
Abstract
Objectives
Cryptic secondary metabolites from bacteria mostly remain undetected under standard cultivation conditions because their biosynthetic gene clusters are not expressed or are only weakly expressed. The present study aimed to discover cryptic secondary metabolites from Burkholderia plantarii using a high-throughput elicitor screening (HiTES) platform.
Methods
HiTES was applied to activate cryptic biosynthetic gene clusters in B. plantarii. Culture extracts derived from elicitor-treated conditions were subjected to repeated column chromatography for compound isolation. The chemical structures of the isolated compounds were elucidated using a combination of 1D and 2D NMR spectroscopic experiments, HR/MS data, and Marfey`s method. All isolated compounds were evaluated for their antibacterial and antifungal activities against pathogenic bacterial and yeast strains.
Results
In the HiTES approach, a cryptic secondary metabolite, plantazine (
Conclusions
HiTES represents an efficient, comprehensive, and broadly applicable approach for the discovery of cryptic secondary metabolites by inducing the expression of silent biosynthetic gene clusters from bacteria.
Keywords
Introduction
Microbial natural products play a crucial role in various ecological functions, including defense mechanisms, communication, and competition for resources and have been a rich source of bioactive compounds with diverse structures and functions, making them important in fields such as medicine, agriculture, and industry.1-3 In the last decade or so, it has become evident that many biosynthetic gene clusters (BGCs) associated with the production of microbial natural products are not consistently expressed under standard laboratory conditions. 4 These “silent” or “cryptic” BGCs, which can be identified computationally through genomic analyses, often do not result in detectable quantities of the corresponding natural products using conventional cultivation methods. This phenomenon poses a challenge for researchers aiming to isolate and elucidate the chemical structures of these potentially valuable compounds. To overcome these challenges, researchers have been investigating alternative cultivation strategies and innovative techniques. 5 This involves employing co-culturing methods, modifying growth conditions, utilizing genetic engineering to activate silent BGCs, and harnessing advanced technologies such as metagenomics and synthetic biology.6-8 Among these efforts, high-throughput elicitor screening (HiTES) was developed to efficiently enable the rapid detection and identification of cryptic secondary metabolites from genetically intractable bacteria.9-11 In the HiTES approach, bacteria are exposed to 500-1000 external elicitors, and the production of natural products is tracked using genetic reporters, bioactivity assays, or mass spectrometry-based analysis.9-11
The Burkholderia genus consists of a diverse group of bacteria, encompassing species that exhibit a broad range of interactions, from beneficial relationships such as plant growth promotion and biocontrol, to pathogenic roles, particularly in causing infections in humans, animals, and plants.
12
Some Burkholderia species are known for their plant growth-promoting properties, while others can be associated with plant diseases.13-15 The genome sequences of B. plantarii revealed the presence of more than 20 BGCs.
16
Various methods, such as the insertion of active promoters, heterologous expression, and reporter-guided mutant selection, have been employed to explore their extensive metabolomes.17-19 As part of our continuing efforts to find structurally and/or biologically novel compounds from various microbes, we applied the HiTES approach on the mesophilic plant pathogen Burkholderia plantarii for the discovery of new cryptic secondary metabolites. As a result, plantazine ( Workflow for HiTES approach on B. plantarii and the resulting induced secondary metabolome. (A) Overnight spore suspension of B. plantarii is suspended in 96 well plates along with elicitors from a small molecule library to stimulate the expression of silent BGCs. After the incubation period, the filtered extracts from each well undergo analysis through the UPLC-Q-TOF-MS experiment. (B) The HiTES approach induces the secondary metabolome of B. plantarii using 320 elicitors. The globally induced metabolome is visualized in a 3D plot that displays the m/z and abundance of each compound as a function of the elicitor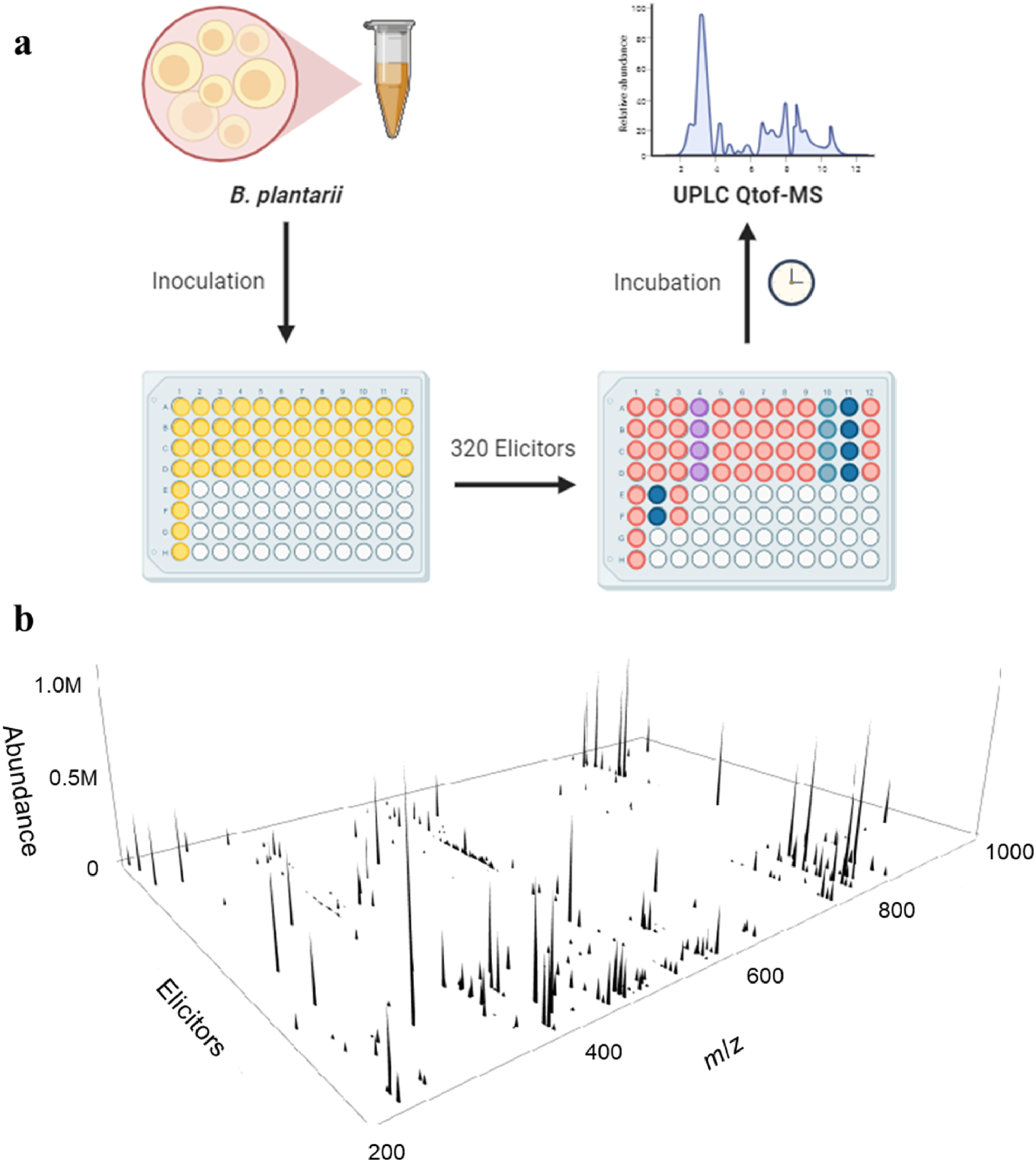
Materials and Methods
General Experimental Procedures
Optical rotations were calculated using a Jasco P-1020 polarimeter (Jasco, Easton, MD, USA). Low-resolution high-performance liquid chromatography-mass spectrometry (HPLC-MS) was carried out on an Agilent 1260 Infinity Series HPLC system coupled with an automated liquid sampler, a diode array detector, and a 6120 Series ESI mass spectrometer. Samples were resolved on an analytical Kinetex C18 column (Phenomenex, 5 µm, 4.6 x 100 mm) with a gradient solvent system of 10-100 % CH3CN over 17 min. CH3CN and H2O contained 0.1 % (v/v) formic acid (FA). High resolution (HR) HPLC-MS was performed on an Agilent 6540 Accurate Mass Q-TOF LC-MS, consisting of an automated liquid sampler, a 1260 Infinity Series HPLC system, a diode array detector, a JetStream ESI source, and a 6540 Series Q-tof. An analytical Kinetex C18 column (Phenomenex, 5 µm, 4.6 x 100 mm) was used to resolve samples. Elution was carried out at 0.5 mL/min with a gradient of 10-100% CH3CN over 18 min. In addition, HR-ESI MS spectra for the analysis of secondary metabolome were recorded using an Agilent 1290 Infinity II system equipped with a 6545 UPLC/QTOF mass spectrometer (Agilent Technologies) coupled with an automated liquid sampler, a diode array detector, and a JetStream ESI source. An analytical Poroshell EC C18 column (Agilent, 2.7 µm, 3.0 x 50 mm) was utilized to resolve samples. Elution was carried out at 0.5 mL/min with a gradient of 10-100% CH3CN over 2.5 min. HPLC purifications were carried out on an Agilent 1260 Infinity Series analytical or preparative HPLC system equipped with a photodiode array detector and an automated fraction collector. NMR spectra, including 1H-1H COSY, HSQC, and HMBC experiments, were carried out using a Bruker 500 MHz NMR spectrometer equipped with a DCH double resonance cryoprobe. For the elicitor validation, all drug stocks were purchased from Sigma-Aldrich or Fisher Scientific. Merck precoated silica gel F254 plates were used for thin-layer chromatography. Spots were detected on TLC under UV light or by heating after spraying with anisaldehyde-sulfuric acid. The present study was conducted at the College of Pharmacy, Pusan National University (Busan, Republic of Korea) from December 2024 to January 2026.
Bacterial Strain
B. plantarii ATCC43733 was obtained from the ATCC. The following media were routinely used to cultivate B. plantarii. MM9 medium (1 L) was prepared as follows; Solution A [350 g K2HPO4 and 100 g KH2PO4 dissolved in 1 L distilled water] and solution B [29.4 g NaCl, 50 g (NH4)2SO4, 5 g MgSO4 dissolved in 1 L distilled water] were autoclaved. 2 g of an amino acid mixture [
High-Throughput Elicitor Screening
High-throughput elicitor screening was performed as previously described with minor modifications.9-11 B. plantarii was inoculated onto an MM9 agar plate (60 mm × 15 mm) from a -80°C frozen culture stock and cultivated at 30°C for 2 days. Fresh spores were collected from the MM9 agar plate and inoculated into 4 mL of fresh MM9 medium in a 14-mL sterile culture tube and then cultured for 2 days at 30°C/250 rpm. The overnight spore suspension was diluted 1000-fold into MM9 liquid medium. Each 150 μL of fresh MM9 medium was distributed into four sterile round bottom 96 well plates (Corning, 0.35 mL per well). Subsequently, elicitors (0.5 μL per well) were added from a commercially available FDA-approved drug library (Enzo Scientific, BML-2843) to give a final concentration of 33 μM. Then, each 96 well plate was covered with Breath-Easy sealing membrane (Sigma) and cultured at 30°C/250 rpm for 3 days. Humidity was sustained in the incubator by incubating several flasks containing distilled water. After 3 days, each well of the 96 well plates was extracted for 4 h using 150 μL of 100% MeOH at 4°C. And then, the 100 μL MeOH extract of each well was loaded onto a 96-well Strata C8-resin plate (Phenomenex) and eluted with 300 μL of 100% MeOH into fresh 96 well plates.
Analysis by UPLC-Q-TOF-MS
The prepared 96 well plates containing the MeOH extract were covered with 96-well cap mats (Thermo Scientific) and 2 μL injected into analytical Poroshell EC C18 column (Agilent, 2.7 µm, 3.0 x 50 mm) on UPLC-Q-TOF-MS. Elution was performed with CH3CN and water as mobile phase (each containing 0.1%, v/v FA) and a gradient system of 10-100% CH3CN over 2.5 min at 0.5 mL/min. HR-MS data was acquired in positive mode (scan range m/z 100 to 3300). After HR-MS data collection, m/z and retention times for secondary metabolites were extracted and aligned across all samples in 96 well plates utilizing Agilent Profinder v. B.06.00. The threshold parameters were set to an ion count of ≥105, affording arranged ions with discrete m/z and retention time. The multi-dimensional datasets were analyzed with MetEx, an in-house application developed for this purpose, as previously described. 20 All detected ions above a selected abundance threshold were binned and subtracted from twice the average value for that bin in the vehicle control-treated sample. Positive values of the resulting difference matrix were displayed in a 3D plot, which shows the m/z and abundance for all induced metabolites as a function of the elicitor. Simple visual examination of 3D plots allowed us to easily identify cryptic secondary metabolites induced by given elicitors.
Validation of HiTES Elicitors
Validation of HiTES elicitors was performed following the previously described HiTES procedure with minor modifications.9-11 In order to validate the production of the plantazine (
Large-Scale Cultivation of B. plantarii
Large-scale cultivation was performed following a similar procedure to that for the small-scale cultivations as described above. For large-scale fermentation of B. plantarii, a -80°C frozen culture stock was used to inoculate onto MM9 agar plate (60 mm × 15 mm) and cultivated at 30°C for period times. Fresh spores on the MM9 plate were transferred into several 4 mL of fresh MM9 medium in a 14-mL sterile culture tube to prepare liquid seed cultures. The seed cultures were transferred into 8 x 50 mL MM9 media (in 125 mL Erlenmeyer flask) and incubated at 30°C/250 rpm for period times. After period time, 8 x 50 mL liquid media were added to 8 x 1 L MM9 liquid media (in 4 L Erlenmeyer flask), corresponding to an inoculum size of 5% (v/v). Each culture was then supplemented with the optimal concentration of the elicitors. The culture was incubated at 30°C/250 rpm for period times.
Purification of Compounds From B. plantarii
All compounds were isolated from 8 L MM9 liquid fermentation of B. plantarii in the existence of ipratropium bromide at a final concentration of 60 μM. After a 3-day fermentation, B. plantarii grown in the MM9 liquid media were solvent-partitioned with ethyl acetate (16 L) and the ethyl acetate extract was completely dried under reduced pressure. The ethyl acetate-soluble extraction (520 mg) was loaded onto a Diaion HP-20 chromatography column and fractionated with 500 mL of each solvent system of 0%, 50%, and 100% MeOH in H2O. Based on the LC/MS analysis, the 100% MeOH miscible fraction was separated using preparative reverse-phase HPLC (Phenomenex Luna phenyl-hexyl, 250 × 21.2 mm, 5 μm, at a flow rate of 10 mL/min) with a gradient solvent system of CH3CN-H2O (1:9 to 1:0, v/v) to afford three subfractions (C1-C5). Fraction C3 (15.5 mg) was isolated by semi-preparative reverse-phase HPLC (Phenomenex Luna C18, 250 × 10.0 mm, 5 μm, at a flow rate of 2 mL/min) with an isocratic solvent system of 51% MeOH to give compound
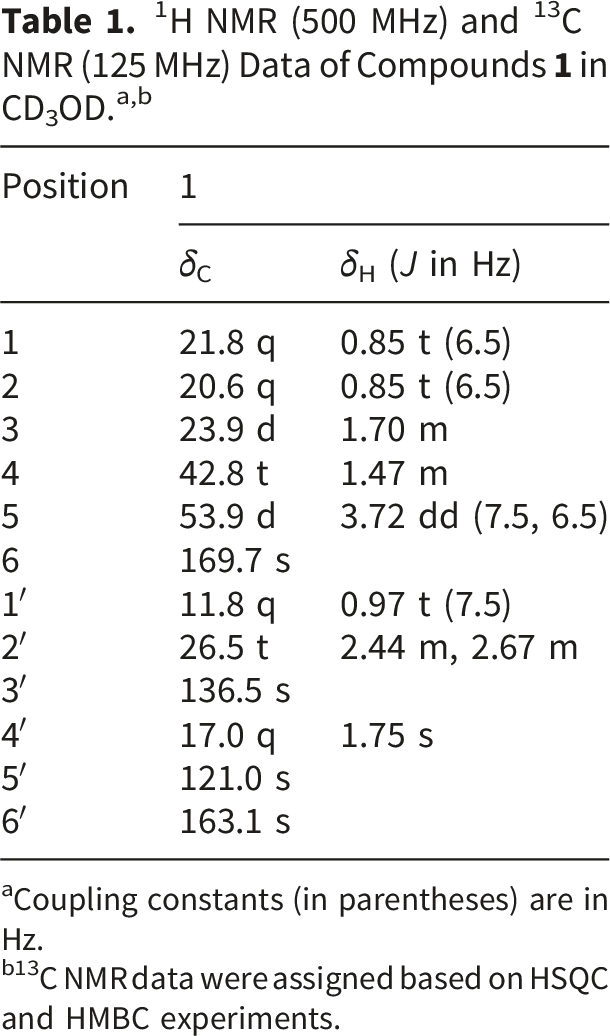
aCoupling constants (in parentheses) are in Hz.
b13C NMR data were assigned based on HSQC and HMBC experiments.
Computational Analysis
Preliminary conformational searches were performed using the Marvin module (ChemAxon). The generated conformers were subjected to energy minimization using the MMFF94 and Dreiding force fields. Conformers located within a 10 kJ/mol energy window, as evaluated by the MMFF94 force field, were chosen for subsequent geometry optimization. Geometry optimizations were performed using Tmolex 4.3.1 at the DFT level with the B3LYP functional and M3 grid, and the convergence criteria were set to 10-6 hartree for energy and 10-3hartree/bohr for the gradient norm (|dE/dxyz|). A mixed basis set was assigned to all atoms.
NMR shielding tensors were additionally calculated at the same level of theory using optimized geometries. The corresponding NMR chemical shifts for each isomer were obtained through Boltzmann-weighted averaging of the calculated 13C NMR shifts for the energetically accessible conformers. Chemical shifts were calculated using the conventional relationship between the computed shielding values and the reference shielding constant: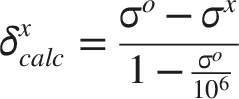
Marey’s Analysis of 1
Compound
Antibacterial and Antifungal Assays
Minimum inhibitory concentration (MIC) and half-maximal inhibitory concentration (IC50) measurements were carried out according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI) using the broth-dilution method. E. coli K12 (37°C), V. cholera (37°C), P. aeruginosa PAO1 (30°C), B. gladioli (30°C), S. aureus (37°C), E. faecalis OG1RF (30°C), and B. subtilis 168 (30°C) were cultured in Muller-Hinton Broth at the indicated temperatures. S. cerevisiae and S. pombe were cultivated in potato-dextrose medium at 30°C. All tested strains were grown in 5 mL overnight cultures in sterile 14-mL culture tubes in the media and at the temperatures indicated. They were then diluted in the same medium to an initial OD600 of 0.005 and dispensed into 96-well plates (100 μL per well). The wells received the following final concentrations of the compounds; 100, 50, 25, 12.5, 6.25, 3.13, 1.56, 0.78, 0.39, 0.19, and 0 μM. The plates were incubated for 18 h without shaking. OD600 values of the plates were obtained using a plate reader. IC50 and MIC values were obtained using a dose-response curve fit and the assays were conducted in triplicates with all isolated compounds.
Results
To perform the HiTES assay in a 96-well setup, we started by pouring liquid media into microtiter plates. Next, a robot added a set of 320 compounds from a candidate elicitor library (FDA-approved drug library) and then we incubated them for 3 days at 30°C/250 rpm. The contents were then extracted by treating each well with methanol, and we analyzed the resulting secondary metabolomes using ultra-high-performance liquid chromatography with quadrupole time-of-flight mass spectrometry (UPLC-Q-TOF-MS) and the recently developed metabolomics explorer (MetEx) analytical software.
We created three-dimensional maps to visualize the induced secondary metabolome of B. plantarii in response to 320 elicitors. For each well, ions exceeding a chosen abundance threshold were grouped and subtracted from double the average value for that bin in the control treated with the vehicle. Positive values in the resulting difference matrix were then showcased in a 3D plot, illustrating the m/z and intensity of all cryptic metabolites concerning the elicitor. The outcomes reveal a rich metabolome for B. plantarii, affirming it as an excellent natural source for discovering concealed secondary metabolites (Figure 1).
In the three-dimensional map of B. plantarii, our attention was directed to one ion signal with m/z 225 (Figure 2). 2D slices extracted from the 3D map demonstrated that the most significant induction of the ion signal occurred when exposed to plant-derived tropane alkaloids like atropine and ipratropium bromide, as well as the antimigraine triptan drug, zolmitriptan (Figure 2). We confirmed the production of the compound in larger liquid media, noting a substantial production increase induced by the addition of ipratropium bromide as an elicitor (Figure 3). Interestingly, the compound was not detected in the DMSO-treated culture (no elicitor). We purified the compound from extensive liquid cultures and these cultures were enriched with ipratropium bromide as an elicitor. Discovery of a cryptic secondary metabolite, plantazine ( Elicitor validation and chemical structures of all isolated compounds. (A) Validation of production of 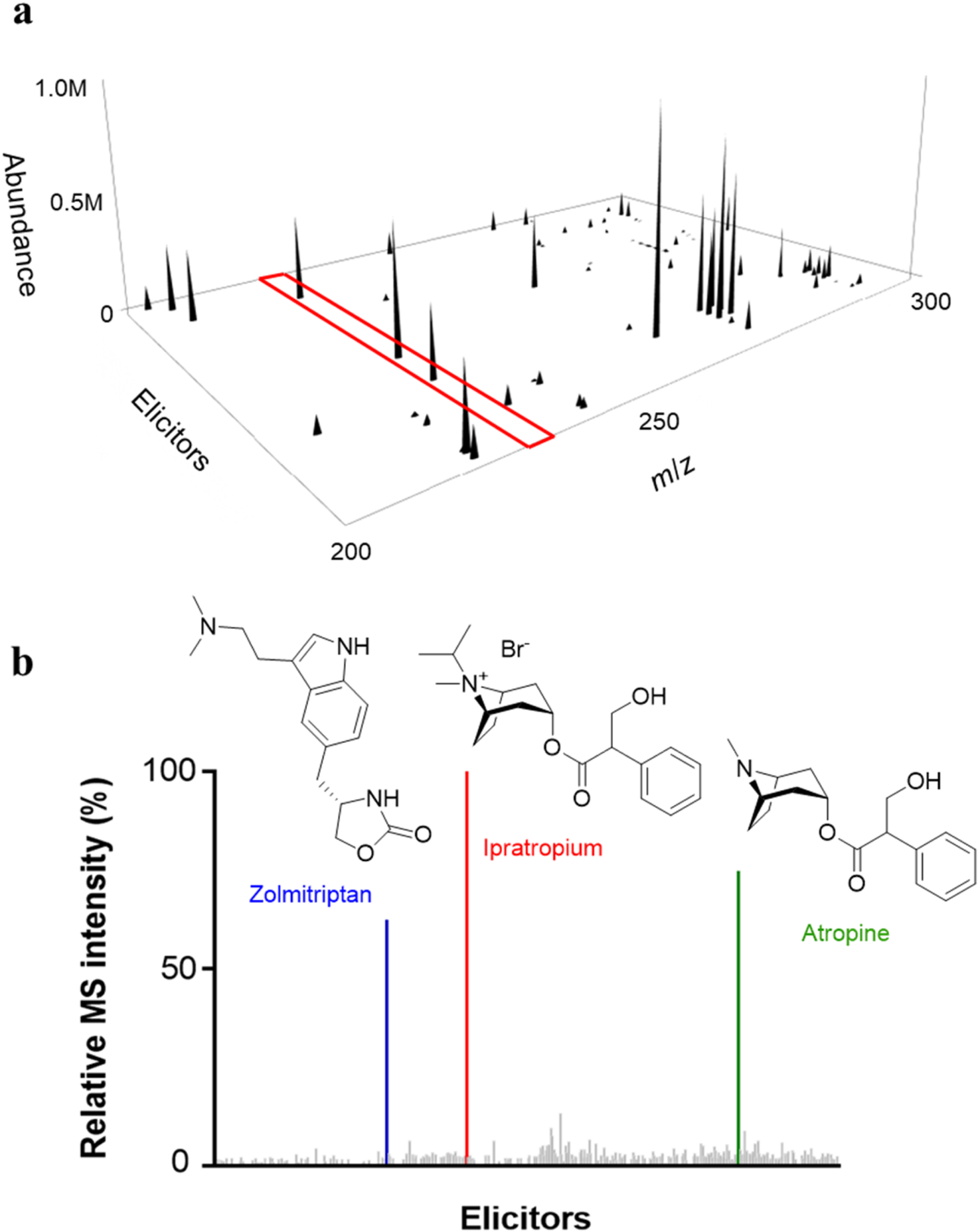
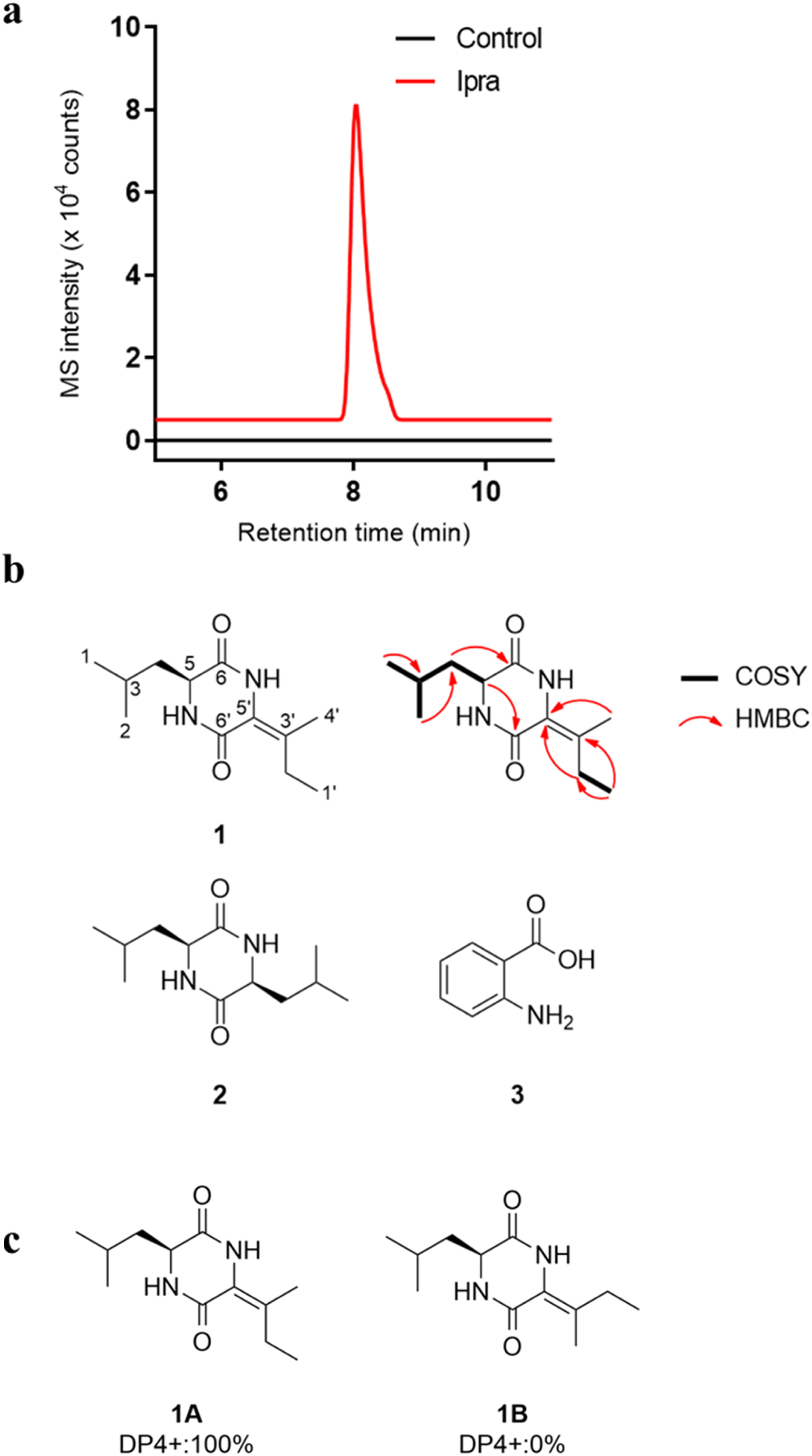
Compound
The known compounds isolated from B. plantarii were identified as (3S,6S)-3,6-disobutylpiperazine-2,5-dione (
All isolated compounds
Discussion
Silent BGCs in bacteria have the potential to encode the biosynthesis of structurally and/or biologically active compounds.22-24 Despite harboring the genetic information for these biosynthetic pathways, these clusters remain inactive or silent under typical laboratory growth conditions. We applied UPLC-Q-TOF-MS-based HiTES, a rapid and comprehensive method that permits the simultaneous detection of globally induced secondary metabolites.25,26 With one well-studied bacterial strain, the LC/MS-based HiTES approach led to the purification and structural elucidation of a cryptic secondary metabolite, plantazine (
Conclusions
In conclusion, the UPLC-Q-TOF-MS-based HiTES approach enabled the discovery of a new cryptic secondary metabolite, plantazine (
Supplemental Material
Supplemental material - Discovery of a Cryptic Secondary Metabolite, Plantazine, From the Mesophilic Plant Pathogen Burkholderia plantarii
Supplemental material for Discovery of a Cryptic Secondary Metabolite, Plantazine, From the Mesophilic Plant Pathogen Burkholderia plantarii by Yeo Jin Lee, Eun Seo Jang, Hyeon-Jeong Hwang, Youngsang Nam, Tae Gyun Kim, Chaeyun Hong, Eun La Kim, Mohammad R. Seyedsayamdost, and Seoung Rak Lee in Natural Product Communications.
Footnotes
Acknowledgements
The authors are grateful to the College of Pharmacy, Pusan National University, South Korea for NMR analysis of the isolated compounds.
Consent to Participate
There are no human subjects in this article and informed consent in not applicable.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by 2025 Junior Faculty Support Program of Pusan National University and the Korea Basic Science Institute (National research Facilities and Equipment Center) grant funded by the Korea government (MSIT) (RS-2024-00403999). This work was also supported by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIT) (RS-2025-23525419).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Statement of Human and Animal Rights
This article does not contain any studies with human or animal subjects.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.