
Abstract
Introduction
Breast cancer remains a leading cause of cancer-related morbidity worldwide, necessitating continued exploration of alternative therapeutic agents. Plant-based resources have been increasingly exploited because of their pharmacological usefulness. This study aimed to comparatively evaluate the in vitro cytotoxic and gene-expression modulatory effects of glycyrrhizin (GL) and tamoxifen (TAM) in MCF-7 breast cancer cells, with particular emphasis on transcriptional changes associated with tumor suppressor and oncogenic pathways.
Methodology
MCF-7 cells were treated with increasing concentrations of GL and TAM, and cell viability was assessed using MTT assay to determine relative inhibitory concentrations (IC25, IC50, and IC75). Quantitative real-time PCR was employed to evaluate the expression of selected genes related to cell proliferation, apoptosis, and tumor suppression, including p53, PTEN, RAN, c-Myc, Bcl-2, stathmin, INFγ, hsp84, cyclin D1, and α-tubulin.
Results
Both GL and TAM demonstrated dose-dependent inhibition of MCF-7 cell viability, with tamoxifen exhibiting significantly greater cell inhibition potency, reflected by a lower IC50 (28.71 ± 2.97 µM) value compared to that of glycyrrhizin (37.70 ± 3.10 µM). Gene expression analysis revealed downregulation of several oncogenes, including RAN, c-Myc, Bcl-2, stathmin, INFγ, hsp84, cyclin D1, and α-tubulin, while upregulation of the tumor suppressor p53 gene. Notably, glycyrrhizin uniquely induced PTEN upregulation, suggesting a transcriptionally distinct mechanism compared to tamoxifen.
Conclusion
This exploratory in vitro study demonstrates that glycyrrhizin modulates key cancer-related gene expression pathways in MCF-7 cells, despite exhibiting lower cell inhibition potency than tamoxifen. The findings suggest that GL may exert mechanistically distinct transcriptional effects, particularly involving PTEN regulation. However, as this study is based solely on gene-expression data, further investigations incorporating protein-level validation, functional assays, and in vivo models are required to confirm these observations and assess translational relevance.
Introduction
Breast cancer remains one of the most frequently diagnosed malignancies worldwide, with approximately one in eight women at risk of developing invasive disease during their lifetime. 1 In 2022, an estimated 2.3 million new cases were reported globally, accounting for nearly 666,000 deaths. 2 Although historically more prevalent in industrialized nations, the incidence has increased substantially in developing countries, including Pakistan, which reported 34,038 new cases in 2012 and the highest breast cancer–related mortality rate (25.2%) among Asian countries. 3 These trends emphasize the growing global burden of breast cancer and the urgent need for more effective therapeutic and preventive strategies. 4
Breast cancer is a multifactorial disease resulting from complex interactions among genetic, hormonal, and environmental factors. 5 Recent evidence suggests that endocrine regulators beyond estrogen may contribute to breast tumorigenesis. Elevated levels of thyroid hormones, particularly fT3 and fT4, have been associated with increased breast cancer risk, especially in HER2-positive tumors. 6
At the molecular level, breast cancer progression is driven by dysregulated gene expression involving proto-oncogenes and tumor suppressor genes that promote uncontrolled proliferation, resistance to apoptosis, invasion, and metastasis.7,8 Recent genomic studies have identified additional breast cancer susceptibility genes beyond BRCA1/BRCA2, including CHEK2, PALB2, and TP53, expanding the genetic landscape of breast cancer risk. 9
Environmental factors may also influence tumorigenesis. Persistent pollutants such as polychlorinated biphenyls (PCBs) have been implicated in endocrine disruption and breast cancer development through modulation of oncogenic pathways including PI3K-Akt, MAPK, and HIF-1, involving genes such as BRCA1, AKT1, EGF, and IL6. 10
Diagnosis typically relies on clinical evaluation supported by imaging and histopathology. 11 Recent advances in artificial intelligence–assisted imaging have improved breast cancer diagnosis and treatment planning. Deep learning models analyzing tumor and axillary lymph node ultrasound images can accurately predict pathological response after neoadjuvant therapy, potentially reducing unnecessary axillary surgery. 12
Standard treatment strategies include surgery, chemotherapy, radiotherapy, and targeted therapies such as endocrine therapy for hormone receptor–positive disease. 13
Tamoxifen, a selective estrogen receptor modulator, has long been a cornerstone therapy for hormone-responsive breast cancer; however, prolonged treatment frequently leads to drug resistance. 14 Similarly, Aromatase inhibitor therapy in ER+/HER2+ tumors can induce early molecular responses, including reduced Ki67 proliferation and shifts from Luminal B to Luminal A subtype, which are associated with improved outcomes. 15
Dysregulated cell-cycle control is a hallmark of cancer. CDK4/6 inhibitors targeting the G1–S transition are effective in HR-positive/HER2-negative breast cancer, but resistance can arise through activation of MAPK/PI3K-AKT pathways and mutations such as NF1 that disrupt RAS signaling. 16
In addition to resistance, tamoxifen therapy has been associated with adverse effects including endometrial hyperplasia, thromboembolic complications, and hepatic dysfunction, which has prompted interest in alternative or adjunct therapeutic approaches. 17 Natural products and plant-derived compounds have therefore gained increasing attention as potential anticancer agents. Bioactive phytochemicals—including flavonoids, alkaloids, terpenoids, and polyphenols—can modulate multiple cellular processes involved in cancer progression, including oxidative stress, inflammation, and gene expression. 18
Several plant-derived metabolites, including epiberberine, crocin, evodiamine, and Halodule uninervis extracts, show antiproliferative and pro-apoptotic effects in cancer models, while nanoformulations may enhance their bioavailability and targeted delivery. 19
Among these natural compounds, Glycyrrhiza glabra (licorice root) and its principal bioactive constituent glycyrrhizin have attracted considerable interest due to their anti-inflammatory, antioxidant, and antiproliferative properties observed in several cancer models. 20 Glycyrrhizin has been reported to modulate tumor suppressor genes and oncogenic signaling pathways involved in cell survival and apoptosis and may enhance anticancer activity by promoting DNA damage, apoptosis, and G2-phase cell-cycle arrest. 21
The present study aimed to comparatively evaluate the in vitro antiproliferative and gene-expression modulatory effects of glycyrrhizin and tamoxifen in the MCF-7 breast cancer model. Specifically, the study investigated their effects on cell viability and the transcriptional regulation of selected oncogenes and tumor suppressor genes involved in breast cancer progression, with the objective of elucidating potential mechanistic differences and therapeutic implications.
Materials and Methods
Reagents and Chemicals
Cell culture media - RPMI 1640 (Gibco), 0.25% trypsin EDTA (Gibco), fetal bovine serum (FBS) (Gibco), Penicillin + Streptomycin (Gibco), dimethyl sulphoxide (sigma), doxorubicin (sigma), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide dye (MTT dye) (Gibco), glycyrrhizin (Sigma), tamoxifen (ICI), 95% ethanol (Sigma), phosphate buffer saline (Bioworld), TRIzol reagent (Sigma), Chloroform (Merck), Isopropyl Alcohol (Merck), Primers (Eurofins genomics), RNAse free water (ThermoScientific), methanol (Merck).
Cell Culture and Cell Line Maintenance
MCF-7 human breast cancer cells were obtained from ATCC (RRID: CVCL_0031). Cells were revived from cryopreservation and cultured in RPMI-1640 medium supplemented with 10% FBS and 1% Penicillin-Streptomycin under standard conditions (37°C, 5% CO2, humidified incubator). Cells were used within a limited number of passages after revival to minimize drift and phenotypic variation.
Mycoplasma contamination was monitored through routine microscopic examination during passaging, and RT-PCR–based testing was performed after initial acquisition and during long-term culture. Although no independent STR profiling was performed, reliance on an authenticated commercial source has been acknowledged.
Cell Viability Assay
For cell viability assessment, MCF-7 cells were seeded in 96-well plates at a density of 1 × 104 cells/well and allowed to adhere overnight. Cells were then treated with escalating concentrations of glycyrrhizin and tamoxifen (1.56 µM to 200 µM) for 48 hours. Positive control wells received Doxorubicin (1uM) as standard while untreated wells served as negative controls. All treatments were performed in six technical replicates per concentration.
After 48 hours of treatment, cell viability was evaluated using the MTT assay. Briefly, MTT reagent (5 mg/mL stock) was added to each well and incubated for 3–4 hours at 37°C to allow formation of formazan crystals. The supernatant was carefully removed, and the crystals were dissolved in DMSO. Absorbance was measured at 570 nm using a microplate reader and percentage inhibition was calculated relative to negative controls and expressed as mean ± SD. Concentrations corresponding to IC25, IC50, and IC75 were determined by nonlinear regression using GraphPad Prism.
RNA Isolation and RT-qPCR Analysis
For gene expression analysis, MCF-7 cells were seeded in 6-well plates at 1 × 106 cells/well and grown to 80–90% confluence. Culture medium was replaced with maintenance medium, and cells were treated with IC25, IC50, and IC75 concentrations of glycyrrhizin and tamoxifen for 48 hours. Control wells without drug treatment were included for comparative analysis, and each condition was performed in six replicates.
Sequence of Gene Specific Primers for qRT-PCR With Their Annealing Temperatures and Product Size
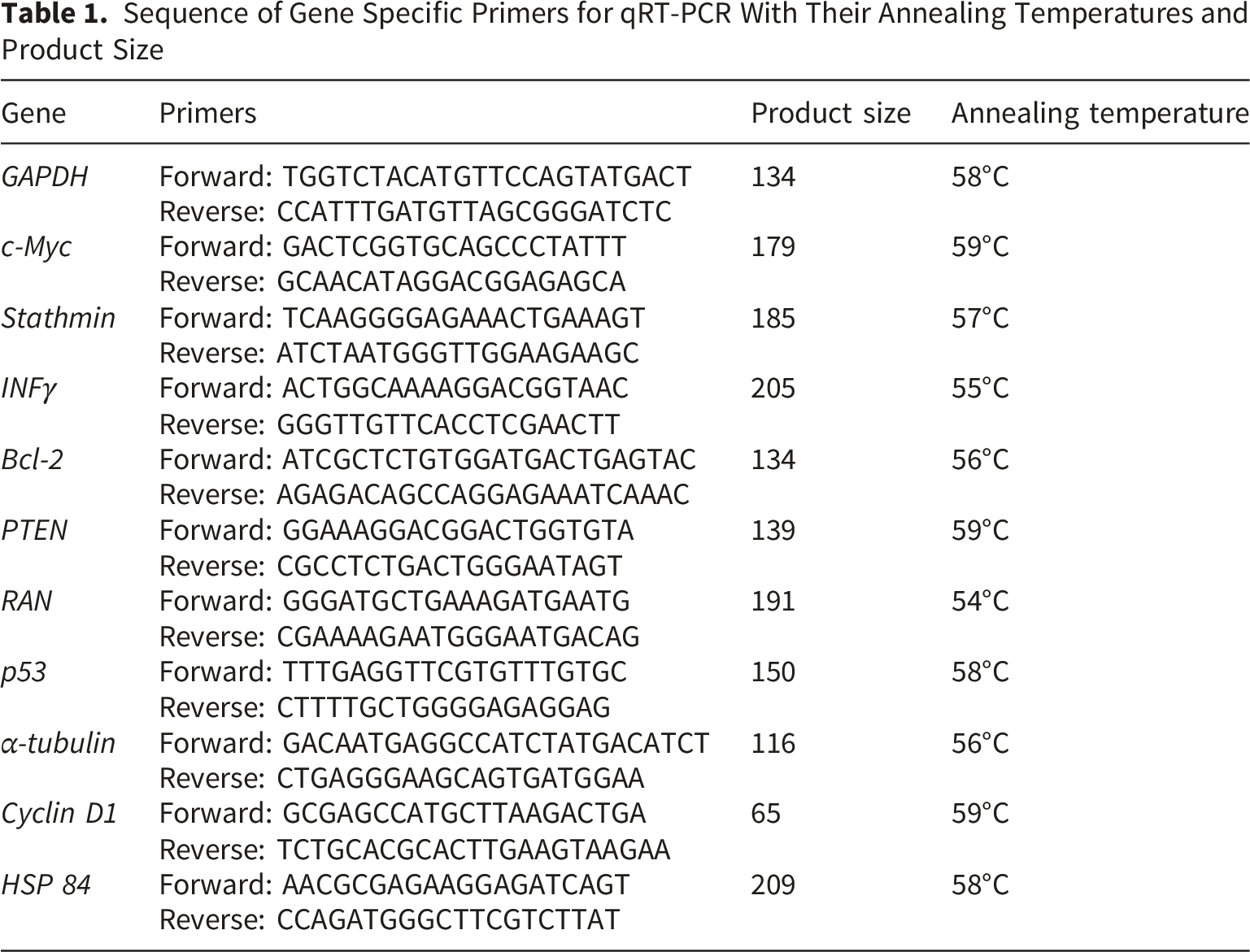
Quantitative real-time PCR was performed using SYBR Green chemistry on the Applied Biosystems QuantStudio™ 1 Real-Time PCR System. The amplification protocol included initial denaturation at 95°C for 10 minutes, followed by 40 cycles of denaturation (95°C for 20 seconds), annealing (Specific primer temperature for 40 seconds), and extension (72°C for 40 seconds).
Relative gene expression was calculated using the 2−ΔΔCT method, normalized to GAPDH as an internal control. Melt curve analysis was performed to confirm amplification specificity, and only reactions demonstrating a single peak were included in the final analysis.
Statistical Analysis
All statistical analyses were performed using Statistical Package for the Social Sciences (SPSS, version 24) and GraphPad Prism (version 6). Quantitative variables were expressed as mean ± standard deviation (SD). Treatment effects on cell viability and gene expression were analyzed using independent samples t-test and one way analysis of variance (ANOVA), followed by Tukey’s post hoc test to evaluate pairwise group differences. The normality of distributions for all dependent variables was assessed using the Kolmogorov–Smirnov/Shapiro–Wilk tests. Nonlinear regression analysis (log[inhibitor] vs. response) was used to estimate IC values and the results are reported with 95% confidence intervals (CI). A p-value ≤ 0.05 was considered statistically significant.
Results
Inhibitory Concentration (IC) Analysis of Glycyrrhizin and Tamoxifen
Both glycyrrhizin (GL) and tamoxifen (TAM) exhibited a concentration-dependent increase in the percentage inhibition of MCF-7 breast cancer cells across the tested range (1.56–200 µM). (Figure 1). The mean inhibitory concentrations (IC25, IC50, and IC75) were calculated by nonlinear regression using GraphPad Prism software. For glycyrrhizin, the IC25 was 9.64 ± 1.08 µM, IC50 was 37.70 ± 3.10 µM, and IC75 was 189.52 ± 4.79 µM. For tamoxifen, the IC25 was 4.92 ± 0.43 µM, IC50 was 28.71 ± 2.97 µM, and IC75 was 143.77 ± 6.34 µM. Statistical comparison using a t-test revealed that tamoxifen had significantly lower IC25 (p<0.01), IC50 (p<0.05), and IC75 (p<0.001) values compared to glycyrrhizin, indicating that tamoxifen is a more potent cytotoxic drug compared to glycyrrhizin. Percentage cell inhibition of MCF-7 cells in the presence of increasing log concentrations of glycyrrhizin and tamoxifen.
Tumor Suppressor Gene Expression
Both glycyrrhizin and tamoxifen significantly upregulated p53 expression in MCF-7 cells at all tested concentrations (IC25, IC50, IC75) compared to control (p<0.001). The highest expression was observed at IC75 for both drugs. No significant difference was found between IC25 of glycyrrhizin (1.66 ± 0.05 fold) and IC25 of tamoxifen (1.69 ± 0.06 fold), but significant increases (p<0.001) were observed when comparing higher doses within and between treatment groups (Figure 2A). Glycyrrhizin significantly upregulated PTEN expression in a concentration-dependent manner (p<0.001), whereas tamoxifen significantly downregulated PTEN expression at all concentrations compared to the control (p<0.001). The difference between glycyrrhizin and tamoxifen groups was highly significant at corresponding concentrations (Figure 2B). Comparison of effect of different inhibitory concentrations (IC25, IC50 and IC75) of glycyrrhizin (GL) and tamoxifen (Tam) on mRNA expression levels (fold change) of (A) p53 and (B) PTEN in MCF-7 cells. “a” symbolizes comparison of control with other groups. “b” symbolizes comparison of “GL IC25” with other groups. “c” symbolizes comparison of “Tam IC25” with other groups. “d” symbolizes comparison of “GL IC50” with other groups. “e” symbolizes comparison of “Tam IC50” with other groups. “f” symbolizes comparison of “GL IC75” with other groups. “a,b,c,d,e,f” symbolize p 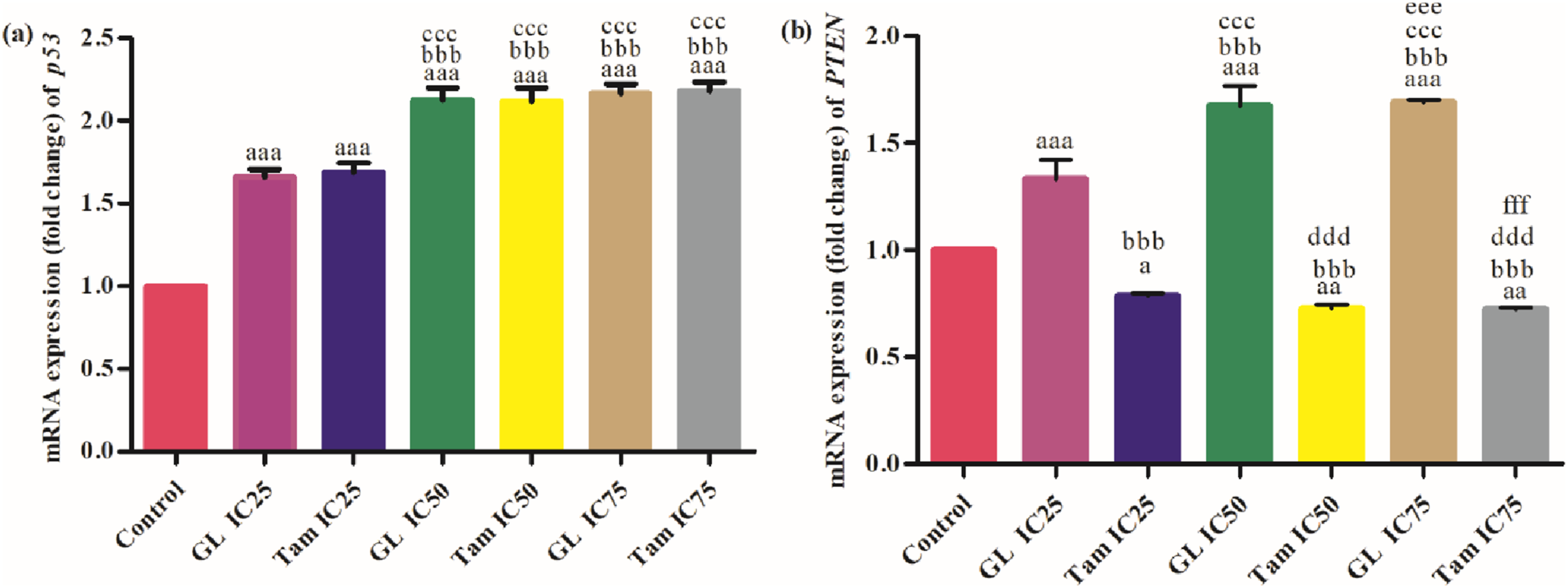
Oncogenes and Apoptosis-Related Genes
Treatment with both glycyrrhizin and tamoxifen resulted in significant downregulation of RAN mRNA levels (p<0.001). The strongest downregulation was observed at IC75 for both compounds. No significant difference was observed between IC25 of glycyrrhizin and IC25 of tamoxifen, but progressively stronger effects were seen at higher concentrations (Figure 3A). A significant (p<0.001) dose-dependent downregulation of c-Myc was observed in cells treated with both drugs. IC75 of glycyrrhizin (0.71 ± 0.01 fold) and tamoxifen (0.74 ± 0.01 fold) produced the strongest reductions. Statistically significant differences were noted between low and high concentrations of both agents (Figure 3B). Comparison of effect of different inhibitory concentrations (IC25, IC50 and IC75) of glycyrrhizin (GL) and tamoxifen (Tam) on mRNA expression levels (fold change) of (A) RAN, (B) c-Myc, (C) Bcl-2 and (D) INFγ in MCF-7 cells. “a” symbolizes comparison of control with other groups. “b” symbolizes comparison of “GL IC25” with other groups. “c” symbolizes comparison of “Tam IC25” with other groups. “d” symbolizes comparison of “GL IC50” with other groups. “e” symbolizes comparison of “Tam IC50” with other groups. “a,b,c,d,e” symbolize p 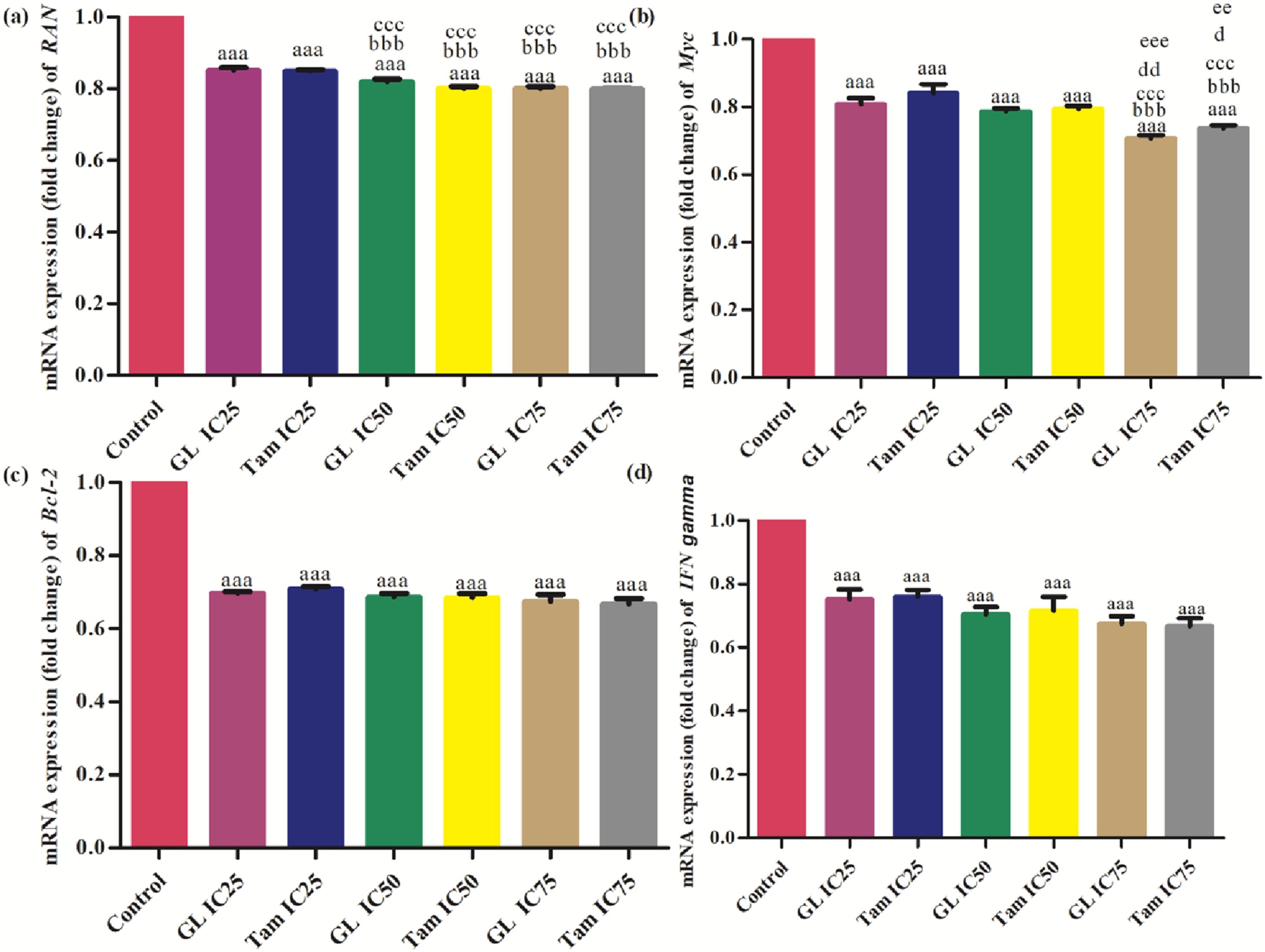
Both treatments significantly (p<0.001) reduced Bcl-2 expression compared to control. IC75 of glycyrrhizin (0.67 ± 0.02 fold) and tamoxifen (0.67 ± 0.02 fold) showed greater inhibition compared to their IC25 counterparts. Significant differences between groups were observed primarily at higher doses (Figure 3C). A consistent, significant (p<0.001) downregulation of INFγ was seen with both treatments. IC75 concentrations resulted in the most substantial reduction (0.67 ± 0.02 fold). IC25 of glycyrrhizin (0.75 ± 0.03 fold) was similar to IC25 of tamoxifen (0.76 ± 0.02 fold), but differences became more pronounced at higher doses (Figure 3D).
Cytoskeletal and Cell Cycle Regulatory Genes
Treatment with glycyrrhizin and tamoxifen significantly (p<0.001) downregulated Hsp84 expression. IC75 of tamoxifen (0.71 ± 0.01 fold) and glycyrrhizin (0.72 ± 0.01 fold) yielded the lowest expression levels. No significant difference was found between IC25 levels of both drugs, but inter-group comparisons at IC50 and IC75 showed statistically significant reductions (Figure 4A). A significant (p<0.001) decrease in stathmin expression was observed with increasing concentrations of glycyrrhizin and tamoxifen. At IC75, glycyrrhizin reduced stathmin expression to 0.64 ± 0.02 fold, while tamoxifen reduced it to 0.67 ± 0.02 fold. Significant inter-group differences were noted at all dose levels (Figure 4B). Comparison of effect of different inhibitory concentrations (IC25, IC50 and IC75) of glycyrrhizin (GL) and tamoxifen (Tam) on mRNA expression levels (fold change) of (A) heat shock protein 84, (B) stathmin, (C) cyclin D 1 and (D) α-tubulin in MCF-7 cells. “a” symbolizes comparison of control with other groups. “b” symbolizes comparison of “GL IC25” with other groups. “c” symbolizes comparison of “Tam IC25” with other groups. “d” symbolizes comparison of “GL IC50” with other groups. “a,b,c,d” symbolize p 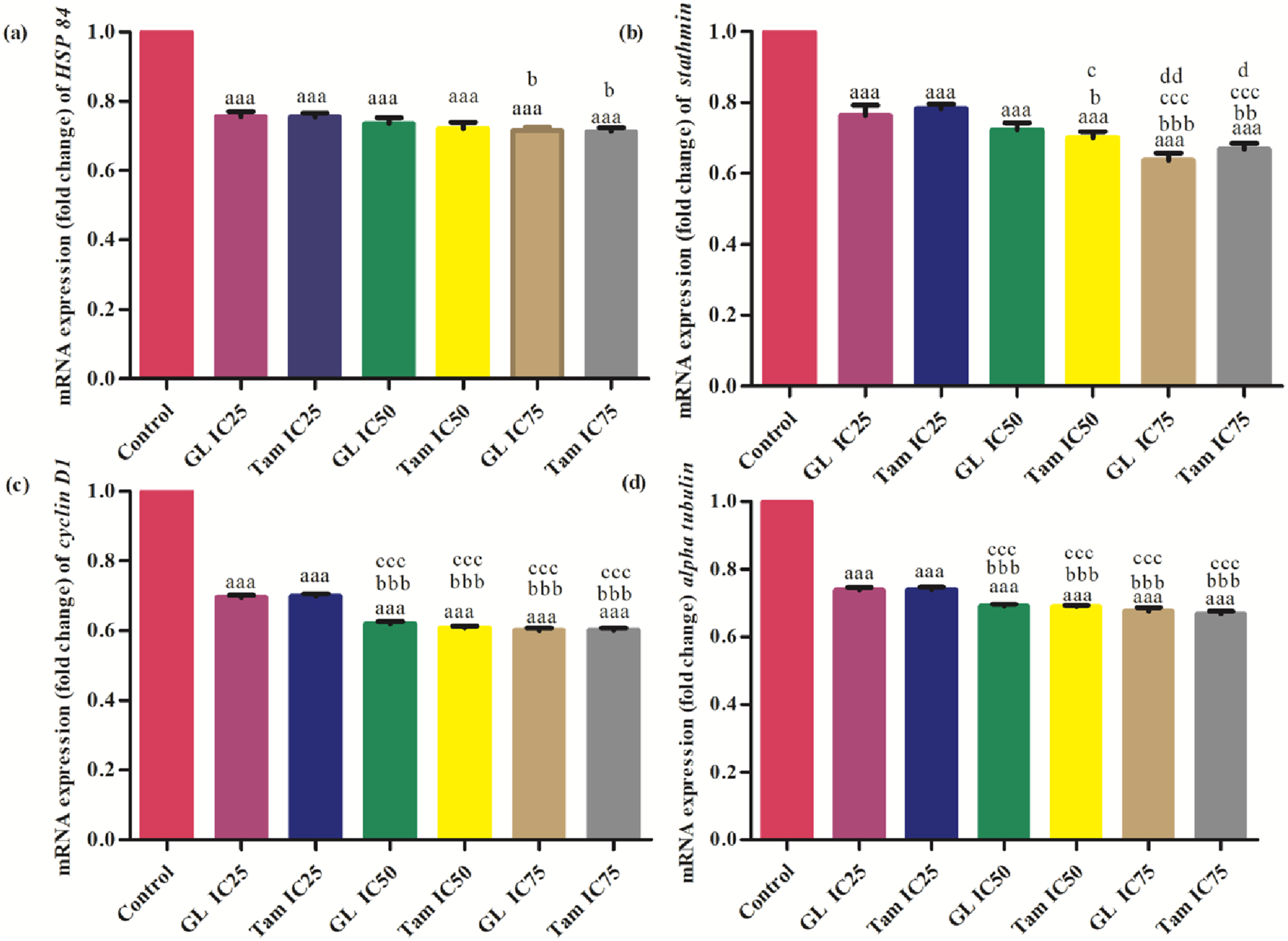
Both compounds significantly reduced cyclin D1 mRNA levels in a concentration-dependent manner (p<0.001). Glycyrrhizin at IC75 (0.60 ± 0.01 fold) and tamoxifen at IC75 (0.60 ± 0.01 fold) had the strongest effects. Differences between IC25 and IC75 treatments were statistically significant (Figure 4C). α-tubulin expression was significantly downregulated (p<0.001) by both glycyrrhizin and tamoxifen. IC75 of both agents (0.67 ± 0.01 fold) demonstrated maximum suppression. Inter-group comparisons showed a consistent trend of decreasing expression with increasing concentrations (Figure 4D).
Discussion
Breast cancer remains a leading cause of cancer-related morbidity and mortality among women worldwide, 2 and drug resistance and adverse effects associated with long-term tamoxifen therapy14,17 continue to drive interest in alternative or adjunct therapeutic agents. Natural compounds have attracted attention due to their potential anticancer effects and comparatively favorable safety profiles. 18 Glycyrrhizin has previously demonstrated anti-proliferative and pro-apoptotic properties across several malignancies, 20 prompting its evaluation as a potential transcriptionally active compound in breast cancer models.
The present study comparatively evaluated the cell viability and gene-expression modulatory effects of glycyrrhizin and tamoxifen in the MCF-7. Both agents demonstrated concentration-dependent inhibition of cell viability, with tamoxifen exhibiting lower IC25, IC50, and IC75 values, consistent with its established clinical potency.21,22 These findings confirm tamoxifen’s role as an appropriate pharmacological comparator and position glycyrrhizin as a compound with measurable, albeit less potent, inhibitory activity.
Beyond cell viability, the primary objective of this study was to examine transcriptional modulation of cancer-related genes. Rather than acting on a single molecular target, both compounds influenced multiple pathways associated with proliferation, apoptosis, and cell-cycle regulation, suggesting coordinated regulatory effects at the gene-expression level.
Upregulation of p53 observed with both treatments is biologically relevant given p53’s central role in DNA damage sensing, cell-cycle arrest, and apoptosis induction.23,24 Transcriptional activation of p53 may reflect cellular stress responses triggered by treatment exposure. However, as protein stabilization and post-translational modifications are critical determinants of p53 activity, the present findings indicate transcriptional engagement rather than confirmed functional activation.
A key distinguishing observation was the significant upregulation of PTEN following glycyrrhizin treatment, which was not comparably observed with tamoxifen. PTEN negatively regulates the PI3K/AKT survival pathway and is frequently downregulated in breast cancer.25,26 The selective enhancement of PTEN transcription by glycyrrhizin suggests potential engagement of growth-inhibitory signaling distinct from estrogen receptor antagonism. Given that tamoxifen primarily acts through estrogen receptor modulation, this divergence may reflect alternative upstream regulatory inputs, consistent with reports of differential downstream signaling responses to endocrine therapies.27,28 Although PI3K/AKT pathway activity was not directly measured, the transcriptional upregulation of PTEN represents a biologically meaningful finding aligned with tumor suppressor pathway restoration.
Suppression of RAN by both agents is noteworthy, as RAN plays a critical role in nuclear transport, mitotic spindle assembly, and chromosomal stability, and its overexpression correlates with tumor aggressiveness.29-31 Downregulation may therefore indicate impaired mitotic progression and reduced proliferative potential.
Downregulation of c-Myc and cyclin D1, two pivotal regulators of G1/S cell-cycle progression, suggests coordinated interference with proliferative signaling.32-36 In parallel, reduced expression of stathmin and α-tubulin indicates potential disruption of microtubule dynamics and mitotic spindle organization.37,38 As α-tubulin is a principal structural component of microtubules required for chromosome segregation, and its overexpression has been associated with aggressive and treatment-resistant breast cancer phenotypes,39-41 its transcriptional suppression—together with decreased stathmin—suggests coordinated modulation of cytoskeletal regulatory networks. Collectively, these transcriptional alterations imply convergence on cell-cycle and mitotic control pathways rather than isolated gene-specific effects. However, functional validation through cell-cycle or cytoskeletal assays would be required to confirm downstream biological consequences.
The decrease in Bcl-2 transcripts aligns with previously reported apoptosis-associated gene modulation following glycyrrhizin42,43 and tamoxifen44,45 treatment. Although functional apoptosis assays were not performed, reduced anti-apoptotic signaling at the mRNA level may contribute to the observed reduction in cell viability.
Heat shock protein modulation provides additional insight. Suppression of hsp84, encoding the β-subunit of Hsp90, may have broader implications, as Hsp90 stabilizes numerous oncogenic client proteins and supports malignant phenotypes.46,47 Transcriptional reduction could theoretically destabilize multiple signaling axes simultaneously, reinforcing the multi-targeted nature of glycyrrhizin’s activity.
Downregulation of IFN-γ also warrants consideration. While classically associated with antitumor immunity, IFN-γ can contribute to immune checkpoint induction and tumor-promoting inflammation under certain contexts.48-51 In a monoculture epithelial system such as MCF-7, the biological implications remain uncertain; nevertheless, the observed modulation suggests that glycyrrhizin may influence cytokine-associated transcriptional networks beyond direct proliferative pathways.
Collectively, the gene-expression profile observed following glycyrrhizin treatment indicates simultaneous modulation of tumor suppressor pathways (p53, PTEN), proliferative drivers (c-Myc, cyclin D1), survival regulators (Bcl-2), mitotic regulators (RAN, stathmin, α-tubulin), and stress-associated proteins (hsp84). When interpreted in an integrated manner, these changes suggest a coordinated shift toward reduced proliferative capacity and altered survival signaling. While tamoxifen exhibited greater potency, glycyrrhizin demonstrated a partially overlapping yet transcriptionally distinct profile, particularly in relation to PTEN regulation, consistent with the study’s comparative objective.
Importantly, these interpretations remain limited to mRNA-level measurements. Protein quantification, apoptosis assays, cell-cycle profiling, and pathway-specific analyses would be necessary to confirm whether the transcriptional alterations translate into functional biological outcomes.
From a translational standpoint, glycyrrhizin’s pharmacokinetic limitations—including poor oral bioavailability and potential adverse effects such as pseudoaldosteronism—must be considered in therapeutic development. In addition, the exclusive use of the MCF-7 luminal A, estrogen receptor–positive model provides relevance for hormone-responsive disease but limits generalizability. Evaluation in additional subtypes, such as triple-negative models, and incorporation of in vivo systems will be essential to determine whether the transcriptional effects observed here extend across broader breast cancer contexts.
Conclusion
In this exploratory in vitro study, glycyrrhizin and tamoxifen were comparatively evaluated for their effects on cell viability and gene expression in the MCF-7. Both agents demonstrated concentration-dependent cell inhibition, with tamoxifen exhibiting greater potency. However, glycyrrhizin produced a distinct pattern of transcriptional modulation, including upregulation of the tumor suppressor gene PTEN, alongside regulation of other cancer-related genes.
These findings indicate that although glycyrrhizin shows less cell inhibition than tamoxifen under the tested conditions, it differentially modulates molecular pathways relevant to breast cancer biology. As the present study is limited to mRNA-level analysis in a single in vitro model, the results should be interpreted as preliminary and hypothesis-generating. Further investigations incorporating protein-level validation and functional assays are required to determine the biological and therapeutic significance of these transcriptional effects.
Footnotes
Ethical Considerations
This study was approved by the Ethical Review Committee (UHS/REG-19/ERC/2812) of the University of Health Sciences, Lahore (Pakistan). Cell line used in this study was commercially obtained from certified cell repositories; therefore, no additional ethical approval was required.
Author Contributions
MA and MS designed the study. All authors contributed to the collection of data. MA, UA and TA conducted the statistical analysis. MA drafted the first version of the manuscript and SA and TA contributed to subsequent versions. Final manuscript is read and approved by all authors. The authors declare that all data were generated in-house and that no paper mill was used.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data used to support the findings of this study is available from the corresponding author upon request.