group) of fetal losses, fetal weights, placental, and maternal (prior to sacrifice) weights observed on gestation day 20. Gestational choline supplementation improved (P < .05) pregnant outcomes of the experimental inflammation-induced (LPS treated vs LPS + C) pregnant rats. Placenta and fetal weights were decreased in the LPS-treated group compared to other groups (P < .05). There were also significant differences (P < .05) in the fetal and placenta weights between the P group versus the P +C group. However, there were no significant differences (P > .05) in maternal weights among the groups. Different superscript letters (b, c, and d) next to means (SEM) indicate significant differences (P < .05) between mean values for the groups.
Effects of LPS and Choline Supplementation on Cytokines
The effects of LPS and choline supplementation during pregnancy on serum pro-inflammatory cytokines in the groups are shown in Figure 3A. Lipopolysaccharide treatment significantly elevated maternal blood levels of TNF-α (P < .01), IL-1β (P < .01), IL-6 (P < .05), IL-17A (P < .01), IFN-γ (P < .05), and GM-CSF (P < .05) compared to controls. In addition, choline supplementation during pregnancy significantly inhibited LPS-induced release of TNF-α (P < .05), IL-1β (P < .05), IL-6 (P < .05), IL-17A (P < .01), and IFN-γ (P < .05), except for GM-CSF. The blood levels of IL-1α, IL-2, IL-5, IL-12P70, IL-13, and G-CSF showed no significant differences (P > .05) among the groups (data not shown). The effects of choline supplementation during pregnancy on placental pro-inflammatory cytokines in LPS rat model were also investigated (Figure 3B). In the placenta, the levels of TNF-α, IL-1β, IL-6, IL-17A, GM-CSF, and IL-2 were significantly (P < .05) increased after LPS administration. Choline supplementation significantly attenuated LPS-induced TNF-α, IL-1β, IL-6, and IL-17A concentrations (P < .05), except for IL-2 and GM-CSF. The placental levels of IL-1α, IL-5, IL-12P70, IL-13, IFN-γ, and G-CSF showed no significant differences (P > .05) among the groups (data not shown). The anti-inflammatory levels of cytokines in maternal serum and placenta for the various groups are shown in Figure 4. The levels of placental (P < .05) and serum IL-4 (P < .01) were significantly decreased after LPS injection and choline treatment significantly attenuated (P < .05) the LPS-induced decrease in maternal serum and placental IL-4 levels. The levels of IL-10 showed no significant differences (P > .05) in serum and placenta among the groups (data not shown).
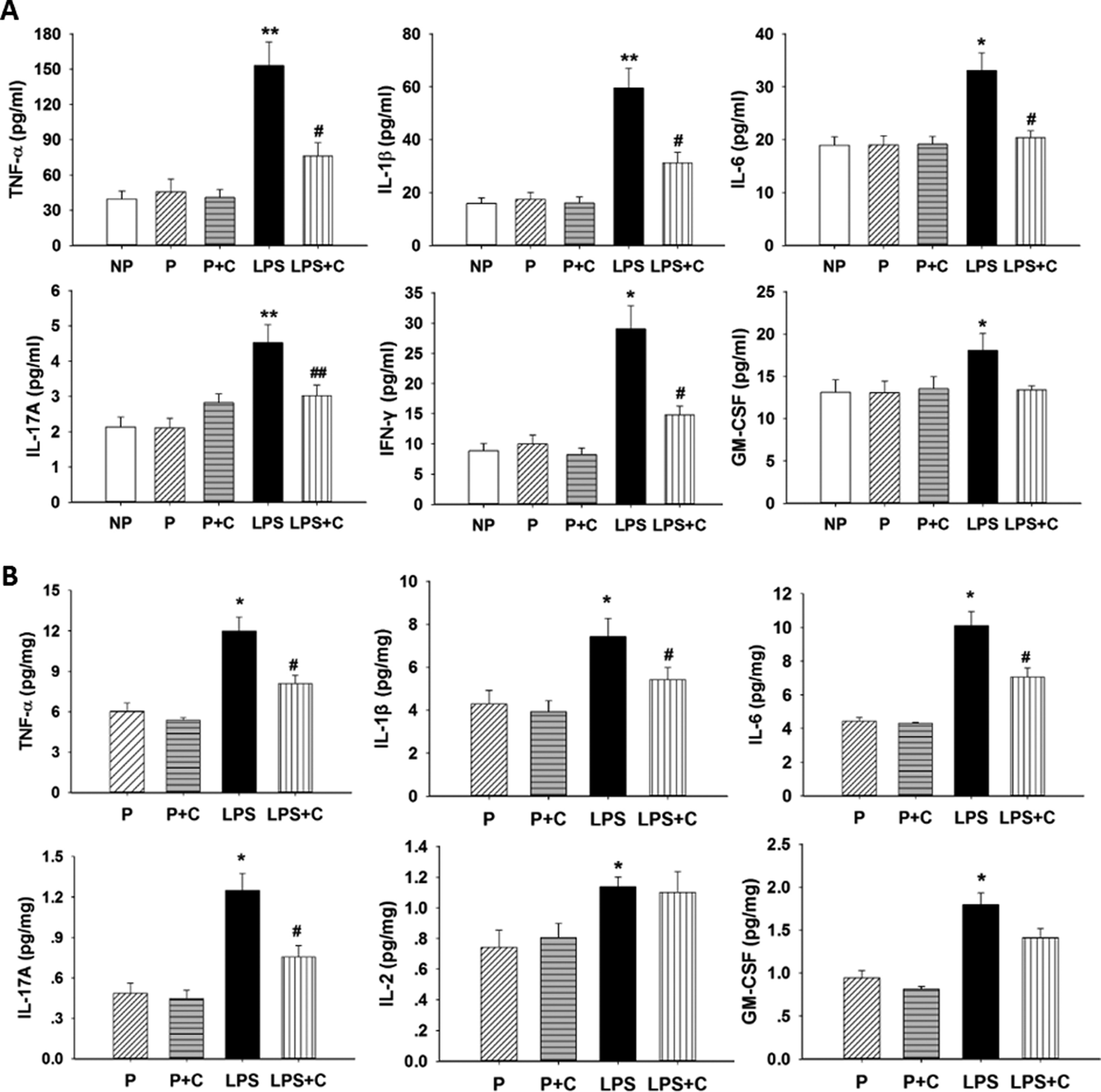
A, Effects of lipopolysaccharide (LPS) and choline supplementation on serum pro-inflammatory cytokines. Data on serum pro-inflammatory cytokines of the groups (expressed as mean (standard error of the mean, n = 6/group) are shown. Serum levels of tumor necrosis factor α (TNF-α; P < .01), interleukin (IL)-1β (P < .01), IL-6 (P < .05), IL-17A (P < .01), interferon γ (IFN-γ; P < .05), and granulocyte-macrophage colony-stimulating factor (GM-CSF; P < .05) collected on gestation day (GD) 20 were significantly increased in the LPS group compared to the pregnant (P) group. Serum levels of TNF-α (P < .05), IL-1β (P < .05), IL-6 (P < .05), IL-17A (P < .01), and IFN-γ (P < .05) were significantly decreased in the LPS + C group compared to the LPS group. *P < .05, LPS group compared to nonpregnant (NP), P, and P + C groups. **P < .01, LPS group compared to NP, P, and pregnant rats on 5.0 g/kg choline diet (P + C) groups. #P < .05, LPS + choline (C) group compared to the LPS group. ##P < .01, LPS + C group compared to the LPS group. B, Effects of LPS and choline supplementation on placental pro-inflammatory cytokines. It shows the effects of LPS and choline supplementation on placental pro-inflammatory cytokines, means (SEM), n = 6/group. Placental levels of TNF-α, IL-1β, IL-6, IL-17A, IL-2, and GM-CSF were significantly increased (P < .05) in the LPS group and levels of TNF-α, IL-1β, IL-6, and IL-17A were significantly decreased (P < .05) in the LPS + C group compared to the LPS group. *P < .05, LPS group compared to the P and P + C groups. #P < .05, LPS + C group compared to the LPS group.
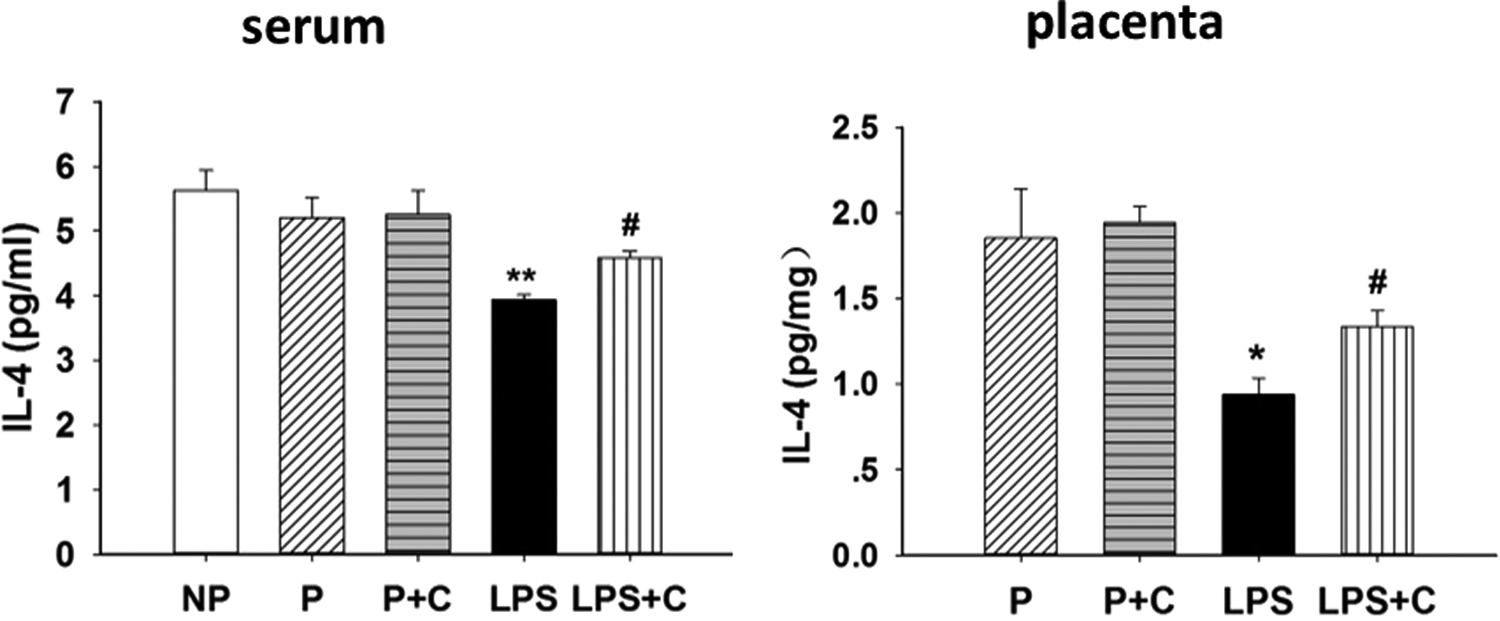
Effects of lipopolysaccharide (LPS) and choline supplementation on interleukin 4 (IL-4), an anti-inflammatory cytokine. Levels of placental and serum anti-inflammatory cytokines, means (standard error of the mean [SEM]), n = 6, on gestation day (GD) 20 are presented. Placental (P < .05) and serum levels (P < .01) of IL-4 were significantly decreased in the LPS group compared to the pregnant (P) group. In contrast, placental and serum levels of IL-4 were significantly increased (P < .05) in the LPS + choline (C) group compared to the LPS group. *P < .05, LPS group compared with P and pregnant rats on 5.0 g/kg choline diet (P + C) groups. **P < .01, LPS group compared to nonpregnant (NP), P, and P + C groups. # P < .05, LPS + C group compared to the LPS group.
Effects of LPS and Choline Supplementation on the CAP
The effects of LPS and choline supplementation on α7nAChR protein expression in placenta in LPS-induced rat model are demonstrated in Figure 5. There was a significant increase (P < .05) in α7nAChR expression in the LPS group compared to the control group. Choline supplementation significantly decreased (P < .05) the level of α7nAChR expression in the LPS + C group compared to the LPS group, but choline did not increase the levels of α7nAChR in the P + C group compared to the P group (P > .05).
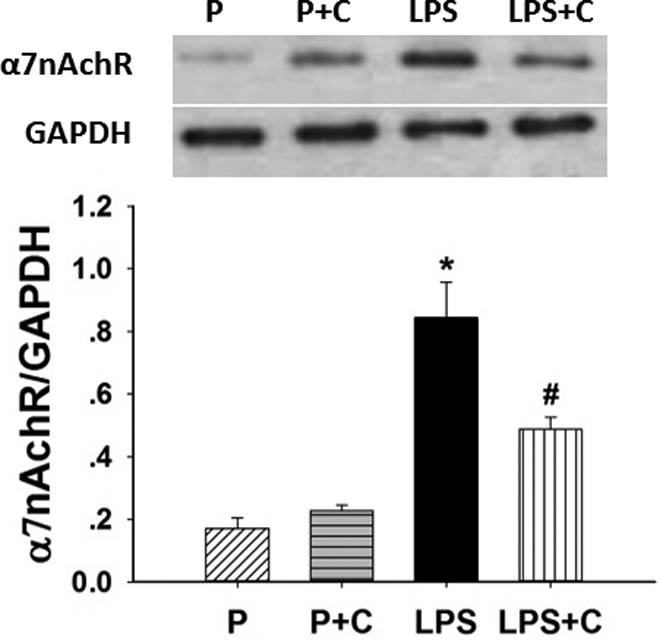
Choline supplementation decreases placental α7 nicotinic acetylcholine receptor (α7nAChR) expression. Western blots showing the differential protein expression of placental α7nAChR among the 4 groups on gestational day (GD) 20. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control to show loading of protein. The ratio of α7nAChR–GAPDH expression was used to normalize protein levels in the blots (bottom of the figure). There was a significant increase (P < .05) in α7nAChR expression in the lipopolysaccharide (LPS) group compared to the pregnant (P) and pregnant rats on 5.0 g/kg choline diet (P + C) groups. Choline treatment dramatically decreased α7nAChR expression in the LPS+ choline (C) rats (P < .05) compared to the LPS-treated rats. *P < .05, LPS group compared to P and P + C groups, #P < .05, LPS + C group compared to the LPS group. The values are means (standard error of the mean [SEM]) of 6 samples per group.
To investigate whether maternal LPS injection activates NF-κB in placenta, the levels of pIκBα, IκBα, and NF-κB p65 were measured by Western blotting (Figures 6A and B). Although the levels of placental IκBα showed no significant difference (P > .05) among the groups, maternal exposure to LPS significantly increased (P < .01) the level of pIκBα in placenta, which may indicate that placental IκBα was degraded after maternal LPS exposure. The LPS-induced placental IκBα phosphorylation was attenuated (P < .05) by gestational choline supplementation. Western blotting also indicates that the level of placental nuclear NF-κB p65 was significantly increased (P < .05) in rats with LPS treatment (Figure 6B), and this was decreased (P < .05) by choline supplementation.
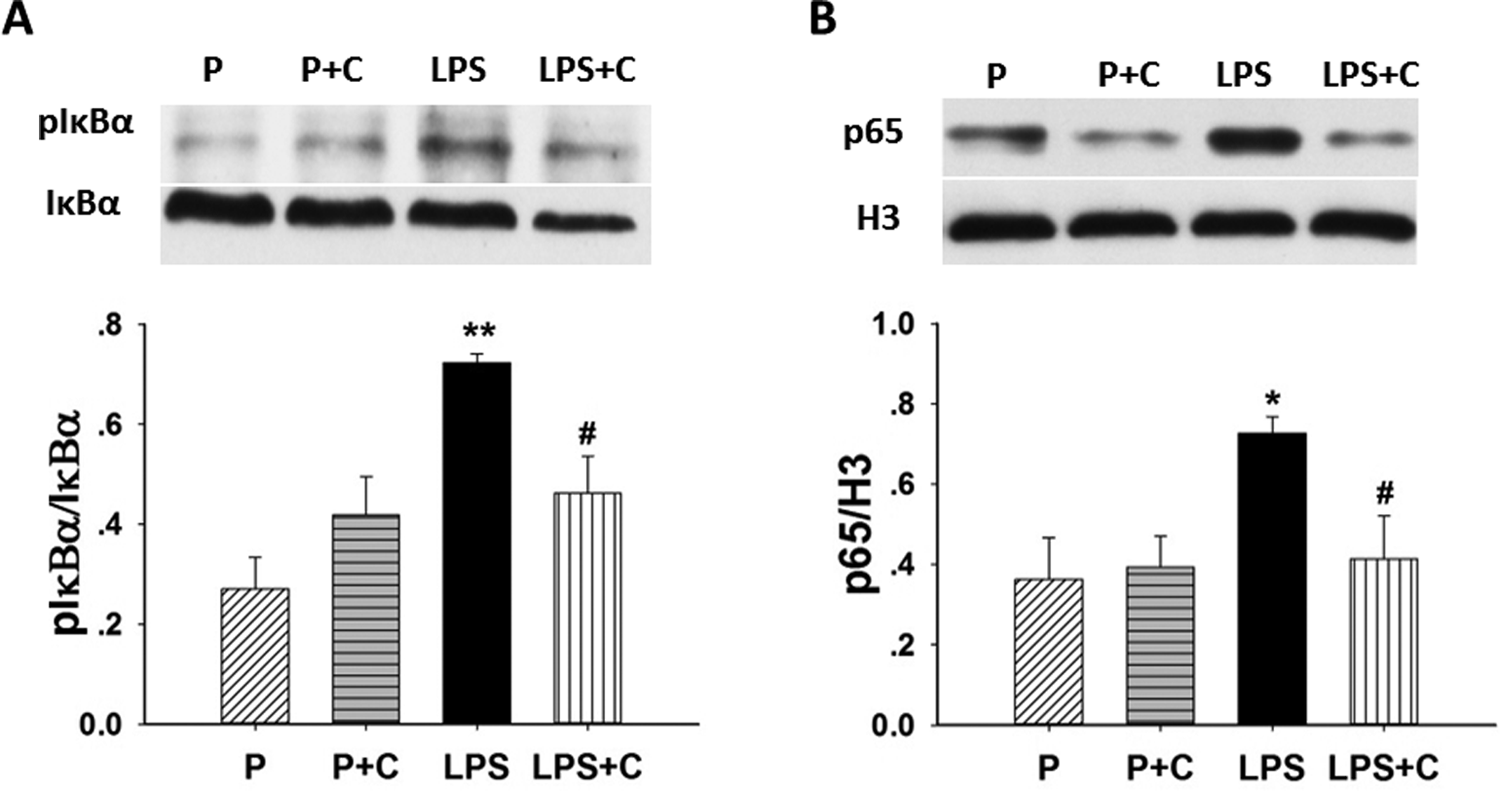
Choline inhibits activation of placental nuclear factor κB (NF-κB). Western blots showing the differential protein expression of placental NF-κB activation, including phosphorylation of IκBα and p65, among the 4 groups on day 20 of gestation. A, Placental IκBα and pIκBα were measured using Western blotting. Representative gels for pIκBα and IκBα and ratios of pIκBα–IκBα (lower panel) are shown. B, The nuclear fractions were prepared from placenta. Representative gels for p65 and H3 ratios of p65–H3 (lower panel) are shown. There were significant increases in expression pIκBα (P < .01) and NF-κB p65 (P < .05) in the lipopolysaccharide (LPS) group compared to the pregnant (P) and pregnant rats on 5.0 g/kg choline diet (P + C) groups. Added gestational choline dramatically decreases (P < .05) pIκBα and NF-κB p65 expression of LPS-treated rats. *P < .05, LPS group compared to the P and P + C groups. **P < .01, LPS group compared with the P and P + C groups. #P < .05, LPS+ choline (C) group compared to the LPS group. The values are means (standard error of the mean [SEM]) of 6 samples per group.
Figure 7A to D shows representative photographs of immunocytochemical studies of nuclear translocation of NF-κB p65 in the various groups. NF-κB p65 was mainly localized in the mononuclear sinusoidal trophoblast giant cells of the labyrinth zone. Lipopolysaccharide treatment increased the localization of placental mononuclear staining (Figure 7C). Further, choline supplementation significantly inhibited LPS-induced nuclear translocation of NF-κB p65 in the giant cells (Figure 7D).
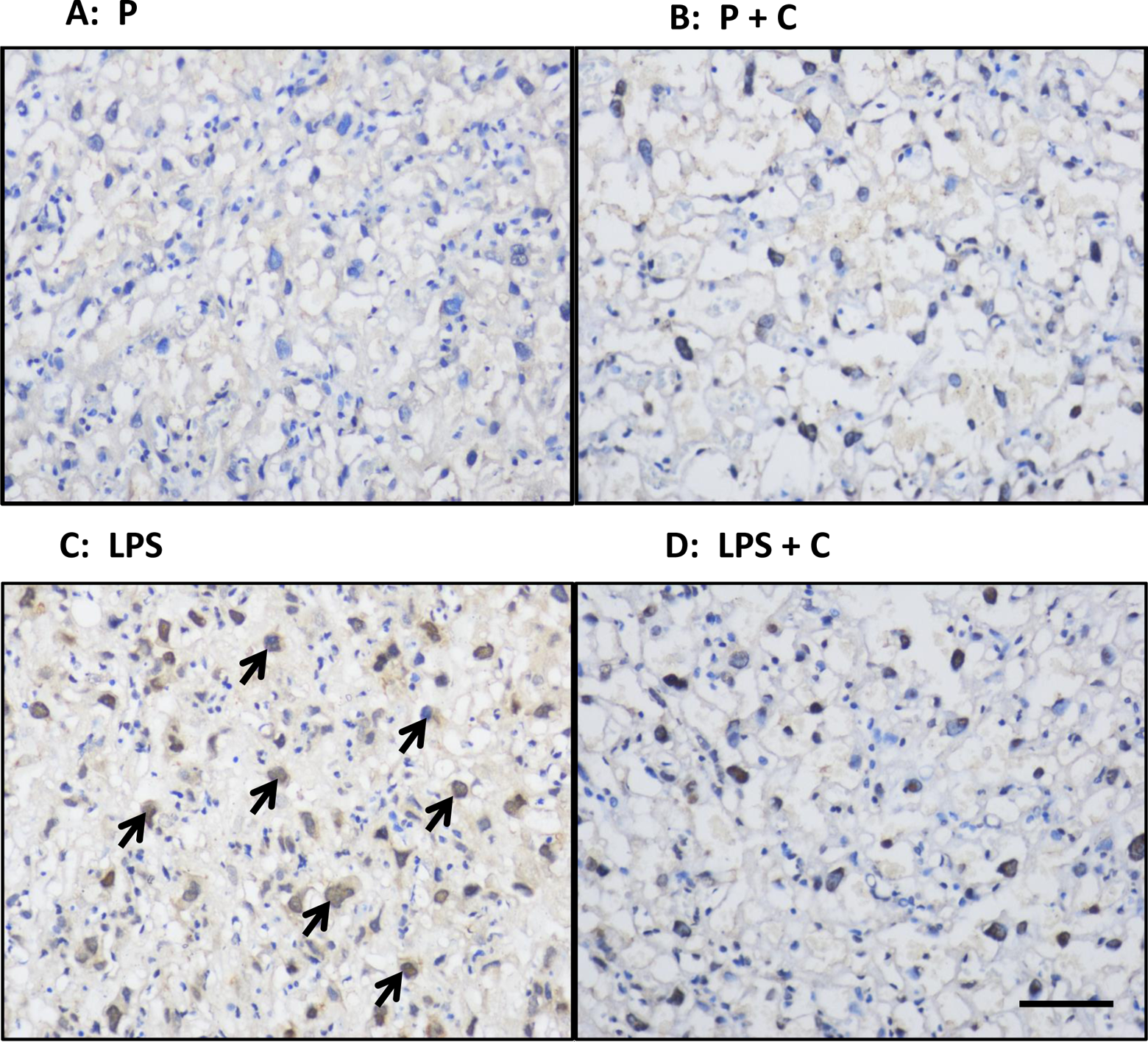
Effects of lipopolysaccharide (LPS) and choline (C) on nuclear translocation of nuclear factor κB (NF-κB) p65 in the placenta. A-D, Representative photomicrographs of placental localization of NF-κB p65 in the nuclei of mononuclear sinusoidal trophoblast giant cells of the labyrinth zone of the various groups using immunohistochemistry in tissues collected on gestation day (GD) 20. Shown are placental photos from rats treated with (A) saline + normal choline (P group); (B) saline + supplemented choline (P + C); (C) LPS + normal choline (LPS); (D) LPS + supplemented choline (LPS + C). Scale bar on right bottom of Figure 7D = 120 μm for all photos. Nuclear location of NF-κB p65 was most prominent in cells from the LPS-treated group (arrows).
The effects of choline on AKT phosphorylation in placenta in control and LPS rat model are shown in Figure 8. No significant difference (P > .05) was found in the LPS group compared to the P control group. In addition, choline supplementation did not affect (P > .05) the level of AKT in placenta. However, the level of pAKT was significantly increased (P < .01) in the placenta of rats administered LPS. Choline supplementation dramatically inhibited (P < .05) LPS-induced AKT phosphorylation.
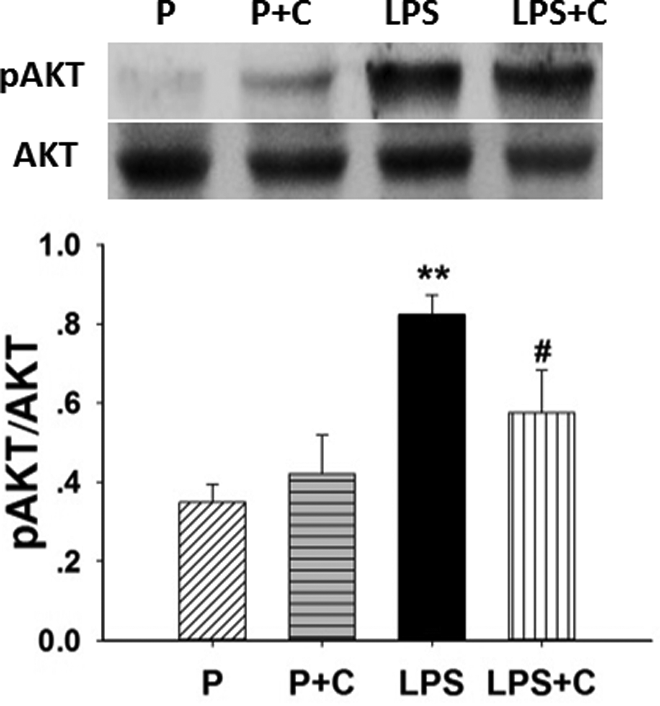
Choline inhibits phosphorylation of placental protein kinase B (AKT). Western blots showing the differential protein expression of placental AKT phosphorylation activation among the 4 groups on gestation day (GD) 20. Shown are mean values and standard error of the mean (SEM) from 6 samples/group. Placental AKT (standard marker) and phosphorylated AKT (pAKT) were measured using Western blotting. Representative gels for pAKT and AKT are shown (top of the figure). Expression of pAKT/AKT was significantly increased (P < .01) in the lipopolysaccharide (LPS) group compared to the pregnant (P) and pregnant rats on 5.0 g/kg choline diet (P + C) groups (bottom of the figure). Gestational choline supplementation dramatically decreased (P < .05) pAKT expression of LPS-treated rats. **P < .01, LPS group compared with the P and P + C groups. #P < .05, LPS + C group compared to the LPS group. The values are means (SEM) of 6 samples per group.
Discussion
This study clearly demonstrates the effects of LPS and added choline administration during pregnancy. Lipopolysaccharide treatment induces inflammatory conditions that are mostly reversed by choline supplementation, and this protects rats from inflammatory symptoms, such as adverse fetal outcomes (fetal loss and growth restriction), hypertension, and proteinuria. Moreover, choline supplementation inhibits inflammatory responses in LPS-treated rats by decreasing placenta α7nAChR, steps in the CAP placental pathways (namely, activation of NF-κB and localization in mononuclear cells and AKT phosphorylation), and serum and placenta pro-inflammatory cytokines. These observations suggest that added gestational choline treatment reverses, at least partially, inflammation by suppression of steps in the CAP and thereby has positive therapeutic effects in the inflammatory model. The results of this study indicate that additional choline in the diet or treatment with other agonists to the α7nAChR may be an effective clinical strategy for treatment of inflammatory conditions associated with pregnancy such as PE and infection-related preterm birth.
Previous studies 12 –14,17 demonstrate that LPS treatment produces inflammatory responses in animal models which are associated with elevated levels of cytokines. Previously, we have also demonstrated that nicotine decreases LPS-induced increases in cytokines and reverses increases in LPS-induced symptoms in rats 12,15,17 and decreases fetal loss. In this study, we show that added choline in the diet, like injections of nicotine, reduces inflammatory conditions produced by LPS and decreases adverse symptoms and also decreases fetal loss. Therefore, we conclude that choline may be a very important way to reverse pregnancy-associated inflammation and a variety of adverse responses in a clinical setting. Exactly why choline treatment did not completely inhibit the LPS-stimulated increase in many of inflammatory symptoms and the steps involved in the CAP may be due to the dose of choline available for our study or the weak agonistic activity of choline. Perhaps cholinergic agonists with higher affinity for the α7nAChR, such as carbachol or GTS-1, or agents that achieve greater concentrations at the receptor would be more effective in reversing the effects of LPS and be more appropriate for clinical treatments.
According to this study, LPS treatment, representing a manifestation of endotoxin and infection, produces signs and symptoms similar to PE (ie, increased blood pressure, decreases in fetal and placenta weights, and proteinuria). However, there are no data to suggest that PE in women is induced by bacterial endotoxin, and the diagnosis of PE is no longer dependent upon the presence of proteinuria. 36 Lipopolysaccharide has been used in this and other studies as a model system for infection-induced preterm birth in animals, 9,11,12 but the findings that LPS also increases blood pressure and proteinuria may be due to the effects of the toxin on the vasculature and kidneys, which may be irrelevant to the glomerular endotheliosis of the clinical manifestation of PE. In any event, our study demonstrates that choline supplementation alleviates intravenous LPS-induced increases in blood pressure and proteinuria, pro-inflammatory gene expression, and adverse fetal outcomes. Surprisingly, our results also suggest that choline in the absence of LPS significantly increased fetal and placental weights and LPS in the absence of choline stunted fetal and placental weights. These findings may be attributed to increased perfusion of the placenta and fetuses by choline and a decreased perfusion by LPS. There were no differences in maternal weights between the groups suggesting that choline does not increase fetal and placenta weights, at least in the LPS untreated groups where fetal loss is not apparent, by increasing the maternal weight.
Cholinergic stimulation with agents such as nicotine, GTS-21, and choline are considered to be an effective treatment for the overproduction of pro-inflammatory cytokines which result in deleterious effects in P and NP diseases. 37,38 An early report illustrated that choline administration reduces TNF-α levels and modulated the redox status of bronchoalveolar cells to ameliorate oxidative stress in a mouse model. 33 A cross-sectional survey reported that healthy adults with higher choline and betaine intakes showed lower concentrations of C-reactive protein, IL-6, and TNF-α. 39 Numerous studies revealed that maternal LPS exposure during pregnancy stimulates the production of pro-inflammatory cytokines, which exerts part of a severe systemic inflammatory response. 17,40 Other studies also showed that pregnancy-induced hypertension, such as PE, is associated with increased maternal circulating pro-inflammatory factors, including TNF-α, IL-6, and IL-8, as well as enhanced expression of TNF-α in placentas and increased IL-1β, IL-6, and IL-8 in peripheral monocytes. 41 The present study illustrates that LPS treatment significantly elevates maternal blood and placenta levels of several pro-inflammatory cytokines and choline supplementation during pregnancy significantly inhibited them. These observations indicate that choline mediates a protection against LPS-induced inflammation, and this can be attributed to its anti-inflammatory effects.
Both IL-10 and IL-4 are important anti-inflammatory cytokines, and we and others had demonstrated that IL-4 and IL-10 in peripheral blood are significantly decreased in PE conditions. 16,40,42,43 We have also found that treatment with nicotine statistically increased the level of the maternal serum IL-4. 16 The finding in this study that choline supplementation significantly increases the level of serum and placenta IL-4 suggests that IL-4 might play an important role in choline-mediated protection against LPS-induced inflammation.
Many reports suggest that the CAP can suppress the overproduction of pro-inflammatory cytokines. 44 Accumulating evidence has pointed out that α7nAChR plays an essential role in CAP. The α7nAChR has been found in the nervous system, such as the hippocampus, and has also been reported in human and rat placenta. 45 Dietary choline supplementation has also been reported to be neuroprotective and have potential use in the treatment or prevention of brain neurodegeneration by acting as a selective activator of α7nAChR. 46 Long-term improvements in sensory inhibition with gestational choline supplementation has been linked to α7 nicotinic receptors. 47 Maternal LPS exposure resulted in an increase in α7nAChR expression and choline supplementation significantly decreases the level of α7nAChR expression indicating the anti-inflammatory effects of choline. The present study also demonstrates that choline supplementation inhibits many of the steps involved in the CAP pathway and thus accounts for the reversal of symptoms and changes in cytokines seen with added choline treatment. Steps in the CAP have been thoroughly reviewed. 44,48 Exactly how LPS and choline function to produce their effects is not clear, but an increase in the α7nAChR by LPS and a decrease in the receptor by choline may account for most of the other changes in the intermediate steps resulting in altered cytokine levels.
Nuclear factor-κB plays a central role in LPS-induced upregulation of inflammatory genes. In unstimulated cells, NF-κB resides in the cytoplasm as an inactive NF-κB–IκBα complex. In the activation condition, I-κB phosphorylation causes translocation of NF-κB to the nucleus and then activates the transcription of various inflammatory gene products. There is increasing evidence that cholinergic stimulation can inhibit cytokines by inhibiting the NF-κB pathway through an α7nAChR-dependent anti-inflammatory pathway. 49,50 An early study showed that NF-κB p65 was involved in the effects of choline supplementation on reducing oxidative stress in a mouse model of allergic airway disease. 33 We found that the levels of phosphorylated IκBα and nuclear NF-κB p65 were significantly increased in the placenta of rats treated with LPS. Moreover, nuclear translocation of NF-κB p65 was mainly observed in mononuclear sinusoidal trophoblast giant cells of the placenta. These observations are similar to a study of nuclear translocation of NF-κB p65 in the placenta of a LPS-induced fetal growth restriction and demise mice model. 51 Interestingly, these effects were reversed by choline supplementation in this study. These observations support our contention that cholinergic stimulation attenuates LPS-induced elevation in placental nuclear NF-κB p65. Protein kinase B serine–threonine kinase is thought to be involved in the activation of NF-κB. 52 Also, maternal LPS exposure results in placental AKT phosphorylation. 51 A previous report demonstrated that GTS-21 suppressed the inducible activation of NF-κB by inhibiting the PI3K/AKT pathway in LPS-induced inflammatory cytokine production in RAW264.7 cells. 53 Importantly, the present study shows that choline supplementation during pregnancy dramatically inhibits LPS-induced placental AKT phosphorylation. These results indicate that choline-mediated repression of LPS-induced placental NF-κB activation might be associated with the inhibition of AKT phosphorylation. To our knowledge, this is the first report to investigate the protected effects of gestational choline supplementation on LPS-induced inflammatory responses in vivo through the α7nAChR and CAP. Our previous study on effects of nicotine supports the view that the CAP may be useful to improve the manifestation of inflammatory responses. However, nicotine has well-known side effects and therefore nicotine may not be the ideal treatment for clinical treatment of inflammation. Choline may be a much better therapeutic choice for the treatment of inflammation because choline is an essential nutrient and it plays an important role in reducing the risk of neural tube defects, fatty liver disease, and other pathologies. In addition, unlike nicotine, choline is safe, orally effective, and has few side effects if an appropriate dose is used. There are a number of reports that demonstrate beneficial perinatal effects of choline supplementation in rodent models, from improved learning and memory to protection from the behavioral effects of fetal alcohol exposure acting through the α7nAChR. 46,47 In addition, supplementation of maternal choline in rodents during pregnancy significantly yields lasting improvements in offspring cognitive functioning and reduces vulnerability to mammary tumors in the female offspring. 35,54 Thus, choline not only has protective effects against inflammation during pregnancy as shown here, but it has other valuable effects during pregnancy.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by Science and Technology Program of Guangdong Province, China, to Huishu Liu (Grant #2016A020218002); Natural Science Foundation of Guangdong Province, China, to Huishu Liu (Grant #2014A030313722); Innovation of Science and Technology Program of Guangzhou, China, to Junjie Bao (Grant #172201607010315).