
Abstract
Progestins have been recommended for preterm birth prevention in high-risk women; however, their mechanism of action still remains an area of debate. Medroxyprogesterone acetate (MPA) has previously been shown to significantly inhibit tumor necrosis factor α (TNFα)-induced matrix metalloproteinase 9 (MMP9) messenger RNA (mRNA) expression and activity in primary amnion epithelial cells, a process that may lead to preterm premature rupture of membranes. A mechanism that explains MPA’s inhibition of TNFα-induced MMP9 mRNA expression and activity in primary amnion epithelial cells is unclear since these cells lack the classic nuclear progesterone receptor but express a membrane-associated progesterone receptor—progesterone receptor membrane component 1 (PGRMC1) along with the glucocorticoid receptor (GR). Primary amnion epithelial cells harvested from healthy term pregnant women at cesarean section were treated with PGRMC1 (to knockdown PGRMC1 expression), GR (to knockdown GR expression), or control small interfering RNA (siRNA; 10 nm) for 72 hours, pretreated with ethanol or MPA (10−6 M) for 6 hours, and then stimulated with or without TNFα 10 ng/mL for 24 hours. Real-time quantitative polymerase chain reaction and gelatin zymography were used to quantify MMP9 mRNA expression and activity, respectively. Experimental groups were compared using 1-way analysis of variance. Both TNFα-induced MMP9 mRNA expression and activity were significantly inhibited by pretreatment with MPA; however, only the inhibition of TNFα-induced MMP9 activity was partially reversed with PGRMC1 siRNA. However, GR siRNA reversed both the inhibition of TNFα-induced MMP9 mRNA expression and activity by MPA. This study demonstrates that MPA mediates its anti-inflammatory effects primarily through GR and partially through PGRMC1 in primary amnion epithelial cells.
Keywords
Introduction
Preterm premature rupture of membranes (PPROM) is responsible for up to a third of preterm births, and currently, there are no known therapies for its prevention due in part to a lack of understanding of the mechanisms that lead to PPROM. Progestins are clinically recommended for the prevention of preterm births in women with a prior history of spontaneous preterm birth. 1 However, the mechanism of action still remains an area of debate. Furthermore, the role of progestins in the prevention of PPROM still remains unclear. 1 –5 While clinical evidence for the use of progestins for the prevention of PPROM is lacking, in vitro studies have demonstrated that progestins may be able to inhibit inflammation-induced pathological changes in human fetal membranes that lead to fetal membrane weakening and PPROM. 6 –8 It has also been recently suggested that progestins other than 17α hydroxyprogesterone caproate (17OHP) may be more effective at attenuating inflammation-induced fetal membrane weakening leading to PPROM. 9
An increase in matrix metalloproteinase 9 (MMP9) activity and expression in human fetal membranes, and specifically in the amniochorion, reduces fetal membrane tensile strength and has been observed in both term labor and PPROM patients. 10 –13 Furthermore, a polymorphism in the MMP9 promoter region in primary amnion epithelial cells that increases promoter activity may be associated with an increased risk of PPROM. 14 An increase in MMP9 activity in the amnion results in degradation of extracellular matrix proteins such as type IV collagen, which is a component of the basement membrane for cell adhesion. Degradation of type IV collagen by MMP9 leads to amnion epithelial cell detachment from the basement membrane, apoptotic cell death, a reduction in fetal membrane tensile strength, and ultimately fetal membrane rupture. 15 In an in vitro fetal membrane model that mimics PPROM, tumor necrosis factor α (TNFα)-induced MMP9 expression is associated with a significant reduction in the tensile strength of human fetal membranes. 16 Interestingly, TNFα-induced fetal membrane weakening in vitro is partially inhibited by pretreatment with progestins including MPA. 8 We previously described that the progestin, MPA, inhibits TNFα-induced MMP9 activity and messenger RNA (mRNA) expression in the primary amnion epithelial cells but not in primary chorion cells. 6 It is unclear how MPA inhibits TNFα-induced MMP9 mRNA expression and activity in primary amnion epithelial cells. Primary amnion epithelial cells do not express the classic nuclear progesterone receptor (PGR) but do express other membrane-associated progesterone receptors. 17,18 We have demonstrated that the primary amnion epithelial cells express progesterone receptor membrane component 1 (PGRMC1). 17 We have also previously demonstrated that PGRMC1 may partially mediate the effect of MPA on TNFα-induced MMP9 activity in cytotrophoblast cells in vitro. 17 Furthermore, it has also been implicated in regulating the inflammatory response to Ureaplasma parvum exposure in primary amnion and chorion cells harvested from term fetal membranes. 19 The functional role of PGRMC1 is supported by the fact that PGRMC1 protein expression is diminished in the fetal membranes of women with PPROM, suggesting that it plays a role in molecular pathways that lead to PPROM. 20
Medroxyprogesterone acetate is a mixed effect progestin that also mediates some of its effects via the glucocorticoid receptor (GR). 21 In fact GR is ubiquitously expressed in both the decidua and fetal membranes at term. 22,23 Even though GR signaling has been implicated in pathways that lead to spontaneous preterm births, its effects also modulate downstream pathways involved in matrix degradation, inflammation, and collagen metabolism that may also be involved in PPROM. 24 Furthermore, multiple mechanisms which may lead to PPROM such as prostaglandin synthesis, the expression of lysyl oxidase by amnion mesenchyme cells, and apoptosis in the amnion epithelial cells by glucocorticoids are regulated through GR. 25,26 Additionally, GR is partially responsible for progestin-mediated effects in vitro. For example, in term primary myometrium cells, the suppression of interleukin 1β (IL1β)-induced COX2 expression both by progesterone and MPA is partly mediated through GR. 27
In this study, our objective was to determine whether the inhibition of TNFα-induced MMP9 activity and mRNA expression by MPA was mediated through PGRMC1. Alternately we sought to determine whether this progestin-mediated effect was also mediated through GR. Our working hypothesis was that MPA would inhibit TNFα-induced MMP9 activity and gene expression primarily through a PGRMC1-dependent mechanism.
Methods
Patients and Samples
Fetal membranes were collected from healthy term patients having elective cesarean section without labor (n = 11). These samples were deidentified preventing any link to clinical information, and as a result, this study was covered by a waiver of consent from the institutional review board of Duke University.
Primary Amnion Epithelial Cell Isolation
Primary amnion epithelial cells were harvested from fetal membrane samples using a modification of the previously described protocol. 28 Briefly the amnion was peeled off from the underlying choriodecidua using a pair of forceps and washed in Dulbecco's Modified Eagle Medium: Nutrient Mixture F12 (DIMEM/F12) media containing antibiotics–antimycotics (ThermoFisher, Waltham, Massachusetts) to remove blood and debris. To remove amnion epithelial cells, the tissue was minced using a pair of scalpel blades and then digested in DMEM/F12 media with antibiotics–antimycotics and 1 g of 1:250 trypsin (MilliporeSigma, St Louis, Missouri) for 30 minutes at 37°C with intermittent shaking. The digest was filtered using a metal strainer and the filtrate was centrifuged at 2000g for 5 minutes and the cells resuspended in DIMEM/F12 with antibiotics-antimycotics and 10% fetal bovine serum (FBS; ThermoFisher, Waltham, Massachusetts). This process was repeated an additional 2 times on the resulting undigested tissue fragments. The cells collected from the first and second digestions were pooled and centrifuged at 2000g and resuspended in DIMEM/F12 with antibiotics–antimycotics and 10% FBS. The primary amnion epithelial cells were then plated in 10-cm culture dishes and incubated in humidified air and 5% CO2 for 7 days until they grew to confluence with cell culture media change every 48 hours. Primary amnion epithelial cells were passaged using 0.25% trypsin with EDTA (ThermoFisher, Waltham, Massachusetts) and plated at approximately 0.5 to 1.0 × 106 cells/mL for subsequent experiments in 6-well culture plates. Cells were passaged only once for subsequent experiments.
Immunofluorescence
Primary amnion epithelial cells were plated on 3-well chamber slides at 1 × 105 cell/mL in DIMEM/F12 with antibiotics–antimycotics and 10% FBS for 24 to 48 hours. The cells were fixed in ice-cold 100% methanol at −20°C for 10 minutes. The cells were first incubated with Image -IT Signal Enhancer (ThermoFisher, Waltham, Massachusetts) for 30 minutes and then permeabilized and blocked with 5% goat serum and 0.3% Triton X for 1 hour and then incubated with antirabbit polyclonal PGRMC1 antibody 1:100 (catalog No. HPA08277; MilliporeSigma, St Louis, Missouri) and antimouse monoclonal GR antibody 1:250 (catalog no. SAB4800041; MilliporeSigma, St Louis, Missouri) overnight at 4°C in a humidified slide chamber. An isotype monoclonal antimouse (catalog no. MA5-14453; ThermoFisher, Waltham, Massachusetts) and polyclonal antirabbit antibody (catalog no. ab27472; Abcam, Cambridge, Massachusetts) were used as negative controls. The cells were then incubated with the Alexa Fluor 488 goat antimouse ReadyProbes (ThermoFisher, Waltham, Massachusetts), and Alexa Fluor 594 goat antirabbit ReadyProbes (ThermoFisher, Waltham, Massachusetts) diluted according to the manufacturer’s specifications for 1 hour. The cells were then incubated with 4', 6-Diamidino-2-Phenylindole, Dihydrochloride (DAPI) 1:1000 for 5 minutes. The slides were mounted with ProLong Diamond Antifade mounting media (ThermoFisher, Waltham, Massachusetts) and imaged with the Zeiss Axio Imager fluorescence microscope.
Progesterone Receptor Membrane Component 1 and GR Small Interfering RNA Knockdown
Primary amnion epithelial cells were plated at a density of 0.5 to 1.0 × 106 cells for 24 to 48 hours. The cells were then transfected in serum and antibiotic-free DMEM/F12 with control small interfering RNA (siRNA, ID: AM4611; ThermoFisher, Waltham, Massachusetts), PGRMC1 siRNA (ID: S21310; ThermoFisher, Waltham, Massachusetts), or GR siRNA (ID: AM51331; ThermoFisher, Waltham, Massachusetts) using Lipofectamine RNAimax (ThermoFisher, Waltham, Massachusetts). The final concentration of the siRNA used was 10 nM. The cells were incubated for 72 hours but were supplemented with serum containing media after 24 hours (1 mL of DMEM/F12 with 20% FBS). At the end of the 72-hour incubation, the cells were pretreated with ethanol (unstimulated control) or MPA (10−6 M) (MilliporeSigma, St Louis, Missouri) for 6 hours followed by stimulation with or without TNFα 10 ng/mL (RnD Systems, Minneapolis, Minnesota) for an additional 24 hours in DMEM/F12 with 1% FBS. At the end of the experiments, cell culture media were harvested for MMP9 quantification using gelatin zymography, and TRIzol (Qiagen, Germantown, Maryland) lysates were harvested for MMP9 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) mRNA quantification using realt-time quantitative polymerase chain reaction (RT-qPCR). The efficiency of siRNA transfection and inhibition of protein expression were quantified by Western blot for PGRMC1, GR, and GAPDH on protein lysates from parallel experiments harvested at the end of the experimental treatment conditions.
Polymerase Chain Reaction
Total RNA was isolated from TRIzol lysates according to the manufacturer’s protocol using the RNeasy Mini Kit (Qiagen, Germantown, Maryland). Total RNA concentration was quantified with the Nanodrop spectrophotometer, and 1000 ng of RNA was reverse transcribed into complementary DNA (cDNA) using the Superscript III first-strand RNA synthesis system (ThermoFisher, Waltham, Massachusetts). Fifty nanograms of cDNA was used as the template for real-time polymerase chain reaction (PCR) using prevalidated Taqman gene expression probes targeting MMP9 (Assay ID: Hs00234579_m1) and GAPDH (assay ID: HS03929097_g1) with the IQ Supermix Master Mix (Bio-Rad, Hercules, California). Real-time quantitative PCR was performed using the iCycler IQ real-time PCR detection system (Bio-Rad, Hercules, Californial), using the following protocol: initial denaturation at 95°C for 3 minutes, followed by a 2-step amplification process of 95°C for 30 seconds and 60°C for 40 seconds for a total of 40 cycles. All samples were run in duplicate and the gene expression for the genes of interest was evaluated using the 2− ▵▵CT method normalizing MMP9 mRNA expression to the housekeeping gene GAPDH.
Gelatin Zymography
Matrix metalloproteinase 9 activity was quantified using gelatin zymography as previously described. 6,17 Harvested cell culture media from each experimental group were mixed in a 1:1 ratio with Novex Tris–glycine sodium dodecyl sulfate (SDS) sample buffer (ThermoFisher, Waltham, Massachusetts) and 10 to 20 μL was loaded on 10% Novex zymogram gels (ThermoFisher, Waltham, Massachusetts) and electrophoresed at 125 V for 90 minutes on ice. The gels were then incubated in Novex renaturing buffer (ThermoFisher, Waltham, Massachusetts) for 30 minutes followed by 30 minutes of incubation in Novex developing buffer (ThermoFisher, Waltham, Massachusetts). The gels were incubated in fresh developing buffer for an additional 18 hours at 37°C. They were then stained with a SimplyBlue Safestain (ThermoFisher, Waltham, Massachusetts) for 1 hour and then destained by washing the gels in deionized water for 1 hour twice at room temperature. The total MMP9 activity was quantified by analyzing band densities at 88 and/or 92 kDa using Image J (NIH, Bethesda, Maryland). For each gel, the bands were normalized to the TNFα stimulated control in the control siRNA group.
Western Blot
At the termination of the experiments, cell culture media were removed and cells were washed with ice-cold PBS and then lysed in radioimmunoprecipitation assay buffer (MilliporeSigma, St Louis, Missouri) with cOmplete mini protease cocktail inhibitor (MilliporeSigma, St Louis, Missouri) for 5 minutes on ice. The cells were removed from the plate using a cell scraper and placed in a fresh 1.5-mL microcentrifuge tube for an additional 30 minutes on ice. The tubes were centrifuged at 13 000g at 4°C for 20 minutes and the supernatant was harvested. The harvested supernatant was frozen at −80°C until analyzed. Protein concentrations were quantified using the Bradford assay and 10 to 15 μg of total protein was loaded onto 4% to 12% Bis–Tris gels and electrophoresed at 125 V for 60 minutes and then transferred onto a polyvinylidene difluoride membrane. The membranes were incubated with rabbit antihuman GR (1:1000 dilution, catalog No. 3660 S; Cell Signaling, Danvers, Massachusetts), PGRMC1 (1:2000 dilution, catalog No. HPA08277; MilliporeSigma, St Louis, Missouri), and GAPDH (1:10,000 dilution, catalog No.2118 S; Cell Signaling, Danvers, Massachusetts) in blocking buffer (5% nonfat milk in Tris-buffered saline and Tween 20 buffer) overnight at 4°C followed by incubation with the secondary antibody (1:1000, horseradish peroxidase-linked antirabbit; Cell Signaling, Danvers, Massachusetts) for 1 hour at room temperature. The membranes were then incubated with SuperSignal West Pico Chemiluminescent Substrate (ThermoFisher, Waltham, Massachusetts) and exposed on film. The density of the bands was analyzed using ImageJ and both GR and PGRMC1 were normalized to GAPDH.
Statistical Analysis
Experimental groups were compared using 1-way analysis of variance with post hoc pairwise comparisons using the Šidák test. Each P value was adjusted for multiple comparisons. A P < .05 was considered significant. Data were analyzed using GraphPad Prism. Data are presented as mean ± standard error of the mean.
Results
Progesterone Receptor Membrane Component 1 and GR Localization in Primary Amnion Epithelial Cells
In primary amnion epithelial cells, PGRMC1 localized primarily to the cytoplasm, particularly in the perinuclear region (Figure 1A and D). The GR localized to the cytoplasm and nucleus (Figure 1B and E). Both receptors localized to the perinuclear region (Figure 1C and F).
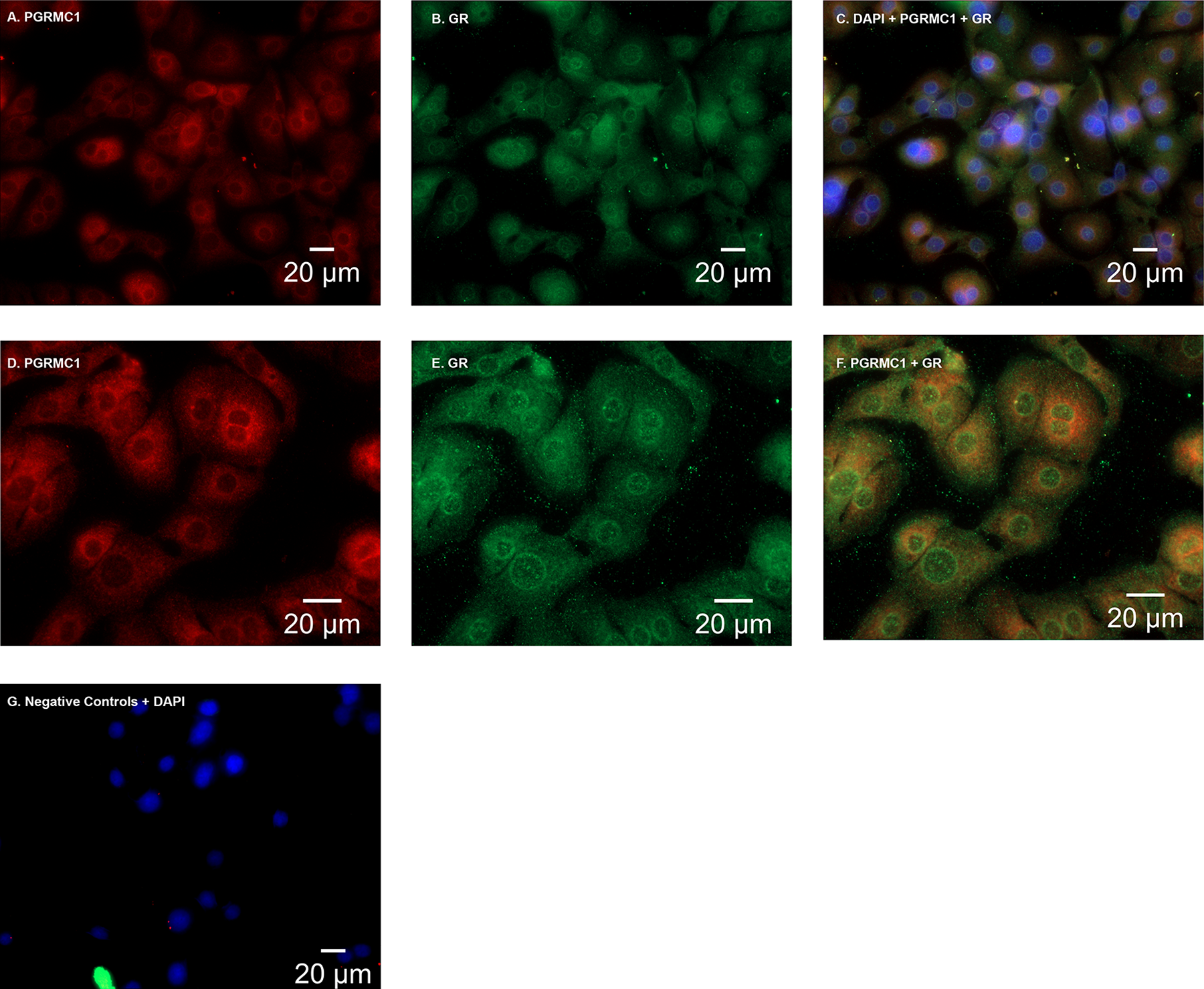
A and D, In primary amnion epithelial cells, progesterone receptor membrane component 1 (PGRMC1) was localized (red) primarily to the perinuclear region and to the cytoplasm. B and E, Glucocorticoid receptor (GR; green) localized to the cytoplasm and the nucleus. C and F, Both PGRMC1 and GR localized in primary amnion epithelial cells with 4', 6-Diamidino-2-Phenyindole Dihydrochloride (DAPI) in the perinuclear region. G, Primary amnion epithelial cells stained with isotype controls (a monoclonal mouse and polyclonal rabbit antibody) plus DAPI. Images (A-C and G) were captured at a magnification of 40×. Images D to F were captured at a magnification of 63×.
Progesterone Receptor Membrane Component 1 siRNA Inhibited PGRMC1 Protein Expression and GR siRNA Inhibited GR Protein Expression
Treatment of the primary amnion epithelial cells with PGRMC1 siRNA significantly inhibited PGRMC1 protein expression in primary amnion epithelial cell cultures by approximately 59 ± 0.06% (P < .001) when compared to the control (Figure 2A). Interestingly, we also observed a 37 ± 0.05% reduction in GR protein expression when PGRMC1 protein expression was inhibited with PGRMC1 siRNA (P < .001). Treatment of primary amnion epithelial cells with GR siRNA significantly inhibited GR protein expression by 96 ± 0.05% (P < .001) when compared to the control (Figure 2B).
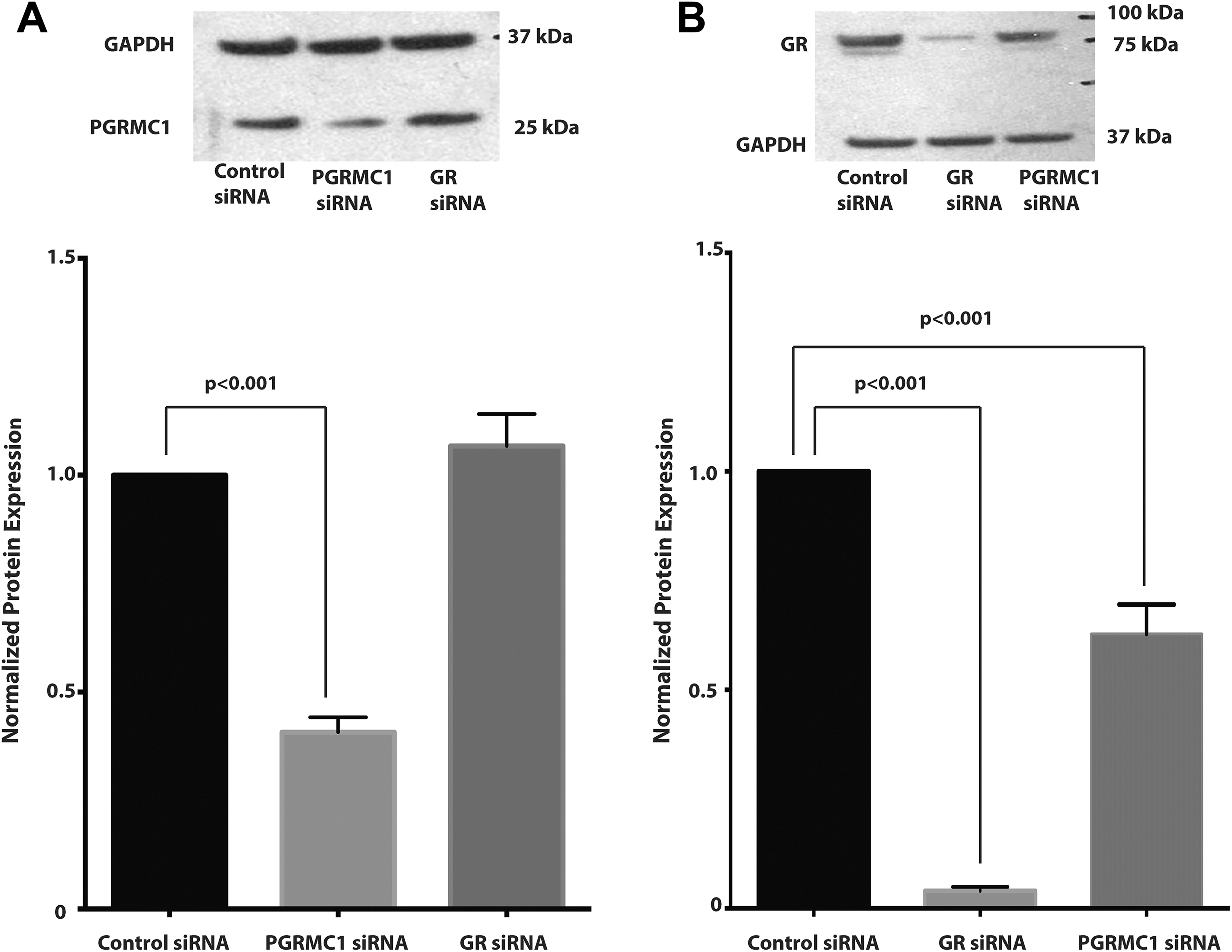
A, Progesterone receptor membrane component 1 (PGRMC1) small interfering RNA (siRNA) significantly inhibits PGRMC1 protein expression. Protein expression was normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and then normalized to the control siRNA group. A representative Western blot is shown above. B, Glucocorticoid receptor (GR) siRNA significantly inhibits GR protein expression and PGRMC1 siRNA also partially inhibits GR protein expression (n = 6 patient samples). Data represent mean ± standard error of the mean (SEM).
The Effect of PGRMC1 and GR siRNA on Basal MMP 9 mRNA Expression and Activity
When compared to the unstimulated control, MPA did not inhibit basal MMP9 mRNA expression (Figure 3) or activity within the control, PGRMC1, or GR siRNA-treated groups (Figure 4). There were no significant differences in basal MMP9 mRNA expression when comparing the unstimulated control in the control siRNA with those in the PGRMC1 siRNA group or GR siRNA group. Similarly, there were no significant differences in basal MMP9 mRNA expression when comparing cells treated with MPA only in the control siRNA group with those in the PGRMC1 siRNA or GR siRNA group. Glucocorticoid receptor protein expression inhibition was associated with a significant increase in basal MMP9 activity when compared with the control group (Figure 4A and B). Basal MMP9 activity was significantly increased in the unstimulated control in the GR siRNA group when compared to the unstimulated control in the control siRNA group (unstimulated control in control siRNA vs unstimulated control in GR siRNA, P = .044; Figure 4A and B). Similarly, basal MMP9 activity was significantly increased in the cells pretreated with MPA only in the GR siRNA when compared with the similar group in the control siRNA group (MPA only in control siRNA vs MPA only in GR siRNA, P = .007; Figure 4A and B).
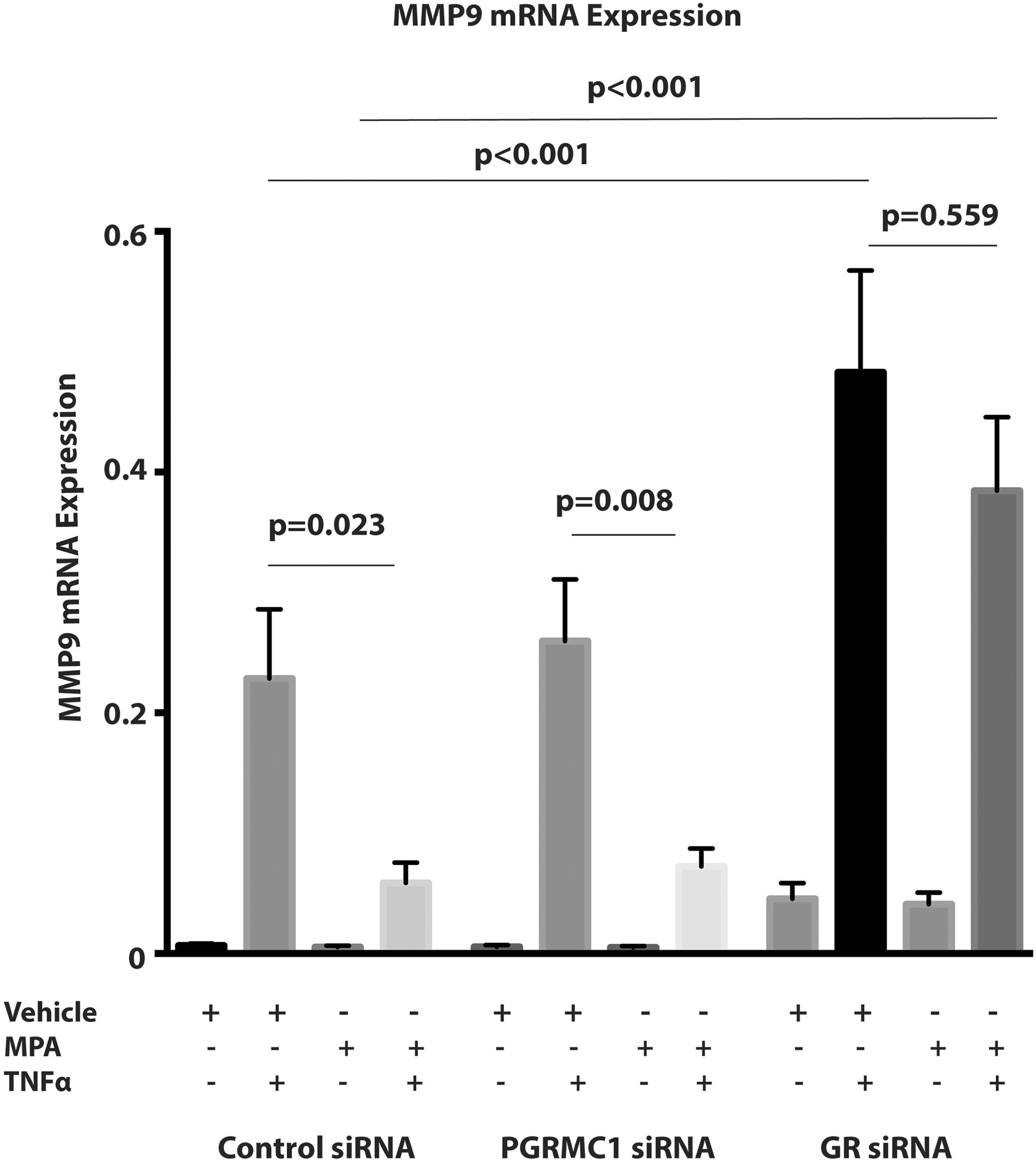
The effect of progesterone receptor membrane component 1 (PGRMC1) small interfering RNA (siRNA) and glucocorticoid receptor (GR) siRNA on both basal matrix metalloproteinase 9 (MMP9) and tumor necrosis factor α (TNFα)-induced MMP9 mRNA expression. The MMP9 mRNA expression was normalized to Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and the y-axis represents the 2−▵▵CT values (n = 11 patient samples). Data represent mean ± standard error of the mean (SEM).
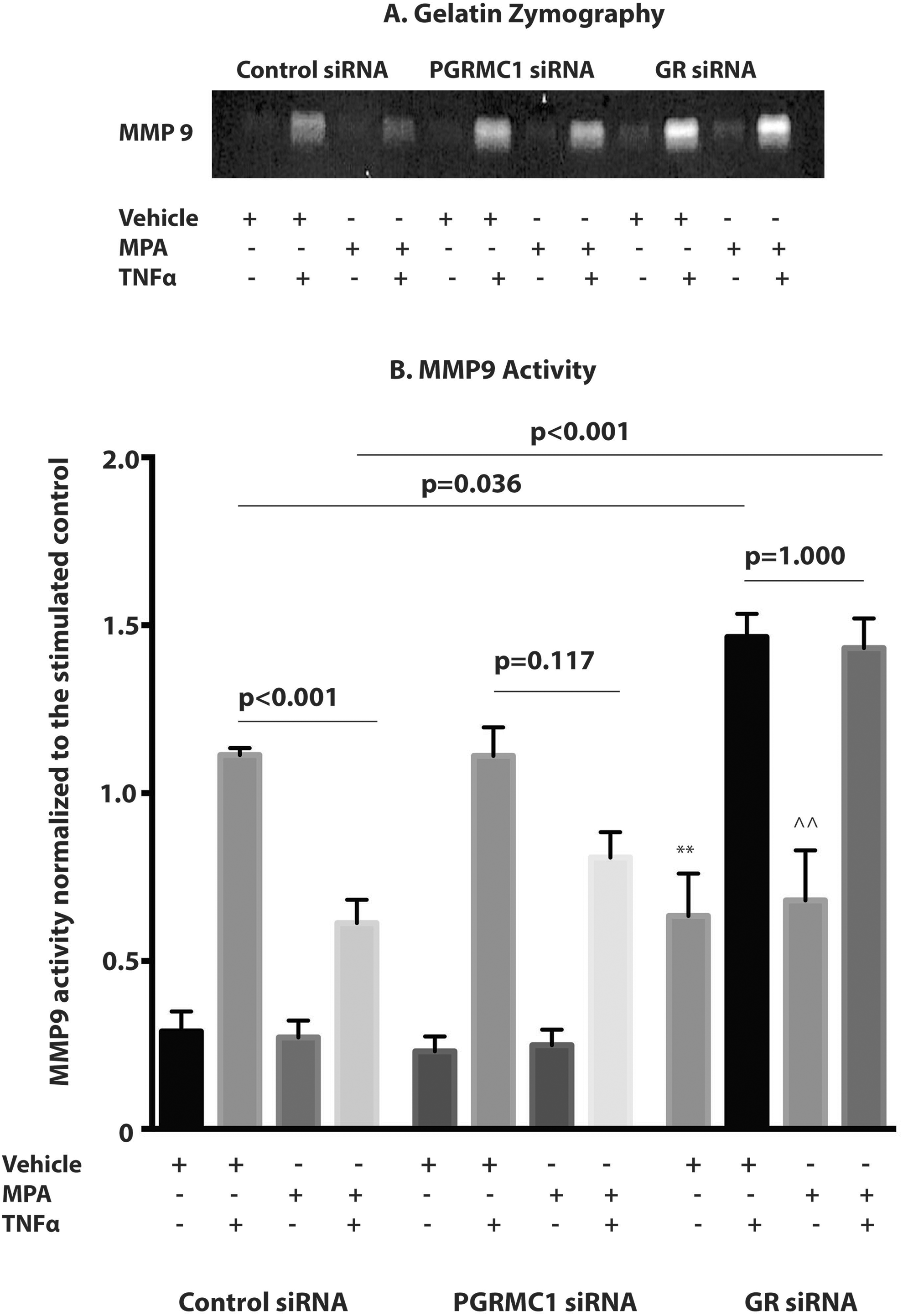
The effect of progesterone receptor membrane component 1 (PGRMC1) small interfering RNA (siRNA) and glucocorticoid receptor (GR) siRNA on both basal matrix metalloproteinase 9 (MMP9) and tumor necrosis factor α (TNFα)-induced MMP9 activity. A, A representative zymogram gel is shown with the white areas proportional to the MMP9 activity. B, The MMP9 activity was normalized to vehicle + TNFα sample (stimulated control) in the control siRNA group. **P = .044 vehicle (unstimulated control) in control siRNA group versus vehicle (unstimulated control) in GR siRNA group. ^^P = .007 MPA in the control siRNA group versus MPA in the GR siRNA group (n = 9 patient samples). Data represent mean ± standard error of the mean (SEM).
The Effect of PGRMC1 and GR siRNA on TNFα-Induced MMP9 mRNA Expression and Activity
Tumor necrosis factor α significantly induced MMP9 mRNA expression (Figure 3) and activity (Figure 4) in the control, PGRMC1, and GR siRNA groups (P < .001 for unstimulated control vs TNFα in all groups). Pretreatment with MPA significantly inhibited TNFα-induced MMP9 mRNA expression (P = .023; Figure 3) and activity (P < .001; Figure 4) when compared with the stimulated control in the control siRNA group. However, when GR protein expression was inhibited, the inhibition of TNFα-induced MMP9 mRNA expression (P = .559; Figure 3) and MMP9 activity (P = 1.000; Figure 4) by pretreatment with MPA when compared to the stimulated control, observed in the control siRNA group, were both attenuated. Additionally, following GR protein expression inhibition, TNFα induced a significantly greater increase in both MMP9 mRNA expression and activity in the stimulated control (stimulated control in control siRNA vs stimulated control in GRsiRNA, P < .001 for mRNA, P = .036 for MMP9 activity) when compared to the control siRNA group (Figures 3 and 4). Inhibition of GR protein expression was also associated with a significantly greater increase in both TNFα-induced MMP9 mRNA expression (Figure 3) and MMP9 activity (Figure 4) in cells pretreated with MPA and then stimulated with TNFα (MPA +TNFα in control siRNA vs MPA +TNFα in GRsiRNA, P < .001 for mRNA, P < .001 for MMP9 activity) when compared to the control siRNA groups. Pretreatment with MPA significantly inhibited TNFα-induced MMP9 mRNA expression (P = .008) when compared with the stimulated control in the PGRMC1 siRNA group (Figure 3). However, the inhibition of TNFα-induced MMP9 activity by pretreatment with MPA observed in the control siRNA group was partially attenuated by PGRMC1 inhibition (P = .110; Figure 4).
Discussion
The main finding from our study was that the inhibition of TNFα-induced MMP9 activity and mRNA expression by MPA is primarily mediated by GR. This finding represents an extension of our previous work which demonstrated that MPA significantly inhibited TNFα-induced MMP9 activity and mRNA expression in primary amnion epithelial cells. 6 Inhibition of PGRMC1 protein expression also resulted in partial attenuation of TNFα-induced MMP9 activity by MPA. We have previously demonstrated that the partial inhibition of TNFα-induced MMP9 activity in the HTR8/SVneo cytotrophoblast cell line by MPA was also PGRMC1 mediated. 17 Another interesting finding was that though PGRMC1 and GR both have different expression patterns they do both appear to localize to the cytoplasm, particularly around the nucleus in primary amnion epithelial cells. The localization of GR to both the cytoplasm and nucleus has previously been described. 29,30 Progesterone receptor membrane component 1 has a transmembrane domain and has been shown to localize to multiple subcellular locations including the cytoplasmic and nuclear membranes, the nucleolus, the golgi apparatus, endosomes, and secretory vesicles. 31 –34 These multiple locations suggest that PGRMC1 may play a role in regulating multiple cellular processes.
One of the proposed mechanisms by which progestins may prevent fetal membrane weakening is through inhibition of molecular mechanisms by interaction with membrane-associated progesterone receptors such as PGRMC1. 8 Progesterone receptor membrane component 1 is known to bind progesterone; however, only functional studies have demonstrated that MPA may mediate some of its effects through PGRMC1, which would be in line with its ability to bind to progesterone receptor. 35 Of the progestins, MPA is known to also have significant affinity for GR. 21 Meng et al demonstrated that MPA and dexamethasone at a dose 10−7 M stimulate GR in primary amnion epithelial cells, chorion cells, and decidual stromal cells. 36,37 Medroxyprogesterone acetate is not used in clinical practice for preterm birth prevention; however, recent work by Kumar et al demonstrated that progestogens including MPA significantly inhibit both TNFα and thrombin-induced fetal membrane weakening in vitro. 8 Furthermore, they demonstrated that this weakening effect mediated by the inflammatory mediator GM-CSF was attenuated by RU-486. 8 RU-486 is known to be a GR antagonist, substantiating the possibility that some of the inhibition of inflammation-induced fetal membrane weakening inhibited by progestins in fetal membranes may also be mediated through GR. 38 In fact, the induction of IL1β-induced COX2 gene expression was significantly inhibited by both progesterone and MPA primarily through GR, further highlighting the role of GR in mediating the anti-inflammatory effects of progestins. 27
Both the role of GR in fetal membranes and its expression in placental and fetal membranes are still areas of debate. In placental decidual cells and interstitial trophoblasts, GR is constitutively expressed but its expression is unaffected by labor status. 22 Interestingly, GR is highly expressed in amnion epithelial cells, amnion mesenchyme cells, and the chorion laeve trophoblasts. 23 Glucocorticoid receptor nuclear staining in fetal membranes was increased in preterm labor patients when compared to term labor and term cesarean section patients in all the above cell types. 23 However, the authors did not investigate GR expression in fetal membranes harvested from PPROM patients. Amnion epithelial cells when compared to the choriodecidua also express higher levels of steroid receptor coactivators (steroid receptor coactivator 1, p300, and p300/CBP-associated factor), which may also regulate GR activity and are known to play a role in labor initiation and regulation. 27
Glucocorticoid receptor signaling may play an important role in labor initiation as evidenced by the surge in glucocorticoid levels in maternal, fetal, and amniotic fluid levels as gestation ends. 39 –41 A local increase in cortisol may also initiate fetal membrane rupture by stimulating amnion epithelial cell apoptosis via the tissue-type plasminogen (tPA)/plasmin system activation of cleaved caspase-3. 26 This regulation of tPA/plasmin system is now thought to be mediated through GR and demonstrates that GR may also regulate pathways leading to PPROM. 26 One interesting finding from our study was that inhibition of GR protein expression was associated with significant increase in both basal MMP9 activity and TNFα-induced MMP9 activity and mRNA expression in amnion epithelial cells when compared with the control group. This exaggerated inflammatory response with and without TNFα stimulation suggests that GR may play a role in regulating MMP9 expression and activity and inflammatory pathways that lead to PPROM. This response may also partially explain how chronic psychosocial stress may lead to spontaneous preterm births and highlights another physiological role of GR. Chronic psychosocial stress is associated with an increased risk of preterm births and transgenerational stress in animal models is also associated with preterm births. 42,43 Pathway analysis studies have demonstrated that GR may be implicated in molecular pathways that lead to spontaneous preterm births. 24 However, candidate gene studies and gene linkage studies investigating associations of single nucleotide polymorphisms in GR with spontaneous preterm births have to date demonstrated no significant associations. 44,45 However, none of these studies specifically focused on PPROM patients or their offspring.
Progesterone receptor membrane component 1 is a membrane associated with progesterone receptor which regulates genes associated with apoptosis and may play a role in cell cycle regulation and cholesterol biosynthesis, and through its interaction with the epidermal growth factor receptor and cytochromes P450, it may regulate cancer cell proliferation and chemotherapy drug resistance. 46 –48 Recently, it has been demonstrated that PGRMC1 forms a complex with 2 other membrane-associated progesterone recetors —PGRMC2 and the progestin and adipoQ receptor family member-7. The resulting cytoplasmic complex was found necessary for some of progesterone’s function, such as its ability to suppress the ability of ovarian granulosa cells to enter the cell cycle. 49 Although our findings demonstrate that the inhibition of PGRMC1 does not attenuate the inhibition TNFα-induced MMP9 mRNA expression by MPA, it does not rule out the possibility that MPA’s effects could be mediated through PGRMC2 and its interaction with other membrane-associated progesterone receptors. Our group has previously demonstrated that PGRMC1 protein expression is significantly diminished in fetal membranes of PPROM patients, particularly at rupture sites, when compared with term and preterm no-labor patients. 20 However, determining expression patterns of the other membrane-associated PGRs in fetal membranes has not been comprehensively investigated in the different preterm birth phenotypes, which could shed further light on the role of these receptors in preterm births.
Our data demonstrated that PGRMC1 inhibition partially attenuates MPA’s inhibition of TNFα-induced MMP9 activity, but not MMP9 expression. This concurs with the findings of Feng et al who also demonstrated that PGRMC1 inhibition partially attenuated the Ureaplasma parvum’s induction of MMP9 activity but not mRNA expression in amnion epithelial cells. 19 One possible explanation for this effect is that PGRMC1 may exert its effect posttranslationally or on modulators of MMP9 activity. For example, PGRMC1 is now known to regulate MMP9 activity through its effect on the expression and production of neutrophil gelatinase-associated lipocalin—a glycoprotein which modulates MMP9 activity. 50 Another possible explanation is that PGRMC1 may exert its effect indirectly on modulators of MMP9 activity through functional interaction with GR as evidenced by the partial downregulation in GR protein expression when PGRMC1 protein expression was inhibited despite sharing no sequence homology. Recently, it was also demonstrated that MPA may upregulate PGRMC1 protein expression in fetal membranes possibly through a GR-dependent mechanism. 36 While these effects may reflect off target effects of siRNA treatment, the fact that both PGRMC1 and GR are expressed in similar subcellular locations in amnion epithelial cells suggests that functional interactions between these receptors may play a role in regulating cellular processes, particularly in cells that lack the classic nuclear PGR.
One limitation of this study is that we did not investigate the effects of progestins on other clinically relevant mechanisms that may lead to PPROM. For example, placental abruption and the subsequent production of thrombin have been implicated in the pathogenesis of PPROM. 51 –54 Thrombin has also been shown to directly weaken the isolated amnion layer in vitro both through the induction of MMP9, cleaved poly (ADP-ribose) polymerase and a reduction in tissue inhibitor of matrix metalloproteinase 3 protein expression mediated through the thrombin receptor—protease activated receptor-1 (PAR-1). 53 However, primary amnion epithelial cells do not express PAR-1 and thrombin may not induce MMP9 mRNA expression or activity in this cell type. 55 Instead recent evidence demonstrates that in the amnion, thrombin may exert its effects primarily on amnion mesenchymal cells which express PAR-1. 55 Investigating the effect of progestins on thrombin-mediated effects in amnion mesenchymal cells may further determine the role of progestins in preventing PPROM in the setting of placental abruption. Another limitation of this study is that we utilized MPA as the progestin of interest, but this was based on our previous findings that demonstrated that MPA but neither progesterone nor 17OHP suppressed TNFα-induced MMP9 activity or mRNA expression of primary amnion epithelial cells in vitro. 6 However, MPA is not used clinically for preterm birth prevention. Clinical trials investigating its use for preterm birth prevention did not demonstrate a significant clinical benefit, but these trials are outdated, have small sample sizes, and are methodologically flawed. 56 –58 A recent in vitro study has also questioned the role of 17OHP for the prevention of PPROM. 9 In this study, while 17OHP inhibited the generation of GM-CSF from the choriodecidua, it did not prevent the subsequent weakening of the amnion layer by GM-CSF. 9 Our findings in the current study suggest that GR may be the primary mediator of MPA’s effect in vitro. While systemic administration of GR agonists in vivo could have unintended consequences given GRs ubiquitous expression, 59 further studies are warranted to investigate fetal membrane–specific GR agonism either through modulation of GR function or by targeting GR signaling pathways to reduce any adverse effects.
Conclusion
In summary, our findings demonstrate that the progestin MPA inhibits TNFα-induced MMP9 activity and mRNA expression primarily through GR and partially through PGRMC1. Furthermore, GR regulates the amnion’s inflammatory response to cytokines and may play a central role in modulating pathways in fetal membranes that lead to fetal membrane weakening and PPROM. Modulating GR function or targeting GR regulated pathways for the prevention of PPROM may be a useful therapeutic strategy which warrants reevaluation.
Footnotes
Authors’ Note
Terrence K. Allen studied the concept and design. He performed experiments, data analysis, and interpretation. He drafted the manuscript and contributed to the approval of the final version of the manuscript. Matthew N. Nazzal performed experiments, data analysis, and interpretation. He drafted the manuscript and contributed to the approval of the final version of the manuscript. Liping Feng studied the concept and design. She drafted the manuscript and contributed to the approval of the final version of the manuscript. Irina A. Buhimschi studied the concept and design. She drafted the manuscript and contributed to the approval of the final version of the manuscript. Amy P. Murtha studied the concept and design. She drafted the manuscript and contributed to the approval of the final version of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the