
Abstract
Objective
The increasing incidence of fungal contamination, mycotoxin-related hazards, and antifungal resistance has intensified the search for sustainable alternatives to conventional fungicides. This study investigated the antifungal and antisporulation activity of thyme essential oil (Thy-EO) against Aspergillus flavus, Aspergillus parasiticus, Penicillium expansum, and Penicillium verrucosum.
Methods
The activity of Thy-EO was assessed through agar well diffusion method, disk-volatilization method, broth microdilution method, fungal fitness, and sporulation assays. Antifungal efficacy was evaluated by measuring growth inhibition zones, minimum inhibitory concentrations (MICs) and minimum fungicidal concentrations (MFCs), fungal survival over time, and changes in spore production following treatment with increasing Thy-EO concentrations.
Results
Thy-EO inhibited the growth of all tested fungi, producing inhibition zones ranging from 10.3 to 42.8 mm, at tested concentrations 5 µL/well and 10 µL/well in the diffusion assays. MIC/MFC values were 1/2.5 µL/mL for A. flavus, 0.5/1 µL/mL for A. parasiticus, 0.25/1 µL/mL for P. expansum, and 0.25/0.5 µL/mL for P. verrucosum. The volatile fraction exhibited a more marked antifungal activity, generating zones of inhibition up to 36.3 mm against P. verrucosum at the highest concentration (5 µL/disc) used in the disk-volatilization assays performed in this study. Fungal fitness decreased in a concentration- and time-dependent manner, with P. verrucosum and A. parasiticus showing the highest susceptibility. Sporulation was strongly affected in Penicillium spp., with complete inhibition observed at 5 µL/mL for P. expansum and at ≥2.5 µL/mL for P. verrucosum. In contrast, Aspergillus spp. showed approximately 50% reduction in spore production only at the highest concentration tested.
Conclusion
Thy-EO demonstrated both antifungal and antisporulation activities, supporting its potential application as a natural and sustainable tool for fungal control in food preservation and indoor hygiene.
Keywords
1. Introduction
Fungal infections are increasingly difficult to manage due to the rise of multidrug-resistant strains, which has reduced the effectiveness of available antifungal therapies and highlighted the need for alternative and sustainable control strategies within the broader context of antimicrobial resistance. This issue represents a growing global public health concern and underscores the urgency of developing new antimicrobial approaches.1-3 In particular, airborne fungi represent one of the most relevant components of atmospheric bioaerosols. Humans are continuously and involuntarily exposed to fungal spores through daily inhalation, with estimated exposure levels ranging from 1,000 to 10 billion spores per individual per day, resulting in constant interaction between the respiratory system and potentially pathogenic microorganisms. 4 Despite this constant exposure, most antifungal studies still primarily evaluate inhibition of mycelial growth, while the impact on sporulation and spore viability remains comparatively underexplored, even though these parameters are key determinants of environmental persistence, transmission, and contamination risk. 5 Among the fungal genera most frequently detected in both indoor and outdoor air, Aspergillus and Penicillium are particularly prominent, owing to their high sporulation capacity and remarkable adaptability to diverse environmental conditions.6,7 The genus Aspergillus encompasses a highly diverse group of fungi that are ubiquitous in the environment and among the most widespread worldwide. Within this genus, Aspergillus section Flavi includes several species, notably Aspergillus flavus, Aspergillus parasiticus, and Aspergillus nomius, which are commonly associated with soil and plant-derived substrates and are well known for their ability to produce aflatoxins, highly toxic and carcinogenic secondary metabolites. 8 In parallel, the increasing emergence of antifungal resistance among Aspergillus species has progressively compromised the efficacy of conventional therapies, representing a major global public health concern. 9 Beyond antifungal resistance and vegetative growth, the high sporulation capacity of Aspergillus species plays a central role in environmental persistence and airborne dissemination. However, sporulation and spore viability are still rarely considered primary endpoints in antifungal efficacy studies,10,11 despite their relevance for transmission dynamics.
Beyond Aspergillus, filamentous fungi belonging to the genus Penicillium are also widely distributed in indoor and agricultural environments, where they constitute a significant source of concern due to their capacity to contaminate food products and produce toxic secondary metabolites. Spores of Penicillium species are frequently detected in indoor air, and several species are known to synthesize mycotoxins such as citrinin, patulin, and penicillic acid, posing potential risks to food safety and human health. 12 Among these, Penicillium expansum represents a particularly problematic postharvest pathogen of plant host tissues, especially fruits, due to its widespread occurrence and intrinsic resistance. As a necrotrophic fungus and a major producer of patulin, P. expansum can rapidly colonize plant host tissues following spore contact, especially through minor surface injuries, thereby contributing to food spoilage and potential toxin-mediated risks for consumers. 13 In addition, the ecological success and pathogenic potential of Penicillium species are strongly linked to their ability to produce large quantities of resilient airborne conidia, which serve as primary vectors for food contamination and environmental spread.
Fungal contamination also represents an emerging challenge in the field of food safety, not only because of the deterioration of the organoleptic properties of food products, but also due to the potential development of toxin-mediated foodborne illnesses. 14 Filamentous fungi belonging to the genera Aspergillus and Penicillium are able to colonize a wide range of food commodities, particularly under conditions of elevated temperature and humidity. 15 Several species within these genera produce mycotoxins, a heterogeneous group of toxic secondary metabolites that represent a major concern for food safety and pose a serious threat to both human and animal health.16-18
In response to these challenges, the use of natural compounds, such as essential oils, has emerged as a promising area of research due to their richness in bioactive molecules and broad-spectrum antimicrobial properties. Thyme essential oil (Thymus vulgaris L.) has attracted increasing interest for its broad-spectrum antimicrobial activity, largely attributable to phenolic compounds such as thymol and carvacrol.19-22 In vitro studies have demonstrated that these molecules can interfere with multiple cellular processes in pathogenic microorganisms, supporting a multitarget mode of action that may be particularly effective against strains resistant to conventional antifungal agents.23,24 The antifungal activity of essential oils is complex and multifactorial. Plant volatile organic compounds contained in essential oils are small, lipophilic molecules characterized by low molecular weight and high vapor pressure, which enable them to readily diffuse across cell membranes. 25 Their antifungal effects are primarily associated with the disruption of fungal cell membrane integrity, alteration of lipid organization, and increased membrane permeability, leading to leakage of intracellular components, collapse of membrane potential, and ultimately impaired cell viability or fungal cell death.26,27 In addition to these direct effects, thyme essential oil has been shown to inhibit key virulence- and resistance-related mechanisms, including biofilm formation, mycotoxin production, and quorum sensing, which are critical for fungal pathogenicity and persistence.24,28 The main bioactive constituents—thymol, carvacrol, and p-cymene—exhibit strong lipophilic properties that facilitate their interaction with fungal membranes. 29 Consequently, the antifungal efficacy of essential oils is likely the result of synergistic interactions among multiple components rather than the action of a single molecule. 30 Despite the extensive literature on the antifungal properties of thyme essential oil, most studies have focused primarily on inhibition of mycelial growth, while its effects on sporulation and spore viability remain comparatively underexplored. This represents an important knowledge gap, as fungal spores are the principal agents of environmental dissemination, airborne transmission, food contamination, and human exposure.
In this framework, the present study evaluated the activity of thyme essential oil (Thymus vulgaris L.) against selected Aspergillus and Penicillium isolates of environmental and food relevance. Although the antifungal properties of thyme essential oil have been widely documented, its impact on fungal sporulation and spore viability remains comparatively underexplored. Therefore, the novelty of the present work lies in the integrated assessment of antifungal, anti-sporulation, and sporicidal activities. By simultaneously targeting mycelial growth, conidial production, and spore viability, the study explores a dual-action antifungal strategy aimed at reducing not only fungal colonization but also the generation and dissemination of infective propagules. This approach is particularly relevant for indoor environments and food-related settings, where spore-mediated contamination often represents a greater challenge than vegetative fungal growth alone. The findings provide a more comprehensive evaluation of the biocontrol potential of T. vulgaris essential oil and support the development of sustainable strategies for controlling fungal bioaerosols and spore-mediated contamination, thereby contributing to environmental quality, food safety, and human health within a One Health framework.
2. Materials and Methods
This in vitro study was conducted between January 2025 and April 2026 at the Department of Science and Technology, University of Sannio, Benevento, Italy. All antifungal assays were performed under controlled laboratory conditions.
2.1. Thyme Essential Oil (Thy-EO)
The thyme essential oil (Thy-EO) used in this study was purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany; product details and code: thyme oil – white FG, W306509) and stored in the dark at 25 °C until use to preserve its chemical composition and prevent potential alterations due to light or temperature fluctuations. According to the manufacturer’s specification sheet, the major constituents of the tested oil were thymol (43.52%), p-cymene (31.65%), linalool (5.38%), and carvacrol (5.11%). No additional in-house GC-MS characterization was performed in the present study. Therefore, the chemical composition of Thy-EO was reported according to the manufacturer’s specification sheet. In particular, the physicochemical characterization of the thyme essential oil, including the ATR–FTIR spectrum (attenuated total reflectance Fourier-transform infrared spectroscopy), was obtained from the technical documentation provided by the manufacturer and is available at the following link: https://www.sigmaaldrich.com/deepweb/assets/sigmaaldrich/quality/spectra/263/215/ATIR0018422.pdf. The product was supplied as a food-grade, Halal- and Kosher-certified essential oil and complied with IFRA guidelines, EU Regulations 1223/2009 and 1334/2008, and FDA 21 CFR 182.2 requirements. According to the manufacturer’s specification sheet, the oil was characterized by a density of 0.917 g/mL at 25 °C, refractive index (n20/D) of 1.502, optical rotation [α]20/D of −1° (neat), and boiling point of 195 °C. Thy-EO was used either undiluted, for direct-contact and vapor-phase assays, or directly dispersed, in fungal growth media. Prior to microbiological assays in aqueous systems, the oil was thoroughly homogenized by vortex mixing (3 min), according to previously validated procedures,19-21 to obtain a transiently uniform dispersion phase. Given the intrinsic hydrophobicity of essential oils, this approach results in a temporary dispersion under agitation, which was immediately used for downstream assays to minimize phase separation effects. All dilutions were freshly prepared immediately before use to ensure stability and reproducibility of the experimental conditions. Since no organic solvents or emulsifiers were used, in order to avoid potential interference with the antifungal activity of Thy-EO, no additional vehicle control was required. Untreated controls consisted of Thy-EO-free medium or distilled water, processed under the same experimental conditions.
2.2. Fungal Isolates and Growth Conditions
The antifungal activity of thyme essential oil (Thy-EO) was evaluated against four environmental fungal strains: Aspergillus flavus IZPS1, Aspergillus parasiticus IZPS2, Penicillium expansum IZPS3 and Penicillium verrucosum IZPS4. The fungal isolates were characterized and provided by the Istituto Zooprofilattico Sperimentale (IZPS, Portici, Naples, Italy). Aspergillus flavus IZPS1 and Aspergillus parasiticus IZPS2 were isolated from environmental samples, whereas Penicillium expansum IZPS3 and Penicillium verrucosum IZPS4 were isolated from fruit and wheat samples, respectively. Species-level identification was confirmed using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) with the VITEK MS Prime system and the VITEK MS MOULD Kit (bioMérieux, Marcy-l’Étoile, France), according to the manufacturer’s instructions. Identification was based on characteristic protein spectral fingerprints, and a confidence value of 99.9% was obtained for all isolates, representing the maximum score achievable by the system. Fungal cultures were maintained under aerobic conditions at 25 ± 2 °C using Sabouraud Dextrose Agar (SDA) or Broth (SDB; CONDA, Madrid, Spain). Stock cultures were stored at −80 °C in SDB supplemented with 10% (v/v) glycerol. Prior to use in experiments, working cultures were reactivated on solid or liquid media at 25 °C for 48 hours to ensure active growth for subsequent in vitro antifungal assays.
2.3. Antifungal Assays
2.3.1. Agar Well Diffusion Method
To assess the in vitro antifungal activity of thyme essential oil (Thy-EO) against filamentous fungi, an agar well diffusion assay was performed according to standardized protocols, with minor modifications (Perez et al, 1990).20,21,31 A standardized spore suspension (1 × 106 spores/mL) was prepared from 5-day-old fungal cultures. Briefly, fungal mycelium was gently scraped from the culture surface, suspended in phosphate-buffered saline (PBS, 1X; Sigma Aldrich, St. Louis, MO, USA), and subsequently filtered through sterile gauze to recover spores entrapped within the fungal biomass. Spore concentration was determined using a Bürker counting chamber (Bürker–Türk hemocytometer, Knittel Glass, Braunschweig, Germany) and further confirmed by measuring the optical density at 600 nm. Aliquots (100 µL) of the standardized spore suspension were uniformly spread onto Sabouraud dextrose (SD) agar plates using a sterile cotton swab. After inoculation, wells of 6 mm in diameter were aseptically punched into the agar using a sterile glass Pasteur pipette. Each well was filled with defined volumes of Thy-EO (5 and 10 µL). Distilled water was used as negative control, while tioconazole (≥97%, Sigma-Aldrich, St. Louis, MO, USA) at a final concentration of 1 mg/well was used as positive control. Plates were incubated at 25 ± 2 °C for 48 h. After incubation, antifungal activity was evaluated by measuring the mean diameter of the inhibition zones (MDIZ), expressed in millimeters, around each well, including the diameter of the well. All experiments were performed in triplicate using independent fungal cultures.
2.3.2. Disk-Volatilization Method
To evaluate the antifungal activity of the volatile components of thyme essential oil (Thy-EO), an agar disk-volatilization assay was performed, following the method described by Gaare et al., 32 Sateriale et al and Di Rosario et al.,19,22 with minor modifications. Briefly, 100 µL of a standardized spore suspension (1 × 106 spores/mL), obtained from 5-day-old fungal cultures, was uniformly spread onto the surface of Sabouraud dextrose (SD) agar plates using a sterile cotton swab. Sterile filter paper disks were impregnated with different volumes of Thy-EO (2.5 and 5 µL) and placed on the inner surface of the Petri plate lid, ensuring that the essential oil did not come into direct contact with the agar medium.
A 5% sodium hypochlorite solution (NaClO; Sigma-Aldrich, St. Louis, MO, USA) was used as positive control, while distilled water served as negative control. Plates were incubated upside down at 24 ± 2 °C for 48 h, allowing the volatile compounds to diffuse and act on the fungal culture. Antifungal activity was assessed by measuring the mean diameter of the inhibition zones (MDIZ), expressed in millimeters, produced by the volatile compounds of Thy-EO. All assays were conducted in triplicate using independent fungal cultures.
2.3.3. Broth Microdilution Method
The susceptibility of Aspergillus and Penicillium isolates to increasing concentrations of thyme essential oil (Thy-EO) was evaluated using a broth microdilution assay adapted from the Clinical and Laboratory Standards Institute (CLSI) M38 guideline for filamentous fungi. 33 For each strain, standardized spore suspensions (1 × 106 spores/mL) were prepared in phosphate-buffered saline (PBS, 1X; Sigma Aldrich, St. Louis, MO, USA). Aliquots (20 µL) of the spore suspensions were inoculated into wells containing Sabouraud dextrose (SD) broth with increasing concentrations of Thy-EO (0.25, 0.5, 1, 2.5, and 5 µL/mL). Cultures were incubated at 25 ± 2 °C for 48 h under constant agitation to ensure homogeneous distribution of the essential oil micelles in the broth. The minimum inhibitory concentration (MIC) was defined as the lowest concentration of Thy-EO that completely inhibited visible growth of spores and mycelium in the wells. To determine the minimum fungicidal concentration (MFC), aliquots from each well were inoculated onto SD agar plates and examined for mycelial growth after incubation. All assays were performed in triplicate using independent cultures.
2.3.4. Fungal Fitness Method
To evaluate the effect of thyme essential oil (Thy-EO) on the growth and survival capacity of environmental fungal isolates, a fungal fitness assay was performed by adapting previously reported colony-count-based fungal viability procedures.33,34 Standardized spore suspensions (1 × 106 spores/mL) obtained from 5-day-old cultures were filtered through sterile gauze and incubated at 25 ± 2 °C in the presence or absence of Thy-EO at concentrations corresponding to ½ MIC, MIC, 2× MIC, and 4× MIC. At time intervals of 0, 24, 48, and 72 h, decimal dilutions of each sample were prepared and plated onto Sabouraud dextrose (SD) agar. Plates were incubated at 25 ± 2 °C for 48–72 h until visible mycelial growth was observed. Fungal fitness was determined by calculating colony-forming units (CFU) and comparing the growth of Thy-EO-treated samples with untreated controls. All assays were performed in triplicate using three independent fungal cultures to ensure reproducibility of the results.
2.4. Fungal Sporulation Assay
To evaluate the effect of thyme essential oil (Thy-EO) on spore production by the tested fungal isolates, fungal sporulation inhibition assays were performed, with modifications to the method described by Sateriale et al. 34 Fungal strains were initially grown on Sabouraud dextrose (SD) agar for 5 days at 25 ± 2 °C. Small agar fragments covered with mycelium were then transferred into 1 mL of SD broth containing increasing concentrations of Thy-EO (½ MIC, MIC, 2× MIC, and 4× MIC). After one week of incubation under appropriate growth conditions, samples were filtered through sterile gauze to remove excess agar, and 1 mL of sterile distilled water was added to maintain the original concentration of Thy-EO in each tube. Following an additional 24 h incubation at 25 ± 2 °C, 10 µL aliquots from each sample were collected for spore counting under a light microscope (Nikon Eclipse E600, Nikon Instruments, Tokyo, Japan) using a Bürker counting chamber (Bürker–Türk hemocytometer, Knittel Glass, Braunschweig, Germany). All assays were performed in triplicate using independent fungal cultures.
2.5. Statistical Data Analysis
Statistical analyses were not adapted from an experimental protocol but were performed according to standard statistical procedures implemented in GraphPad Prism 8 software (GraphPad Software, San Diego, CA, USA). All experiments were performed using three independent biological replicates, each consisting of three technical replicates (n = 9 per experimental condition for antifungal assays). Prior to statistical analysis, data distribution was assessed using the Shapiro–Wilk normality test. Homogeneity of variances was evaluated using the ANOVA assumption tests implemented in GraphPad Prism to ensure the validity of parametric testing. Statistical significance was assessed using one-way ANOVA followed by Dunnett’s post hoc multiple-comparison test for comparisons between treated samples and controls, and two-way ANOVA followed by Bonferroni’s post hoc correction for time- and concentration-dependent analyses. Exact adjusted p-values are reported in the Supplementary Material (Table S1).
3. Results
3.1. In vitro Antifungal Activity of Thyme Essential Oil Against Aspergillus and Penicillium spp.
The in vitro assays demonstrated a notable antifungal activity of thyme essential oil (Thy-EO) against the tested environmental isolates of Aspergillus and Penicillium spp.
The efficacy of Thy-EO was first evaluated using the agar well diffusion method, which revealed clear inhibition zones of mycelial growth. The mean diameters of inhibition zones (MDIZ) ranged from 25.33 ± 0.47 mm (Thy-EO 10 µL vs A. parasiticus, Figure 1A) to 10.33 ± 0.47 mm (Thy-EO 5 µL vs A. flavus, Figure 1B), compared to the positive control (tioconazole, 17.33 ± 0.94 mm for A. flavus and 23.67 ± 0.47 mm for A. parasiticus). For Penicillium, MDIZ ranged from 42.83 ± 0.62 mm (Thy-EO 10 µL vs P. verrucosum, Figure 1C) to 31.00 ± 0.82 mm (Thy-EO 5 µL vs P. expansum, Figure 1D), with the positive control yielding 27.33 ± 0.47 mm for P. expansum and 30.00 ± 0.82 mm for P. verrucosum. In vitro antifungal activity of thyme essential oil (Thy-EO) against Aspergillus flavus IZPS1 (A), Aspergillus parasiticus IZPS2 (B), Penicillium expansum IZPS3 (C), and Penicillium verrucosum IZPS4 (D), evaluated using the agar diffusion assay. Inhibition zone diameters (mm) are reported as mean ± SD of three independent experiments performed in technical triplicate (n = 9). Statistical significance compared with the positive control was determined by one-way ANOVA followed by Dunnett’s post hoc test (p *< 0.05; **p < 0.01; ***p < 0.001; ***p < 0.0001). MDIZ, mean diameter of inhibition zones; Thy-EO, thyme essential oil; TCZ, tioconazole (1 mg/well)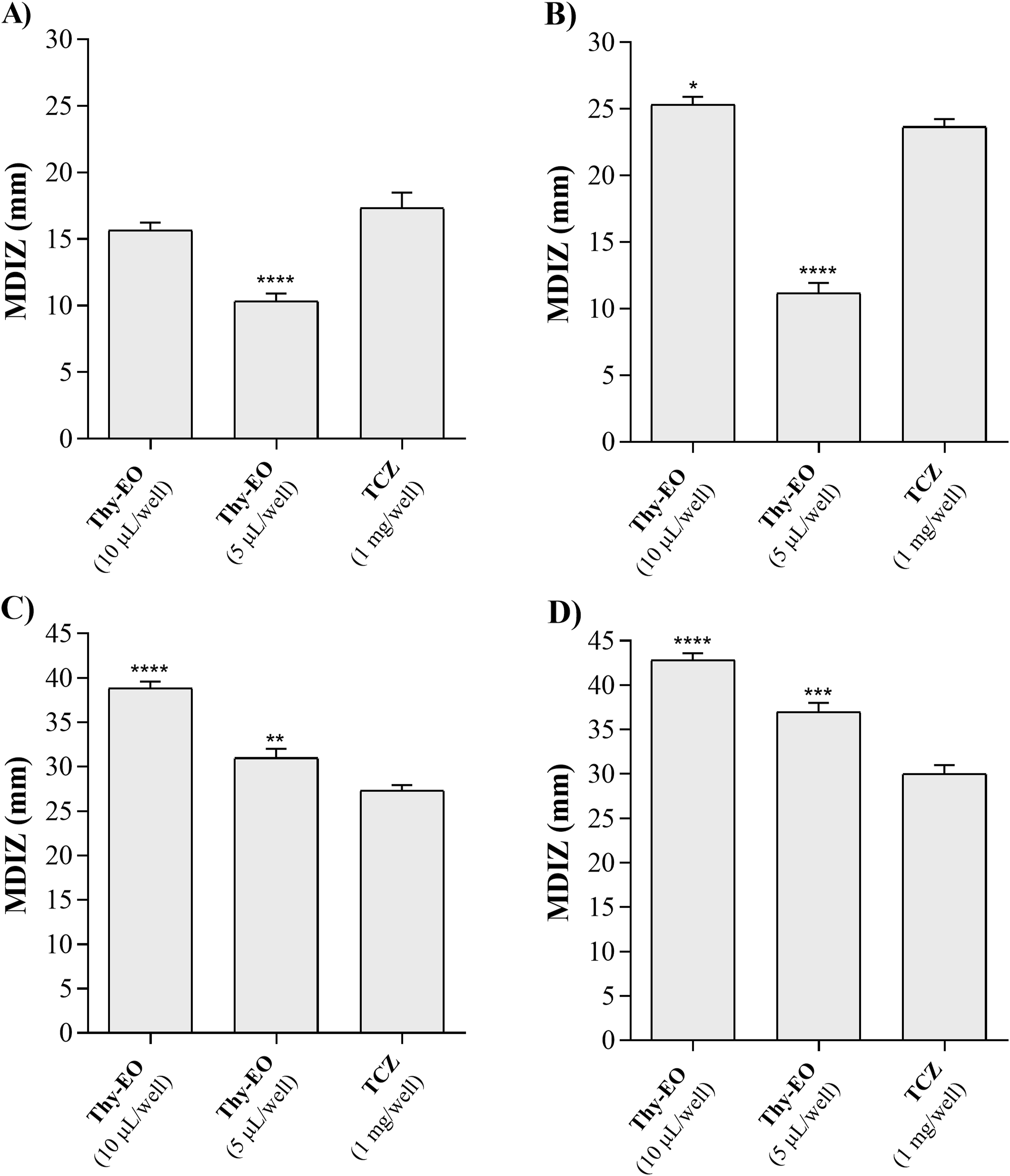
Representative images of the in vitro antifungal activity of Thy-EO, assessed using the agar well diffusion method against A. flavus, A. parasiticus, P. expansum, and P. verrucosum, are shown in Figure S1.
The disk-volatilization assay was performed to assess the antifungal activity of the volatile components of Thy-EO (Figure 2). In vitro antifungal activity of thyme essential oil (Thy-EO) against Aspergillus flavus IZPS1 (A), Aspergillus parasiticus IZPS2 (B), Penicillium expansum IZPS3 (C), and Penicillium verrucosum IZPS4 (D), evaluated using the disk volatilization method. Inhibition zone diameters (mm) are reported as mean ± SD of three independent experiments performed in technical triplicate (n = 9). Statistical significance compared with the positive control was assessed by one-way ANOVA followed by Dunnett’s post hoc test (**p < 0.01; ***p < 0.001). MDIZ, mean diameter of inhibition zones; Thy-EO, thyme essential oil; NaClO 5%, sodium hypochlorite 5% (10 µL/disk)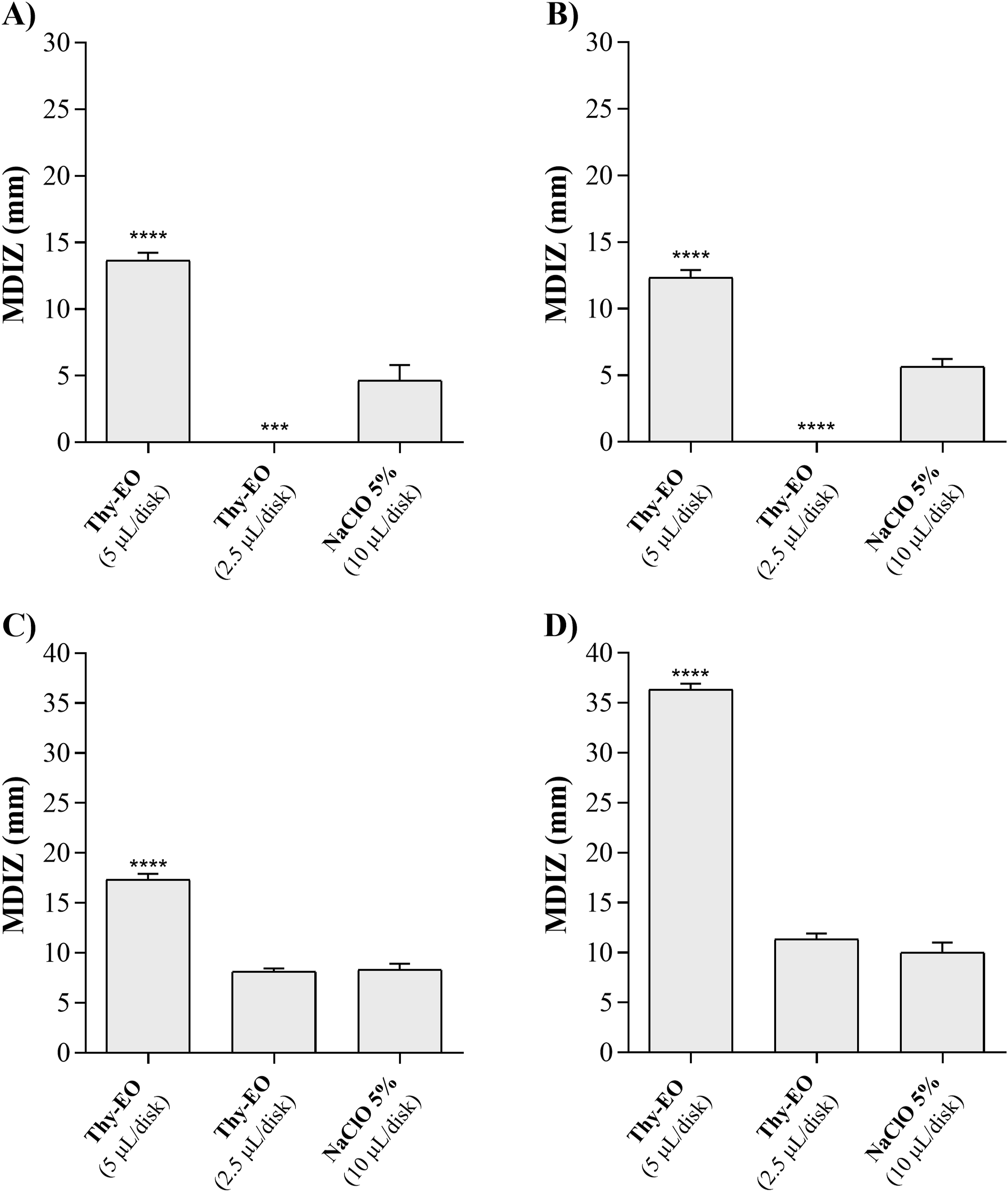
For Aspergillus, no significant inhibition was observed at the lower volume tested (2.5 µL), whereas at 5 µL, inhibition zones of 13.67 ± 0.47 mm for A. flavus and 12.33 ± 0.47 mm for A. parasiticus were recorded, exceeding the effect of the positive control (5% sodium hypochlorite, commonly used for indoor bioaerosol disinfection; Figure 2A and B; Figure S2). For Penicillium, no inhibition was observed at 2.5 µL, while 5 µL of Thy-EO produced zones of 17.33 ± 0.47 mm for P. expansum and 36.33 ± 0.47 mm for P. verrucosum, which was statistically significant compared to the positive control (Figure 2C and D; Figure S2).
Antifungal Activity of Thyme Essential Oil (Thymus vulgaris) Against Environmental Strains of Aspergillus flavus IZPS1, Aspergillus parasiticus IZPS2, Penicillium expansum IZPS3, and Penicillium verrucosum IZPS4
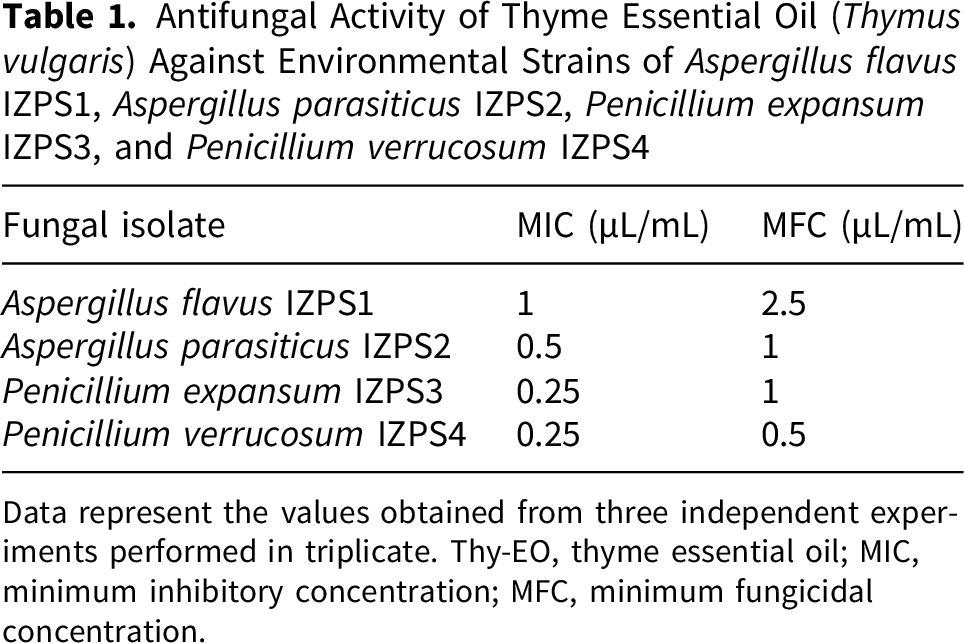
Data represent the values obtained from three independent experiments performed in triplicate. Thy-EO, thyme essential oil; MIC, minimum inhibitory concentration; MFC, minimum fungicidal concentration.
Representative images illustrating the fungistatic and fungicidal effects of Thy-EO on these environmental isolates after 48 h incubation are shown in Figure S3. These results indicate that Thy-EO exerts both fungistatic effects, limiting mycelial growth, and fungicidal activity against the tested environmental Aspergillus and Penicillium isolates.
3.2. Effect of Thyme Essential Oil on Fungal Fitness
The tested fungal strains, Aspergillus flavus, A. parasiticus, Penicillium expansum, and P. verrucosum, were exposed to increasing concentrations of thyme essential oil (Thy-EO; 0, 0.25, 0.5, 1, 2.5, and 5 µL/mL) and monitored over 72 hours to evaluate its impact on fungal fitness.
As shown in Figure 3, both Aspergillus strains were affected by Thy-EO treatment, displaying a concentration-dependent reduction in fungal fitness. A. parasiticus exhibited a significant decrease in fitness after 24 hours of exposure at 0.5 µL/mL, which was statistically significant compared to the untreated control (Figure 3B). In contrast, A. flavus showed a more gradual decline, indicating higher tolerance to the essential oil. Inhibitory effect of thyme essential oil (Thy-EO) on the fitness of the fungal strains Aspergillus flavus IZPS1 (A), Aspergillus parasiticus IZPS2 (B), Penicillium expansum IZPS3 (C), and Penicillium verrucosum IZPS4 (D). Microbial survival was monitored over 72 h in the absence and presence of increasing concentrations of Thy-EO (0.25–5 µL/mL). Results are expressed as log colony-forming units per mL of fungal inoculum (log CFU/mL). Experiments were performed in triplicate, and statistical significance was assessed by two-way ANOVA followed by Bonferroni’s post hoc test. Data are presented as mean ± SD of three independent experiments performed in technical triplicate (n = 9). Asterisks (p *< 0.05; **p < 0.01; ***p < 0.001; ***p < 0.0001) indicate statistically significant differences between treated and control cultures; ns, not significant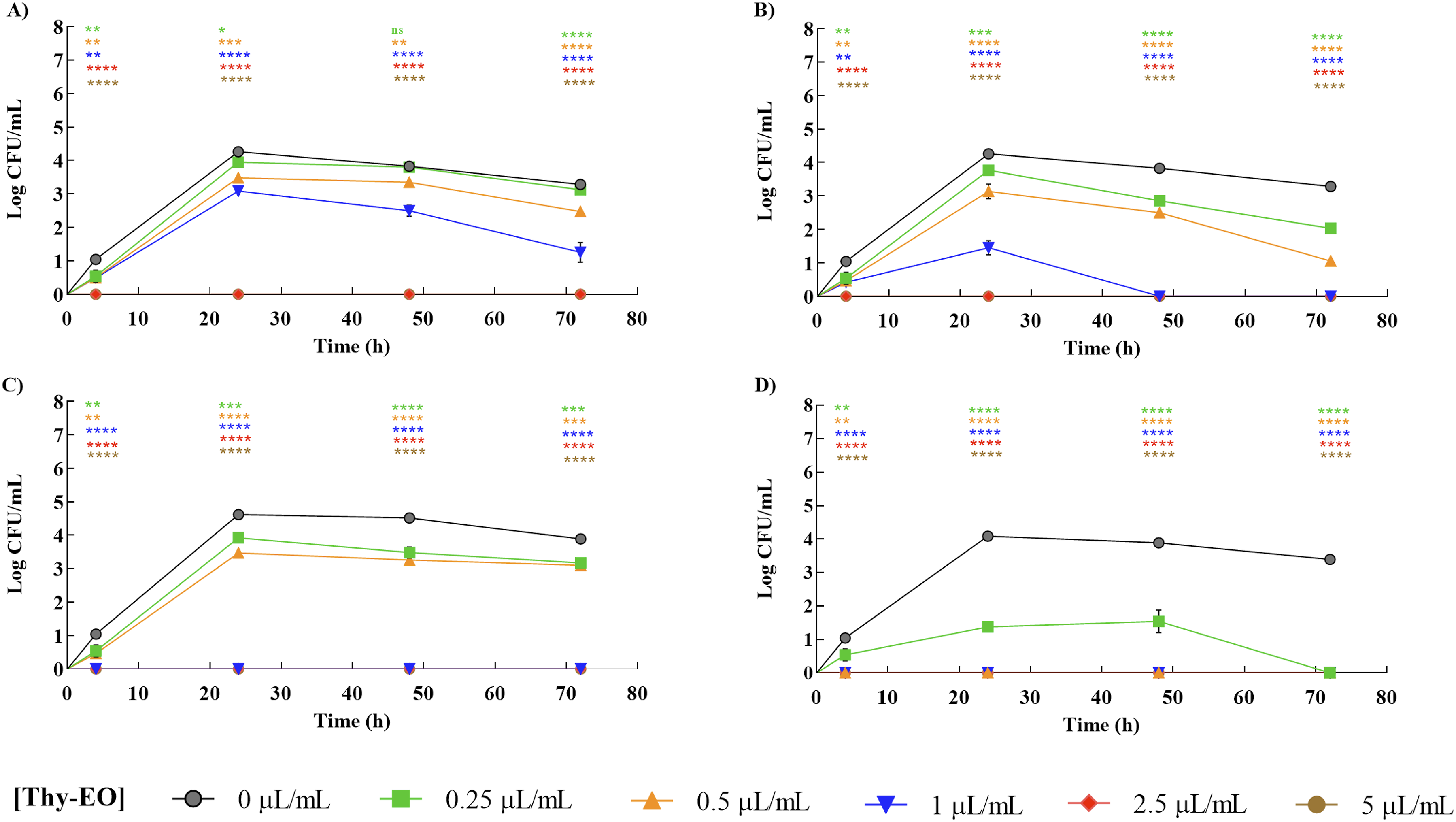
For the Penicillium strains, a similar concentration-dependent reduction in fungal fitness was observed (Figure 3C and D). However, the two species displayed different sensitivities to Thy-EO. P. verrucosum was the most susceptible, showing a marked decrease in fitness even at the lowest tested concentration (0.25 µL/mL) after 24 hours (Figure 3D). Conversely, P. expansum exhibited greater initial resistance, with a notable reduction in survival only at higher concentrations and prolonged exposure times (Figure 3C).
Overall, these results demonstrate that Thy-EO exerts a dose-dependent and sustained antifungal effect on both Aspergillus and Penicillium species. The more pronounced effect observed against A. parasiticus and P. verrucosum indicates species-specific differences in susceptibility, highlighting the greater resistance of A. flavus and P. expansum to Thy-EO.
3.3. Inhibition of Fungal Sporulation by Thyme Essential Oil
The in vitro assays evaluating the effect of thyme essential oil (Thy-EO) on fungal sporulation are shown in Figure 4. Sporulation was quantified as log spores/mL following exposure to increasing concentrations of Thy-EO (0, 0.25, 0.5, 1, 2.5, and 5 µL/mL). Inhibitory effect of thyme essential oil (Thy-EO) on sporulation of Aspergillus flavus IZPS1 (A), Aspergillus parasiticus IZPS2 (B), Penicillium expansum IZPS3 (C), and Penicillium verrucosum IZPS4 (D). Statistical significance compared with the control (untreated cultures) was evaluated by one-way ANOVA followed by Dunnett’s post hoc test. Data are presented as mean ± SD of three independent experiments performed in technical triplicate (n = 9). Asterisks (**p < 0.01; ***p < 0.001; ***p < 0.0001) indicate statistically significant differences between treated and control cultures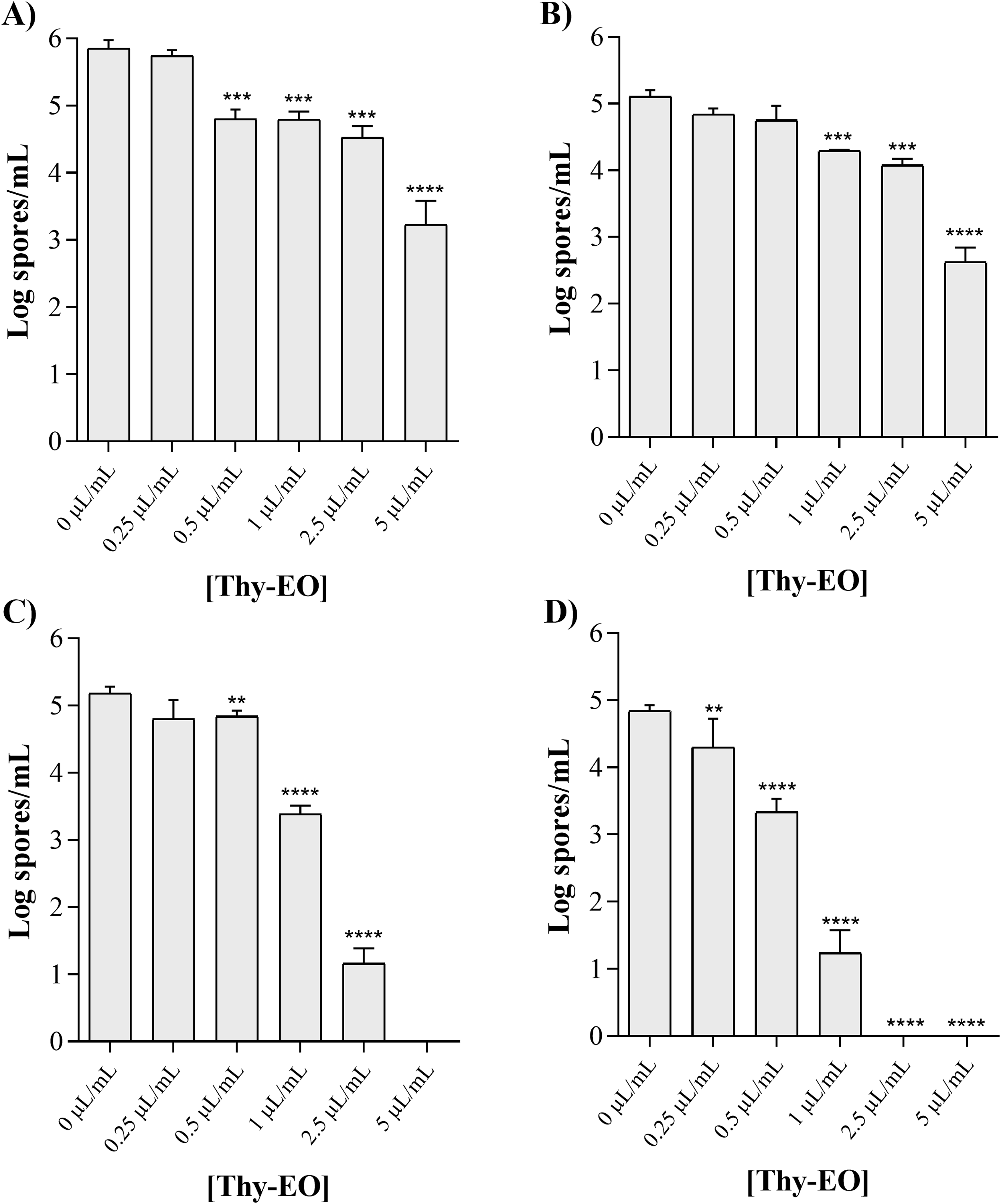
For Aspergillus flavus and A. parasiticus, Thy-EO did not markedly affect sporulation at the lower concentrations tested (0.25–2.5 µL/mL), with spore counts remaining in the range of approximately 4.5–6.0 log spores/mL (Figure 4A and B). A more pronounced reduction in sporulation was observed only at the highest concentration tested (5 µL/mL), corresponding to an approximate 50% decrease in spore production compared to the untreated control, with spore levels reduced to about 3.0 log spores/mL for A. flavus and 2.5 log spores/mL for A. parasiticus. Overall, these data indicate a threshold-like response, characterized by limited or no measurable effects at low Thy-EO concentrations and a marked inhibition occurring only at the highest dose.
In contrast, the results obtained for Penicillium expansum and P. verrucosum clearly demonstrated a dose-dependent inhibition of sporulation (Figure 4C and D). For both species, increasing Thy-EO concentrations resulted in a progressive and significant reduction in spore production, expressed as a marked decrease in log spores/mL. Complete suppression of sporulation was observed at Thy-EO concentrations of 5 µL/mL for P. expansum and at 2.5 and 5 µL/mL for P. verrucosum. Notably, P. verrucosum exhibited higher sensitivity to Thy-EO, showing a statistically significant reduction in sporulation already at relatively low concentrations (0.5 and 1 µL/mL), whereas P. expansum required higher doses to achieve a comparable inhibitory effect. These findings indicate that, although both species are susceptible to Thy-EO, P. verrucosum is overall more sensitive to its antisporulation activity.
Representative microscopic observations performed using a Bürker counting chamber are shown in Figure 5. In the absence of Thy-EO, high spore densities were observed for both Aspergillus species, while with increasing Thy-EO concentrations, a clear reduction in spore counts was evident. For the Penicillium species, an intermediate Thy-EO concentration (1 µL/mL) already resulted in a strong decrease in sporulation, while at the highest concentration tested (5 µL/mL) spores were completely absent. Light microscopy images showing sporulation of Aspergillus flavus IZPS1, Aspergillus parasiticus IZPS2, Penicillium expansum IZPS3, and Penicillium verrucosum IZPS4 in the absence and presence of thyme essential oil (Thy-EO). Spore suspensions obtained from untreated cultures (control) and from cultures treated with an intermediate concentration (1 µL/mL) and a higher concentration (5 µL/mL) of Thy-EO are shown
Collectively, these results confirm that Thy-EO effectively inhibits fungal sporulation, exerting a strong antisporulation effect against Aspergillus spp. and a pronounced sporicidal activity against Penicillium spp., supporting its potential application as a natural antifungal agent for controlling spore-mediated contamination in agri-food and indoor environments.
4. Discussion
The increasing incidence of fungal contamination in indoor environments and agri-food systems, together with growing concerns regarding antifungal resistance and chemical residues, has intensified the search for effective and sustainable alternatives to conventional fungicides. The relevance of fungal contamination is further supported by the ability of several species to produce toxic and potentially carcinogenic mycotoxins, contaminate food and beverages, and contribute to respiratory disorders, particularly in susceptible or immunocompromised individuals. In this context, essential oils represent a promising class of natural compounds due to their broad-spectrum antimicrobial activity and volatility, although their safety and efficacy must be carefully assessed according to the specific formulation, dose, and intended application. However, most existing studies primarily focus on inhibition of mycelial growth, while comparatively limited attention has been devoted to fungal sporulation and spore viability, despite their central role in environmental dissemination and exposure dynamics. In this framework, the present study provides an integrated evaluation of the antifungal, anti-sporulation, and sporicidal activities of thyme essential oil (Thy-EO) against environmental isolates of Aspergillus and Penicillium of relevance for human health and food safety. No emulsifiers or organic solvents were used in the present study, in order to avoid potential interference with the antimicrobial activity of the essential oil and to support a more sustainable experimental design. This approach is consistent with the growing interest in solvent-free or minimal-interference methodologies in essential oil research. It should be noted that the agar diffusion and disk volatilization assays were designed as preliminary comparative screenings under direct-contact and vapor-phase exposure conditions, whereas the dose-dependent response was further investigated using the liquid assay, in which a broader concentration range was tested.
Thy-EO exerted a strong inhibitory activity against Aspergillus spp., with antifungal effects comparable to those of the reference antifungal drug tioconazole and, in some cases, significantly higher. Tioconazole, a reference azole antifungal, was included as a clinically relevant benchmark to contextualize the in vitro antifungal performance of Thy-EO with respect to a standard pharmacological agent. These findings are particularly noteworthy, suggesting that a natural product widely used in food and phytotherapeutic contexts may represent a valid alternative or complementary approach to synthetic antifungals. The strong antifungal activity observed against Aspergillus spp. is consistent with previous studies attributing the efficacy of thyme essential oil primarily to its high content of thymol and carvacrol. Previous studies have suggested that these phenolic compounds may disrupt fungal cell membrane integrity through alterations in ergosterol biosynthesis, increased membrane permeability, and impairment of essential cellular functions. In particular, Khatoon et al. 35 reported minimum inhibitory concentration (MIC) values of 31 µL/mL and minimum fungicidal concentration (MFC) values of 125–250 µL/mL for thyme essential oil against aflatoxigenic strains of A. flavus, which are comparable to those obtained in the present study. Previous studies have proposed that thymol may contribute to ergosterol depletion, membrane destabilization, and leakage of intracellular components, whereas carvacrol has been reported to affect H+-ATPase activity, potentially compromising proton gradients and nutrient uptake. However, these mechanisms were not investigated in the present study.35,36
The comparison between agar diffusion and disk volatilization assays revealed that, at equal oil volumes, Thy-EO exerted a stronger inhibitory effect against Aspergillus spp. in the vapor phase. This observation is consistent with literature reports indicating that the antifungal activity of essential oils in the gaseous phase may be 8- to 128-fold higher than in the liquid phase. 37 In particular, Inouye et al. 38 demonstrated that thyme essential oil displayed more than a 100-fold higher efficacy in vapor form compared to direct contact against Aspergillus fumigatus. Similarly, Tian et al. 37 showed that complete inhibition of A. flavus mycelial growth was achieved with 20 µg/mL of Thy-EO in the vapor phase, whereas a 20-fold higher concentration was required in liquid medium.
The antifungal efficacy of Thy-EO against Penicillium expansum and P. verrucosum was also remarkable, with inhibition zones comparable to or significantly higher than those of the selected controls. In disk volatilization assays, Thy-EO further outperformed 5% sodium hypochlorite, which was selected as a positive control for this assay because of its widespread use as a disinfectant in environmental sanitation and indoor surface decontamination practices. This comparison supports the potential of Thy-EO volatile compounds as a promising alternative or complementary strategy for controlling fungal contamination under vapor-phase exposure conditions. The comparison between agar diffusion and vapor-phase assays showed that inhibition of P. verrucosum was nearly comparable between the two exposure modes, whereas P. expansum exhibited slightly higher sensitivity in the liquid phase. This pattern aligns with previous reports indicating a pronounced susceptibility of Penicillium spp. to the vapor phase of thyme essential oil, even at low doses. 39 The enhanced efficacy of the gaseous phase is likely attributable to the higher bioavailability of volatile compounds such as thymol and p-cymene, whose interaction with agar matrices may reduce their effective concentration in solid media. Microscopic observations and representative images (Figure S3) further confirmed both fungistatic and fungicidal effects of Thy-EO against P. expansum and P. verrucosum at concentrations corresponding to MIC and MFC values. These findings are consistent with previous studies reporting strong antifungal activity of thyme essential oil against Penicillium spp. For instance, Kręcidło et al. 40 demonstrated that thyme oil exhibited the highest antifungal activity among tested essential oils against environmental Penicillium isolates, with MIC and MFC values of 1 µL/mL, while P. expansum was identified as the most resistant species but still fully inhibited under the same conditions.
Fitness assays revealed a clear dose- and time-dependent antifungal effect of Thy-EO on both Aspergillus and Penicillium species. Among Aspergillus, A. parasiticus exhibited a more pronounced reduction in fitness compared to A. flavus, suggesting species-specific differences in susceptibility. The biological factors underlying these differences were not investigated in the present study, although variations in membrane composition, stress responses, or detoxification pathways could represent possible explanations. Similarly, Thy-EO markedly affected the fitness of Penicillium spp., with P. verrucosum showing a faster and stronger decline in survival compared to P. expansum, confirming its overall higher sensitivity. These findings indicate that Thy-EO exerts sustained antifungal pressure that extends beyond growth inhibition, progressively impairing fungal viability over time. From a mechanistic perspective, thymol may affect ergosterol biosynthesis and membrane-associated functions. However, these mechanisms were not directly evaluated in the present work. Previous investigations have reported that thymol may interfere with ergosterol biosynthesis and that thyme essential oil constituents may affect H+-ATPase activity, potentially altering membrane homeostasis, proton gradients, and nutrient transport.35,36 These multi-target effects likely contribute to the persistent reduction in fungal fitness observed in both genera. Importantly, these findings indicate that the antifungal activity of Thy-EO extends beyond growth inhibition, exerting sustained pressure on fungal viability and biological performance over time. This reinforces the concept of a dual-action antifungal strategy targeting both vegetative growth and long-term fungal fitness.
Beyond the effects of Thy-EO on fungal growth and fitness, a key novel aspect of the present study lies in the systematic evaluation of sporulation and spore viability, endpoints that are rarely considered in previous investigations on thyme essential oil despite their ecological and epidemiological relevance. Thy-EO was shown to modulate sporulation, a key determinant of fungal dissemination and environmental persistence. The in vitro sporulation assays revealed distinct genus-specific responses. For Aspergillus spp., Thy-EO exerted a limited antisporulation effect, with a significant reduction in conidial production observed only at the highest concentration tested. This finding is consistent with previous studies reporting variable effects of thyme essential oil on Aspergillus sporulation. In particular, Tian et al. 41 showed that thyme oils rich in p-cymene inhibited mycelial growth of A. flavus without significantly affecting conidiogenesis, as confirmed by microscopic analyses. The chemical composition of the Thy-EO used in this study—characterized by a phenolic fraction of approximately 48.6% (thymol 43.5% and carvacrol 5.11%) combined with a relatively high p-cymene content (31.65%)— may represent one possible explanation for the limited antisporulation effect observed for Aspergillus spp.; however, this hypothesis was not directly tested. In contrast, Thy-EO exhibited a strong and dose-dependent inhibition of sporulation in both P. expansum and P. verrucosum, culminating in complete suppression of spore production at higher concentrations. P. verrucosum proved to be particularly sensitive, showing significant reductions in sporulation even at relatively low Thy-EO concentrations. This behavior is consistent with the findings of Özcakmak et al., 42 who reported a 50–60% reduction in sporulation of P. verrucosum within 48 h and fungal lethality of approximately 78–81% following treatment with essential oils, including thyme. Similarly, more recent evidence indicates that exposure to essential oil vapours effectively suppresses sporulation in Penicillium spp., confirming that the gaseous phase directly interferes with conidiation processes. In particular, Hlebová et al. 43 demonstrated that the vapor phase of thyme essential oil significantly inhibited both mycelial growth and sporulation of Penicillium commune at low concentrations, highlighting the high sensitivity of conidiation processes to volatile antifungal compounds. At the molecular level, recent evidence have suggested that p-cymene may affect membrane integrity and membrane potential in Penicillium spp., potentially facilitating the activity of thymol and carvacrol, while γ-terpinene has been proposed to interfere with nutrient transport and cellular homeostasis. 44 Collectively, these literature-based mechanisms may provide possible explanations for the pronounced sporulation inhibition observed in Penicillium spp.; however, their contribution was not experimentally assessed in the present study. Taken together, these results support the notion that Thy-EO does not merely inhibit fungal growth, but can also interfere with conidiation processes and spore viability, thereby potentially limiting environmental dissemination and contamination at its source.
The present study has some limitations that should be acknowledged.
First, the molecular mechanisms underlying the antifungal and antisporulation activities of Thy-EO were not directly investigated. Therefore, the proposed involvement of membrane disruption, ergosterol depletion, alterations in membrane permeability, and H+-ATPase inhibition remains based on evidence reported in previous studies rather than on direct experimental confirmation. Nevertheless, the study provides robust phenotypic evidence of antifungal, fungicidal, and antisporulation effects across multiple complementary assays, establishing a solid biological basis for future mechanistic investigations.
Second, the antifungal assessment was performed on a limited number of fungal isolates. Although the four selected isolates belong to species of high relevance for food spoilage, mycotoxin contamination, and indoor environmental exposure, this experimental design does not allow the full variability of filamentous fungi to be captured, particularly at the intra-species level. Indeed, different strains belonging to the same fungal species may show different susceptibility profiles depending on their genetic background, physiological state, origin, and previous environmental adaptation. Therefore, the present findings should be interpreted as preliminary evidence obtained from representative target fungi, rather than as a comprehensive definition of the antifungal spectrum of Thy-EO. This approach is consistent with preliminary antifungal studies using one or a few representative fungal strains.45,46 Future studies should therefore include a larger and more diverse panel of isolates, including multiple strains per species and isolates from different environmental or food-related sources, in order to confirm the robustness and generalizability of the observed antifungal effects.
Third, the study was conducted using a commercially available thyme essential oil with a manufacturer-reported chemical composition. Although no independent in-house GC-MS analysis was performed, the essential oil was accompanied by physicochemical documentation provided by the manufacturer, including the ATR–FTIR spectrum. Since the phytochemical profile of essential oils can vary according to geographical origin, cultivation conditions, extraction methods, and storage parameters, some variability in antifungal efficacy may be expected among different thyme oil preparations. At the same time, the use of a well-characterized commercial product enhances the reproducibility and potential translational relevance of the findings, as such formulations are readily available for practical applications and further validation studies.
Furthermore, mycotoxin production was not evaluated. Since Aspergillus and Penicillium species may continue to produce toxins under certain stress conditions, future studies should determine whether the observed inhibition of fungal growth and sporulation is accompanied by a reduction in mycotoxin biosynthesis. Despite this limitation, the demonstrated suppression of fungal fitness and reproductive capacity constitutes an encouraging first step toward the development of integrated strategies aimed at reducing both fungal contamination and its associated risks.
Finally, all experiments were performed under controlled in vitro conditions. Such models cannot fully reproduce the complexity of real food systems or indoor environments, where environmental variables, substrate composition, and microbial interactions may influence antifungal performance. However, in vitro investigations represent an essential preliminary stage for identifying promising biocontrol candidates under standardized and reproducible conditions. In this regard, the consistent activity of Thy-EO across different experimental approaches, including direct contact assays, vapor-phase exposure, fitness analyses, and sporulation assays, strengthens the evidence supporting its potential application and justifies further validation in more complex real-world settings.
5. Conclusions
Overall, this study provides a comprehensive in vitro evaluation of the antifungal potential of T. vulgaris essential oil, demonstrating its combined effects on fungal growth, fitness, sporulation, and spore viability. This multifaceted antifungal activity represents an advancement over previous studies primarily focused on vegetative growth inhibition, highlighting the ability of Thy-EO to also interfere with spore-mediated dissemination.
From an applicative perspective, these findings suggest that Thy-EO may represent a promising natural candidate for the development of alternative antifungal strategies aimed at improving indoor air quality and reducing surface contamination by Aspergillus species in controlled environments. However, these implications remain preliminary, as the evidence is based exclusively on in vitro assays and requires further validation under more realistic conditions. Likewise, the observed activity against Penicillium spp. indicates potential relevance in agri-food applications, where these fungi are major contributors to food spoilage and mycotoxin contamination. Nevertheless, translation into practical applications will require additional studies in complex food matrices, together with assessments of stability, safety, and efficacy under real-use conditions.
In conclusion, these results expand current knowledge on the bioactivity of thyme essential oil and support its potential role in reducing environmental contamination, food spoilage, and associated exposure risks. Further investigations are warranted to confirm these findings and to evaluate their applicability in real-world systems.
Supplemental Material
Supplemental Material -Bio-Control of Food- and Indoor-Associated Filamentous Fungi: Fungicidal and Antisporulation Action of Thymus vulgaris Essential Oil
Supplemental Material for Bio-Control of Food- and Indoor-Associated Filamentous Fungi: Fungicidal and Antisporulation Action of Thymus vulgaris Essential Oil by Giuseppina Forgione, Manuela Siciliano, Elena Scaglione, Giuseppe Mantova, Paola Salvatore, Caterina Pagliarulo, and Daniela Sateriale in Natural Product Communications.
Footnotes
Acknowledgments
The authors would like to thank the Istituto Zooprofilattico Sperimentale (IZPS, Portici, Naples, Italy) for providing the fungal isolates.
Author Contributions
Conceptualization, D.S. and C.P.; methodology, D.S., G.F., M.S; E.S., G.M.; data curation, D.S. and C.P.; writing—original draft preparation, D.S., C.P. and G.F.; writing—review and editing, D.S., C.P. and G.F.; visualization, P.S.; supervision, C.P. and P.S.; project administration, C.P.; funding acquisition, C.P. and P.S. All authors have read and agreed to the published version of the manuscript.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by Fondi Ricerca di Ateneo (FRA) 2024 for C.P. and EU funding within the MUR PNRR Extended Partnership initiative on Emerging Infectious Disease (Project no. PE00000007, INF-ACT) for P.S.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.