
Abstract
Mesenchymal stem cells are multipotent progenitor cells that can differentiate into osteogenic, adipogenic, and chondrogenic lineages. Existing methods for modifying mesenchymal stem cells differentiation rely heavily on chemical inducers and synthetic growth factors; however, these strategies are often associated with high costs, poor stability, and potential adverse effects. Therefore, growing interest has been devoted to natural bioactive compounds as safer, more readily available alternatives. Epigallocatechin gallate, the most abundant catechin in green tea, has been identified as a promising regulatory agent of mesenchymal stem cells fate in multiple original studies. This mini-review summarizes and provides a critical analysis of current in vitro evidence regarding the impact of unmodified epigallocatechin gallate on human mesenchymal stem cells differentiation, specifically highlighting osteogenic stimulation and adipogenic suppression—thereby elucidating its potential utility in bone regeneration.
Keywords
Introduction
Stem cells, residing in both embryonic and adult tissues, possess the innate capacity for self-renewal and differentiation into diverse cell lineages. 1 However, not all stem cells exhibit the same differentiation potential, and based on this characteristic, stem cells are classified into several types: these are totipotent, pluripotent, multipotent, oligopotent, and unipotent. 2 Among these, totipotent stem cells such as zygotes exhibit the highest differentiation potential, as they are capable of forming both embryonic and extra-embryonic structures. Pluripotent stem cells—such as embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs)—can differentiate into cells of the ectodermal, mesodermal, and endodermal germ layers; while they lack the capacity to generate extra-embryonic structures such as the placenta3,4. Multipotent stem cells are capable of differentiating into multiple related cell types within a specific tissue or lineage. Furthermore, oligopotent stem cells can differentiate into a limited range of closely related cell types, whereas unipotent stem cells are capable of differentiating into only a single cell type. 5
Mesenchymal stem cells (MSC) were initially harvested from bone marrow and subsequently isolated from various other tissues. 6 While MSC differentiation has traditionally been modulated by synthetic growth factors, cytokines, and small molecules; these agents may elicit adverse reactions or, in some instances, induce malignant transformation7-11 Consequently, phytochemicals have been extensively investigated as alternative regulators of stem cell fate due to their comparative safety, cost-effectiveness, and potent biological activity.11,12 Some phytochemicals have demonstrated promising results in enhancing stem cell proliferation, survival, and differentiation.13-15 Among these, epigallocatechin gallate (EGCG) is a major polyphenolic compound found in green tea which has been widely studied for its antioxidant, anti-inflammatory, and osteoprotective properties. 16 Emerging evidence indicates that EGCG can modulate human MSC (hMSC) differentiation into osteogenic and adipogenic lineages by influencing critical signaling pathways and transcriptional networks.17-20 The present review provides a concise synthesis of current findings regarding the role of unmodified EGCG in directing hMSC fate toward these specific lineages.
Mesenchymal Stem Cell Characteristics and Differentiation
MSC are non-hematopoietic stem cells that can give rise to cells of ectodermal (neurocytes), endodermal (hepatocytes), and mesodermal (adipocytes, osteocytes, chondrocytes) lineages. Typical sources for their isolation include bone marrow, adipose tissue, wharton’s jelly, peripheral blood, umbilical cord, placenta, and amniotic fluid. 21 MSC in culture exhibit typical features, including plastic adherence and positive expression of CD105, CD73, and CD90. Hematopoietic markers—including CD45, CD34, CD14, CD11b, CD79α, CD19, and HLA-DR—are not detected in these cells. 22
MSC differentiation is often initiated by adding lineage-specific factors to the culture medium. Dexamethasone, ascorbic acid, and bone morphogenetic proteins such as BMP2 are frequently needed for osteogenic differentiation, while dexamethasone, insulin, 3-isobutyl-1-methylxanthine, and indomethacin are used to induce adipogenic differentiation. 23 Additionally, the number of passages affects the differentiation ability, with early-passage cells maintaining higher multipotency than extensively passaged cells. 24
Within living organisms, MSC undergoes osteogenic differentiation through a tightly regulated series of stages. This process is initiated by the recruitment and proliferation of progenitor cells to the bone formation site, and then progresses through their transformation into preosteoblasts. Subsequently, preosteoblasts transform into mature osteoblasts. Finally, the deposition of Ca2+ mineralizes the extracellular matrix. The mineralized matrix traps certain osteoblasts, which then develop into osteocytes.25-27 Mature osteoblasts secrete alkaline phosphatase (ALP), 28 which hydrolyzes phosphate esters into phosphate and contributes to the mineralization of the extracellular matrix. 29 ALP serves as an early indicator of osteogenesis. 30 Osteoblast differentiation and maturation are controlled by key transcription factors, such as Runx2 and Osterix.31,32 Signaling pathways such as BMP/Smad, Wnt/β-catenin also regulate the process.33-35 Several microRNAs are also involved in the osteogenic process by targeting specific signaling pathways. For example, miR-27a, miR-346, and miR-142-3p regulate the Wnt/β-catenin pathway,36,37 while miR-21 modulates the PI3K/Akt/β-Catenin pathway. 38 Conversely, two important transcription factors, C/EBPα and PPARγ, play central roles in controlling adipogenic differentiation. 39 Activation of C/EBPβ commences the adipogenic program, increases the expression of PPARγ and C/EBPα, and induces adipocyte-specific genes such as adiponectin.40-43
Moreover, MSC exhibit therapeutic properties through the production of bioactive molecules and possess significant immunoregulatory capabilities.
44
MSC evade attack by alloreactive T cells due to their lack of major histocompatibility complex
Epigallocatechin gallate—green Tea Polyphenol
Green tea is manufactured from the leaves of Camellia sinensis. These leaves contain flavanols, which are typically converted by the enzyme polyphenol oxidase into dark polyphenolic compounds responsible for the characteristic color of black tea. In the production of green tea, the leaves undergo a steaming process to deactivate polyphenol oxidase, thereby preserving the green color. Another key step is rolling, which involves cutting and twisting the leaves. 48 The primary active components in green tea are polyphenols, which constitute approximately 8–12% of the tea’s dry weight. Specifically, green tea polyphenols are classified as catechins. The four primary catechins found in green tea are epicatechin (EC), epigallocatechin (EGC), epicatechin gallate (ECG), and epigallocatechin-3-gallate (EGCG). 49 Approximately 59% of the total catechins consist of EGCG, making it the predominant catechin in green tea.50,51 A 250 ml cup of green tea typically provides between 50 and 100 mg of EGCG; However, a daily intake greater than or equal to 800 mg when taken as a food supplement could be hepatotoxic.52,53 The bioactivities of EGCG—including osteogenic, cardioprotective, antioxidant, anti-inflammatory, anti-obesity, and neuroprotective effects—are critically influenced by the eight hydroxyl groups present in its chemical structure. 54 Green tea polyphenols were effective in mitigating bone loss in preclinical models of age-related, estrogen-deficient, and inflammation-induced bone degeneration.55-57 The ovariectomized (OVX) rat model is widely used to study osteoporosis, exhibiting clinical features of postmenopausal estrogen deficiency and subsequent bone loss.58,59 In OVX rats, EGCG attenuated bone loss and downregulated the expression of Semaphorin 4D, a known inhibitor of bone formation. 60 EGCG not only attenuated bone loss but also improved bone microarchitecture and upregulated BMP2 expression in OVX rats. 61 EGCG effectively preserved bone density in OVX rabbits, while simultaneously reducing bone marrow adiposity and normalizing bone turnover rates. 62 EGCG restored bone mechanical strength in rats with femoral deformities or tibial fractures by upregulating BMP-2.63,64 In a rat model of dexamethasone-induced osteoporosis, EGCG treatment attenuated the deterioration of bone quality and improved bone microarchitecture. 65 Specifically, it downregulated the expression of ALP, Runx2, OCN, and PPARγ, while upregulating the protein levels of β-catenin and Wnt3a in mice with secondary osteoporosis. 66 Taken together, these findings provide a robust rationale for investigating the impact of EGCG on the lineage-specific differentiation of MSC.
However, EGCG shows low bioavailability because of its instability and extensive metabolism. Following oral administration, it undergoes hydrolysis in the oral cavity, followed by hepatic biotransformation and further modification by gut microbiota. Various strategies have been employed to improve bioavailability, such as co-administration with dietary components (e.g., vitamin C, xylitol, or certain foods) or the use of advanced delivery systems, including double emulsions and nanoparticle-based formulations. 67
Literature Selection
The databases Scopus, Web of Science, and PubMed were searched until 1st January 2025 using the following keywords: (((Green tea polyphenol) OR (Green tea flavonoids) OR (epigallocatechin gallate) OR (epigallocatechin-3-gallate) OR (EGCG)) AND ((stem cell differentiation) OR (mesenchymal stem cell differentiation))). An updated search was conducted on 19 April 2026. The PRISMA 2020 flow diagram was used in the search process.
68
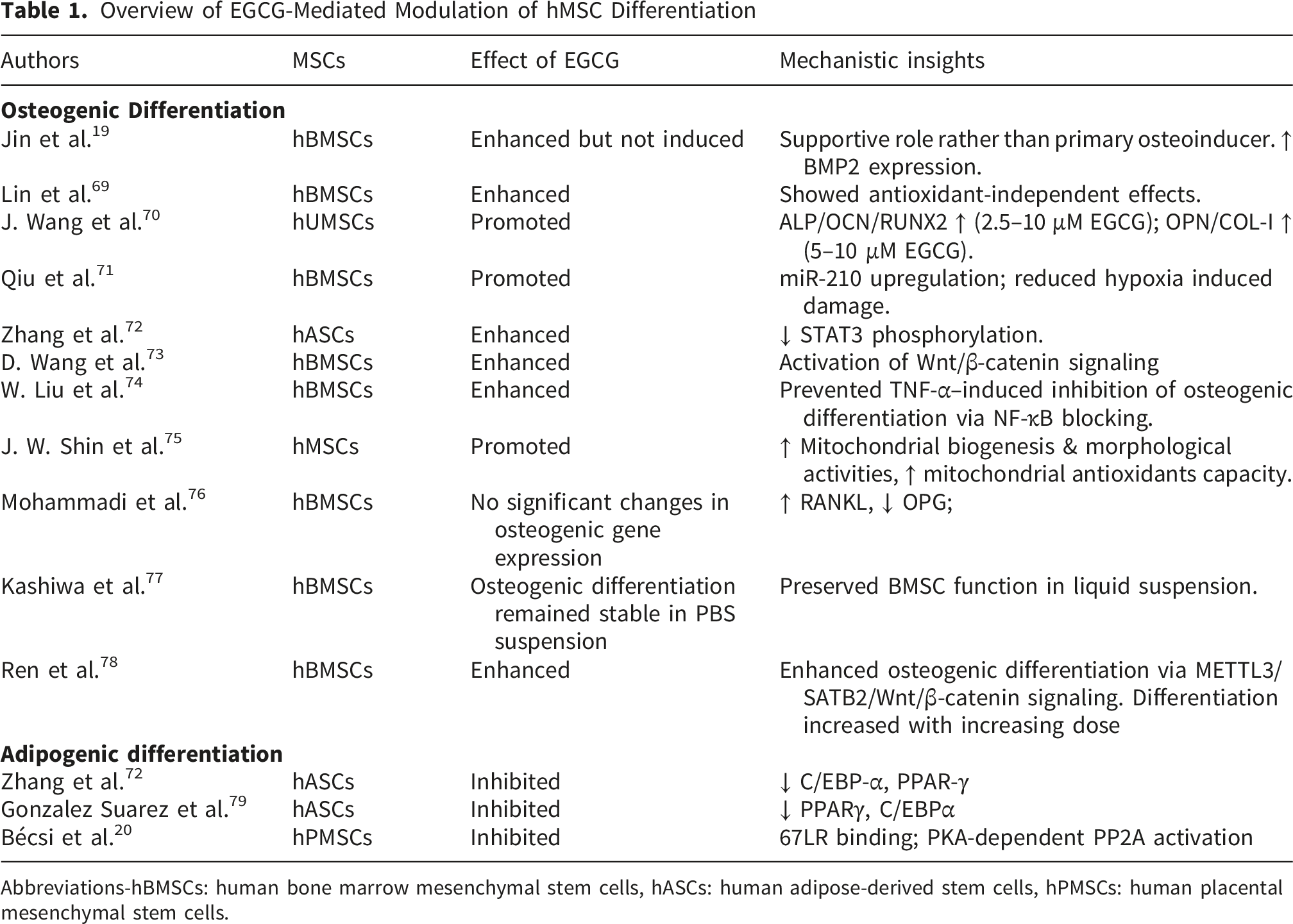
The process of searching for articles is shown in Figure 1. This review included studies (Table 1) that evaluated the impact of EGCG on the in vitro differentiation of hMSCs into osteogenic or adipogenic lineages. Studies involving modified EGCG or co-treatment regimens were excluded. Ultimately, 13 articles met the eligibility criteria. Flow diagram of the literature search process Overview of EGCG-Mediated Modulation of hMSC Differentiation Abbreviations-hBMSCs: human bone marrow mesenchymal stem cells, hASCs: human adipose-derived stem cells, hPMSCs: human placental mesenchymal stem cells.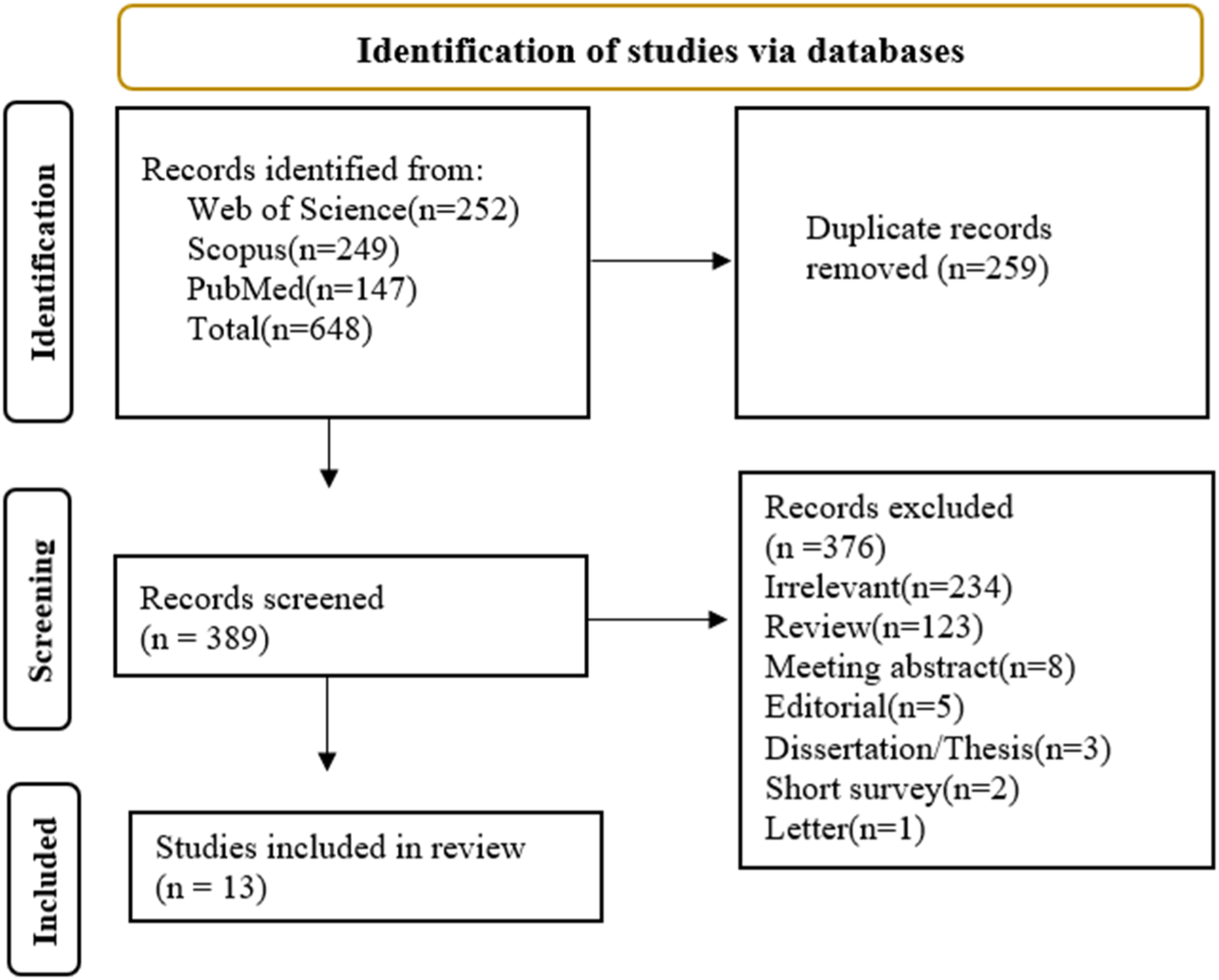
Effects of EGCG on hMSC Viability and Proliferation
Low concentrations of EGCG (5–10 μM) enhanced cell viability in human adipose-derived mesenchymal stem cells (hASCs), 72 human bone marrow-derived MSCs (hBMSCs). 73 Another study reported that in hBMSCs, cell viability progressively increased with higher EGCG concentrations (10 < 20 < 40 < 80 μM). 78 In human umbilical cord-derived mesenchymal stem cells (hUMSCs), EGCG (<10 μM) did not affect the cell viability. 70 Additionally, EGCG markedly attenuated the H2O2-mediated reduction in cell viability in hBMSCs 73 and also mitigated the inhibitory effects of TNF-α on hBMSCs, thereby restoring normal viability and reducing cell death. 74 EGCG (5 µM) also promoted the proliferation of hBMSCs, 19 while concentrations below 10 µM enhanced growth in hUMSCs. 70 Notably, EGCG treatment preserved the high viability and proliferative capacity of hBMSCs, even after being suspended in phosphate-buffered saline (PBS) for 4 days at 4°C 77 .
Effects of EGCG on Osteogenic Differentiation of hMSC
Epigallocatechin gallate has been extensively investigated as a regulator of osteogenic differentiation in hMSCs. Current evidence indicates that it does not function as a primary osteoinductive agent but rather as a potent enhancer of osteogenesis, particularly under osteogenic conditions. 19 Consistent with this role, multiple in vitro studies using hBMSCs have demonstrated significant modulation of key osteogenic transcription factors and markers including RUNX2, BMP2, ALP, osteocalcin, osteonectin, and COL1A1; alongside increased alkaline phosphatase activity and extracellular matrix mineralization.19,69,71 Consistent with these findings, Lin et al showed that osteogenic differentiation persists even when its antioxidant activity was suppressed, suggesting mechanisms that extend beyond redox regulation. 69 In line with this, several studies reported a dose-dependent promotion of osteogenesis.69,70,72,74 Nevertheless, one study reported reduced OPG and increased RANKL expression in hBMSCs, without significant changes in osteogenic gene expression. 76
At the molecular level, these effects appear to involve multiple signaling pathways. For instance, osteogenic differentiation in hBMSCs has been linked to activation of METTL3-mediated m6A methylation of SATB2, which subsequently stimulates the Wnt/β-catenin signaling pathway. 78 Additionally, the compound counteracts oxidative stress–induced inhibition of osteogenesis by promoting β-catenin and cyclin D1 expression in hBMSCs, 73 while in hASCs it enhanced osteoblast differentiation through downregulation of STAT3. 72
Beyond its pro-osteogenic activity, it also exhibits notable cytoprotective properties under adverse conditions. Under hypoxic condition, it restored osteogenic potential of hBMSCs by upregulating miR-210 and suppressing its target EFNA3, while simultaneously reduced apoptosis and functional impairment. 71 In inflammatory conditions, EGCG prevented TNF-α-mediated suppression of osteogenesis by inhibiting NF-κB signaling in hBMSCs. 74
Interestingly, at higher concentrations (25 μM), it enhanced osteogenic differentiation in hMSCs, as evidenced by increased expression of VEGF and BMP2. This effect was accompanied by upregulation of mitochondrial biogenesis markers (PGC-1α and mtTFA) and antioxidant genes (SOD1, SOD2, CAT, and GPX1), suggesting a coordinated improvement in both osteogenic and mitochondrial function. However, the specific hMSC source used in this study was not specified.
75
Mechanistic roles of EGCG in osteogenic differentiation were provided in Figure 2. Mechanisms of EGCG-mediated osteogenesis in hMSCs. EGCG enhances differentiation and mineralization by modulating METTL3/SATB2, Wnt/β-catenin, and STAT3 pathways. Furthermore, EGCG counteracts the inhibitory effects of hypoxia and inflammation on osteogenic potential by regulating miR-210 and the NF-κB pathway, respectively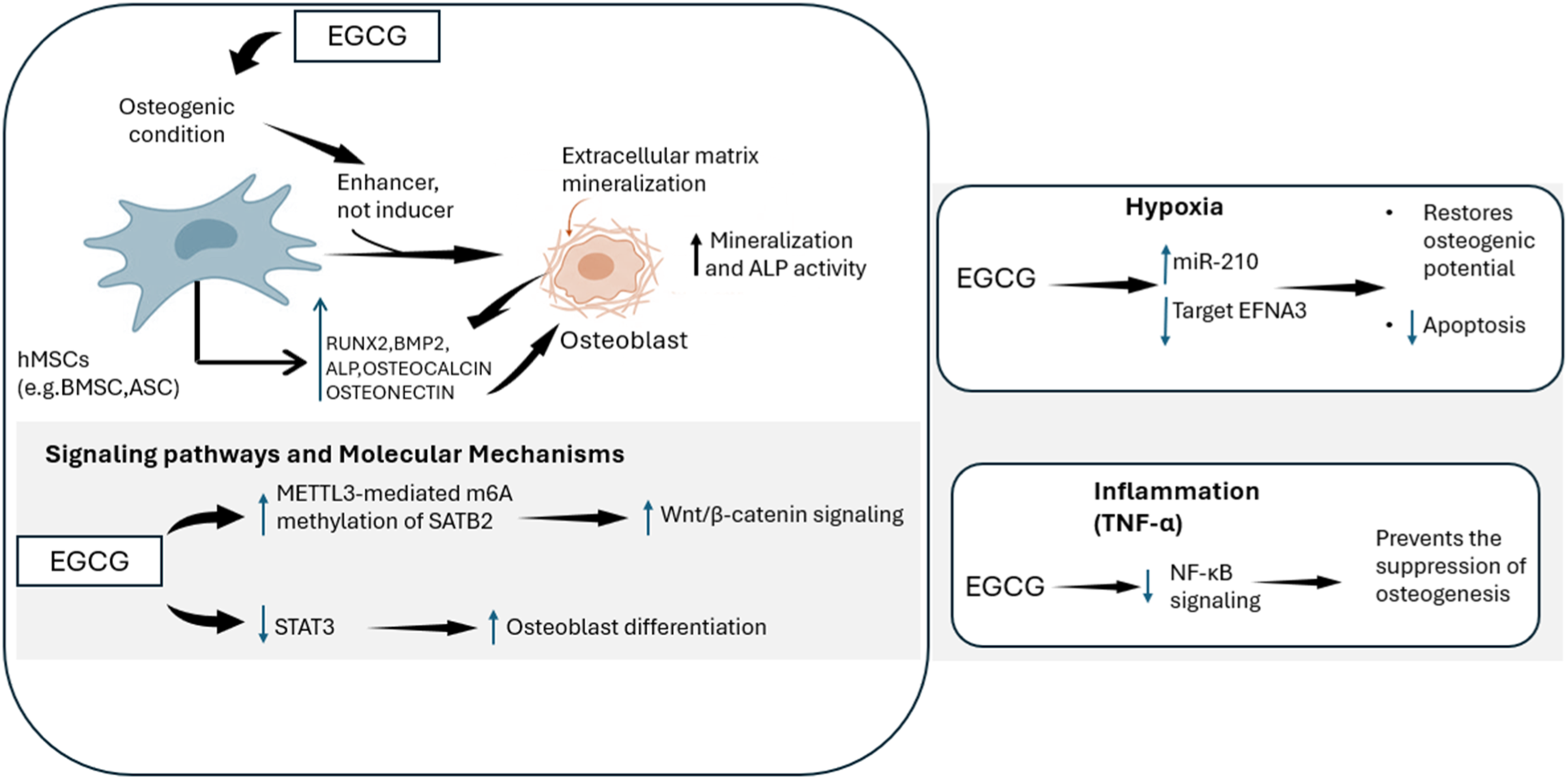
Effects of EGCG on Adipogenic Differentiation of Human MSC
EGCG suppressed adipogenesis in hASCs by attenuating PPARγ and C/EBPα, leading to diminished lipid droplet accumulation and decreased expression of adipocyte-specific genes.72,79 EGCG bound to the 67-kDa laminin receptor (67LR), causing PKA-dependent activation of protein phosphatase-2A (PP2A). This signaling cascade decreased myosin phosphatase activity and impaired adipogenic transcriptional pathways, resulting in the inhibition of PPARγ-driven differentiation. 20
Conclusion
EGCG is a potent natural modulator of hMSC differentiation, stimulating osteogenesis while simultaneously inhibiting adipogenesis. It upregulates osteogenic markers (RUNX2, BMP2, ALP, osteocalcin) and increases matrix mineralization via BMP–Smad and Wnt/β-catenin signaling. Moreover, it mitigates oxidative stress, inflammation, and hypoxia-induced dysfunction. However, its impacts are dose-dependent (optimal at 5–10 μM). EGCG mainly acts as an enhancer, not as an inducer. Future studies should clarify its molecular targets, pathway crosstalk, long-term effects, and translational potential. EGCG represents a low-cost, biologically active agent with promise for stem cell–based bone regenerative therapies.
Footnotes
Author Contributions
MMH & ST contributed equally in conceptualization, data extraction, manuscript writing, review, and editing. YT reviewed the scientific contents described in the manuscript. All authors read and approved the final submitted version of the manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Figure Declarations
Figure 1 was developed using the PRISMA flow diagram template. ![]() was original work designed and created by the authors.
was original work designed and created by the authors.