
Abstract
Introduction:
Obesity is an epidemic affecting more than one-third of adults in the United States. Obese women experience decreased fertility, explained in part by oocyte quality. Since follicular fluid (FF) provides an important microenvironment for oocyte growth, we sought to evaluate the effect of increased body weight on FF levels of 11 metabolic hormones and fatty acid metabolism.
Methods:
The FF was collected from 25 women (10 normal weight, 10 overweight, and 5 obese) with diminished ovarian reserve undergoing in vitro fertilization (IVF) following a minimal stimulation protocol. Hormone levels were determined by multiplex immunoassay using the MAGPIX (Luminex, Austin, Texas) instrument. Fatty acid metabolites were determined using gas and ultra-high pressure liquid chromatography coupled with mass spectrometry.
Results:
Levels of hormones related to glucose and energy homeostasis and regulation of fat stores (insulin, glucagon, glucagon-like peptide-1, C-peptide, and leptin) were increased significantly in FF from obese women compared to FF from nonobese(normal weight and overweight) women. Interestingly, FF levels of branched-chain amino acids (BCAA) isoleucine, leucine, and valine as well as uric acid, isocaproic acid, butanoic acid, tyrosine, threonine, glycine, and methionine correlated positively with body mass index.
Conclusion:
This pilot study demonstrates significant alterations in the FF milieu of obese women undergoing IVF, which may contribute to the decreased fecundity of obese women. Although the impact of this environment on oocyte and embryo development is not fully realized, these changes may also lead to imprinting at the genomic level and long-term sequelae on offspring.
Keywords
Introduction
Obesity is an epidemic affecting more than one-third of all adults in the United States. Two-thirds of all women and 50% of reproductive-age women are either overweight or obese. 1
In addition to the long list of medical complications of excess body weight, there are significant consequences on reproductive health. Obese women have higher rates of ovulatory dysfunction, decreased ovarian response to ovulation induction, altered oocyte and endometrial function, and children with lower birth rates. Interestingly, even in the setting of normal ovulation, obese women can still experience decreased fecundity, and this can be partly explained by the decrease in oocyte quality. 2 –4
Follicular fluid (FF) provides an important microenvironment for the developing oocyte. It is a product of both granulosa and theca cells secretions as well as the transfer of plasma components across the blood–follicular barrier. 5 The biochemical characteristics of the FF surrounding the oocyte play a critical role in determining oocyte quality and the subsequent potential to achieve fertilization and embryo development. 6
The objective of this pilot study was to evaluate the metabolic characteristics of FF as a function of body weight in women undergoing in vitro fertilization (IVF). Considering the extensive involvement of metabolic hormones in ovarian follicle and oocyte development, as well as hormone production and secretion, we sought to investigate the impact of body weight on FF concentrations of 11 metabolic hormones and fatty acid and amino acid metabolites. An evaluation of the available literature and the pathways of lipid metabolism lead us to select insulin, glucagon, leptin, ghrelin, and adiponectin as hormones of interest in the FF. The metabolic hormones kit commercially available to test for these hormones also tests for the remaining 6 hormones; hence, all these results are reported in this study.
Materials and Methods
This study was conducted at the Fertility and Advanced Reproductive Medicine clinic of University of Texas Southwestern in Dallas, Texas, during the period of January 2016 to July 2017. Institutional Review Board exemption was granted (reference STU 012012-235). Informed consent was obtained from all individual participants included in the study. We recruited 25 patients: 10 normal weight [body mass index (BMI) 18.5-24.9 kg/m2], 10 overweight (BMI 25-29.9 kg/m2), and 5 obese women (BMI ≥30 kg/m2). All participants in this study were comprised of women with diminished ovarian reserve as defined by the Bologna criteria 7 undergoing IVF following our minimal stimulation protocol, aged 35 to 45, and BMI of 18.5 to 40 kg/m2. All embryos were frozen in preparation for transfer in another cycle. Exclusion criteria included the following: patients with associated medical illnesses (eg, diabetes, hypothyroidism, malignancy, hypercortisolism, arthritis, or inflammatory diseases), polycystic ovary syndrome (PCOS), or endometriosis.
Minimal stimulation protocols were conducted as described previously. 8 Briefly, clomiphene citrate (100 mg daily) was administered for 10 consecutive days starting on cycle day 3. Human menopausal gonadotropin (HMG; 150 units) was added every other day starting on cycle day 5. Luteinizing hormone (LH) levels were followed throughout the cycle, and GnRH antagonist was administered as needed if LH levels were >5 IU/L. Patients received a dual trigger shot of leuprolide acetate (2 mg) and human chorionic gonadotropin (5000 U) when the lead follicle reached 18 mm. Oocyte retrieval took place 35 hours after trigger. This type of “low-intensity” stimulation lacks many of the unwanted characteristics of high-dose gonadotropin stimulation protocols typically used in IVF cycles of similar patient populations, such as a possible increase in aneuploidy rate. 9 –11 During oocyte retrieval, FF was collected using a 17G Cook single lumen oocyte retrieval needle (Brisbane, Australia) under suction. The first follicle to be collected was the largest, and after the oocyte was isolated from the FF under microscopic guidance, FF from this follicle was processed for analysis. The remaining FF was discarded after the oocytes were isolated. The FF processing took place within 1 hour of collection. All patients with grossly bloody FF were excluded from the study. The FF was centrifuged at 1000×g for 10 minutes after which the supernatant was isolated and kept at −80°C until time of analysis.
Analysis of FF for Metabolic Hormones
Concentrations of insulin, gastric inhibitory polypeptide (GIP), glucagon, visfatin, ghrelin, glucagon-like peptide-1 (GLP-1), plasminogen activator inhibitor-1 (PAI-1), resistin, C-peptide, and leptin were assayed using the Bio-Plex Pro human diabetes 10-plex immunoassay (171A7001M; Bio-Rad Laboratories, Hercules, California). The concentration of adiponectin was assayed using Bio-Plex Pro human diabetes adiponectin assay (171A7003M, Bio-Rad Laboratories). Following the manufacturer’s instructions, a 96-well plate was initially prewet with washing buffer. After the buffer was discarded, microsphere magnetic beads coated with monoclonal antibodies against the specific target analytes were added to the wells. Samples, blank, and standards were added to the wells and incubated at room temperature on a shaker at 850 rpm for 60 minutes. Wells were washed using a magnetic manifold, and detection antibodies were added. After incubation for 30 minutes, streptavidin-PE was added to the beads and incubated for 10 minutes. Adiponectin samples were diluted 100-fold prior to testing. This dilution was taken into consideration when values were entered in the MAGPIX instrument (Luminex, Austin, Texas, USA). Measurements were performed using the Bio-Plex MAGPIX Multiplex Reader (Bio-Rad Laboratories). Inter- and intra-assay coefficients of variations for the assays ranged between 2% and 6%. 12
Metabolite extraction and detection using gas and ultra-high-pressure liquid chromatography coupled to mass spectrometry (GC [Gas chromatography–mass spectrometry]- and UHPLC-MS[Ultra performance liquid chromatography - mass spectrometry])
Sample Preparation
Sample volume varied from 125 μL to 1 mL. Extraction solvent was normalized to the sample volume assayed. Methyl-tert-butyl ether (MTBE) biphasic extraction was performed. To a 400-μL sample, for example, 1.2 mL of methanol (MeOH) was added to form a 75% MeOH (25% of water was from the sample) and then mixed with 2.4 mL of MTBE for the first extraction. After 20 minutes of vortexing at 4°C, 1 mL of water was added to the monophase of MTBE-MeOH-water to induce phase separation. The sample was centrifuged after a brief vortexing at 3250g and 4°C for 10 minutes. The upper organic layer was transferred to a glass vial, to be used for lipid profiling on a UPLC-MS analysis. The lower aqueous phase was transferred to a second glass vial, to be used for GC-MS analysis of polar small molecules.
Data Acquisition
GC/MS
Aqueous extract (1.6 mL) was dried under nitrogen, resuspended in 50 μL of pyridine containing 25 mg/mL of methoxyamine hydrochloride, incubated at 60°C for 45 minutes, vigorously vortexed for 30 seconds, sonicated for 10 minutes, and incubated for an additional 45 minutes at 60°C. Next, 50 μL of N-methyl-N-trimethylsilyltrifluoroacetamide with 1% trimethylchlorosilane (MSTFA + 1% TMCS; Thermo Scientific, Waltham, Massachusetts, USA ) was added, and the samples were vigorously vortexed for 30 seconds and incubated at 60°C for 30 minutes. For samples having less than 1.6 mL of aqueous layer, the volume of derivatization reagents was reduced proportional to the sample volume.
Metabolites were detected using a Trace 1310 GC coupled to a Thermo ISQ mass spectrometer. Samples (1 μL) were injected at a 40:1 split ratio to a 30 m TG-5MS column (Thermo Scientific; 0.25 mm id, 0.25 μm film thickness) with a 1.2 mL/min helium gas flow rate. The GC inlet was held at 285°C. The oven program started at 80°C for 30 seconds, followed by a ramp of 15°C/min to 330°C, and an 8 minutes hold. Masses between 50 and 650 m/z were scanned at 5 scans/sec under electron impact ionization. Transfer line and ion source were held at 300°C and 260°C, respectively. Pooled QC samples were injected after every 6 actual samples.
UHPLC/MS
Aqueous extract of 2 mL was dried under nitrogen gas and resuspended in 100 μL of toluene/methanol (1/1, v/v). Extract of 2 μL was injected onto a Waters Acquity UPLC system in discrete, randomized blocks with a pooled QC injection after every 6 sample injections and separated using a Waters Acquity UPLC CSH Phenyl Hexyl column (1.7 μmol/L, 1.0 × 100 mm), using a gradient from solvent A (2 mmol/L ammonium hydroxide, 0.1% formic acid) to solvent B (acetonitrile, 0.1% formic acid). Injections were made in 100% A, held at 100% A for 1 minute, ramped to 98% B over 12 minutes, held at 98% B for 3 minutes, and then returned to starting conditions over 0.05 minutes and allowed to reequilibrate for 3.95 minutes, with a 200 μL/min constant flow rate. The column and samples were held at 65°C and 6°C, respectively. The column eluent was infused into a Waters Xevo G2 Q-TOF-MS with an electrospray source in positive mode, scanning 50 to 2000 m/z at 0.2 seconds per scan, alternating between MS (6 V collision energy) and MSE mode (15-30 V ramp). Calibration was performed using sodium iodide with 1 ppm mass accuracy. The capillary voltage was held at 22.
For each sample, raw data files were converted to .cdf format, and matrix of molecular features as defined by retention time and mass (m/z) was generated using XCMS software in R for feature detection and alignment. Raw peak areas were normalized to total ion signal in R, and outlier injections were detected based on total signal and PC1 of principal component analysis. Features were grouped based on a novel clustering tool, RAMClustR, which groups features into spectra-based coelution and covariance across the full data set, whereby spectra are used to determine the identity of observed compounds in the experiment. The peak areas for each feature in a spectrum were condensed via the weighted mean of all features in a spectrum into a single value for each compound. Compounds were annotated based on spectral matching to in-house (for LC–MS experiments only), National Institute of Standards and Technology MS databases (NISTv14), and Golm metabolite databases (for GC-MS experiments only).
Statistics
Data are expressed as mean ± standard error of mean. Patients and cycle characteristics in the 3 groups were analyzed using ANOVA for continuous variables and the Kruskal-Wallis test for ordinate data. Differences between levels of insulin, GIP, glucagon, visfatin, ghrelin, GLP-1, PAI-1, resistin, C-peptide, leptin, and adiponectin were determined using the Student t test comparing 2 groups (obese vs nonobese).
For GC-MS and UPLC-MS, ANOVA for 3 groups (normal weight, overweight, and obese) was conducted on each compound using the aov function in R, and P values were adjusted for false positives using the Bonferroni-Hochberg method in the P.adjust function in R. PCA was conducted on mean-centered and Pareto variance-scaled data using the pcaMethods package in R. Differences were considered to be significant at P < .05.
Results
Patients and Cycle Characteristics
We recruited 25 patients at our IVF center: 10 normal weight, 10 overweight, and 5 obese women. Patient and cycle characteristics are represented in Table 1. In brief, mean age was 40.5 years, mean AMH was 1.17 ng/mL, and mean HbA1C was 5.47%. All patient and cycle characteristics were similar among groups except BMI.
Patients and Cycle Characteristics.
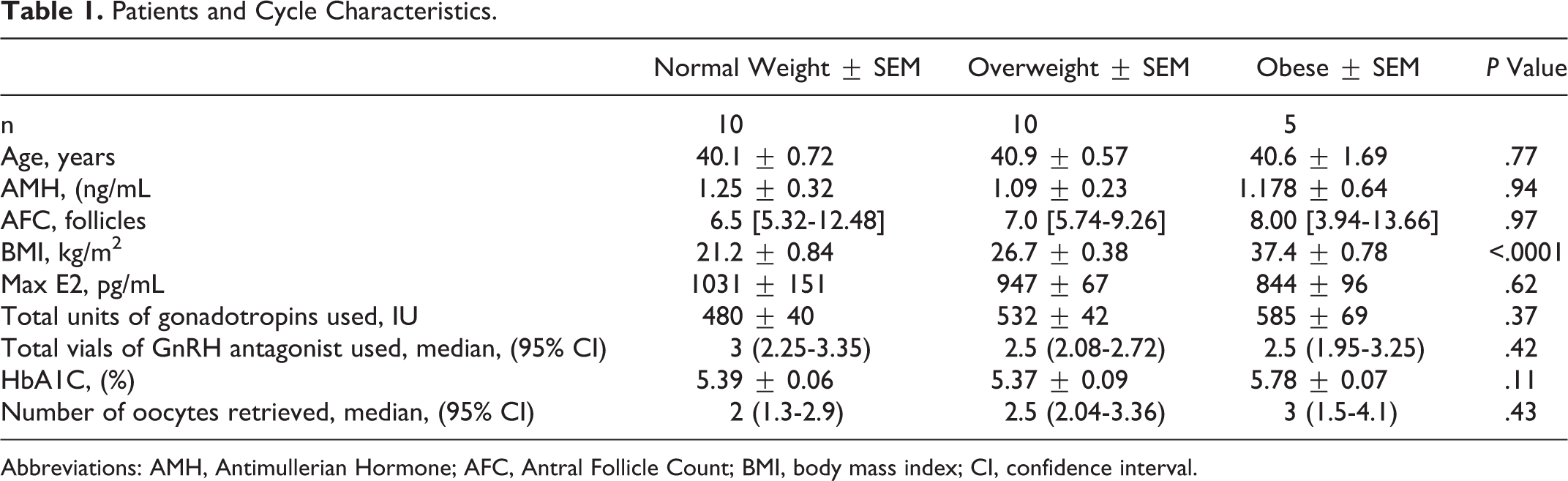
Abbreviations: AMH, Antimullerian Hormone; AFC, Antral Follicle Count; BMI, body mass index; CI, confidence interval.
FF Levels of Metabolic Hormones
An initial analysis was conducted to compare metabolic hormone levels between normal weight and overweight participants. Levels of all metabolic hormones were indistinguishable between these 2 groups. Hence, comparisons shown in Table 2 were conducted between obese and nonobese (normal weight combined with overweight) participants. Interestingly, levels of certain metabolic hormones were strikingly different in FF from obese women. Levels of hormones related to glucose and energy homeostasis and regulation of fat stores including insulin, glucagon, GLP-1, C-peptide, and leptin were increased significantly in FF from obese women compared to FF from nonobese women. In contrast, levels of GIP, ghrelin, PAI-1, resistin, and adiponectin were similar between the 2 groups. Levels of visfatin, an enzyme in the nicotinamide adenine dinucleotide (NAD+) salvage pathway formerly known as PBEF1 that promotes B cell maturation and inhibits neutrophil apoptosis, was only detectable in the FF of 7 of 25 patients, and the levels did not correlate with BMI.
Levels of Metabolic Hormones in Follicular Fluid of Women Undergoing IVF.

Abbreviations: GIP, gastric inhibitory polypeptide; GLP-1, glucagon-like peptide-1; PAI-1, plasminogen activator inhibitor-1; IVF, in vitro fertilization; BMI, body mass index.
Metabolite Variation
The ANOVA results indicated that few metabolites reached the levels of statistical significance (eg, isoleucine, leucine, valine, and uric acid). These metabolites were increased in FF of obese women.
While ANOVA is an excellent statistical tool for categorical data, a curve fit is better suited for numerical data, such as the BMI values. We fit a quadratic curve for BMI versus each metabolite. The quadratic regression revealed significant associations between certain metabolites and BMI. Specifically, the levels of isoleucine, leucine, valine, uric acid, isocaproic acid, butanoic acid, tyrosine, threonine, glycine, and methionine in FF correlated with BMI. Thus, results of ANOVA and quadratic regression analyses both emphasize a positive correlation between BMI and levels of the branched amino acids isoleucine, leucine and valine as well as uric acid (Figure 1).
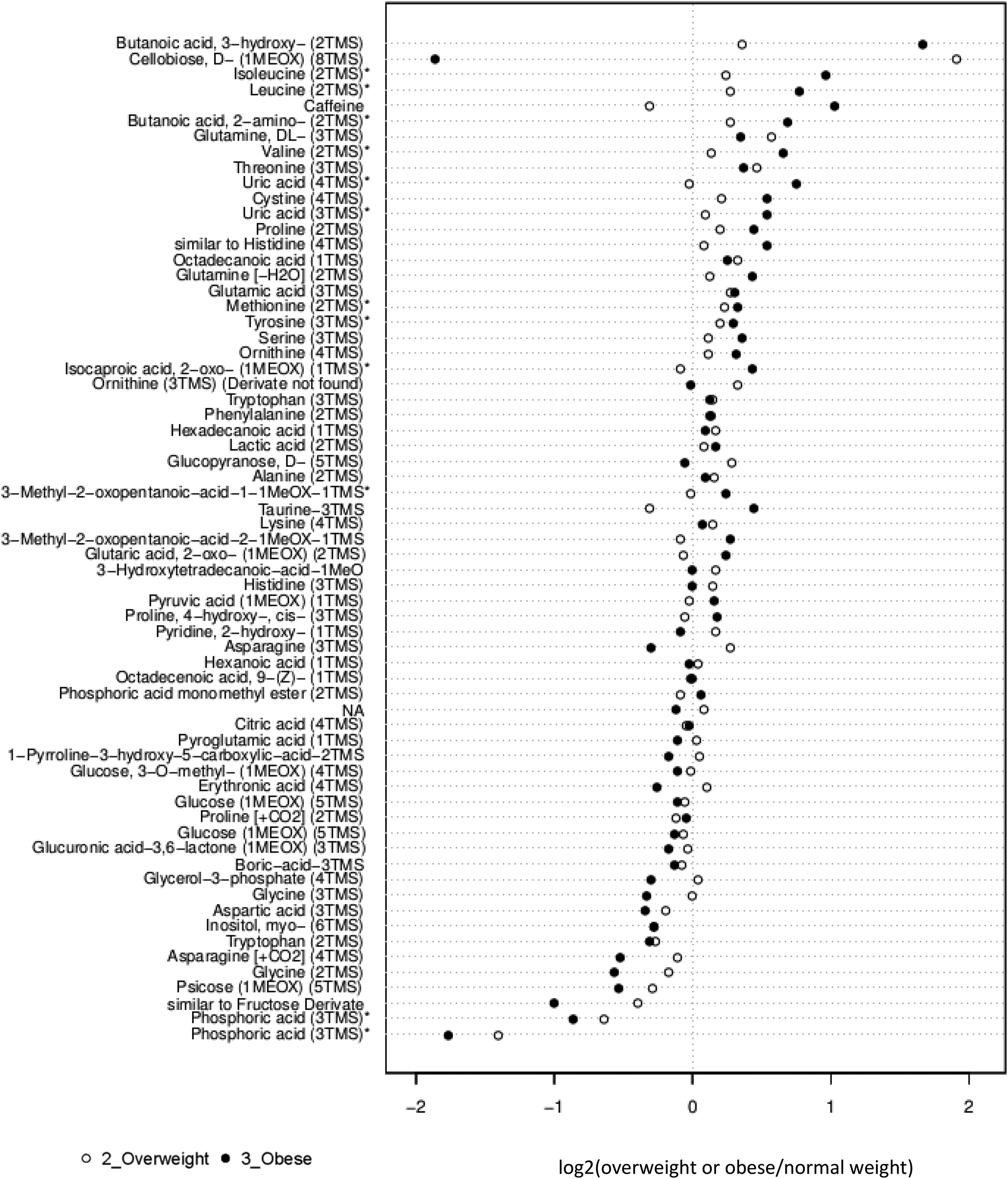
Fold change in follicular fluid fatty acid metabolites in obese or overweight women relative to normal weight women undergoing IVF. The center line (0) represents median levels in FF from normal weight women. Open symbols represent log2 (overweight or obese/normal weight) data from overweight participants (BMI 25-29.9) and solid symbols from obese (BMI ≥30) participants. *P < .05. IVF indicates in vitro fertilization; FF, follicular fluid; BMI, body mass index; TMS, trimethylsilyl; MEOX, methoxyamine hydrochloride.
Figure 1 represents the fold change between levels of each metabolite in FF of the overweight or obese groups compared to patients of normal weight. Briefly, isoleucine, leucine, valine, butanoic acid, uric acid, methionine, tyrosine, isocaproic acid, and oxopentanoic acid demonstrate a statistically significant increase with obesity, while phosphoric acid is decreased significantly with obesity.
We include the embryo outcomes of the patients in our study for the sake of completeness. Given our small sample size, no statistical analysis was performed between the groups. In the group of patients of normal weight, 5 patients obtained blastocysts, of which 4 underwent preimplantation genetic testing (PGT-A). Two of the biopsied embryos were euploid and transferred and resulted in live births in both patients. In the group of patients who are overweight, 7 patients obtained blastocysts all of which received PGT-A. Three of these patients had euploid embryos, but only 1patient underwent a frozen embryo transfer. This embryo transfer resulted in a live birth. Of the 5 obese patients, 1 obtained 3 day-3 embryos which were frozen at this stage. The remaining 4 patients obtained blastocysts that were cryopreserved. One of the patients underwent PGT-A which resulted in a euploid embryo but, however, did not result in a pregnancy upon frozen embryo transfer.
Discussion
Our results demonstrate alterations in the levels of metabolic hormones and fatty acid metabolites in the FF of obese women undergoing IVF. In particular, levels of insulin, glucagon, GLP-1, C-peptide, and leptin are elevated in the FF of obese women, and levels of branched chain amino acids (BCAA) leucine, valine, and isoleucine in FF are increased with obesity. Similar findings of elevated insulin and leptin levels have been previously reported in obese mares. 13 These significant alterations may play a role in oocyte quality and maturation, fertilization, and embryo quality and development, thereby affecting fertility in obese women. It is of particular interest to us that while these levels are similar in the normal weight and overweight women, the striking difference starts at a BMI of 30 and above.
Metabolic hormones have several described functions in follicular growth, oocyte development and maturity, and hormone production and secretion. Insulin for instance has a role in stimulating granulosa cell aromatase activity and steroid production and may have a part in regulating oocyte maturation. In animal studies, insulin exposure during bovine in vitro oocyte maturation demonstrated a decrease in blastocyst development rate and embryo quality, as well as a perturbance in gene expression. 14,15 Furthermore, in bovine granulosa cells in vitro, low doses of insulin stimulated aromatase activity and favored secretion of estradiol while high doses of insulin did not stimulate aromatase activity and instead favored the secretion of progesterone and androgens over the secretion of estradiol. 16 –19
Leptin, on the other hand, is produced in the adipose tissue and released into circulation according to energy stores. Levels of leptin in plasma have been demonstrated to be higher in participants with a higher BMI and a higher body fat percentage. 20 FF leptin is thought to play an important role in fertility. Levels of leptin in FF of women with unexplained infertility were found to be higher than in the FF of fertile women. 21 In addition, in patients undergoing ART, it was demonstrated that an FF leptin level correlated negatively with oocyte and embryo numbers, 22 embryo quality, and pregnancy success. 23 Several studies have also demonstrated a reduced ovarian response to gonadotropins with elevated FF leptin, suggesting an explanation for the higher amounts of gonadotropins generally needed in obese patients. 23,24 A systematic review of the literature 25 suggests its potential interest as a predictor of IVF outcome.
The GC- and HPLC-MS revealed that BCAA leucine, isoleucine, and valine are increased significantly with body weight. The BCAA have several metabolic functions including a role in fatty acid metabolism and regulation of serum glucose levels. 26 In mice, BCAA have a role in regulating blastocyst development and embryo implantation as well as enhancing fetal development through mTOR signaling pathway. The mTOR signaling is required for the implantation of murine embryos and induces trophectoderm motility and differentiation. 27 –29 With published data showing that leucine activates the mTOR signaling pathway, evidence is accumulating to indicate that leucine may play a role in blastocyst development. 27 –32 Our study did not investigate embryo implantation but rather focused on FF environment which may be associated with human oocyte competence in general. In a study performed in women with PCOS, the FF levels of BCAA, glutamic acid, phenylalanine, alanine, and arginine increased with BMI irrespective of PCOS status. 33 This same study found increased levels of leucine, valine, and glutamic acid in insulin-resistant PCOS compared to controls and non–insulin-resistant PCOS. Furthermore, increased FF BCAA levels correlated with decreased pregnancy rates and increased miscarriage rates. In our study, women with obesity showed both increased FF insulin and BCAA levels compared to overweight and normal weight women which may correlate with reported inferior IVF outcomes. 4
Our study has a number of strengths. All of our patients have received the same low amounts of gonadotropins per minimal stimulation IVF protocol ruling out potential effects of high-dose gonadotropins typically used in patients with advanced reproductive age and/or diminished ovarian reserve. In addition, most studies looking at the effects of obesity on the FF milieu are biased by the fact that obese women require higher doses of gonadotropins for stimulation. As a result, the reported differences between the obese and nonobese patients could be originating from the increased doses of medications administered rather than the sole presence of obesity. Our study eliminates this bias by limiting study patients to those undergoing a minimal stimulation protocol who have received similar doses of gonadotropins, thus decreasing the bias caused by medication effects on the FF milieu. Furthermore, in an effort to eliminate interpatient variability, all patients received the same stimulation, FF was collected by the same physician, in the same laboratory, and processed by the same individual. All of the patients recruited for this study have Diminished ovarian reserve (DOR), in order to account for the bias that DOR may have on FF metabolism. While this decreases interpatient variability, it is important to note that this may render our conclusions not generalizable to the whole pool of patients with infertility. By doing all of the above exclusions, we have limited our total number to 25 patients, keeping it a pilot study, which makes it challenging to generate strong clinical correlations which we see as a weakness of our study.
In conclusion, this study demonstrates significant alterations in the FF milieu of obese women undergoing IVF, which may contribute to the decreased fecundity of obese women. The extent to which these alterations affect the oocyte quality, maturity, and fertilization potential as well as the effect on embryo development has not been yet elucidated. Nonetheless, it is striking that the hormonal and metabolic milieu of FF from obese women undergoing IVF is abnormal with significant alterations in key hormones that regulate energy metabolism and branched chained amino acids that also regulate fatty acid metabolism and glucose. Although the impact of this environment on oocyte and embryo development is not fully understood, these changes may also lead to imprinting at the genomic level and long-term sequelae on offspring. The ability to conserve, acquire, and expend energy is believed to be an innate, ancient trait embedded in the genome. These changes are also believed to possibly be inherited across generations. 34 Recent studies on a more global scale reveal that the molecular mechanisms of generational problems with disease phenotypes are broadly caused by a suboptimal environment in the reproductive tract. 35 Here, we suggest that generational phenotypes may begin early in life through the development of oocytes in an adverse FF environment. With an increase in overweight and obese women around the world and in the United States, studies to clarify the impact of body weight on reproductive potential and offspring are of paramount importance.
Footnotes
Acknowledgments
The authors wish to thank the Proteomics and Metabolomics Facility at Colorado State University for their help with the GC/UPLC-MS analysis and interpretation. We also acknowledge the assistance of Dr Karla Saner-Amigh and Dr Jianming Li, embryologists at Fertility and Advanced Reproductive Medicine Clinic, in isolating the follicular fluid during oocyte retrievals.
Declaration of Conflicting Interests
The author(s) declared no potential conflict of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.