
Abstract
Purpose
Chronic inflammation is a key contributor to the development of various metabolic and cardiovascular diseases, underscoring the importance of identifying effective anti-inflammatory agents. Natural products are gaining increasing attention as potential sources of bioactive compounds with therapeutic value. Gloiopeltis tenax (G. tenax), an edible red alga widely consumed in East Asia, exhibits antioxidant and antimicrobial activities; however, its anti-inflammatory effects remain poorly understood. In this study, we investigated the anti-inflammatory potential of an ethanolic extract of G. tenax in lipopolysaccharide (LPS)-stimulated RAW264.7 macrophages.
Methods
The effects of G. tenax extract on the production of inflammatory mediators and major signaling pathways were investigated in LPS-stimulated RAW264.7 macrophages.
Results
Exposure to G. tenax extract markedly attenuated LPS-stimulated nitric oxide production and the release of pro-inflammatory cytokines, such as tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6), accompanied by downregulation of inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) expression, with no detectable cytotoxicity. Moreover, the extract attenuated nuclear factor-κB (NF-κB) pathway activation by suppressing inhibitor of κB (IκB) phosphorylation and attenuation of NF-κB pathway activation. In addition, phosphorylation of mitogen-activated protein kinase (MAPK) signaling components was markedly attenuated in the cells treated with extract.
Conclusion
Collectively, these results demonstrate that G. tenax extract exhibits anti-inflammatory activity in macrophages by suppressing inflammatory mediator production, including iNOS and COX-2 expression, and by regulating the NF-κB and MAPK signaling pathways, highlighting its promise as a functional food ingredient or a natural anti-inflammatory agent.
Keywords
Introduction
Modern lifestyle factors, including high-fat and high-sugar dietary patterns, insufficient physical activity, and persistent stress, have contributed to the increasing incidence of inflammatory disorders, such as obesity, diabetes, and cardiovascular disease.1-4 Although inflammation is a fundamental component of the immune response, its dysregulation may lead to chronic inflammation, which in turn contributes to tissue damage and pathogenesis of various diseases.5-7 Accordingly, the proper regulation of inflammation is considered a critical factor in the prevention and management of numerous chronic conditions.
Natural product research involves the isolation of bioactive compounds from natural sources, such as plants, microorganisms, and animals, with significant potential for diverse applications, including drug discovery and the development of functional foods.8-12 In recent years, increasing attention has been directed toward evaluating natural anti-inflammatory compounds using the RAW264.7 macrophage model.13,14 Gloiopeltis tenax, a red alga found along the coast of East Asia, is commonly consumed as food.15,16 Although recent studies have explored the antioxidant and antimicrobial properties of G. tenax extracts, 17 their anti-inflammatory effects have not been thoroughly investigated. Therefore, the present study aimed to evaluate the anti-inflammatory potential of G. tenax extract in RAW264.7 macrophages.
Lipopolysaccharide (LPS), a component derived from the cell walls of gram-negative bacteria, is a potent inducer of immune responses and a well-known pro-inflammatory agent.18,19 LPS interacts with toll-like receptor 4 (TLR4) on immune cells, including macrophages, leading to the initiation of inflammatory signaling cascades.20-22 Based on this, we hypothesized that G. tenax extract modulates LPS-induced inflammatory responses. Here, we examined whether the ethanol extract of G. tenax regulates the production of inflammatory mediators in LPS-stimulated RAW264.7 macrophages. Upon LPS exposure, NF-κB signaling is initiated via receptor-dependent activation of the IKK (IκB kinase) complex, which in turn phosphorylates IκB proteins.23-25 IκB phosphorylation triggers its degradation, thereby enabling NF-κB to translocate to the nucleus and induce the expression of pro-inflammatory genes, including TNF-α, IL-6, and COX-2.26-28 LPS stimulation also activates the MAPK signaling pathway, which is critically involved in regulating pro-inflammatory cytokine production and immune cell activation.29-32 Consequently, targeting these signaling pathways to control macrophage activity may offer a promising therapeutic approach for inflammatory disorders. Accordingly, this study explored the anti-inflammatory effects of G. tenax extract in LPS-stimulated RAW264.7 macrophages, with a focus on the underlying signaling pathways.
Materials and Methods
G. tenax Preparation
G. tenax was obtained from Wando-gun, South Korea. Then, 40 g of dried G. tenax were extracted with 360 mL of 70% EtOH in a sonicator (Mujigae, Seoul, Republic of Korea) at a temperature of 60 ± 5 °C for 3 h. Subsequently, the extract was filtered through a Whatman filter paper and a PVDF syringe filter (0.22 μm; Millipore, Burlington, MA, USA). After filtration, the solvent was evaporated completely in a drying oven. G. tenax, from which all the residual solvent had evaporated, was dissolved in DMSO (Duchefa, Haarlem, Netherlands). The extract stock solution was prepared in DMSO and stored in aliquots at −20 °C until use. For all cell-based assays, the stock solution was diluted in culture medium to the indicated concentrations, and the final DMSO concentration was kept constant across all groups (final DMSO, 0.1% v/v).
Cell Culture
RAW264.7 macrophages (purchased from the Korea Cell Line Bank (KCLB; Seoul, Republic of Korea; KCLB No. 40071)) were cultured in DMEM (Dulbecco’s Modified Eagle Medium) (Welgene, Gyeongsan-si, Republic of Korea) with 10% FBS (fetal bovine serum) (Corning Costar, Cambridge, MA, USA) and 1% antibiotic-antimycotic (Gibco, Waltham, MA, USA) in a 37 °C incubator in an atmosphere of 5% CO2. Mycoplasma contamination was tested using the e-Myco plus Mycoplasma PCR Detection Kit (iN-tRON Biotechnology, Seoul, Republic of Korea) and authenticated through short tandem repeat (STR) analysis using the AmpFlSTR Identifiler kit (Applied Biosystems, Waltham, MA, USA) at the Korean Collection for Type Cultures (KCTC, Seoul, Republic of Korea).
Cytotoxicity
Cytotoxicity was assessed using the MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay (Duchefa, Haarlem, Netherlands). RAW264.7 cells were cultured in 96-well plates (2 × 104 cells/well). RAW264.7 cells were pre-treated with G. tenax extract (0.25–128 mg/mL) for 2 h and then treated with or without LPS (200 ng/mL) (Sigma Aldrich, St. Louis, MO, USA) for 24 h. LPS was suspended in Dulbecco’s Buffered Saline (DPBS) (Welgene, Daegu, Republic of Korea). Then, 10 μL of MTT solution was added to every single well in the plate, which was incubated for 2 h at 37 °C. After incubation, the absorbance was measured using a UV spectrophotometer (570 nm) (Tecan, Männedorf, Switzerland).
Nitric Oxide (NO) Production Assays
Cells were treated with G. tenax extract and LPS in the same manner as described for cytotoxicity. In addition, dexamethasone (DEX) (Sigma-Aldrich, St. Louis, MO, USA) was used as a positive control for inhibition of NO production. Dexamethasone was used at a final concentration of 1 μM, based on previously reported effective concentrations for NO inhibition in RAW264.7 macrophages. Nitrite levels in culture supernatants were quantified using the Griess assay (Sigma-Aldrich, St. Louis, MO, USA) according to the manufacturer’s instructions. Briefly, culture supernatants were mixed with Griess reagent at a 1:1 ratio, incubated for 10 min at room temperature, and absorbance was measured at 540 nm using a microplate reader (Tecan, Männedorf, Switzerland). Nitrite concentrations were calculated using a sodium nitrite standard curve.
Quantitative PCR (qPCR) Analysis
The cells were pre-treated with the G. tenax extract for 2 h and incubated with LPS for 24 h. RNAiso Plus (TaKaRa, Kusatsu, Japan) was used for RNA extraction, and DNA Master Mix (ToYoBo, Osaka, Japan) was used for cDNA synthesis. Quantitative PCR was performed with TOPreal SYBR Green qPCR PreMIX (Enzynomics, Daejeon, Republic of Korea). The mRNA expression levels of the target genes were analyzed using the cycle threshold (Ct) value of GAPDH. The fold-change values of the LPS and G. tenax extract groups compared with those of the control group were used to quantify each inflammatory factor. The sequences of primers used in qPCR were as follows: iNOS, 5′-ATGACTCCCAGCACAAAGGG-3′ (forward) and 5′-ACGTAGACCTTGGGTTT GCC-3′ (reverse); COX-2, 5′-TGGGTGTGAAGGGAAATAAGG-3′ (forward) and 5′-CATCATATTTGAGCCTTGGGG-3′ (reverse); IL-6, 5′-CCTTCCTACCCCAATTTCCA-3′ (forward) and 5′-CGCACTAGGTTTGCCGAGTA-3′ (reverse); TNF-α, 5′-GGCCTCTCTACCTTGTTGCC-3′ (forward) and 5′-TAGGCGATTACAGTCACGGC-3′ (reverse); and GAPDH, 5′-TGCACCACCAACTGCTTAG-3′ (forward) and 5′-GGATGCAGGGATGATGTTC-3′ (reverse).
Western Blotting
RAW264.7 cells were lysed in RIPA buffer (Biosesang, Yongin-si, Republic of Korea) containing protease and phosphatase inhibitor cocktails (GenDEPOT, Katy, TX, USA). Protein samples were separated by SDS-PAGE and transferred to PVDF membranes (Millipore, Burlington, MA, USA). Specifically, separate SDS-PAGE gels (8%, 10%, and 12%) were prepared to enable efficient resolution of proteins with different molecular weights, ranging from ∼37 kDa (GAPDH, IκBα, p38) to ∼130 kDa (iNOS). Membranes were blocked with 5% skim milk in PBST (PBS containing 0.05% Tween 20) for 90 min at room temperature, incubated with primary antibodies at 4 °C for 16 h, and then incubated with appropriate secondary antibodies for 90 min at room temperature. Membranes were washed with PBST before and after antibody incubation. Signals were detected using an ECL substrate (Bio-Rad, Hercules, CA, USA). Antibodies included anti-IκB, anti-NF-κB, anti-NOS2 (iNOS), anti-COX-2, anti-JNK, anti-ERK1/2, anti-p38, anti-phospho IκB, anti-phospho NF-κB, anti-phospho JNK, anti-phospho ERK, anti-phospho p38, anti-Lamin A/C (Santa Cruz Biotechnology, Dallas, TX, USA), and anti-GAPDH (Bioworld Technology, Nanjing, China). Antibody dilutions were 1:5000 for GAPDH and 1:1000 for other antibodies.
Nuclear Fraction
Nuclear extracted samples were prepared from RAW 264.7 cells based on previously described methods. 33 The protein concentration was measured by the BSA Protein Assay (Thermo Fisher Scientific, USA).
ELISA (Enzyme-Linked Immunosorbent Assay)
TNF-α and IL-6 protein expression levels in culture supernatants were measured using ELISA kits (Abbkine, Wuhan, China) according to the manufacturer’s protocol. RAW264.7 cells were pretreated with G. tenax extract for 2 h and stimulated with LPS for 24 h. After incubation, culture supernatants were collected and analyzed according to the manufacturer’s protocol.
Statistical Analysis
Data were analyzed using GraphPad Prism (version 8.0.2). Statistical significance was determined by one-way ANOVA followed by Tukey’s multiple comparison test, as appropriate, or by Student’s t-test where applicable. Values are presented as mean ± SEM. A p value < 0.05 was considered statistically significant (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001; ####p < 0.0001 as defined in figure legends). In all figures, n refers to the number of independent biological replicates, and the number of independent experiments is indicated in the corresponding figure legends.
Results
G. tenax Extract Does Not Induce Cytotoxicity and Inhibits LPS-Induced NO Production
The cytotoxic effects of G. tenax extract on RAW264.7 macrophages were examined by treating cells with varying concentrations (0.25–4 mg/mL) for 24 h and assessing viability using the MTT assay. According to the results, G. tenax extract did not affect the viability of RAW264.7 cells (Figure 1A). To further evaluate cytotoxicity at higher concentrations, additional experiments were performed. As shown in Figure S1, cell viability significantly decreased at concentrations ≥ 8 mg/mL, whereas concentrations up to 4 mg/mL did not exhibit cytotoxic effects. NO production was determined in RAW264.7 macrophages after 24 h stimulation with LPS (200 ng/mL). The results showed that G. tenax extract significantly reduced NO levels at concentrations of 1–4 mg/mL in RAW264.7 cells. Notably, dexamethasone, used as a positive control, exhibited a comparable inhibitory effect on NO production to that observed with 4 mg/mL of the extract (Figure 1B). Based on these results, G. tenax extract concentrations of 2 and 4 mg/mL were chosen for subsequent experiments. Taken together, these findings suggest that the G. tenax extract does not induce cytotoxic effects and effectively inhibits NO production in RAW264.7 macrophages. Effects of Gloiopeltis tenax extract on cell viability and nitric oxide (NO) production in LPS-stimulated RAW264.7 macrophages
G. tenax Extract Downregulates iNOS and COX-2 Expression at Both the mRNA and Protein Levels
The mRNA expression of the inflammatory enzymes iNOS and COX-2 was upregulated by LPS stimulation. In contrast, treatment with G. tenax extract significantly reduced the mRNA expression levels of iNOS and COX-2 in LPS-stimulated RAW264.7 cells (Figure 2A). Furthermore, western blot results confirmed that increasing concentrations of G. tenax extract effectively inhibited LPS-induced COX-2 and iNOS protein expression in RAW264.7 macrophages (Figure 2B). In summary, G. tenax extract mediates its anti-inflammatory effects through the coordinated inhibition of iNOS and COX-2 expression at both the mRNA and protein levels. Effects of Gloiopeltis tenax extract on LPS-induced iNOS and COX-2 expression in RAW264.7 cells. Cells were pre-treated with G. tenax extract for 2 h and stimulated with LPS for 24 h
G. tenax Extract Suppresses Transcription and Secretion of Pro-inflammatory Cytokines
The mRNA expression levels of the inflammatory cytokines IL-6 and TNF-α were notably increased by LPS stimulation and markedly reduced by G. tenax extract in LPS-induced RAW264.7 cells (Figure 3A). The secretion of the pro-inflammatory cytokines IL-6 and TNF-α was assessed using ELISA to determine the suppressive effects of G. tenax extract. Pro-inflammatory cytokine levels, including IL-6 and TNF-α, were significantly elevated in RAW264.7 macrophages. However, treatment with 2 and 4 mg/mL of G. tenax extract significantly reduced the secretion levels of IL-6 and TNF-α (Figure 3B). In summary, G. tenax extract alleviated inflammatory responses by downregulating the production and secretion of pro-inflammatory cytokines. Effects of Gloiopeltis tenax extract on LPS-induced IL-6 and TNF-α expression in RAW264.7 macrophages. Cells were pre-treated with G. tenax extract for 2 h and stimulated with LPS for 24 h
G. tenax Extract Attenuates NF-κB and MAPK Signaling in LPS-Stimulated RAW264.7 Macrophages
To examine the effect of G. tenax extract on NF-κB inhibition, its effects on IκB phosphorylation were investigated. G. tenax extract suppressed the phosphorylation of IκB in a concentration-dependent manner in LPS-stimulated RAW264.7 cells. Furthermore, it was confirmed that G. tenax extract inhibited the phosphorylation of NF-κB p65 (Figure 4A). These results demonstrate that G. tenax extract mitigates inflammation by blocking NF-κB pathway activation in RAW264.7 cells stimulated with LPS. Effects of Gloiopeltis tenax extract on LPS-induced NF-κB and MAPK signaling pathways in RAW264.7 macrophages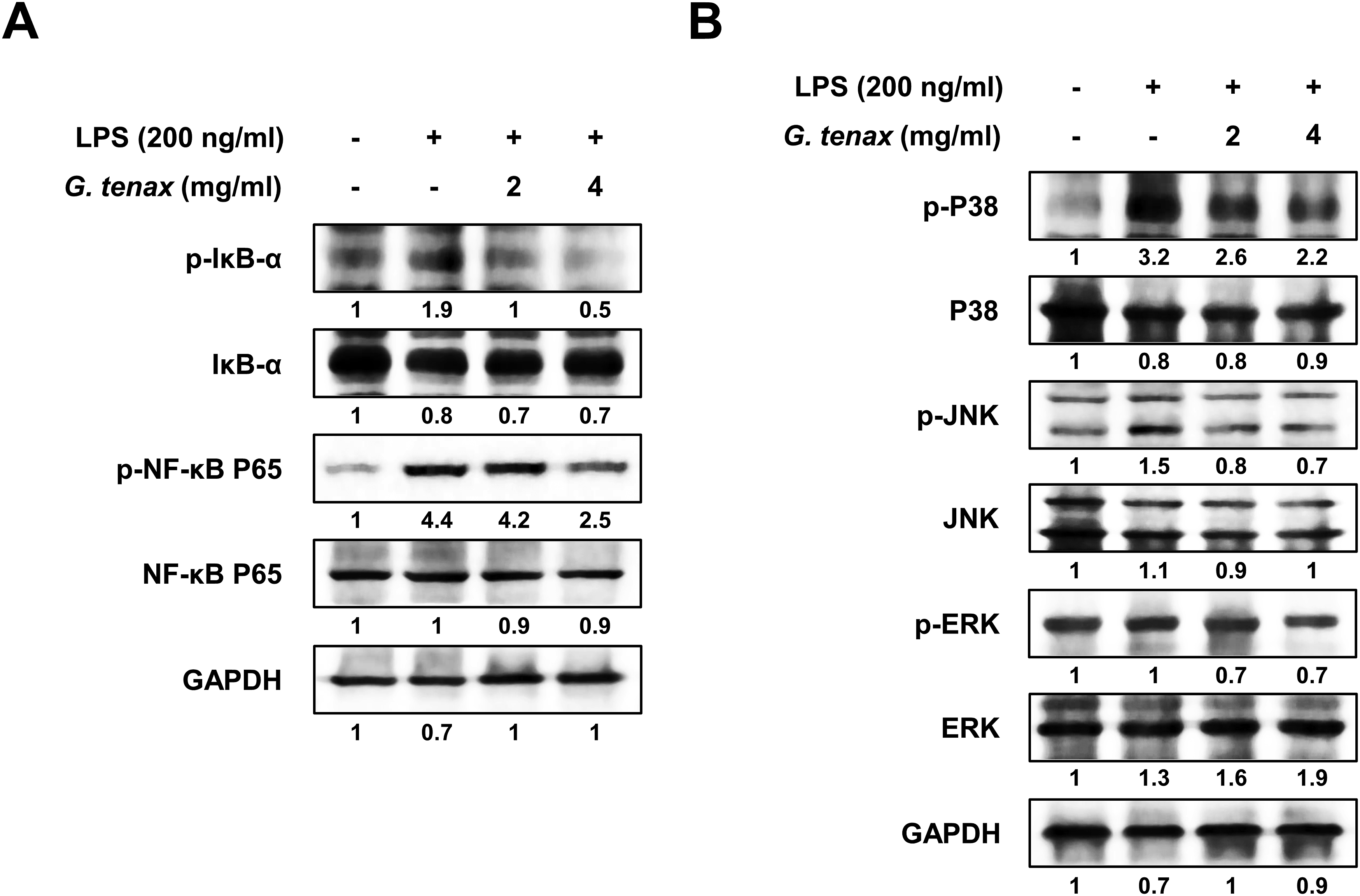
Consistent with these findings, we next examined whether the extract affects NF-κB nuclear translocation using nuclear and whole-cell lysate (WCL) fractionation followed by Western blot analysis. The protein levels of NF-κB in nuclear fractions were analyzed using Lamin A/C as a nuclear marker and GAPDH as a loading control for whole-cell lysates. As shown in Figure S2, treatment with G. tenax extract markedly reduced the nuclear accumulation of NF-κB compared to the LPS-treated control group. These results provide direct evidence that G. tenax extract suppresses NF-κB nuclear translocation.
In addition to NF-κB signaling, we further assessed the effect of G. tenax extract on LPS-triggered MAPK activation. The phosphorylation status of p38, JNK, and ERK was evaluated by Western blot analysis. LPS stimulation significantly increased the phosphorylation of p38, JNK, and ERK in RAW264.7 cells. In contrast, treatment with G. tenax extract reduced the LPS-induced phosphorylation of p38, JNK, and ERK in a concentration-dependent manner (Figure 4B). These data suggest that G. tenax extract exerts anti-inflammatory effects in LPS-stimulated RAW264.7 macrophages through inhibition of p38, JNK, and ERK phosphorylation in the MAPK signaling pathway.
Discussion
Macrophages are key regulators of immune and inflammatory processes, largely because they can secrete pro-inflammatory cytokines and other inflammatory mediators.34-36 Among them, RAW264.7, which is widely used as a standard in vitro model for evaluating anti-inflammatory effects, exhibits robust inflammatory responses upon stimulation with lipopolysaccharide (LPS).34-36 Natural product–based anti-inflammatory agents have attracted attention because of their low toxicity and suitability for long-term use. 37 In particular, bioactive compounds isolated from marine algae, particularly red algae, exhibit strong anti-inflammatory and antioxidant properties.38-41 Previous studies have reported that extracts from various red algae species suppress inflammatory mediators such as NO, iNOS, and pro-inflammatory cytokines through modulation of NF-κB and MAPK signaling pathways. However, studies investigating the anti-inflammatory activity of G. tenax, a red alga, remain limited. Here, we demonstrate that G. tenax extract markedly attenuates LPS-induced inflammatory responses in RAW264.7, highlighting the potential of red algae-derived natural products. These findings are consistent with previous reports on red algae but further expand the current understanding by identifying G. tenax as a relatively underexplored species with anti-inflammatory potential. Collectively, these results provide fundamental evidence supporting the development of G. tenax extract as a functional food ingredient or natural anti-inflammatory candidate.
Several red algae species have been investigated for their anti-inflammatory properties in LPS-stimulated macrophage models, providing a useful framework for contextualizing the present findings. Extracts from Porphyra species have been shown to suppress NO production and iNOS expression through NF-κB pathway inhibition, with effective concentrations typically ranging from 50–200 μg/mL for purified polysaccharide fractions. Gracilaria verrucosa extracts have demonstrated COX-2 and TNF-α suppression at comparable concentrations, primarily through MAPK pathway modulation. Gelidium amansii-derived compounds, including phlorotannins, have been reported to attenuate both NF-κB and MAPK signaling at concentrations below 100 μg/mL. In comparison, G. tenax extract in the present study achieved dual suppression of both NF-κB and MAPK pathways using a crude ethanolic extract, albeit at higher concentrations (2–4 mg/mL), reflecting the unfractionated nature of the preparation. Notably, the simultaneous dose-dependent inhibition of both signaling cascades by G. tenax extract distinguishes it from several previous studies that reported preferential modulation of either NF-κB or MAPK pathways individually. These observations suggest that the crude extract may contain multiple bioactive constituents with complementary mechanisms of action, and underscore the potential value of bioactivity-guided fractionation to identify the specific compounds responsible for these effects.
Here, we investigated whether G. tenax extract modulates inflammatory responses in LPS-stimulated macrophages. Treatment with G. tenax extract significantly and dose-dependently suppressed LPS-induced inflammatory responses without inducing cytotoxicity. The extract markedly suppressed NO production in response to LPS stimulation. NO production upon LPS stimulation primarily occurs in immune cells, especially macrophages, and is mediated by nitric oxide synthase (NOS). Although nNOS and eNOS are constitutively expressed in normal cells, iNOS is strongly induced by inflammatory stimuli, resulting in increased NO production. 42 Excessive NO production is associated with amplified inflammatory signaling and cytokine responses, including TNF-α and IL-6. Cyclooxygenase-2 (COX-2) is an inducible inflammatory enzyme involved in prostaglandin synthesis and is commonly upregulated in activated macrophages.43,44 In our study, LPS stimulation markedly increased COX-2 and iNOS expression at the transcriptional level, which was significantly suppressed by G. tenax extract; western blot analysis further supported these findings.
Inflammatory responses and cytokine expression are highly interrelated, with cytokines playing a pivotal role in regulating and amplifying immune responses during inflammation. Immune and inflammatory responses are orchestrated by cytokines produced by various immune cells, including macrophages, lymphocytes, and neutrophils.45,46 These cytokines are recognized as potent inducers of major inflammatory mediators, including iNOS and COX-2.47,48 The qRT-PCR results demonstrated that increasing concentrations of G. tenax extract progressively suppressed LPS-stimulated transcription of pro-inflammatory cytokines. Additionally, ELISA results showed that cytokine secretion was significantly reduced in the group treated with G. tenax extract compared to that in the LPS-only treated group.
LPS is known to induce NF-κB activation in macrophages, a central mediator of inflammatory responses, while NF-κB normally exists in the cytoplasm as an inactive protein complex. Upon inflammatory stimulation, such as LPS, binding of LPS to its receptor on the cell surface activates the IκB kinase (IKK) complex, which subsequently phosphorylates the IκB protein. IκB phosphorylation triggers its proteasomal degradation, thereby enabling NF-κB to enter the nucleus and activate gene transcription.49,50 Consistent with this mechanism, our results showed that G. tenax extract suppressed the phosphorylation of IκBα and NF-κB p65, suggesting inhibition of NF-κB activation. This phosphorylation-based approach for inferring NF-κB pathway suppression is well-established in the literature; for example, Li et al. (2020) demonstrated simonsinol-mediated NF-κB inhibition in LPS-stimulated RAW264.7 cells using comparable p-IκBα and p-NF-κB p65 western blot data, and Zhong et al. (2022) employed a similar strategy to evaluate flavonol-mediated anti-inflammatory effects. Furthermore, the simultaneous dose-dependent reduction of four independent NF-κB target gene products (iNOS, COX-2, TNF-α, and IL-6) at both transcriptional and translational levels provides robust functional evidence supporting impaired NF-κB transcriptional activity.
In addition to NF-κB suppression, pretreatment with G. tenax extract significantly attenuated the LPS-induced phosphorylation of p38, JNK, and ERK, which are critical components of the MAPK signaling cascade. This simultaneous suppression of both NF-κB and MAPK pathways is consistent with previously reported mechanisms of red algae-derived anti-inflammatory agents, and distinguishes G. tenax extract from species that preferentially modulate either pathway individually.
Collectively, our data show that G. tenax extract reduces inflammatory mediator production and pro-inflammatory cytokine responses while not exhibiting detectable cytotoxicity in RAW264.7 macrophages. This in vitro study provides a foundation for further investigation of G. tenax-derived bioactives. Future work should focus on identifying the active components responsible for these effects, establishing extract standardization parameters, and validating efficacy in in vivo models of inflammation.
Despite these promising findings, several limitations of the present study should be considered when interpreting the results. First, we used a crude ethanolic extract without comprehensive phytochemical characterization or identification of the active constituents. Although previous studies have reported the presence of fatty acids, sterols, and polyphenolic compounds in G. tenax, the specific components responsible for the observed anti-inflammatory effects remain unclear. Therefore, further fractionation and compound-level analyses will be necessary to identify the bioactive molecules responsible for these effects. In this regard, quantification of total phenolic content and total polysaccharide content, which are relatively straightforward yet informative assays for extract characterization, will be prioritized in future studies. Second, although this study provides mechanistic insights into NF-κB and MAPK signaling pathways, additional investigation of upstream signaling components would further strengthen the understanding of the molecular mechanisms involved. Finally, the effective concentrations used in this study are relatively high, which may limit direct physiological relevance. In addition, our findings are based on an in vitro model and may not fully recapitulate the complexity of in vivo inflammatory responses. Therefore, future studies using appropriate animal models will be essential to validate the physiological relevance and therapeutic potential of these findings.
Supplemental Material
Supplemental Material - Gloiopeltis tenax Extract Inhibits LPS-Induced Inflammatory Response in RAW264.7 Macrophage Cells
Supplemental Material for Gloiopeltis tenax Extract Inhibits LPS-Induced Inflammatory Response in RAW264.7 Macrophage Cells by Yu-Seon Lee, Hana Lee, Won-Jin Kim, AReum Choi, Woong Kim, Seok-Jun Kim in Natural Product Communications
Footnotes
Author contributions
All authors have read and agreed to the published version of the manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the Global-Learning & Academic Research Institution for Master’s Ph.D. students and a Postdocs (LAMP) Program of the National Research Foundation of Korea (NRF) grant funded by the Ministry of Education [No. RS-2023-00285353], and the Chosun University (2024).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.