
Abstract
Objective
This study aimed to clarify the inhibitory effects of Spatholobus suberectus Dunn (SSD) on cervical cancer (CC) cells using network pharmacology.
Methods
We explored SSD’s mechanism and targets in CC therapy and identified potential therapeutic targets through comprehensive analysis. Active components of SSD were identified via the TCMSP database, and disease-related targets were sourced from TTD, GeneCards, OMIM, and PharmGKB. A Venn diagram identified intersecting targets, and the STRING database facilitated a protein-protein interaction network. Cytoscape visualized the active C-T-D network. GO and KEGG enrichment analyses were performed using R software. Molecular docking was verified with AutoDock and PyMOL, while the CCK-8 assay assessed SSD’s inhibitory effects on Caski cell proliferation at 24, 48, and 72 hours. The scratch assay evaluated SSD’s impact on cell migration, and RTCA tracked its effects on proliferation and migration over 72 hours. Flow cytometry examined apoptosis and cell cycle distribution post-SSD treatment. Protein expression levels of P27, P21, P53, and those in the PI3K/Akt and Jak2/Stat3 pathways were measured using the JESS automated Western blot system.
Results
Screening identified 24 active components from SSD. Analysis revealed 32,330 CC targets, with 119 overlapping drug-disease targets identified using Jvenn. KEGG pathway enrichment showed SSD significantly affects PI3K/Akt and P53 pathways. Molecular docking indicated strong binding of SSD’s active ingredients to key CC targets. CCK-8, scratch assays, and RTCA demonstrated SSD’s inhibitory effects on Caski cell proliferation and migration. Flow cytometry showed SSD induces apoptosis and cell cycle arrest in Caski cells. Jess assay results suggested SSD may inhibit CC progression by modulating pathways like P27, P21, P53, PI3K/Akt, and Jak2/Stat3.
Conclusion
SSD has a significant inhibitory effect on CC cells, though the mechanisms require further exploration.
Keywords
Introduction
CC is primarily attributed to the persistent infection of high-risk human papillomavirus (HPV) and represents a prevalent malignancy among women. 1 The progression of CC encompasses multiple stages, often asymptomatic in the initial phases and becoming pronounced in the later stages. 2 At present, the cornerstone treatments for early-stage CC include surgical intervention and radiotherapy, with chemotherapy and immunotherapy serving as supplementary approaches. Early detection through screening and HPV vaccination are crucial preventive measures that significantly improve patient survival rates. 3 Recent advancements in CC treatment have been noteworthy, particularly with the introduction of immune checkpoint inhibitors, which offer novel therapeutic options for patients. Anti-PD-1 and anti-PD-L1 antibodies have demonstrated substantial clinical efficacy in CC management, with some patients experiencing prolonged tumor remission. 4 Although T cell immunotherapy exhibits potential in CC treatment, its overall response rate remains limited, benefiting only approximately 10% to 20% of patients. 5 The immunosuppressive nature of the tumor microenvironment has emerged as a significant challenge in CC immunotherapy, frequently involving regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs). These cells suppress the host’s anti-tumor immune response by secreting factors such as IL-10 and TGF-β. 6 Furthermore, CC cells often evade immune detection by upregulating PD-L1 expression and downregulating MHC class I molecules, thereby further diminishing the effectiveness of immunotherapy. 7 The limitations associated with monotherapy have become increasingly apparent, prompting researchers to investigate combination therapies. The integration of immune checkpoint inhibitors with radiotherapy, chemotherapy, or targeted therapies may potentiate the anti-tumor immune response through synergistic effects. 8 Nonetheless, the clinical application of these combined treatments remains contentious, and their therapeutic efficacy requires further validation. In this context, it is crucial to identify novel immunotherapeutic targets and strategies.
Traditional Chinese Medicine (TCM) encompasses a diverse array of anti-cancer active compounds, positioning it as a significant modality in cancer treatment characterized by low toxicity and high efficacy.9-11 TCM’s anti-cancer mechanisms include the induction of apoptosis, inhibition of angiogenesis, overcoming multidrug resistance, enhancement of the immune system, and exerting multi-target and multi-channel effects. 12 Spatholobus suberectus Dunn (SSD), a traditional Chinese herbal medicine from the Leguminosae family, is predominantly found in southern China and Southeast Asia. 13 According to TCM principles, SSD is believed to promote blood circulation, eliminate blood stasis, clear meridians, reduce swelling, and alleviate pain, and is traditionally used to treat conditions such as anemia, irregular menstruation, rheumatism, and arthralgia.14,15 Contemporary pharmacological research has demonstrated that SSD possesses anti-oxidative, 16 anti-inflammatory, 17 anti-tumor, 18 and other beneficial properties. For instance, the mechanism by which genistein, an active constituent of SSD, induces apoptosis in hepatoma cells via mitochondrial mediation and the Fas signaling pathway has been substantiated. 19 SSD is also capable of effectively inducing tumor cell apoptosis by modulating various signaling pathways, including the mitochondrial pathway 20 and the endoplasmic reticulum stress pathway. 21 This indicates that SSD holds significant potential for application in cancer treatment; however, its specific targets and mechanisms in the treatment of CC remain unclear.
With advancements in contemporary research methodologies, network pharmacology offers a novel perspective for elucidating the multi-component and multi-target mechanisms of traditional Chinese medicine. 22 This study aims to investigate the molecular mechanisms and action pathways of SSD in CC through the application of network pharmacology and in vitro experiments.
Method
Database Platform
TCMSP database (https://tcmspw.com/tcmsp. php), GeneCards database (https://www.genecards.org/), PharmGKB database (https://www.pharmgkb.org/), TTD database (https://db.idrblab.net/ttd/), OMIM database (https://omim.org/), DisGeNET database (https://www.disgenet.org/home/), Uniprot database (https://www.uniprot.org/), STRING database (https://string-db.org/), PDB database (https://www.rcsb.org/), Micro-bioinformatics platform (https://www.bioinformatics.com.cn/), Jvenn platform (https://jvenn.toulouse.inra.fr/).
SSD Compound Target and CC Target Acquisition
The primary active constituents and their corresponding protein targets of SSD ( Chinese Pinyin name: Jixueteng, Latin name: Spatholobus Suberectus Dunn) , were identified using the TCMSP database. The search criteria included an oral bioavailability (OB) of ≥ 30% and a drug-likeness (DL) of ≥ 0.18. Subsequently, the Uniprot database was employed to convert these protein targets into gene names.
To identify therapeutic targets associated with CC, databases such as the TTD, GeneCards, OMIM, and PharmGKB were utilized, with “Cervical cancer” serving as the search keyword. Targets were selected based on a score exceeding 1 in these databases.
Network Construction
Using a micro-bioinformatics platform, a network was constructed by identifying the intersection between the targets of SSD’s active components and those related to CC. These intersecting targets were designated as the therapeutic genes for SSD in the treatment of CC. The relationships among SSD’s active components, their target genes, and CC targets were analyzed using Cytoscape 3.8.0 software, resulting in the construction and visualization of an “active component-target-disease' (C-T-D) network.
Construction of Protein-Protein Interaction (PPI) Networks and Screening of Core Targets
Import the intersecting genes into the STRING database, specifying Homo sapiens as the species. Set the confidence level to 0.9 and exclude protein targets lacking correlation. Construct and export the protein interaction network diagram. Subsequently, utilize the STRING tool to download the TSV file containing the PPI analysis results. Import this TSV file into Cytoscape version 3.8.0 for visual analysis. Employ the CytoNCA plugin to perform network characteristic value analysis, focusing on the Degree, Closeness, and Betweenness parameters. Use values exceeding the respective medians of these parameters as criteria to identify network nodes as core targets for SSD intervention in CC.
Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analyses
Utilizing R version 4.4.1 and various RStudio packages, including clusterProfiler, ggplot2, enrichplot, stringi, pathview, colorspace, DOSE, and org.Hs.eg.db, Gene Ontology (GO) enrichment analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis were performed. Only those GO functions and KEGG pathways with a p-value less than 0.05 were considered statistically significant and selected for subsequent investigation.
Molecular Docking
Utilizing network construction, PPI analysis, and KEGG enrichment analysis, the relevant core targets were identified as protein receptors. These receptors were subsequently subjected to molecular docking with small-molecule ligands associated with SSD using AutoDockTools version 1.5.7. The procedure is detailed as follows: Initially, the 2D structures of small molecule ligands are downloaded from PubChem. Subsequently, these 2D structures are converted into 3D structures using Chem3D 14.0 software, where structural energy minimization is performed. The candidate protein’s identifier is then located in UniProt, and its 3D structure is retrieved from the PDB. This structure is imported into PyMOL 2.4.0 software, where water molecules and the original ligand are removed. Following this, the small molecule ligand and protein receptor are imported into AutoDockTools 1.5.7 software for hydrogenation and active pocket determination. Finally, molecular docking between the processed active component and the target protein is conducted using AutoDock Vina, and the results are visualized.
Experimental Validation
Preparation of SSD Extract
The aqueous extract of SSD was provided by Sanyuan Longsheng Biotechnology Co., Ltd., with batch number: LXJXT230215.
Cell Culture
For the experiment, Caski cells were procured from Sewen Innovation Biotechnology Co., Ltd. ( Beijing, China) Brand: Abcells No.: Ac267. These cells were cultured in RPMI 1640 medium, supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin, and maintained in an incubator at 37°C with 5% CO2.
Cell Viability Assay
Following cell quantification, Caski cells were plated at a density of 2×103 cells per well in a 96-well plate. After a 4-hour incubation period, the cells were pre-treated with varying concentrations of SSD (0, 0.2, 0.4, 0.8, and 1.6 mg/mL) for durations of 24, 48, and 72 hours, with three replicate wells for each concentration. Subsequently, 10 μL of CCK-8 solution (KeyGEN BioTECH, Jiangsu, China; Batch number: 20250620) was added to each well, and the plates were incubated at 37°C for 1 hour. Absorbance was then measured at 450 nm using a microplate reader (Biotek, USA).
Cell Scratch Assay
Following cell quantification, Caski cells were plated at a density of 4×105 cells per well in a 6-well plate. After a 4-hour incubation period, a scratch was introduced into the cell monolayer using a sterile pipette tip. Subsequently, the complete medium was aspirated, and the monolayer was rinsed three times with complete medium. The culture medium containing specified concentrations of SSD (0, 0.2, 0.4, 0.8, and 1.6 mg/mL) was then added to each well. Cell migration into the wound area was monitored using a 40x inverted phase contrast microscope ( OLYMPUS, CKX53,Japan) at 0, 24, 48, and 72 hours post-incubation.
Flow Cytometry for Cell Cycle Analysis
Following cell quantification, Caski cells were plated at a density of 3×105 cells per well in a 6-well plate. After a 4-hour incubation period, the cells underwent pre-treatment with varying concentrations of SSD (0, 0.2, 0.4, 0.8, and 1.6 mg/mL) for a duration of 72 hours, with three replicate wells allocated for each concentration. Subsequently, the cells were harvested, and their cell cycle status was assessed utilizing the Cell Cycle Detection Kit (KeyGEN Bio TECH, Jiangsu, China). The cell cycle analysis was conducted using a flow cytometer (Agilent Technologies, NoloCyte), and the resulting data were processed using FlowJo software.
Flow Cytometry for Cell Apoptosis Analysis
Following cell quantification, Caski cells were plated at a density of 1×105 cells per well in a 12-well plate. After a 4-hour incubation period, the cells underwent pre-treated with varying concentrations of SSD (0, 0.2, 0.4, 0.8, and 1.6 mg/mL) for a duration of 48 hours, with three replicate wells allocated for each concentration. Subsequently, the cells were harvested, and apoptosis was assessed utilizing the Annexin V-FITC/PI Apoptosis Detection Kit (KeyGEN BioTECH, Jiangsu, China). Apoptotic cell analysis was conducted using a flow cytometer (Agilent Technologies, NoloCyte), and the resulting data were processed with FlowJo software.
Caski Cell Migration and Proliferation RTCA Assay
The study investigated the impact of varying concentrations of SSD on the migration and proliferation of Caski cells. For the assessment of migration, 165 μL of a drug-containing complete medium was introduced into the lower chamber of the CIM plate, with SSD concentrations set at 0, 0.2, 0.4, 0.8, and 1.6 mg/mL, each concentration tested in triplicate. Subsequently, 30 μL of complete medium was added to the upper chamber, followed by baseline measurement and cell counting. Caski cells were then seeded into the upper chamber at a density of 5×103 cells per well. For the proliferation analysis, 50 μL of cell culture medium was added to each well of a 16-well E-Plate, baseline measurements were taken, and cells were counted. Caski cells were then seeded at a density of 5×103 cells per well, and 10 μL of SSD at varying concentrations (0, 0.2, 0.4, 0.8, and 1.6 mg/mL) was added to each well, with three replicates per concentration. The RTCA system (ACEA, USA) was employed to monitor the effects on cell migration and proliferation every 15 minutes over a 72-hour period.
Jess
Following cell quantification, Caski cells were plated at a density of 4×105 cells per well in a 6-well plate. After a 4-hour incubation period, the cells underwent pre-treated with varying concentrations of SSD (0, 0.2, 0.4, 0.8, and 1.6 mg/mL) for a duration of 72 hours. Subsequently, the cells were lysed using the SevenFast column-based total protein extraction kit (Saiwen Innovation Biotechnology, Lot: 25BN0030) to obtain the protein solution. The protein concentration was then determined utilizing the BCA kit (BOSTER Biological Technology). The protein sample stock solution was combined with 0.1x Sample buffer and Fluorescent 5x Master mix (ProteinSimple) at room temperature, followed by denaturation at 95°C for 5 minutes. The primary antibodies employed in this study included P53 (Cell Signaling Technology, 2527T), P21 (Cell Signaling Technology, 2947S), P27 (Cell Signaling Technology, 3686S), PI3K (Cell Signaling Technology, 4527T), Akt (Cell Signaling Technology, 4691T), Jak2 (Cell Signaling Technology, 3230T), Stat3 (Cell Signaling Technology, 12640T), and GAPDH (Servicebio, Lot: AC250601001). Following the sequential addition of all reagents, the sample underwent centrifugation at 2500 rpm for a duration of 5 minutes at ambient temperature. Subsequently, detection was performed using the Jess system (ProteinSimple, USA). Quantitative analysis of band intensities was conducted utilizing ImageJ software.
Statistical Analysis
All data are expressed as mean ± standard deviation. Statistical analyses were conducted using GraphPad Prism software (version 9.1.0, San Diego, CA, USA). One-way analysis of variance (ANOVA) was used for comparison between groups, and three repeated data were used for each group. A P value of less than 0.05 was deemed indicative of statistical significance.
Result
Screening of Active Ingredients and Prediction of Targets
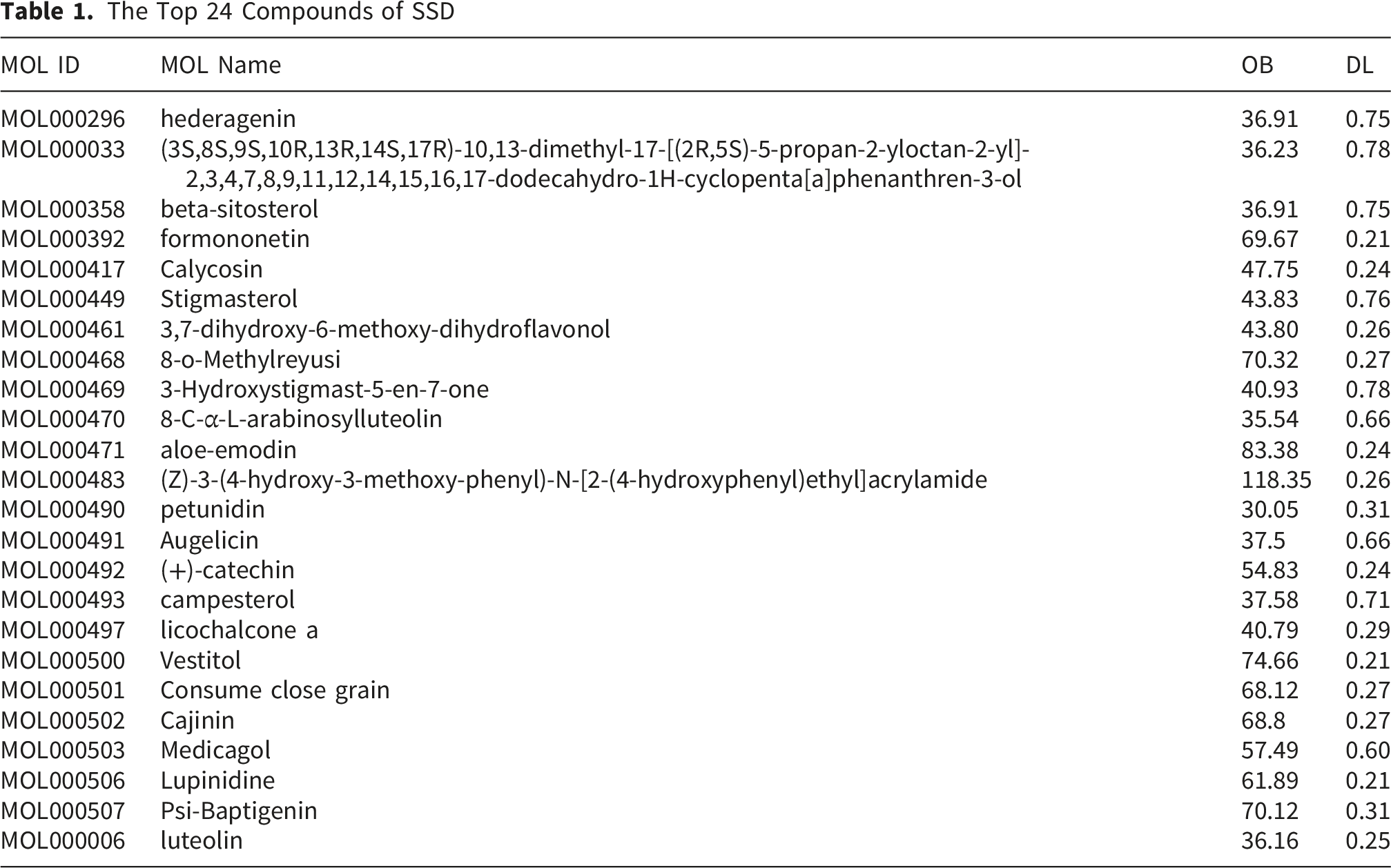
The process and ideas of this experiment are shown in Figure 1. A total of 68 compound molecules were identified in SSD. Following screening criteria of oral bioavailability (OB) ≥ 30% and drug-likeness (DL) ≥ 0.18, 24 compounds were selected for further analysis (see Table 1). These 24 compounds were associated with 427 target genes according to the TCMSP database. Furthermore, a search in the GeneCards database using the keyword “Cervical Cancer' identified 32,330 related genes (Score > 1). Additionally, the OMIM database identified 410 related genes, the PharmGKB database identified 3 related genes, and the TTD database identified 15 related genes, culminating in a total of 32,353 related genes, as illustrated in Figure 2A. Flowchart of network pharmacology analysis of SSD on CC and in vitro experimental validation The Top 24 Compounds of SSD Predicted targets.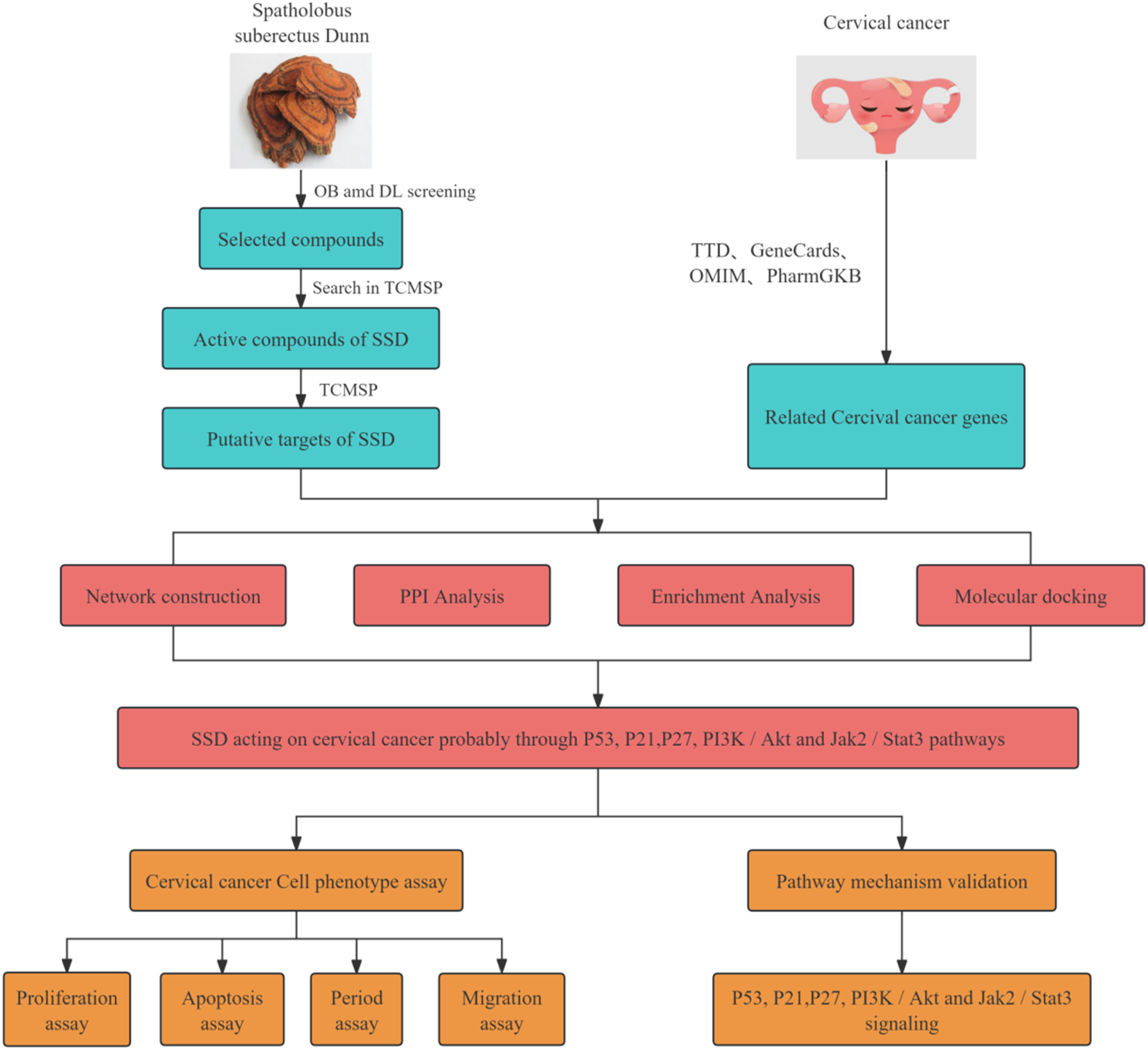
C-T-D Network Analysis
The intersection of the 427 target genes of the compounds with 32,353 CC-related genes resulted in 119 overlapping genes, as illustrated in Figure 2B. The compound-target-disease (C-T-D) network consists of 144 nodes and 385 edges. Within this network, green ellipses denote compounds, red triangles denote genes, yellow hexagons denote SSD herbs, and purplish-red diamonds denote CC, as illustrated in Figure 3. The five compounds with the highest degree centrality are luteolin (MOL000006), formononetin (MOL000392), licochalcone A (MOL000497), beta-sitosterol (MOL000358), and stigmasterol (MOL000449); these compounds are likely to play pivotal roles in exerting anticancer effects against CC. C-T-D Network figure.
PPI Network Analysis
To investigate the interactions between the pertinent targets of SSD and CC, we performed a protein-protein interaction (PPI) analysis, which comprised 89 nodes and 296 edges, as illustrated in Figure 4A. Utilizing Cytoscape 3.8.0 software, we employed the CytoNCA plugin to analyze network characteristic values. We selected the parameters of Degree, Closeness, and Betweenness for this analysis. Nodes and edges exceeding the median values of these parameters underwent an initial screening, resulting in 27 nodes and 123 edges, as illustrated in Figure 4B. A subsequent screening process yielded 8 nodes and 56 edges, identifying these 8 target points as the core targets, as illustrated in Figure 4C. These findings suggest that SSD plays a significant role in the treatment of CC. Illustrates PPI core target selection. Figure A contains 89 nodes and 296 edges, Figure B has 27 nodes and 123 edges, and Figure C includes 8 nodes and 56 edges. Larger shapes indicate higher degree values and greater core significance. Ultimately, 8 core targets were identified
GO and KEGG Enrichment Analysis
The intersection targets of drugs and diseases underwent Gene Ontology (GO) enrichment analysis across three categories: biological process (BP), molecular function (MF), and cellular component (CC). Utilizing a screening threshold of P < 0.05, a total of 2,210 GO terms were identified. Specifically, 1,973 terms were associated with BP, 164 with MF, and 73 with CC. From these, the top 10 terms were selected, revealing that the SSD targets were predominantly localized in membrane rafts, plasma membrane rafts, and membrane microdomains. These targets may influence the proliferation and apoptosis of CC cells by interacting with cell membrane receptors, membrane structure-related proteins, and transcription factors, as illustrated in Figure 5A. Furthermore, KEGG pathway enrichment analysis was conducted on the intersection targets of drugs and diseases, also using a P < 0.05 threshold, resulting in the identification of 181 enriched signaling pathways. The top 20 enriched pathways are depicted in the figure, with the PI3K/Akt signaling pathway being notably enriched, involving 27 targets, as illustrated in Figure 5B. GO and KEGG Enrichment Analysis. Figure A illustrates BP, CC, and MF analyses, with lower q-values and darker colors signifying greater GO term enrichment. Figure B displays the top 20 KEGG enriched pathways, with the x-axis showing the number of target genes per pathway and the y-axis listing the pathways. Red denotes a lower p-value
Molecular Docking Analysis
Utilizing the CTD network, PPI, and KEGG enrichment analyses, we identified the core targets TP53, STAT3, AKT1, BCL2, HSP90AA1, CDKN1A, CASP3, and ESR1 for molecular docking with the top five compounds: luteolin (MOL000006), formononetin (MOL000392), licocalchone a (MOL000497), beta-sitosterol (MOL000358), and stigmasterol (MOL000449). As illustrated in Figure 6, the scoring heatmap of docking binding energies indicates that the most favorable binding combinations are TP53_MOL000449, TP53_MOL000358, TP53_MOL000497, BCL2_MOL000006, and TP53_MOL000006. The docking results are presented in Figure 7A, B, C, D, and E. Stigmasterol exhibits interactions with residues GLU-1598, ARG-1597, ARG-1595, ASN-1594, and GLU-1591 on TP53. Beta-sitosterol engages with GLU-1598, ARG-1597, ARG-1595, and ASN-1594 on TP53. Licocalchone a interacts with GLU-1591, ARG-1597, ARG-1595, and GLU-1598 on TP53. Luteolin demonstrates interactions with TYR-9, ASN-11, ILE-189, and GLN-190 on BCL2, forming hydrogen bonds with TYR-9, ASN-11, HIS-186, and ASP-196, and establishing π-stacking with TRP-195. Additionally, luteolin interacts with LEU-1489, CYS-1535, and ASP-1536 on TP53, forming hydrogen bonds with ARG-1490 and TYR-1600, and π-stacking with TYR-1600. These interactions underpin the stable binding of small molecules to protein receptors. The molecular docking analysis reveals that the compounds exhibit favorable interactions with the protein targets through various bonding mechanisms. The evaluation of binding affinity, as indicated by binding energy, suggests that lower binding energies correspond to enhanced stability, thereby confirming the strong binding affinity of the compounds to the active sites of the protein targets. Binding energy heatmap (Unit: kcal·mol–1). Molecular docking illustration. A, B, C, D, and E correspond to TP53_MOL000449, TP53_MOL000358, TP53_MOL000497, BCL2_MOL000006, and TP53_MOL000006, respectively. Purple lines indicate π-stacking, green lines indicate hydrogen bonds, and blue lines indicate hydrophobic forces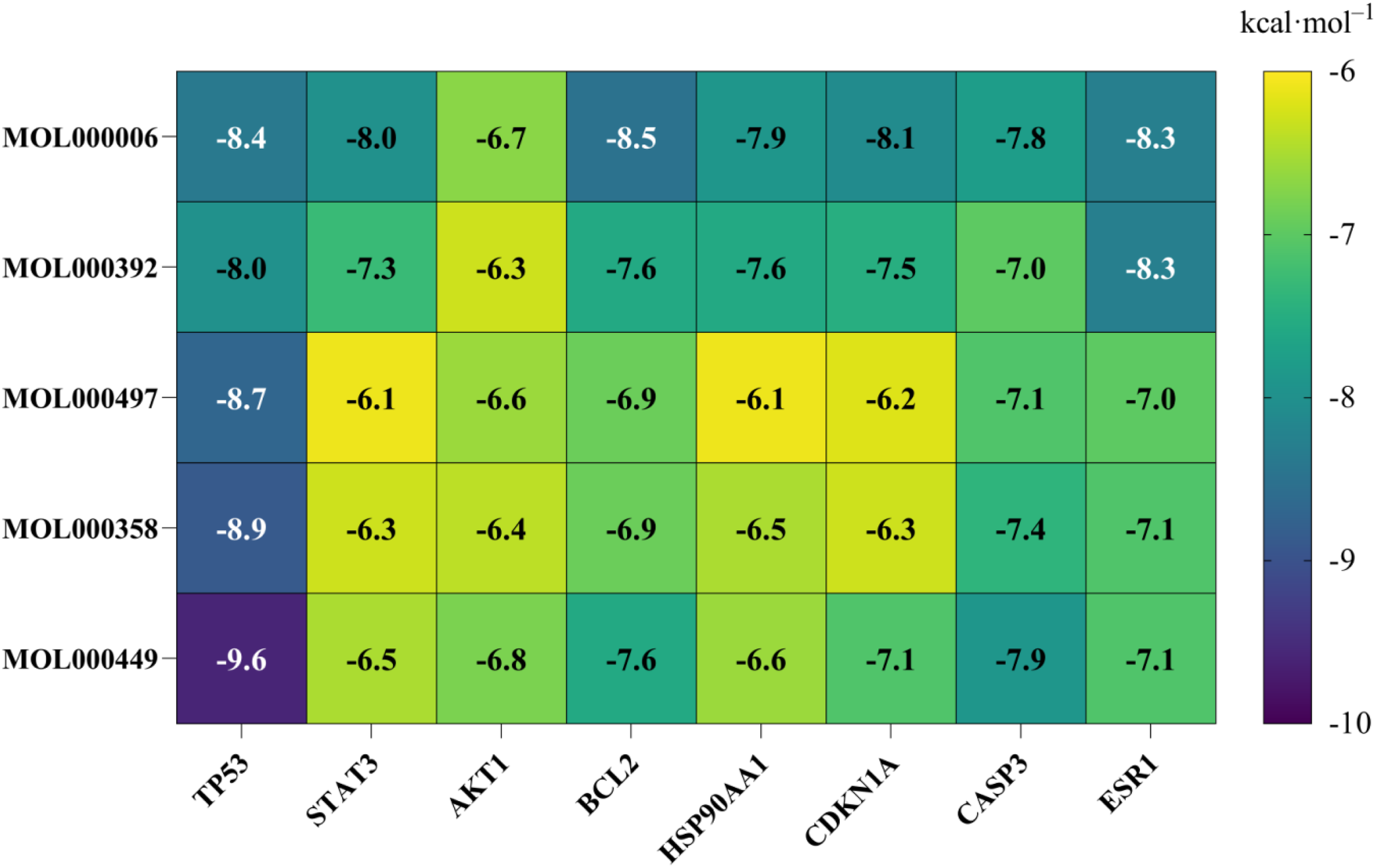

SSD Inhibits the Proliferation of Caski Cells
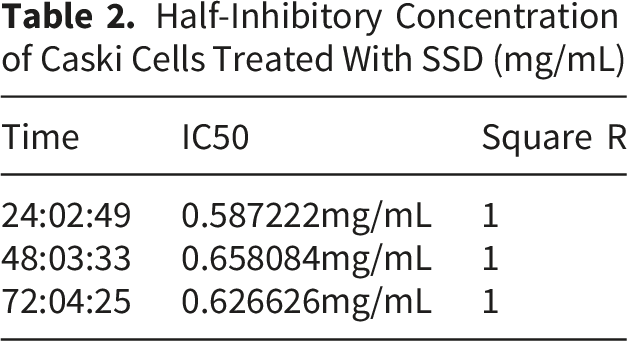
In this study, we aimed to evaluate the inhibitory effect of SSD on cell proliferation by employing CCK8, RTCA, and cell cycle flow cytometry assays. Caski cells were exposed to varying concentrations of SSD for 24, 48, and 72 hours. The results demonstrated that SSD inhibited the proliferation of Caski cells across all tested concentrations and time points, with the 1.6 mg/mL concentration exhibiting the most pronounced effect. Notably, significant inhibition was observed at the 72-hour mark, as illustrated in Figure 8A. In the RTCA assay, 5×103 Caski cells were plated per well and treated with SSD concentrations of 0, 0.2, 0.4, 0.8, and 1.6 mg/mL. At 24, 48, and 72 hours, the cell index for each SSD concentration was consistently lower than that of the control group, displaying a concentration-dependent inhibitory pattern, as illustrated in Figure 8B. The half-maximal inhibitory concentrations (IC50) of SSD on Caski cells at 24, 48, and 72 hours are presented in Table 2. An R-squared value of 1 shows perfect alignment between the experimental data and the fitted dose-response curve, with no deviations. In subsequent experiments, the concentration of SSD will be alternately increased and decreased at the 24-hour IC50 value. Additionally, cell cycle analysis indicated that SSD treatment resulted in increased proportions of Caski cells in the G0/G1 and G2/M phases, suggesting cell cycle arrest and inhibited proliferation activity. At the same time, the growth ratio of G2/M was higher than that of G0/G1 at SSD concentration ( 0.2-0.8mg/mL). This may be associated with DNA damage or the activation of repair mechanisms, as illustrated in Figure 8C. In conclusion, SSD exhibits inhibitory effects on the proliferation of Caski cells through multiple mechanisms. Therefore, SSD holds potential as an effective agent in preventing the occurrence and progression of CC. SSD reduces Caski cell proliferation. Half-Inhibitory Concentration of Caski Cells Treated With SSD (mg/mL)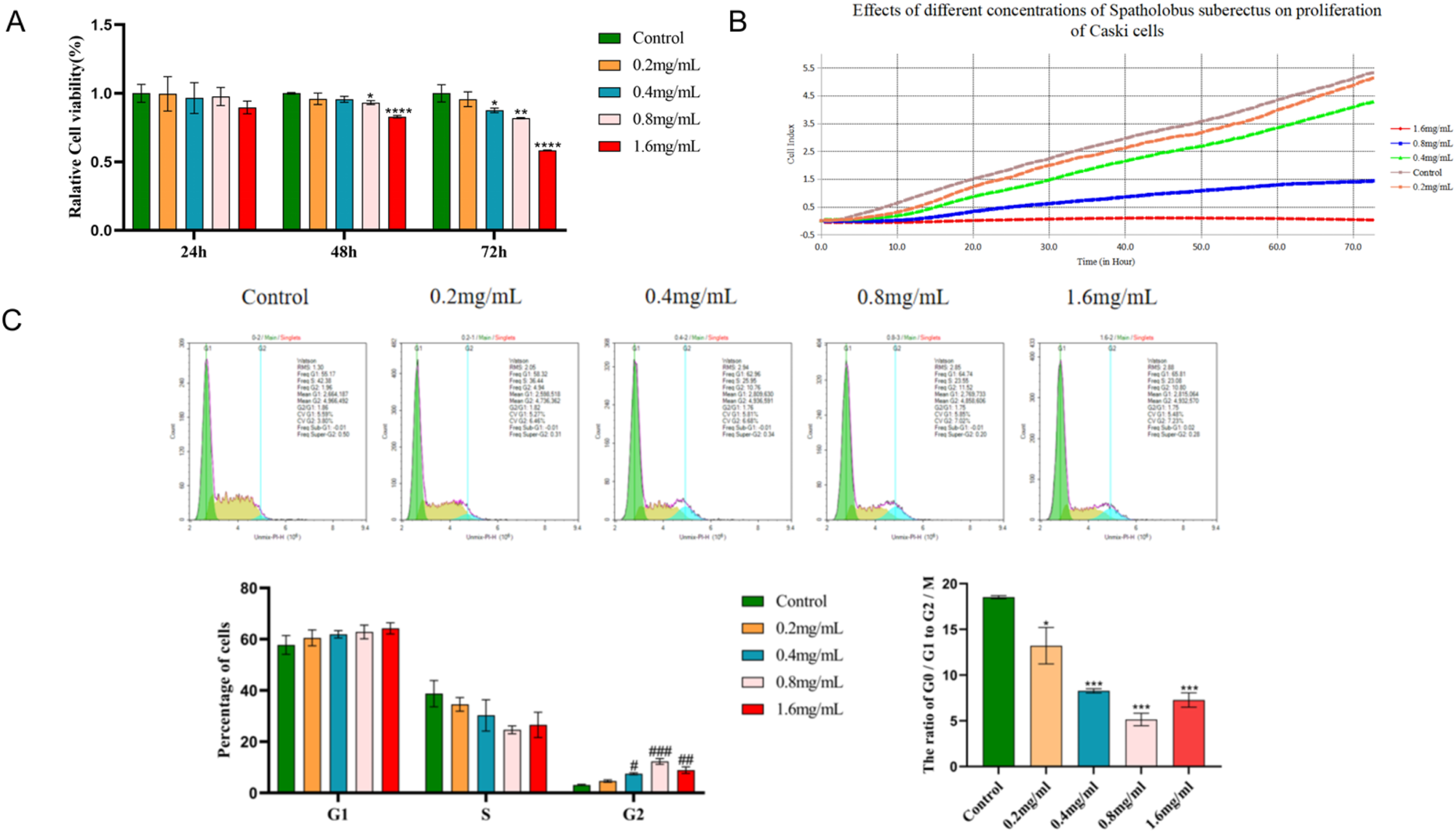
SSD Induces Apoptosis in Caski Cells
Based on the results of the cell viability assay, it was determined that SSD possesses the potential to induce apoptosis in Caski cells. Consequently, an apoptosis flow cytometry assay was conducted. The analysis revealed that both early and late apoptosis rates in Caski cells treated with SSD were elevated compared to the control group. Furthermore, these rates increased proportionally with the concentration of SSD. The total apoptosis rate, defined as the sum of early and late apoptosis rates, was higher in the SSD-treated group (0.2 - 1.6 mg/mL) than in the control group, exhibiting a concentration-dependent trend, as illustrated in Figure 9. These findings suggest that SSD effectively inhibits the migration of Caski cells. Flow cytometry analysis of SSD-induced apoptosis in Caski cells. # # p < 0.01, # represents an increase, compared with the blank group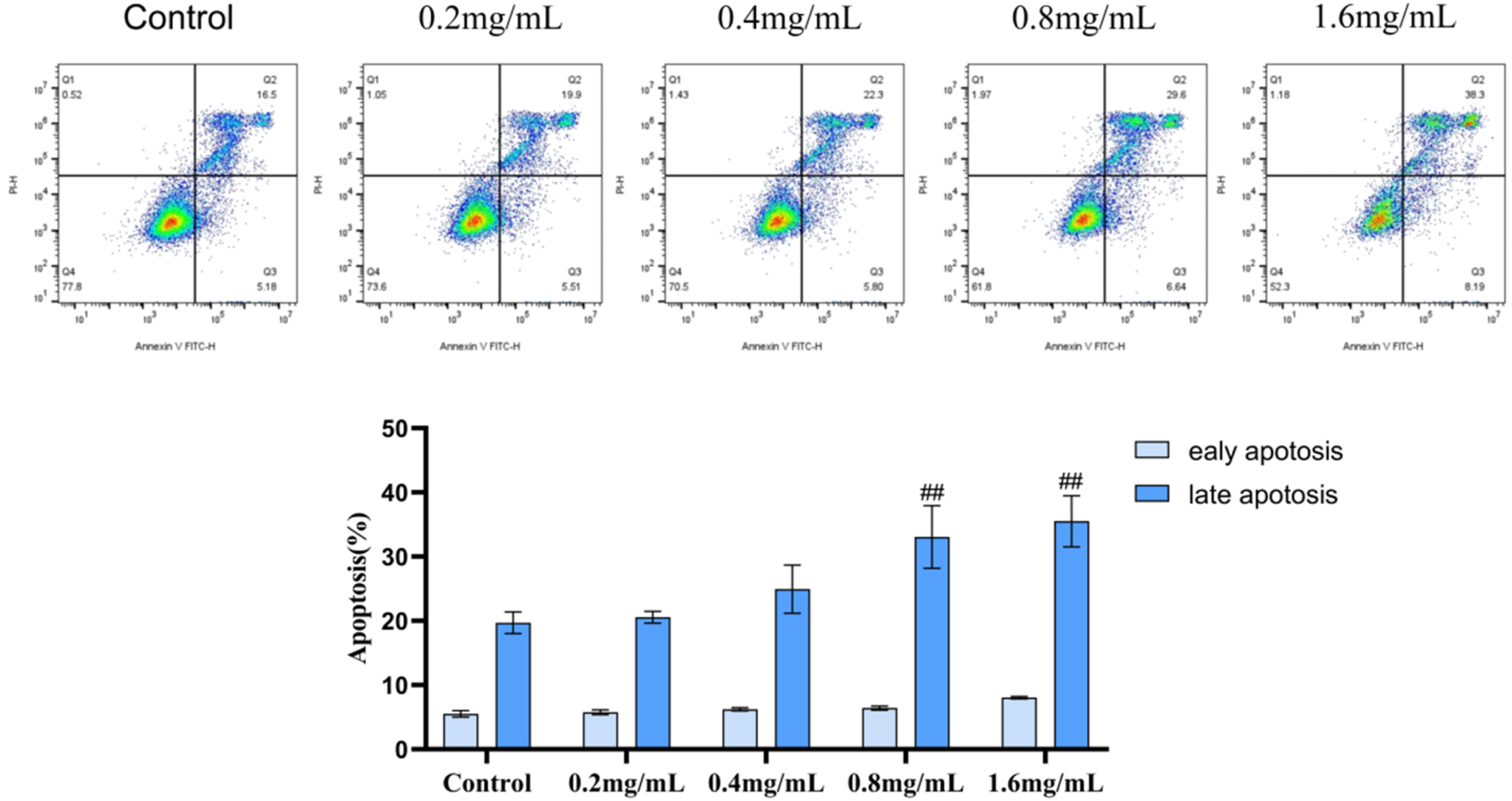
SSD Inhibits the Migration of Caski Cells
To evaluate the inhibitory effect of SSD on the migration of Caski cells, we conducted scratch and RTCA migration assays. Analysis of the scratch assay revealed that the healing area in the 72-hour SSD treatment group was smaller than that in the control group, with a further decrease observed as the SSD concentration increased, thereby indicating a significant inhibition of migration in the SSD-treated group, as illustrated in Figure 10A. Similarly, the RTCA migration results demonstrated that the cell index of the SSD treatment group was lower than that of the control group, exhibiting a concentration-dependent pattern, as illustrated in Figure 10B. These findings collectively suggest that SSD effectively inhibits the migration of Caski cells. SSD inhibits Caski cell migration.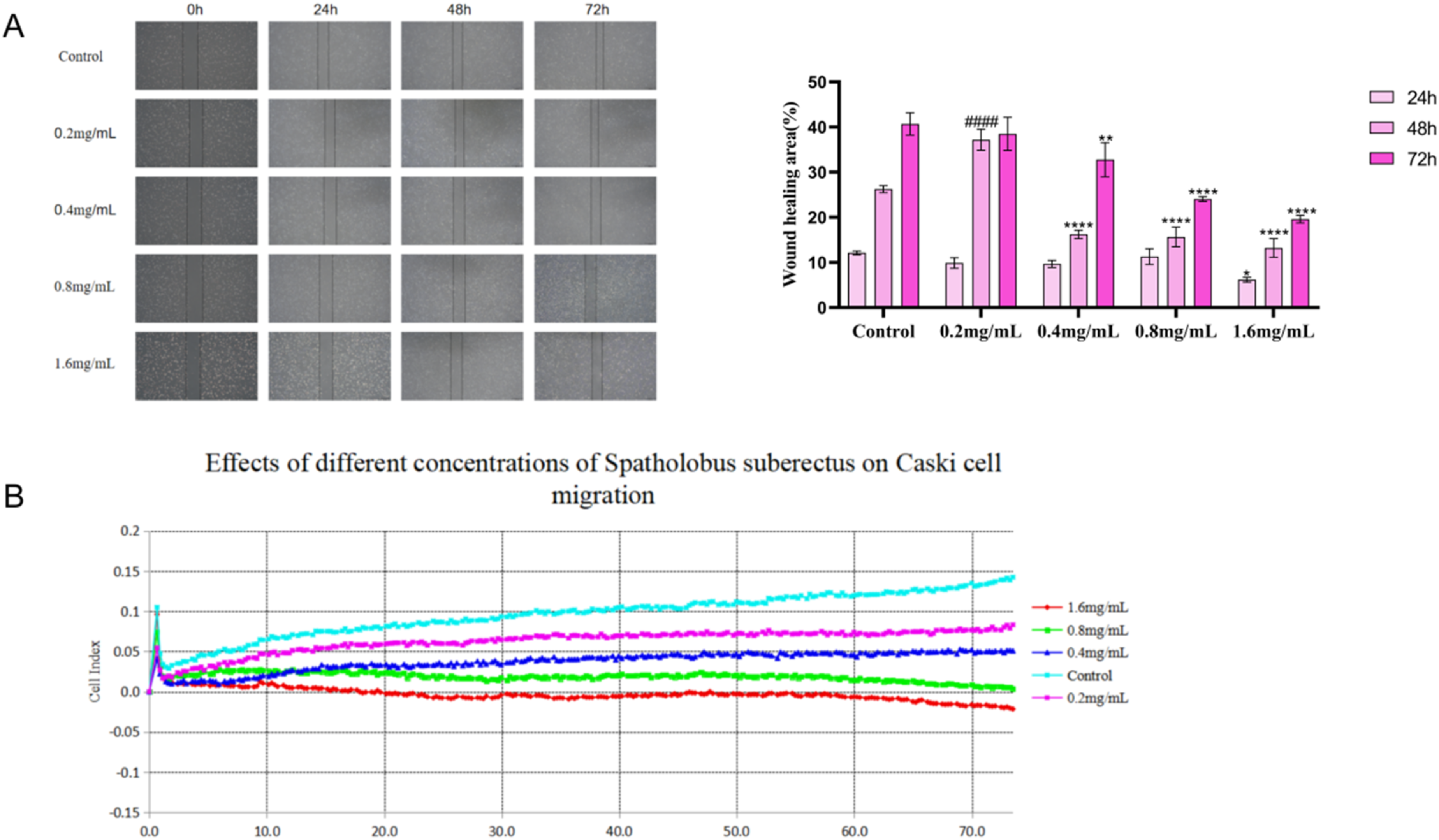
SSD Inhibits the P27, P21, P53, PI3K/Akt, and Jak2/Stat3 Pathways in Caski Cells
Based on network pharmacology analysis, the PI3K/Akt and P53 signaling pathways are likely the central mechanisms through which SSD influences CC. Subsequently, we conducted a detailed examination of the P27, P21, P53, PI3K/Akt, and Jak2/Stat3 pathways utilizing Jess. Post-SSD treatment, there was a significant elevation in the protein levels of P27, P21, and P53 compared to the control group. Furthermore, the expression levels of PI3K and Akt within the PI3K/Akt pathway were reduced in the SSD-treated group relative to the control, and similarly, the expression levels of Jak2 and Stat3 within the Jak2/Stat3 pathway were diminished in the SSD group compared to the control group, as illustrated in Figure 11. These findings suggest that SSD may exert its inhibitory effects on CC through modulation of the P27, P21, P53, PI3K/Akt, and Jak2/Stat3 signaling pathways. Shows Jess-detected expression levels of P27, P21, P53, PI3K/Akt, Jak2/Stat3, and GAPDH proteins across various SSD treatment concentrations. * p < 0.05, * * p < 0.01, * * * p < 0.005, * * * * p < 0.001, # p < 0.05, # # p < 0.01, # # # p < 0.005, # # # # p < 0.001, * represents a decrease, # represents an increase, compared with the blank group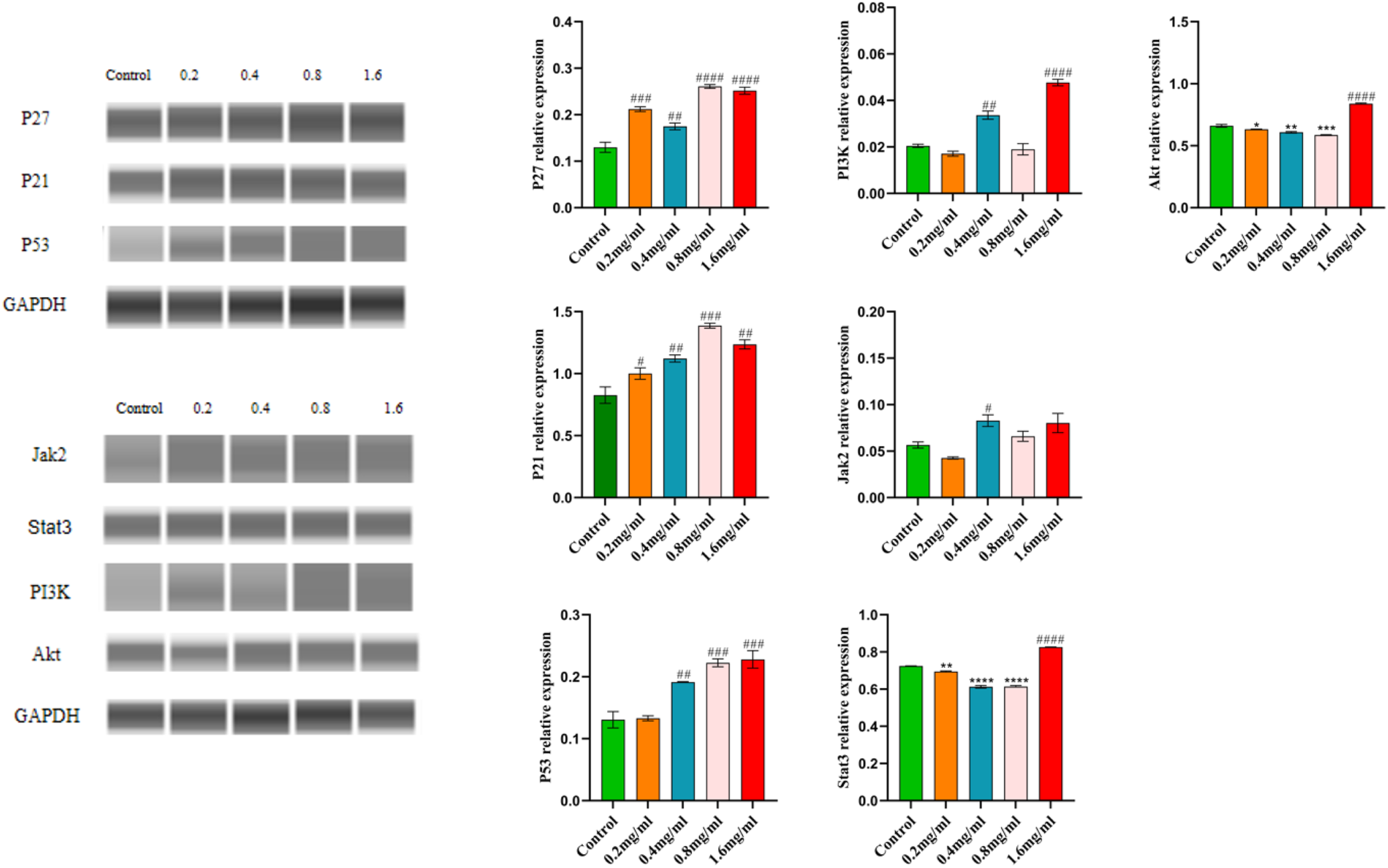
Discussion
CC ranks as the fourth most prevalent malignant neoplasm among women globally 23 . In nations characterized by a high Human Development Index (HDI), the incidence of morbidity is elevated; however, the associated mortality rates remain relatively low. Conversely, countries with low to medium HDI experience both higher morbidity and mortality rates, which can be attributed to limited resources for screening and vaccination programs, as well as an increased mortality risk associated with late-stage diagnoses. 24 Recently, traditional Chinese medicine has attracted increased attention due to its potent anti-cancer properties and minimal toxicity. 25 Thus, exploring the application of traditional Chinese medicine in cancer treatment is currently of considerable importance.
Among traditional Chinese medicinal agents, SSD has demonstrated inhibitory effects on various cancers, including breast cancer, 26 osteosarcoma, 27 ovarian cancer, 28 and lung cancer. 29 The chemical composition of SSD is intricate and varied, primarily comprising flavonoids, terpenoids, and phenolic acids. 30 In this study, the mechanisms and targets of SSD in the context of CC immunotherapy were thoroughly investigated using a network pharmacology approach combined with in vitro experiments, offering novel insights for CC treatment. Network pharmacology introduces a novel paradigm for elucidating and visualizing the potential interaction networks of traditional Chinese medicine in addressing multifactorial diseases. 31 Particularly in the study of traditional Chinese medicine compound prescriptions and natural drugs, network pharmacology exhibits significant advantages. Its “multiple components - multiple targets' characteristic is well-aligned with the holistic mechanism of action inherent in traditional Chinese medicine. 32 During the phase of active ingredient screening and target prediction, a total of 24 compounds and 427 target genes were identified from the SSD, alongside a significant number of genes associated with CC. The constructed compound-target-disease (C-T-D) network elucidated the relationships between the identified drugs and diseases, highlighting the top five compounds based on their Degree values: luteolin (MOL000006), formononetin (MOL000392), licochalcone a (MOL000497), beta-sitosterol (MOL000358), and stigmasterol (MOL000449). These compounds are posited to play a pivotal role in cancer therapeutics. Among the active compounds, luteolin is a flavonoid. 33 Research has demonstrated 34 that luteolin exerts anti-cancer effects by inhibiting tumor cell proliferation, inducing apoptosis in tumor cells, and suppressing their migration and invasion. Formononetin, an isoflavone, 35 has been shown 36 to impede tumor cell growth and survival through various mechanisms. Licochalcone a exhibits notable anti-tumor activity by inhibiting the EGFR signaling pathway. 37 Beta-sitosterol demonstrates anti-cancer properties against breast, prostate, colon, lung, stomach, and ovarian cancers. 38 Additionally, stigmasterol has been found to possess anti-cancer effects across multiple cancer types. 39 These findings suggest that the active constituents of SSD have a pronounced anti-tumor potential; however, further elucidation of the specific targets and pathways involved in the treatment of CC is required.
The analysis of the PPI network identified eight core targets, among which AKT1 and TP53 are significantly associated with biological processes such as cell proliferation and apoptosis. 40 This suggests that SSD may have a pivotal role in the treatment of CC. The KEGG enrichment analysis indicated a high enrichment of the PI3K/Akt and P53 signaling pathways. The PI3K/Akt signaling pathway is crucial in the invasion and metastasis of various solid tumors, 27 and its activation can facilitate tumor cell growth and proliferation.41-43 At the same time, the activation of the PI3K/Akt signaling pathway can substantially increase the expression of PD-L1 on the surface of tumor cells, thereby inhibiting T cell activation and proliferation, which facilitates the evasion of immune surveillance by tumor cells. 44 Conversely, the P53 signaling pathway contributes to tumor suppression by inducing cellular senescence or apoptosis.45-47 Activation of the P53 pathway leads to the upregulation of the downstream target gene P21, thereby arresting the tumor cell cycle. 48 Additionally, the P27 signaling pathway can interact synergistically with the P53/P21 pathway to further inhibit the tumor cell cycle. 49 The activation of the Jak2/Stat3 50 signaling pathway has been shown to facilitate the growth, proliferation, and invasion of tumor cells. At the same time, Stat3 affects the maturation and function of DCs and further affects the activation of T cells by inhibiting the Jak2/Stat5 signaling pathway and reducing the expression of mature markers such as MHC-I, MHC-II, CD80 and CD86 on the surface of DCs, thereby reducing anti-tumor immunity. 51 Molecular docking studies have corroborated the strong binding affinity of the compound to its core target, providing direct evidence for the mechanism of action of SSD. The findings from in vitro experiments were consistent with the results of network pharmacology analysis. SSD was observed to induce apoptosis in Caski cells and significantly inhibit their proliferation and migration. It has been established that the PI3K/Akt and P53 signaling pathways are likely the primary pathways through which SSD exerts its effects on CC. Additionally, it has been demonstrated that SSD may impede the cell cycle of Caski cells via the P53/P21/P27 axis and inhibit the activation of PI3K/Akt and Jak2/Stat3 pathways, thereby obstructing the growth and survival of Caski cells. This experiment has established a foundational basis for the development of novel treatment drugs for CC and is anticipated to advance the clinical application of SSD in CC therapy. At the same time, this study acknowledges several limitations. The mechanisms through which SSD exerts its effects on CC were primarily inferred from network pharmacology, and the functionality of the identified marker gene was validated solely using an in vitro cell model. Furthermore, the verification was restricted to single-cell analyses, which do not adequately replicate the complex in vivo microenvironment and fail to capture the heterogeneity of the cell population. Future research should include multicellular in vitro validations, in vivo animal studies, and clinical trials to comprehensively assess the clinical translational potential of SSD.
Conclusion
In summary, this study conducted a comprehensive analysis of the anti-tumor properties of SSD components utilizing network pharmacology and validated these effects through in vitro experimentation. Additionally, we investigated the potential mechanisms by which SSD exerts its inhibitory effects on CC. The findings suggest that SSD may suppress the proliferation of Caski cells, induce apoptosis, and diminish their migratory capacity via the PI3K/Akt, P53, P27, P21, and Jak2/Stat3 signaling pathways. The precise mechanism will be subject to further verification in future studies. Moreover, this research utilized network pharmacology alongside in vitro experiments to offer novel insights into the treatment of CC.
Supplemental Material
Supplemental Material - Exploring the Role and Targets of Spatholobus suberectus Dunn in Cervical Cancer Therapy Via an Integrated Strategy of Network Pharmacology and in Vitro Experiments
Supplemental Material for Exploring the Role and Targets of Spatholobus suberectus Dunn in Cervical Cancer Therapy Via an Integrated Strategy of Network Pharmacology and in Vitro Experiments by Zhicheng Zhou, Meng Zhang, Ruyan Wen, Meng Xia, Zujie Qin, Yuhan Pu, Hai Long, Yongfang Cheng, Junlong Yu, Xiangling Li in Natural Product Communications
Footnotes
Acknowledgments
I extend my heartfelt gratitude to my mentor, Associate Research Fellow Li Xiangling, for her meticulous guidance throughout the stages of project design, experimental execution, and thesis composition. I am also grateful to Mr. Zhang Meng for his expert guidance in the area of network pharmacology. Furthermore, I acknowledge the financial support provided by the Guangxi Natural Science Foundation (Grant No. 2025GXNSFBA069583), the third batch of the Guangxi University of Traditional Chinese Medicine’s ‘Qihuang Project' high-level talent team training initiative (Project No. 202401), and the National Natural Science Foundation of China (Grant No. 82460921).
Author Contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the [Guangxi Natural Science Foundation] under Grant [number 2025GXNSFBA069583];The Third Batch of “Qihuang Project' High-level Talents Team Training Project of Guangxi University of Chinese Medicine [202401] and National Natural Science Foundation of China [82460921].
Declaration Of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article. We declare that we have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.