
Abstract
Introduction
Chronic inflammatory conditions represent a major global health priority and are closely associated with oxidative stress and metabolic dysfunction. Traditional medicinal plants are known to produce bioactive compounds with diverse properties, including anti-inflammatory activity. This study explored the biological effects of Gnetum parvifolium (G. parvifolium) extract and its main compound, isorhapontigenin (ISO), focusing on inflammation, oxidative stress, cell viability, and adipose cell differentiation.
Methods
HPLC and LC-MS analysis were applied for the identification and quantification of ISO in G. parvifolium extract. Blood leucocytes were exposed to varying concentrations of the extract and ISO to assess the ROS production and cell viability. In parallel, the effect of the extract and ISO on pro-inflammatory cytokine production and markers of adipose cell differentiation were evaluated using ProcartaPlex™ Immunoassays and qRT-PCR analysis.
Results
Treatment with G. parvifolium extract resulted in a significant reduction of intracellular ROS levels (IC50 = 13.9 µg/mL) without affecting leukocyte viability. The extract strongly suppressed the production and gene expression of major pro-inflammatory cytokines, including IFN-γ, IL-12, IL-17a, and IL-2. It also induced a selective downregulation of the adipocyte fatty acid–binding protein aP2, while other markers of adipose cell differentiation remained unchanged. In parallel, ISO markedly decreased ROS generation (IC50 = 7.9 µM) without cytotoxic effects. ISO significantly reduced the secretion of IFN-γ, IL-17a, and IL-2 and was associated with decreased expression of IL-17a, IL-2, and NOX2 genes. Like the plant extract, ISO selectively lowered aP2 expression without significantly modifying other adipogenic markers.
Conclusion
Taken together, these findings demonstrate that G. parvifolium extract and ISO exert combined antioxidant and anti-inflammatory effects while maintaining cell viability and selectively modulating adipose differentiation. Collectively, these effects point to a potential role G. parvifolium extract and ISO in the regulation of biological pathways linked to chronic inflammatory diseases.
Keywords
Introduction
Chronic inflammatory diseases are widely recognized as a leading cause of mortality worldwide. 1 The inflammatory process and immune system are involved in the genesis of the main chronic diseases such as stroke, chronic respiratory diseases, heart disorders, cancer, obesity, and diabetes. Currently, inflammation-related diseases are responsible for 50% of worldwide mortality. 1 The link between inflammation and the development of chronic diseases is explained by the release of pro-inflammatory cytokines such as interferon (INF)-γ, interleukin (IL)-6, IL-12, IL-17, tumor necrosis factor-alpha (TNF-α) 2 and the activation of fatty acid metabolism. 3 The latter stimulates the upregulation of cyclooxygenase-2 (COX-2) expression inducing further secretion of pro-inflammatory cytokines. 4 In addition, the cytokines released during inflammatory process promote the production of reactive oxygen species (ROS), resulting in oxidative stress, which aggravates the development of chronic diseases. 5 Recent research indicates that the simultaneous release of pro- and anti-inflammatory cytokines is essential for a balanced immune response. 6 The blockage of cytokine-mediated inflammatory signaling pathways, such as TNF-α and IFN-γ demonstrated benefit both in treatment of COVID-19 and autoinflammatory diseases. 7
Adipose tissue, included pre-adipocytes, macrophages, endothelial cells, fibroblasts, and leucocytes, is also involved in inflammatory process and metabolic regulation. Adipose tissue releases several adipokines in central and peripheral tissues as TNFα and interleukins. 8 Adipogenesis is a process which consists in the differentiation of preadipocytes into mature adipocytes through the secretion of adipokines, represented mainly by peroxisome proliferator-activated receptor gamma (PPARγ). 9
Isorhapontigenin (ISO) is found in various species of G. parvifolium, a plant from southern Asia, used in traditional chinese medicine to cure rheumatism, painful limbs, acute respiratory infections, and chronic bronchitis regarding to its antioxidant, anticancer, and antibacterial properties.10,11 ISO (Figure 1) has a structure close to resveratrol, a molecule particularly present in grapes in response to stress, and whose effects are no longer to be proven: prevention of cardiovascular diseases, stimulation of the immune system, regulation of the release of pro-inflammatory cytokines, etc.
12
Isorhapontigenin (ISO)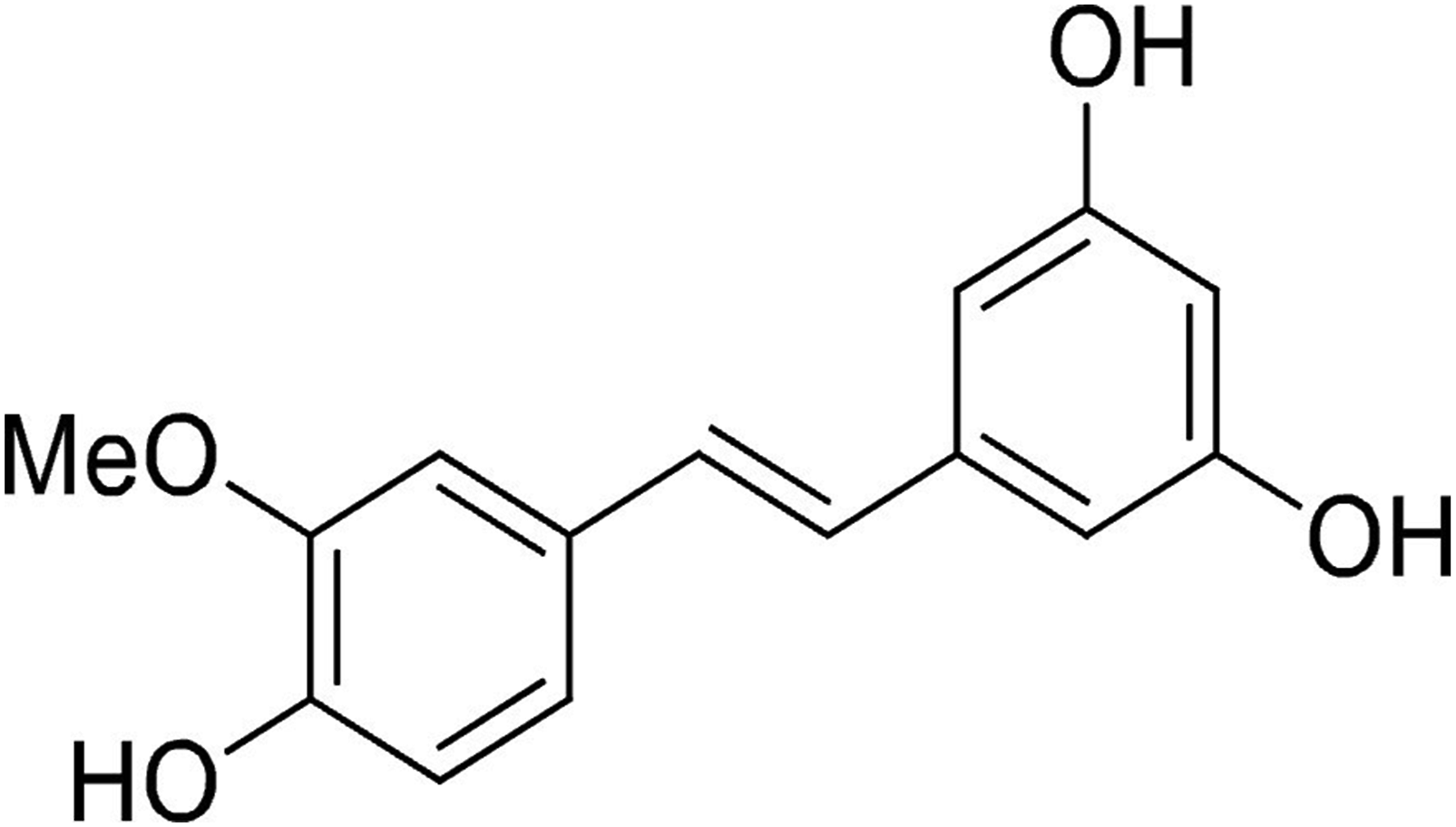
The biological effects of ISO have been extensively studied and summarized in a recent paper. 12 Among its health benefits, it is interesting to mention its anti-inflammatory properties. Indeed, recent studies demonstrated that ISO could suppress IL-1β-induced inflammation and reduced significantly nitric oxide (NO), inducible nitric oxide synthase (iNOS), Prostaglandin E2 (PGE2) COX-2 productions and would be effective in the regulation of markers of inflammation as TNF-α. 13 ISO is also reported to be active against ROS production, by increasing the stimulation of antioxidant enzymes as superoxide dismutase (SOD) 14 and by trapping for hydroxyl (•OH) and hydroperoxyl (•OOH) radicals. Furthermore, ISO is involved in the inhibition of lipogenesis and activation of lipolysis. 15
Although the anti-inflammatory potential of ISO has been previously reported, this study aims to evaluate and compare the effects of G. parvifolium extract and its bioactive constituent, ISO. We specifically explore their ability to modulate inflammatory cytokines, reduce oxidative stress, and influence adipocyte differentiation while preserving cell viability, offering new insights for the management of chronic inflammatory diseases.
Materials and Methods
Plant Extraction
G. parvifolium plant was collected from Guangxi province, in the Southern China, and referenced under G. parvifolium (Warburg) W. C. Cheng, Acta Phytotax. Sin. 9: 386. 1964, in the herbarium Flora of China. G. parvifolium was ground into pieces between 0.1 and 1 cm. A total of 100 g of plant material was subjected to extraction with 2,000 mL of an ethanol–water mixture (50:50, v/v) while stirring at 120 rpm for 4 h at room temperature. The extract was separated from the solid phase by filtration and subsequently evaporated under vacuum at 40 °C to obtain a powdered extract.
Chemicals and Reagents
A 10 mM of ISO (Sigma Aldrich, St. Louis, MO, USA) and G. parvifolium extract (200 mg/mL) stock solutions were prepared in DMSO (Dimethyl sulfoxide, ThermoFisher Scientific, Waltham, MA, USA), aliquoted and then frozen at -20°C. After thawing, dilutions at 1, 5, 10 and 25 µM for ISO and 10, 25, 50 and 100 µg/mL for G. parvifolium extract were prepared in Roswell Park Memorial Institute 1640 Medium (RPMI 1640, Gibco, Life Technologies, Carlsbad, California, USA).
HPLC-Analysis
ISO identification and quantification were carried out using HPLC coupled to a diode-array detector operating between 190 and 800 nm (Agilent, USA). Chromatographic separation was performed on a Luna C18 analytical column (250 × 4.6 mm, 5 µm; Phenomenex) with a C18 pre-column at 20 °C and a constant flow rate of 1 mL/min. The mobile phase was a mixture of 0.1% (v/v) acidified water (phase A) and 0.1% (v/v) acetonitrile (phase B). The gradient of phase A was 80% (0 min), 20% (60 min), 10% (61 min), 10% (66 min), 80% (67 min) and 80% (75 min). The injection volume was 10 µL and sample concentration was 5 mg/mL prepared in methanol. HPLC chromatogram of the sample was acquired at 310 nm.
LC-MS Analysis
LC-MS analysis was performed using a UHPLC Ultimate 3000 RSLC system coupled with an Orbitrap Q-Exactive mass spectrometer (Thermo Fisher Scientific Inc., Waltham, MA, USA). Separation was achieved on an Uptisphere C18-3 column (250×4.6 mm, 5 µm; Interchim, Montluçon, France). Samples were prepared in HPLC-grade methanol (1 mg/mL), filtered through 0.45 µm PTFE filters, and 10 µL was injected. The mobile phase consisted of 0.1% (v/v) formic acid in water (Phase A) and acetonitrile (Phase B). The gradient for Phase A was: 100% (0 min), 80% (10 min), 73% (35 min), 0% (90 min), and returning to 100% (110 min). The flow rate was maintained at 0.8 mL/min.
Cell Culture
Blood Leukocyte Preparation
Human blood samples were obtained from healthy volunteers (n = 3) through the Établissement Français du Sang (EFS, Clermont-Ferrand, France), with written informed consent provided by all donors. This study was conducted under EFS contract no. EFS AURA 25-152, in strict accordance with the French Public Health Code (articles L1222-1, L1222-8, L1243-4, and R1243-61). Peripheral blood leukocytes were isolated via ammonium chloride-induced hemolytic shock using a lysis buffer (NH4Cl 155 μM, NaHCO3 12 μM, and EDTA 0.01 μM). Following lysis, the leukocyte fraction was recovered by centrifugation and resuspended in RPMI 1640 medium (Eurobio Scientific, Saclay, France) supplemented with 50 μg/mL gentamicin and 2 mM L-glutamine (ThermoFisher Scientific).
PBMCs Preparation
Peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats obtained from healthy volunteers (n=3–6). Blood was diluted and carefully layered onto a Ficoll–Histopaque 1077® (Sigma-Aldrich) gradient. After centrifugation at 400×g for 40 min at 20°C, the first layer of plasma was aspirated, yielding a phase of monocytes and lymphocytes ( PBMCs) just above the 1.077 g/mL layer. The cells were washed twice in RPMI 1640 medium, centrifuged at 400×g for 5 min and finally resuspended in complete culture medium consisting of RPMI 1640 supplemented with 10% FBS, 50 μg/mL gentamicin, and 2 mM L-glutamine. The concentration was adjusted to 106 cells/mL for all assays.
Adipose Cells
The murine 3T3-L1 preadipocytes were obtained from the American Type Culture Collection (ATCC No. CL-173. Molsheim, France) and seeded at confluence (33,500 cells/cm2) for differentiation into mature adipocytes in a Dulbecco’s modified Eagle medium (DMEM/F12 (1:1), Gibco) supplemented with 10% FBS, 2mM Gln, gentamycin (50µg/mL), insulin (1µg/mL), dexamethazone (0.25µM), rosiglitazone (0.1µM), isobutyl-methylxanthine IBMX (0.5mM) for 3 days. All the cells used were cultured in a 5% CO2-humidified incubator at 37°C.
Production of ROS by Leukocytes in the Presence of ISO and G. Parvifolium Extract
The leukocyte preparation was adjusted to 106 cells/mL and seeded in 96-well plates. Cells were incubated with ISO (1, 5, 10, and 25 μM) or G. parvifolium extract (10, 25, 50, and 100 μg/mL), as previously described by Cholet et al. 16 The production of ROS by immune cells in response to PMA (phorbol 12-myristate 13-acetate) stimulation (1 µM) for 120 min was measured by the oxidation technique of dihydrorhodamine 123 (1 µM) (DHR123, Sigma Aldrich). DHR123 is a stable non-fluorescent compound that, once inside cells, is oxidized by endogenous ROS to rhodamine 123. Fluorescence was measured using a Tecan Spark® multimode reader (Männedorf, Switzerland). ROS production was expressed as a percentage relative to stimulated untreated control cells (100%).
Cell Viability Study
The cell suspension was adjusted to 106 cells/mL in supplemented RPMI medium (10% FBS, 50 μg/mL gentamicin, and 2 mM L-glutamine) and seeded in 96-well plates. Cells were incubated with G. parvifolium extract or ISO, then treated with PMA (1 μM) and resazurin (25 μg/mL). Fluorescence (excitation/emission:544/590 nm) was measured every 30 min for 2 h using a Tecan Spark® microplate reader (Männedorf, Switzerland).
Determination of the Anti-Inflammatory Activity
Effects of G. parvifolium extract and ISO on cytokine production were studied by Luminex Bio-Plex 200 System (Bio-Rad, France). PBMCs (106 Cells/mL) (n=3 volunteers) were incubated in the presence or not of 5 μg/mL of phytohemagglutinin (PHA, Sigma Aldrich) and treated for 24h. The concentrations of 50μg/mL for the extract and 5 μM for ISO were selected following results obtained in ROS production. The supernatants were taken and frozen until used. A ProcartaPlex Multiplex Immnoassay (ThermoFisher Scientific) kit was used for cytokine assay All samples were run in triplicate and were assayed for 13 human cytokines (INFγ, TNF-α, IL-1β, IL-2, IL-4, IL-6, IL-8, IL-10, IL-12, IL-13, IL-17a, IL-23 and MIP-1α). Cytokine levels were quantified following the manufacturer’s instructions. Data were analyzed with Bio-Plex Manager 4.1 software using a five-parameter logistic (5PL) regression. All assays were performed in triplicate.
Evaluation of Gene Expression by RT-qPCR
PCR Primer Sequences

Statistical Analysis
Statistical analyses were performed using GraphPad Prism v8 (GraphPad Software, USA). Differences between groups were assessed via Student’s t-test. Data are expressed as mean ± SEM of 3–6 independent experiments. P-values< 0.05 (*), < 0.001 (**), and < 0.0001 (***) were considered statistically significant.
Results
HPLC and LC-MS Analysis
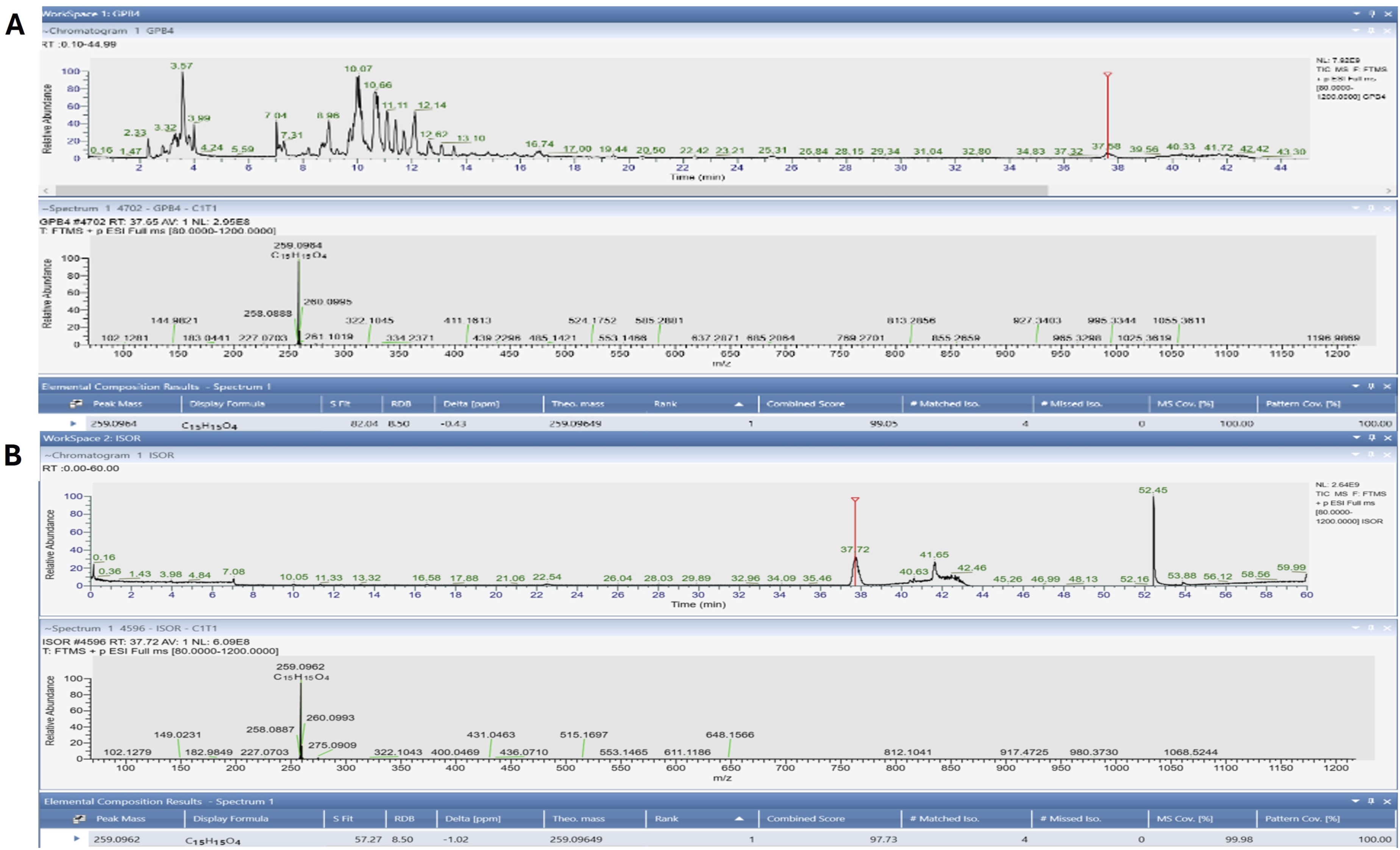
HPLC was applied for the identification and quantification of ISO in G. parvifolium extract (Figure 2) and revealed the presence of ISO (0,014%m/m) at 17.64 min. LC-MS supported the presence of ISO in G. parvifolium extract and demonstrated the peak of ISO at 37.58 min, what agrees with the spectrum of the standard where the ISO appeared in 37.72 min (Figure 3). The HPLC chromatogram of Gnetum parvifolium was acquired at 310 nm LC-MS chromatograms and MS spectrum of Gnetum parvifolium methanolic extract (A) and ISO (B)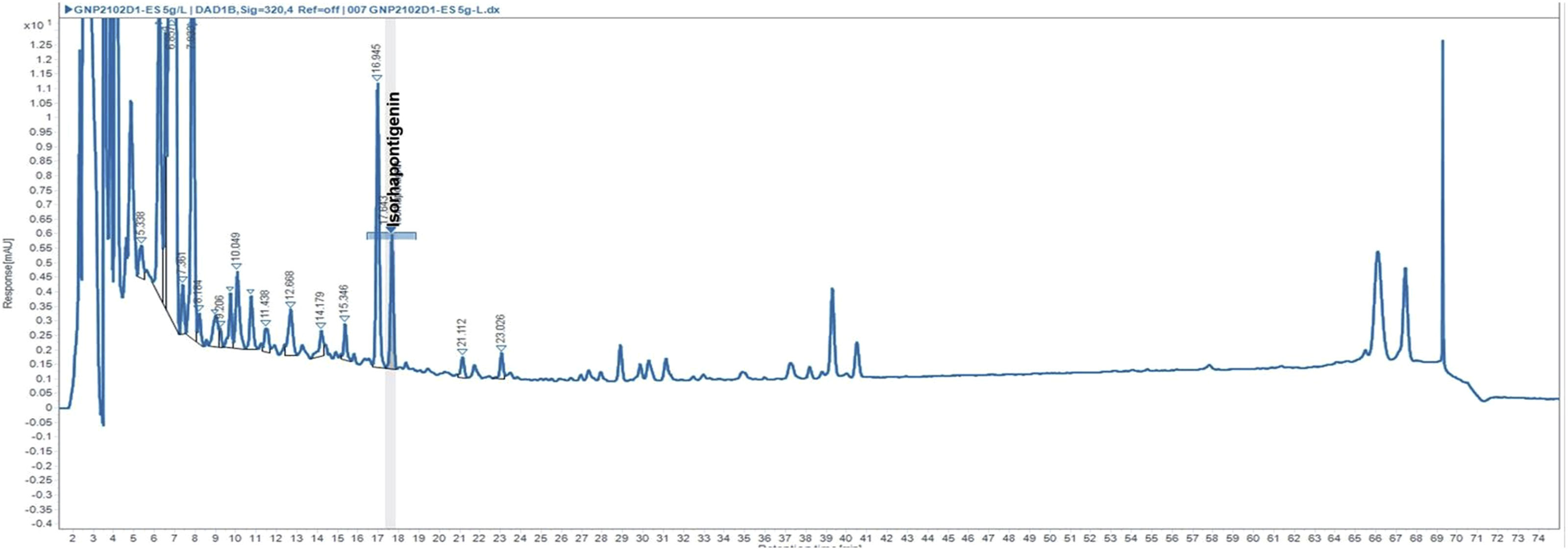
Modulation of Leukocyte ROS Production by G. Parvifolium and ISO
To determine the antioxidant effects of G. parvifolium and ISO on ROS production, blood leukocytes were firstly triggered by PMA, resulting in a significant increase in ROS production after 1h of incubation. Following the addition of G. parvifolium extract, ROS production by blood leukocytes significantly decreased after 30 minutes of exposure at all concentrations tested (Figure 4A). After 2 h, a significant reduction of ROS production was observed reaching 45%, 59%, 59% and 61% of inhibition at 10, 25, 50 and 100 µg/mL, respectively. Antioxidant potential of ISO in blood leukocytes (Figure 4B) demonstrated an antioxidant activity (IC50=7,9 µM). (A) Kinetics of ROS production by blood leukocytes stimulated by PMA in the presence of different concentrations of Gnetum parvifolium extract for 2h. (B) ROS production by blood leukocytes, incubated with ISO (1, 5, 10, 25 µM) and stimulated with PMA for 2 h. Data are expressed as standard mean ± SEM and significance was indicated as *, ** and *** where p ≤0.05, 0.001 and 0.0001 respectively. n =3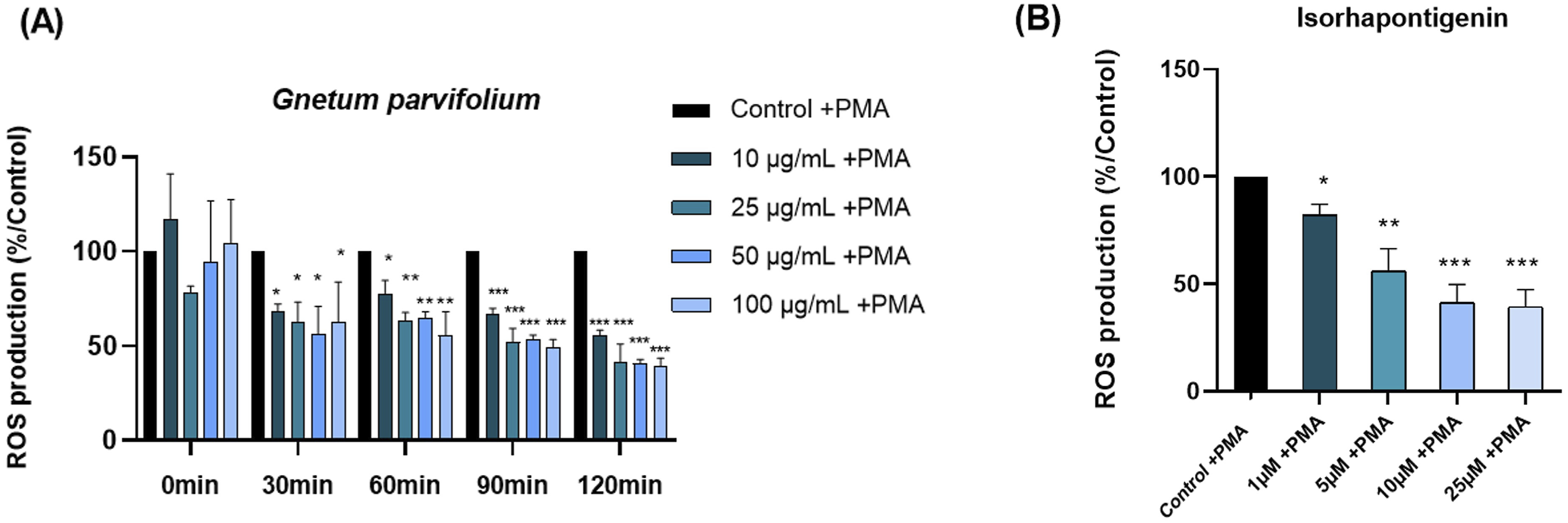
Effects of G. Parvifolium and ISO on Blood Leukocyte Viability
Results indicated that G. parvifolium extract and ISO had no impact on blood leukocyte viability at all plant extract (Figure 5A) or ISO (Figure 5B) concentrations after 2 hours of treatment. So, the inhibition of ROS production was not correlated with viability. Percentage of cell viability of blood leukocytes in the presence of (A) Gnetum parvifolium extract (10-100 µg/mL) for 2h or (B) ISO (1-25 µM) after 2h of incubation. The results are expressed as standard mean ± SEM. n=3-6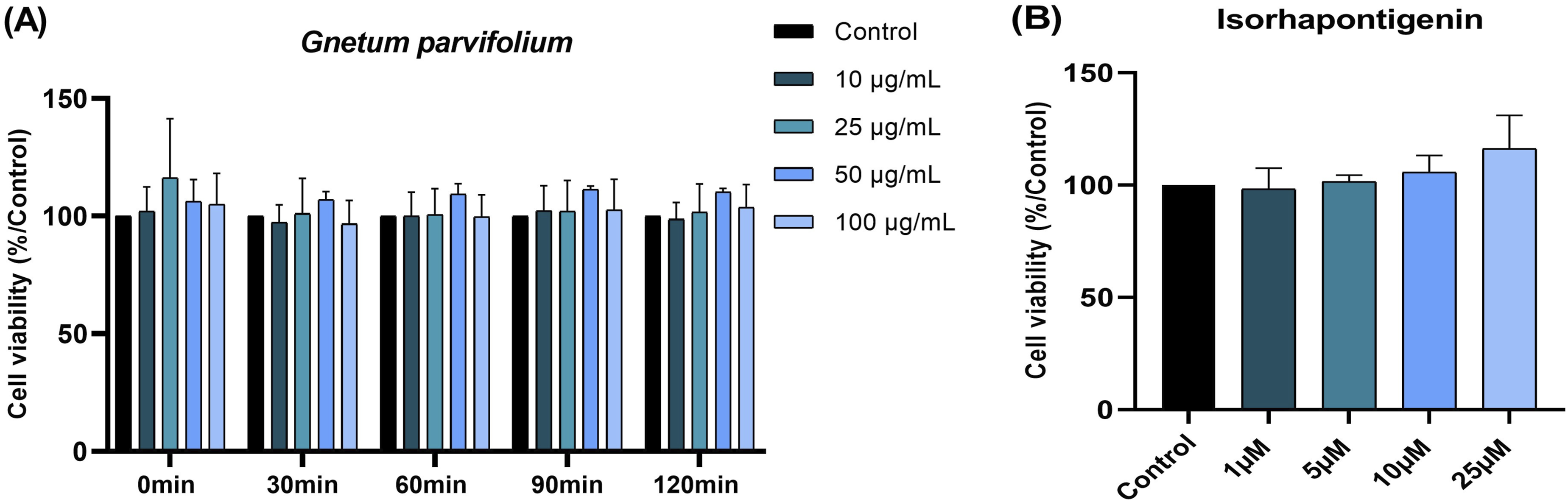
Effects of G. Parvifolium and ISO on the Inflammatory Response
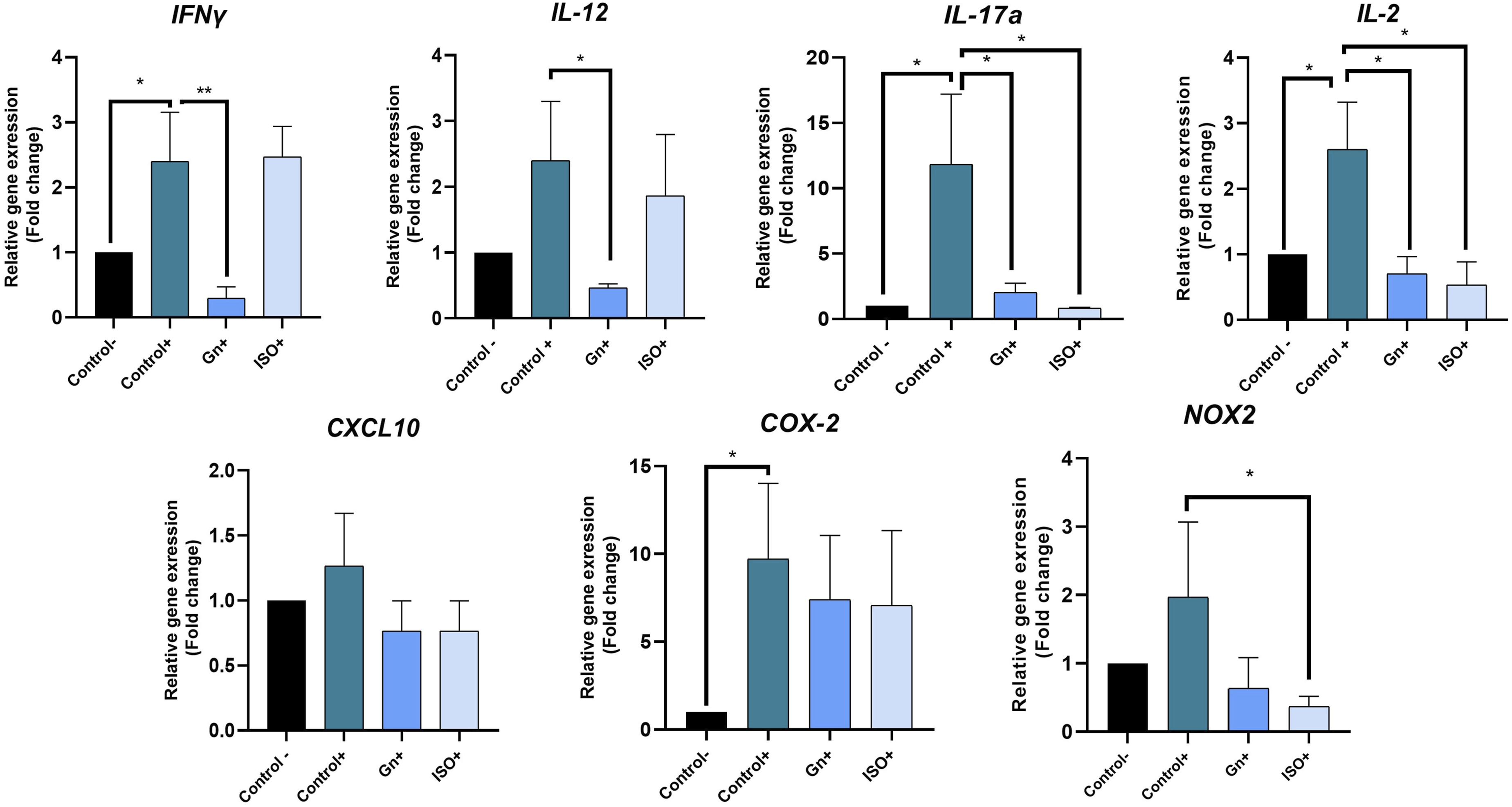
Considering the results above, the concentrations of 50 μg/mL of G. parvifolium extract and 5 µM of ISO were selected for the determination of the inflammatory response. PBMCs were stimulated by PHA in the presence or absence of the tested entity, and the concentration of thirteen cytokines secreted in the supernatant was measured with ProcartaPlex™ Immunoassays.. G. parvifolium extract significantly decreased the secretion of pro-inflammatory cytokines such as IFN-γ (-95%, p<0.0001), IL-12 (-63%, p<0.0001), IL-17a (-87%, p<0.0001) and IL-2 (-56%, p<0.001) (Figure 6A). Similarly, ISO revealed a potent anti-inflammatory effect by the reduction of INF-γ (-94%, p<0.0001), IL-17a (-92.5%, p<0.0001) and IL-2 (-47.4%, p<0.05) (Figure 6B). Furthermore, the treatment with G. parvifolium extract, as well as ISO, significantly decreased some anti-inflammatory cytokines as IL-13 and IL-10 (Figure 6A and B). Consistent with results below, qPCR analysis revealed a significant decrease of INF-γ, IL-12, IL-17a and IL-2 gene expression after 24h of treatment with G. parvifolium extract (0.30±0.10, p<0.001; 0.47±0.01, p<0.05; 2.03±0.03, p<0.05; 0.70±0.23, p<0.05, respectively), compared to the positive control. Significant reduction was also showed with ISO on IL-17a and IL-2 gene expression (0.83± 0.01, p<0.05; 0,53 ± 0.38, p<0.05, respectively) (Figure 7). Furthermore, ISO significantly downregulated NOX2 gene expression (0.4±0.1 p<0.05), a key oxidative effector (Figure 7). Protein inhibition of INF-γ and IL-12 by ISO, without significant changes in gene, could be related to the return of normal gene expression after 24h of treatment. Moreover, no significant modification was observed on CXCL10 and COX-2, both with G. parvifolium extract and ISO. Impact of Gnetum parvifolium extract (50 µg/mL) (A) and ISO (µM) (B) on cytokine secretion profiles in PHA-stimulated PBMCs. The levels of pro-inflammatory cytokines (IFN-γ, IL-1β, IL-12, IL-17a, IL-2, IL-23, IL-6, IL-8, MIP-1α, and TNF-α) and anti-inflammatory cytokines (IL-10, IL-13, and IL-4) were measured following 24 h of stimulation with PHA (5 μg/mL) (n = 3). Data are presented as mean ± SEM. Statistical significance is indicated as *p ≤ 0.05, **p ≤ 0.001, or ***p ≤ 0.0001 compared to the stimulated control Relative gene expression of IFNγ, IL-2, IL-12, IL-17a, CXCL10, COX-2 and NOX2 by RT-qPCR in cells treated or not with 50 μg/mL of Gnetum parvifolium extract or 5 µM ISO and stimulated with LPS/IFN-γ. Results were expressed as mean ± SEM. Significance was indicated as * when p≤0.05 or ** p≤0.001 in comparison to positive control. n=3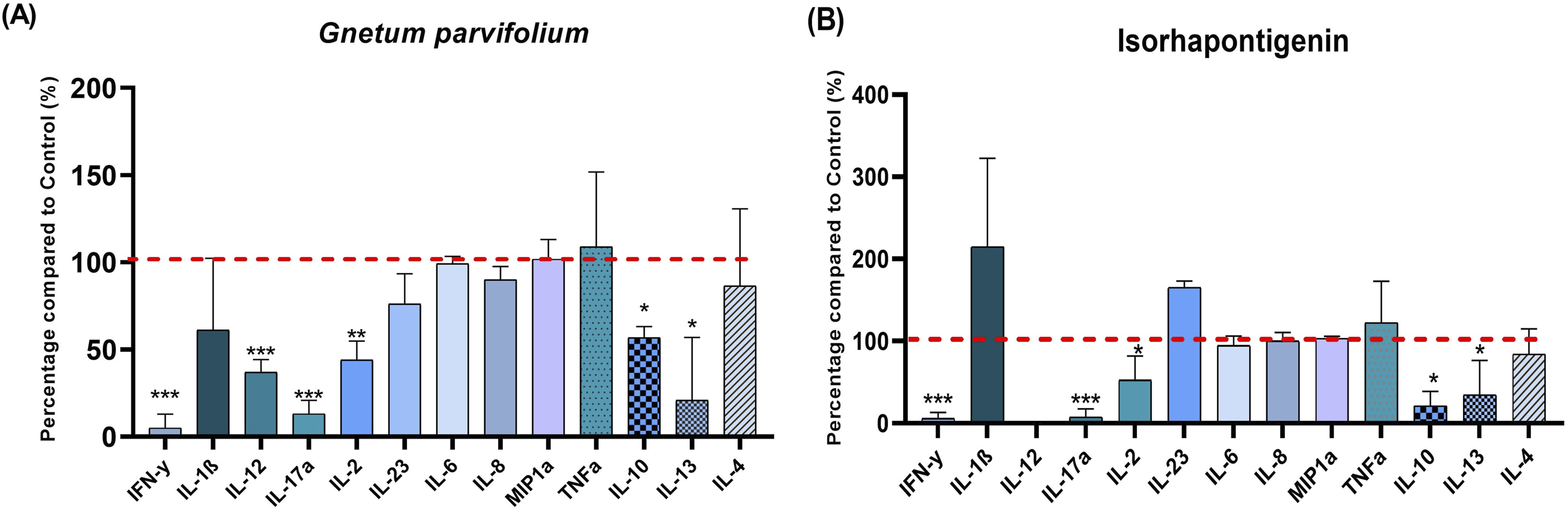
Effects of G. Parvifolium and ISO on Markers of Adipose Cell Differentiation
The secretion of adipokines and inflammatory response constitute a major factor in the development of obesity. For this purpose, we analyzed the gene expression of main markers involved in adipose cell differentiation (aP2, pref1, PPARγ, HSL, Adiponectin and AdipoR1) in the presence or absence of G. parvifolium extract (50µg/mL) or ISO (5µM) for 24h. Surprisingly, as showed in Figure 8A and B, significant decrease was observed only on adipocyte fatty acid binding protein (aP2) with G. parvifolium extract (0.77±0.1, p< 0.05) or ISO (0.6 ±0.1, p< 0.05). Indeed, these treatments failed to reduce the gene expression of Pref1, peroxisome proliferator-activated receptor γ (PPARγ), the hormone-sensitive lipase (HSL), adiponectin expression and its receptor AdipoR1. Effects of Gnetum parvifolium extract (50µg/mL) and ISO (5µM) on gene expression of aP2, Pref1, PPARγ, HSL, Adiponectin and AdipoR1 by qRT-PCR analysis. Experiments were repeated in triplicate and data shown as mean±SEM. Significance was indicated *≤0.05 compared with the control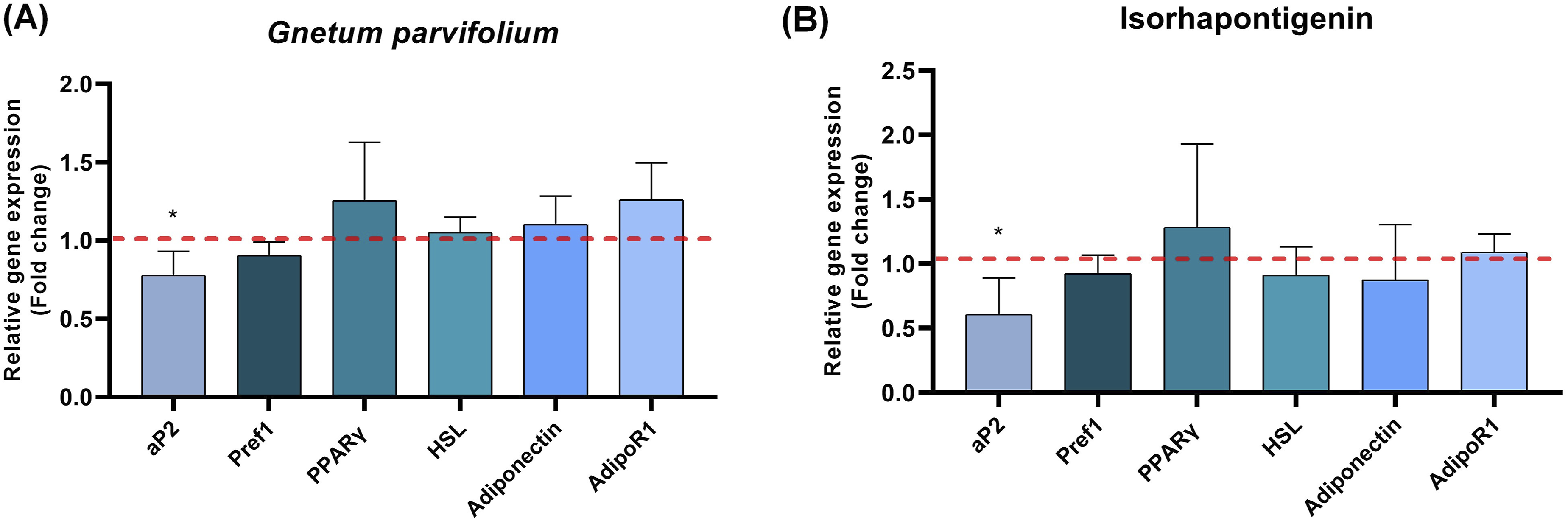
Discussions
The objective of this study was to evaluate the pharmacological potential of G. parvifolium extract and its major constituent, ISO, regarding their antioxidant and anti-inflammatory properties, as well as their impact on adipocyte differentiation. Our findings demonstrate that these compounds exert significant biological activities without compromising cell viability.
Antioxidant Mechanisms and Cellular Safety
A previous study confirmed the presence of ISO in the G. parvifolium extract, characterized by a wide UV absorbance at approximately 310 nm at 18.2 min. 11 The G. parvifolium extract’s robust antioxidant profile is attributed to its high content of polyphenols, namely piceatannol (1.7mg/g), rhaponiticin (60.1 mg/g), resveratrol (37.4 mg/g), and ISO (184.0 mg/g). These compounds are known to inhibit xanthine oxidase (XO) activity and effectively scavenge NO and NO2 radicals through hydrogen atom transfer.11,15,17 Furthermore, resveratrol and ISO have been reported to activate Sirt1, a key modulator of ROS homeostasis.18-21 The specific antioxidant efficacy of ISO may be mediated by the activation of the Nrf2-antioxidant response element (ARE) pathway in THP-1 and the subsequent inhibition of hydroxyl and hydroperoxyl radical production.13,22 Regarding safety, although the effects of G. parvifolium extract on normal cells have not yet been reported, resveratrol, which is the main stilbene of G. parvifolium, had shown no cytotoxic effects on PBMCs. 23 Similarly, a recent study strengthens our results and demonstrated that ISO did not affect the proliferation of RAW264.7 cells stimulated by lipopolysaccharides (LPS) at a concentration of 40 μM. 24 Therefore, G. parvifolium extract and ISO may exert a cytoprotective effect on normal cells.
Anti-Inflammatory Potential and Cytokine Regulation
The anti-inflammatory efficacy of the G. parvifolium extract is likely mediated by its stilbene content. Resveratrol is well documented to regulate the production of IL-1α and IL-6, decrease levels of IFN-γ, IL-2, and IL-12, and diminish IL-17a mRNA expression.25-27 Moreover, ISO demonstrated superior anti-inflammatory effects compared to resveratrol. 13 Indeed, ISO has been reported to effectively suppress chemokine ligand 8 (CXCL8) expression, reduce activation of the PI3K/Akt pathway, and promote Nrf2 signaling, resulting in inhibition of pro-inflammatory mediators.13,22 Nrf2, in turn, can inhibit NADPH oxidase expression, which is associated with the IL-17a mediated oxidative axis.28,29 Furthermore, our data support previous study reporting the suppression of the production of IL-10 and IL-13 by resveratrol during Th2 response. 30 In conclusion, both G. parvifolium and ISO possess anti-inflammatory benefits able to prevent chronic inflammatory diseases.
Selective Modulation of Adipogenesis
A distinctive finding in this study was the selective impact of the treatments on adipogenesis. Indeed, our results showed that G. parvifolium extract and ISO selectively targeted aP2 expression without significant changes in the expression of PPARγ, adiponectin, HSL, and Pref-1, indicating that these treatments do not strongly suppress the overall differentiation process of adipocytes. Moreover, in recent years, various natural compounds, present in G. parvifolium, were investigated for their anti-adipogenic effects. 31 Our results support previous study showed that resveratrol reduced adipogenesis genes such as aP2, FAS and LPL. 32 However, both piceatannol and resveratrol reported inhibition of adipocyte differentiation by suppressing PPARγ activity. 33 In addition, ISO has been reported to inhibe oxidized LDLs (oxLDL) 34 and oxLDL demonstrated to induce ALBP/aP2 gene expression. 35 Taken together, the unchanged expression of PPARγ, adiponectin, HSL, and Pref-1 suggests that G. parvifolium extract and ISO exert limited effects on adipogenesis through aP2 inhibition, without strongly suppressing adipocyte differentiation.
Limitations of the Study
While this study highlights the promising therapeutic potential of G. parvifolium and ISO in modulating inflammation and oxidative stress, some limitations remain. We used a single non-cytotoxic concentration (50 µg/mL for the extract; 5 µg/mL for ISO) based on preliminary tests, which limits the determination of precise pharmacological potency. Additionally, our 3T3-L1 model focused on transcriptional modulation; since only aP2 was reduced without affecting PPARγ, functional assays like Oil Red O staining were not performed. Finally, the 24-hour window may have missed peak mRNA modulation, and further in vivo studies are needed to confirm the effectiveness and bioavailability of G. parvifolium and ISO in more complex biological systems.
Conclusion
In summary, this study highlights the anti-inflammatory and antioxidant properties of G. parvifolium extract and its bioactive compound, ISO. These effects result in a significant inhibition of the production of ROS and pro-inflammatory cytokines, without impact on cell viability. Furthermore, both compounds reduce aP2 expression, suggesting a potential role in the modulation of adipocyte differentiation. These results support the potential of G. parvifolium and ISO as promising candidates for the development of preventive or therapeutic strategies targeting chronic inflammatory diseases.
Footnotes
Acknowledgments
We thank Rebecca Boutin for her technical assistance. We thank Guillaume Tatti and Andreas Farina (Department of Research and Doctoral Studies DRED, UCA) for their Administrative Project Management. We thank Noémie Silva for the preparation of G. parvifolium extracts.We thank the Service de Biochimie et Génétique Moléculaire du CHU Clermont-Ferrand.
Ethical Considerations
This study was approved by the Institutional Review Board/Ethics Committee of Clermont-Auvergne University, Clermont-Ferrand, France.
Consent to Participate
All donors gave their written informed consent for the use of blood samples for research purposes under Établissement Français du Sang contract no. EFS AURA 25-152 (in accordance with articles L1222-1, L1222-8, L1243-4 and R1243-61 of the French Public Health Code).
Author Contributions
Conceptualization, FCC, JYB, KB, PC; methodology: LH, LL, CP, CD, EF; investigation: LH, LL, CD, DF, PC; validation: FCC, KB, CD, LD, PC; supervision: FCC, LD; writing, reviewing, and editing: LH, CD, LD, FCC. All authors have read and agreed to the published version of the manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by Project ANR-19-LCV2-0003-01 Program LABCOM 2019PHYTOPROB’INOV (AV0027085). The project is co-financed by the European Regional Development Fund (FEDER) and AUF IntenSciF Program (FIVAPRONATS).
Declaration of Conflicting Interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Authors “Christelle Parchemin and Jean-Yves Berthon” were employed by the company Greentech. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Data Availability Statement
No data was used for the research described in the article.
Statement of Human and Animal Rights
All of the experimental procedures involving human-derived materials (blood) were conducted in accordance with the Declaration of Helsinki and approved by the Institutional Review Board/ Ethics Committee of Clermont-Auvergne University, Clermont-Ferrand, France.