
Abstract
Background
Psoriasis is a chronic inflammatory skin disease. Quercetin, a natural flavonoid, shows therapeutic potential but lacks comprehensive evidence synthesis.
Objective
To summarize pharmacological mechanisms, efficacy, and innovative formulations of quercetin and its derivatives (rutin, taxifolin) in psoriasis treatment.
Methods
A systematic search was conducted across PubMed, Embase, Web of Science, and Cochrane Library (inception to February 2026). Studies on quercetin, rutin, or taxifolin in psoriasis models or patients were included. Data were synthesized descriptively.
Results
Twenty-two studies were included. Quercetin exerts anti-psoriatic effects by inhibiting NF-κB and JAK/STAT pathways, activating Nrf2 antioxidant defense, regulating PI3K/AKT/Glut1 and Notch signaling to suppress keratinocyte proliferation, modulating AMPK-mediated immunometabolic reprogramming, and targeting TOP2A, AhR, and cell cycle regulators. Rutin acts via JAK2/STAT3 and Keap1/Nrf2 pathways. Taxifolin regulates T-cell differentiation through Notch1/JAK2/STAT3 and cPLA2/PPAR-γ pathways. Innovative formulations (nanoemulsions, liposomes) significantly enhance bioavailability, achieving up to 6-fold increased skin permeation and 56.7% reduction in epidermal thickness, with efficacy comparable or superior to conventional drugs and better safety profiles.
Conclusion
Quercetin and its derivatives exert multi-target therapeutic effects through coordinated regulation of inflammatory, oxidative, proliferative, and metabolic pathways. Advanced delivery systems enhance clinical potential, supporting their development as alternative or adjunctive therapies.
Introduction
Psoriasis is a chronic, recurrent, immune-mediated inflammatory skin disease with a global prevalence of approximately 2%-3%, significantly impacting patients’ quality of life and psychological well-being.1,2 Its typical clinical features are well-demarcated red plaques covered with silvery scales, often accompanied by itching and pain. 3 The pathophysiological mechanisms of psoriasis are complex and are currently understood to involve interactions among genetic susceptibility, environmental factors, and immune system dysregulation. 4 Among these, the abnormal activation of the IL-23/Th17 axis, predominantly driven by Th17 cells, leading to hyperproliferation and abnormal differentiation of keratinocytes and a cascade release of inflammatory factors, is considered the core element in the pathogenesis of psoriasis.5,6 Furthermore, oxidative stress plays a crucial role in the development and progression of psoriasis. Excessive production of reactive oxygen species can further activate pro-inflammatory pathways such as NF-κB, forming a vicious cycle that exacerbates local inflammatory responses. 7
Current treatment strategies for psoriasis are categorized based on disease severity and include topical therapies, phototherapy, systemic immunosuppressants, and biologics. Although biologics have demonstrated remarkable efficacy in severe psoriasis, their high cost, potential risk of infections, and concerns regarding long-term safety limit their clinical application in some patients.8,9 Therefore, identifying safe, effective, and economical natural compounds as adjunctive or alternative therapeutic options has become a focal point in psoriasis research.
Quercetin is a flavonoid widely found in vegetables, fruits, and various medicinal herbs, possessing significant anti-inflammatory, antioxidant, immunomodulatory, and antiproliferative activities. 10 Its natural derivatives, such as rutin (quercetin-3-O-rutinoside) and isoquercitrin, also exhibit similar biological activities both in vitro and in vivo. 11 In recent years, a growing body of preclinical studies and some clinical observations have indicated that quercetin and its derivatives can intervene in the pathological process of psoriasis through multiple targets, including inhibiting the NF-κB signaling pathway, downregulating inflammatory factors such as IL-17 and TNF-α, activating the Nrf2 antioxidant pathway, and inducing keratinocyte apoptosis.12,13 However, current research on quercetin and its derivatives for psoriasis treatment is scattered, and there is a lack of an integrated analysis of their efficacy and mechanisms of action.
Several previous reviews have discussed the anti-inflammatory or antioxidant properties of flavonoids in skin diseases. However, to the best of our knowledge, no review has specifically and comprehensively focused on: (1) the comparative pharmacological mechanisms of quercetin versus its derivatives rutin and taxifolin in psoriasis; (2) the integration of molecular mechanisms (NF-κB, JAK/STAT, Nrf2, PI3K/AKT/Glut1, AMPK) into a cohesive multi-target network; and (3) the emerging role of innovative nanoformulations (nanocapsules, nanoemulsions, cyclodextrin-modified liposomes) that overcome the bioavailability limitations of quercetin. The present review fills these gaps by systematically synthesizing 22 studies to provide an evidence-based foundation for the clinical translation of quercetin-based therapies in psoriasis.
This systematic review aims to systematically summarize the available evidence on the use of quercetin and its derivatives in the treatment of psoriasis, with a focus on elucidating their pharmacological mechanisms and evaluating their efficacy and safety in both preclinical and clinical studies. The goal is to provide an evidence-based foundation for adjunctive therapeutic strategies for psoriasis and to identify directions for future research, while acknowledging the preliminary nature of the current evidence base due to incomplete reporting in several included studies.
Methods
Search Strategy
This systematic review was designed and conducted in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA 2020) statement. 14 and was registered in PROSPERO (registration number: CRD420261428453) retrospectively in 2026/06/18. The completed PRISMA checklist is provided as Supplementary File 1. A systematic search was performed in the following databases: PubMed, Embase, Web of Science, and Cochrane Library, search period will cover from the inception of each database to February 2026. The search strategy combined Medical Subject Headings (MeSH)/Emtree terms with free-text keywords. Chinese search terms included: quercetin, rutin, isoquercitrin, flavonoids, psoriasis, and plaque psoriasis. English search terms included: “Quercetin,” “Rutin,” “Isoquercitrin,” “Flavonoids,” “Psoriasis,” and “Plaque Psoriasis.” Additionally, the reference lists of all included studies weremanually screened to identify potentially eligible studies not captured by the electronic search. The complete search strategies for all databases, including specific keywords, filters, and limits, are provided in Supplementary File 2.
Inclusion and Exclusion Criteria
The inclusion criteria for this review are as follows: (1) Study types: Clinical studies including randomized controlled trials, controlled clinical trials, cohort studies, and case-control studies will be included. In vitro cell experiments and animal model studies will also be included to comprehensively evaluate the mechanisms of action and efficacy of quercetin and its derivatives. (2) Participants/Subjects: This includes patients with a clinically confirmed diagnosis of psoriasis, animal models of psoriasis (e.g., imiquimod-induced mouse models), and psoriasis-related cell models (e.g., HaCaT keratinocytes). (3) Interventions: The intervention must involve quercetin, quercetin derivatives (such as rutin, isoquercitrin), or standardized extracts rich in quercetin. Combination therapy of quercetin or its derivatives with other drugs will be permitted. (4) Comparators: Comparators may include placebo, no treatment (blank control), conventional therapy, or a positive control drug. (5) Outcome measures: Outcomes of interest include, but are not limited to, the Psoriasis Area and Severity Index (PASI), levels of inflammatory factors (e.g., IL-17, TNF-α, IL-23), oxidative stress markers (e.g., ROS, MDA, SOD, Nrf2 expression), and indicators related to keratinocyte proliferation and apoptosis.
The exclusion criteria are: (1) Non-original research articles such as reviews, conference abstracts, comments, case reports, and editorials; (2) Studies not published in English or Chinese due to the authors’ linguistic limitations; however, we acknowledge that restricting to these two languages may introduce language bias, as potentially relevant studies in other languages were not included; (3) Duplicate publications; (4) Studies for which the full text cannot be obtained or data are incomplete.
Data Integrity Consideration
Characteristics of Included Studies
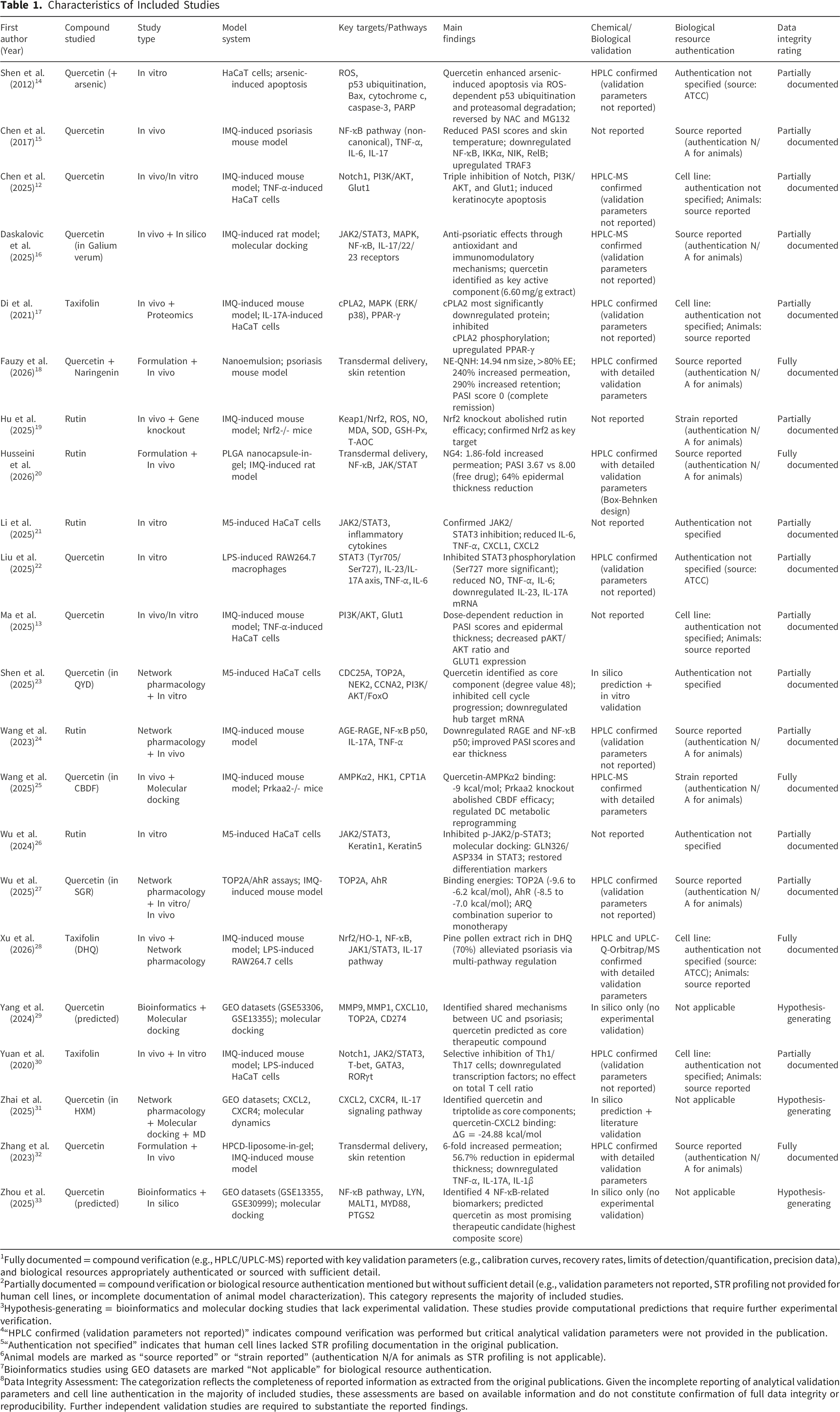
1Fully documented = compound verification (e.g., HPLC/UPLC-MS) reported with key validation parameters (e.g., calibration curves, recovery rates, limits of detection/quantification, precision data), and biological resources appropriately authenticated or sourced with sufficient detail.
2Partially documented = compound verification or biological resource authentication mentioned but without sufficient detail (e.g., validation parameters not reported, STR profiling not provided for human cell lines, or incomplete documentation of animal model characterization). This category represents the majority of included studies.
3Hypothesis-generating = bioinformatics and molecular docking studies that lack experimental validation. These studies provide computational predictions that require further experimental verification.
4“HPLC confirmed (validation parameters not reported)” indicates compound verification was performed but critical analytical validation parameters were not provided in the publication.
5“Authentication not specified” indicates that human cell lines lacked STR profiling documentation in the original publication.
6Animal models are marked as “source reported” or “strain reported” (authentication N/A for animals as STR profiling is not applicable).
7Bioinformatics studies using GEO datasets are marked “Not applicable” for biological resource authentication.
8Data Integrity Assessment: The categorization reflects the completeness of reported information as extracted from the original publications. Given the incomplete reporting of analytical validation parameters and cell line authentication in the majority of included studies, these assessments are based on available information and do not constitute confirmation of full data integrity or reproducibility. Further independent validation studies are required to substantiate the reported findings.
Study Selection and Data Extraction
Two reviewers independently screened the literature. Initially, titles and abstracts were screened to exclude obviously irrelevant studies. Subsequently, the full texts of the remaining studies were reviewed to determine final inclusion. Any disagreements were resolved through discussion or by consulting a third reviewer. Data extraction was performed independently by two reviewers using a standardized data extraction form. The extracted information included: first author, publication year, country, study type, sample size (number of participants/animals/cells), intervention details (specific compound, dose, route of administration, duration), comparator details, outcome measures, main results, and relevant mechanistic findings.
Quality Assessment
The methodological quality of the included studies was assessed using appropriate tools based on the study design. For randomized controlled trials, the Cochrane Risk of Bias tool (RoB 2.0) will be used. For animal studies, the SYRCLE Risk of Bias tool will be applied. As there is no universally accepted standardized tool for assessing in vitro studies, their quality will be evaluated descriptively based on the rigor of the experimental design (e.g., presence of replication, appropriate controls). For randomized controlled trials, the fragility index—a metric reflecting the number of event changes needed to reverse statistical significance—was examined where reported. However, none of the included RCTs explicitly reported the fragility index, which represents a limitation in assessing the robustness of their statistical conclusions.
Data Analysis
Given the anticipated heterogeneity among the included studies (involving clinical research, animal experiments, and cell culture studies) and significant clinical/methodological diversity (e.g., differences in subjects, intervention doses, administration routes, and outcome measures), a descriptive synthesis of the findings was planned. If a sufficient number of clinically and methodologically homogeneous randomized controlled trials are identified, a meta-analysis wasconsidered using software such as RevMan; otherwise, the results will be summarized systematically in a narrative format.
Results
Literature Search Results
All 22 articles were finally included for systematic analysis, including 14 studies on quercetin (Shen et al., 2012
15
; Chen et al., 2017
16
; Chen et al., 2025
12
; Daskalovic et al., 2025
18
; Fauzy et al., 2026
22
; Liu et al., 2025
23
; Ma et al., 2025
13
; Shen et al., 2025
25
; Wang et al., 2025
27
; Wu et al., 2025
29
; Yang et al., 2024
31
; Zhai et al., 2025
32
; Zhang et al., 2023
33
; Zhou et al., 2025
19
), 5 studies on rutin (Hu et al., 2025
20
; Husseini et al., 2026
21
; Li et al., 2025
24
; Wang et al., 2023
26
; Wu et al., 2024
17
), and 3 studies on taxifolin (dihydroquercetin) (Di et al., 2021
28
; Xu et al., 2026
30
; Yuan et al., 2020
34
). The PRISMA flow chart can be seen in the Figure 1. PRISMA Flow chart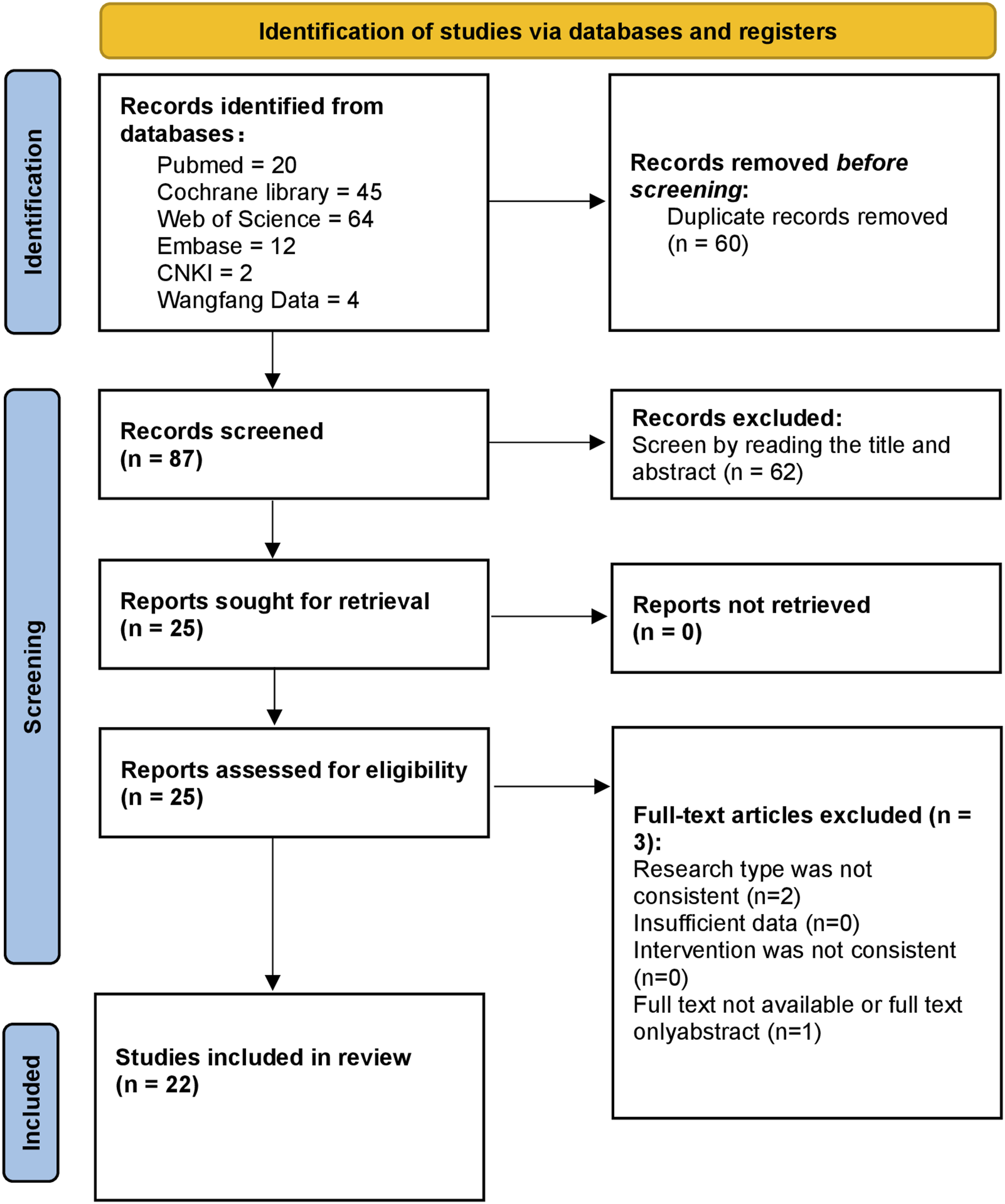
The details of included studies can be seen in Table 1. The data integrity assessments presented in this table are based on the information reported in the original publications. As most studies did not provide complete analytical validation parameters or cell line authentication documentation, these assessments should be interpreted as indicative of reporting completeness rather than definitive confirmation of data integrity.
Pharmacological Mechanisms of Quercetin in Psoriasis Treatment
Anti-Inflammatory Mechanisms
NF-κB Signaling Pathway
Inhibition of the NF-κB pathway by quercetin represents the most consistently reported anti-inflammatory mechanism across the literature. Chen et al. (2017) 16 first reported that quercetin (30, 60, and 120 mg/kg orally) significantly reduced Psoriasis Area and Severity Index (PASI) scores and lesional skin temperature while improving histopathological alterations in an imiquimod (IMQ)-induced psoriasis mouse model. Mechanistic studies revealed that quercetin downregulated key proteins of the non-canonical NF-κB pathway, including NF-κB, IKKα, NIK, and RelB, while upregulating the negative regulator TRAF3. Concurrently, quercetin significantly reduced serum levels of pro-inflammatory cytokines TNF-α, IL-6, and IL-17.
Liu et al. (2025) 23 further demonstrated that quercetin, as the active component of Centella asiatica, inhibits the IL-23/IL-17A inflammatory axis by mediating STAT3 phosphorylation. In an LPS-stimulated RAW264.7 macrophage inflammation model, quercetin (15-60 μmol/L) dose-dependently reduced NO production, TNF-α and IL-6 secretion, and downregulated mRNA expression of IL-23, IL-17A, TNF-α, and IL-6. Western blot analysis showed that quercetin significantly inhibited STAT3 phosphorylation at both sites (Tyr705 and Ser727), with more pronounced inhibition at Ser727.
Zhou et al. (2025) 19 employed a bioinformatics approach to identify NF-κB-related biomarkers in psoriasis. Through LASSO regression analysis of GEO datasets, four characteristic genes—LYN, MALT1, MYD88, and PTGS2—were identified with significant diagnostic value (AUC: 0.975-1.0). Notably, drug prediction using the DSigDB database revealed that quercetin scored highest among all tested compounds, emerging as the most promising therapeutic candidate targeting these NF-κB pathway-related genes, further supporting experimental evidence of quercetin’s NF-κB inhibitory effects.
JAK/STAT Signaling Pathway
Wu et al. (2024) 29 demonstrated in an M5 cytokine mixture (IL-17A, IL-22, TNF-α, IL-1α, oncostatin M)-induced HaCaT cell psoriasis model that rutin (10-30 μM) significantly inhibited activation of the JAK2/STAT3 signaling pathway. Rutin treatment reduced p-JAK2 and p-STAT3 protein levels, and molecular docking predicted that rutin binds to GLN326 and ASP334 residues of STAT3, potentially interfering with STAT3-DNA interactions to inhibit transcriptional activity. Additionally, rutin reversed M5-induced downregulation of keratinocyte differentiation markers Keratin1 and Keratin5.
AGE-RAGE Signaling Pathway
Wang et al. (2023) 26 identified through network pharmacology and experimental validation that rutin exerts anti-inflammatory effects by downregulating the AGE-RAGE signaling pathway. In an IMQ-induced psoriasis mouse model, topical rutin (1.5% cream) treatment significantly improved PASI scores, ear thickness, and histopathological alterations. Western blot and immunohistochemistry confirmed that rutin downregulated RAGE and NF-κB p50 expression while reducing mRNA levels of IL-17A and TNF-α.
Antioxidant Mechanisms
Hu et al. (2025) 20 further confirmed in an IMQ-induced psoriasis mouse model that rutin inhibits oxidative stress injury and inflammatory response through the Keap1/Nrf2 signaling pathway. Rutin treatment significantly reduced ROS, NO, and MDA content and increased T-AOC, SOD, and GSH-Px activities. Importantly, Nrf2 knockout mice exhibited more severe psoriatic symptoms, and the therapeutic effect of rutin was diminished, confirming Nrf2 as a key target for rutin’s antioxidant effects.
Regulation of Cell Proliferation and Apoptosis
PI3K/AKT/GLUT1 Pathway and Notch Signaling Crosstalk
Regulation of the PI3K/AKT/GLUT1 signaling pathway by quercetin to inhibit keratinocyte hyperproliferation is a consistent finding across multiple studies. Chen et al. (2025) 12 first reported that quercetin induces keratinocyte apoptosis through triple inhibition of Notch, PI3K/AKT signaling, and Glut1 in psoriasis treatment. Clinical sample analysis revealed that Notch1, AKT, and Glut1 were highly expressed in psoriatic lesions and CD4+ T lymphocytes, positively correlated with epidermal thickness. In an IMQ-induced psoriasis mouse model, quercetin (30-120 mg/kg orally) significantly reduced PASI scores, epidermal thickness, and Ki67-positive cell numbers while downregulating Notch1, AKT, and Glut1 expression. In TNF-α-induced HaCaT cells, quercetin (IC50=62.48 μM at 48h) significantly induced apoptosis and inhibited Notch1, AKT, and Glut1 expression. Experiments with the Notch inhibitor DAPT confirmed that downregulation of PI3K/AKT and Glut1 was partially dependent on Notch inhibition.
Ma et al. (2025) 13 further confirmed that quercetin treats psoriasis by inhibiting the PI3K/AKT/GLUT1 signaling pathway. In an IMQ-induced mouse model, quercetin (30-120 mg/kg orally) dose-dependently reduced PASI scores, epidermal thickness, and Akt and Glut1 mRNA expression. In TNF-α-induced HaCaT cells, quercetin (IC50=73.28 μM at 48h) significantly induced apoptosis and reduced the pAKT/AKT ratio and GLUT1 protein expression. Experiments with the PI3K inhibitor LY294002 confirmed that quercetin downregulates GLUT1 expression through PI3K/AKT pathway inhibition.
Shen et al. (2012) 15 investigated the synergistic effect of quercetin and arsenic trioxide (As3+) in HaCaT keratinocytes, providing a rationale for improving arsenic-based psoriasis therapy. Quercetin (50-100 μM) significantly enhanced As3+-induced cytotoxicity and apoptosis, as evidenced by increased DNA fragmentation, caspase-3/PARP cleavage, and loss of mitochondrial membrane potential, while rutin showed no such effect. Mechanistically, this effect was mediated by ROS-dependent p53 ubiquitination and proteasomal degradation, as demonstrated by reversal with the antioxidant NAC and the proteasome inhibitor MG132. The study also confirmed increased Bax expression and cytosolic cytochrome c release, implicating the mitochondrial apoptotic pathway.
TOP2A Inhibition and AhR Activation
Wu et al. (2025) 29 identified quercetin as one of three key anti-psoriatic active components (along with astilbin and resveratrol) in Smilax glabra Roxb. extract. Network pharmacology and molecular docking revealed that quercetin exerts synergistic therapeutic effects through TOP2A inhibition and AhR activation. The binding energies of quercetin with TOP2A ranged from -9.6 to -6.2 kcal/mol, and with AhR from -8.5 to -7.0 kcal/mol. In vitro experiments confirmed that quercetin dose-dependently inhibited TOP2A activity and activated AhR. In an IMQ-induced mouse model, the combination of quercetin, astilbin, and resveratrol (ARQ) was significantly superior to individual components in improving PASI scores, reducing splenomegaly, and restoring epidermal structure.
Cell Cycle Regulation
Zhai et al. (2025) 32 performed network pharmacology analysis of Haitang-Xiaoyin Mixture (HXM) and identified quercetin as the core active component with the highest degree value (48). Through machine learning algorithms (LASSO, SVM-RFE, Boruta), two characteristic genes—CXCL2 and CXCR4—were identified as key targets. Molecular docking and molecular dynamics simulations confirmed strong binding between quercetin and CXCL2 (ΔG = -24.88 ± 2.57 kcal/mol). Immune infiltration analysis revealed a significant positive correlation between these characteristic genes and M1 macrophages. Additionally, Shen et al. (2025) 25 identified hub targets including CDC25A, TOP2A, NEK2, and CCNA2 in Qingying Decoction (QYD), genes primarily involved in cell cycle regulation. Molecular docking showed binding energies of quercetin with these hub targets all below -6.5 kcal/mol. In an M5-induced HaCaT cell model, QYD treatment significantly inhibited hyperproliferation and cell cycle progression, downregulated hub target mRNA expression, and suppressed activation of the PI3K/AKT/FoxO pathway.
Immunomodulatory Mechanisms
Dendritic Cell Metabolic Reprogramming
Wang et al. (2025) 27 investigated Cooling Blood and Detoxicating Formular (CBDF) and found quercetin to be a key component binding to AMPKα2. In an IMQ-induced mouse model, CBDF activated AMPK, inhibited glycolysis (reducing HK1), and promoted fatty acid oxidation (increasing CPT1A), thereby suppressing dendritic cell maturation. Molecular docking showed a binding energy of -9 kcal/mol between quercetin and AMPKα2. Prkaa2 (encoding AMPKα2) was significantly downregulated in psoriatic lesions, and Prkaa2 knockout diminished the therapeutic effect of CBDF, confirming AMPKα2 as a key molecule for CBDF/quercetin efficacy.
T Cell Differentiation Regulation
Yuan et al. (2020) 34 investigated the therapeutic effect of taxifolin (dihydroquercetin, 40 mg/kg orally) in IMQ-induced psoriatic mice. Results showed that taxifolin significantly reduced PASI scores, epidermal thickness, and PCNA-positive cell numbers. Flow cytometry analysis indicated that taxifolin selectively inhibited pro-inflammatory Th1 and Th17 cells without affecting total T cell or CD4+ T cell proportions. Mechanistic studies confirmed that taxifolin regulates T cell differentiation by inhibiting Notch1 and JAK2/STAT3 signaling pathways and downregulating transcription factors T-bet, GATA3, and RORγt.
Xu et al. (2026) 30 investigated pine pollen extract rich in dihydroquercetin (DHQ, 70%) and demonstrated that it alleviates IMQ-induced psoriasis through modulation of multiple signaling pathways, including Nrf2/HO-1, NF-κB, and JAK1/STAT3. The study confirmed that DHQ regulates Th17-related gene expression and reduces inflammatory cytokine levels.
Proteomic analysis by Di et al. (2021) 28 revealed that cPLA2 (cytosolic phospholipase A2) was the most significantly downregulated protein following taxifolin treatment in IMQ-induced psoriatic mice, with the PPAR signaling pathway being the most significantly enriched KEGG pathway. Further validation showed that taxifolin inhibited cPLA2 phosphorylation and downstream MAPK pathways (ERK and p38) while upregulating PPAR-γ expression.
Innovative Formulations of Quercetin and Its Derivatives
Nanoformulations for Enhanced Bioavailability
Husseini et al. (2026) 21 optimized rutin-PLGA nanocapsules using Box-Behnken design, achieving an optimal formulation (rutin 30 mg, PLGA 83.5 mg, PVA 4%) with encapsulation efficiency of 82.04%, particle size of 191 nm, and zeta potential of -54.95 mV. Upon dispersion in 1% Carbopol 940 gel (NG4 formulation), ex vivo skin permeation studies showed that NG4 transdermal flux (299.47 μg/h/cm2) was approximately 1.86-fold higher than free rutin gel (161.34 μg/h/cm2). In an IMQ-induced rat psoriasis model, the NG4 treatment group showed significantly lower PASI scores (3.67±0.58) compared to the free rutin gel group (8.00±1.00), with approximately 64% reduction in epidermal thickness, significantly decreased serum inflammatory factors (TNF-α, IFN-γ, IL-6), and restored antioxidant capacity (TAC, SOD).
Fauzy et al. (2026) [17] developed a quercetin-naringenin nanoemulsion (NE-QNH) modified with hyaluronate-phospholipid complex (HA-PC) and coated with thermoresponsive polymers (P188/P407), achieving a particle size of 14.94±0.06 nm and encapsulation efficiency >80%, with HA-PC modification efficiency of 87.4%. Ex vivo skin permeation studies demonstrated 240% improvement in transdermal penetration and 290% enhancement in skin retention. In a psoriasis mouse model, the NE-QNH group achieved a PASI score of 0 (complete remission) compared to 8 (mild) in the free drug group, with histopathology showing complete epidermal recovery.
Cyclodextrin-Liposome-In-Gel System
Zhang et al. (2023) [24] addressed the challenges of poor quercetin solubility (0.2 μg/mL) and low stability by developing a hydroxypropyl-β-cyclodextrin (HPCD)-modified liposome-in-gel system. HPCD formed a coating on the liposomal surface through hydrogen bonding with phospholipid head groups, improving stability. The HPCD/QT Lipo@gel showed 24h cumulative transdermal penetration (17.6 μg/cm2) approximately 6-fold higher than free QT@gel (2.9 μg/cm2), and skin retention (16.2 μg/cm2) approximately 5.8-fold higher. In an IMQ-induced psoriasis mouse model, the HPCD/QT Lipo@gel treatment group showed significantly lower PASI scores (5.2) compared to the free QT@gel group (6.8), with 56.7% reduction in epidermal thickness (model group 244 μm vs treatment group 106 μm), significantly reduced Ki67-positive cells, and downregulated mRNA expression of inflammatory factors (TNF-α, IL-17A, IL-1β).
Rationale for Inclusion of Taxifolin
Although taxifolin (dihydroquercetin) is structurally classified as a flavan-3-ol whereas quercetin is a flavonol, taxifolin was included in this review for the following reasons: (1) taxifolin is a direct metabolic precursor of quercetin in the flavonoid biosynthetic pathway; (2) taxifolin shares key structural features (catechol group at the B-ring, 3-hydroxyl group) that confer similar antioxidant and anti-inflammatory activities; (3) several recent studies have investigated taxifolin in psoriasis models, and excluding it would overlook potentially valuable therapeutic evidence; and (4) comparative analysis of taxifolin and quercetin may reveal structure-activity relationships that guide future drug development.
Synergistic and Comparative Therapeutic Potential
Beyond the investigation of quercetin and its derivatives as single agents, several studies have explored their therapeutic potential in combination with other natural compounds or in comparison with conventional antipsoriatic medications. These investigations provide valuable insights into the potential clinical utility of quercetin-based therapies, either as components of multi-target formulations or as alternatives to existing treatments with improved safety profiles.
Synergistic Effects of Quercetin With Other Natural Compounds

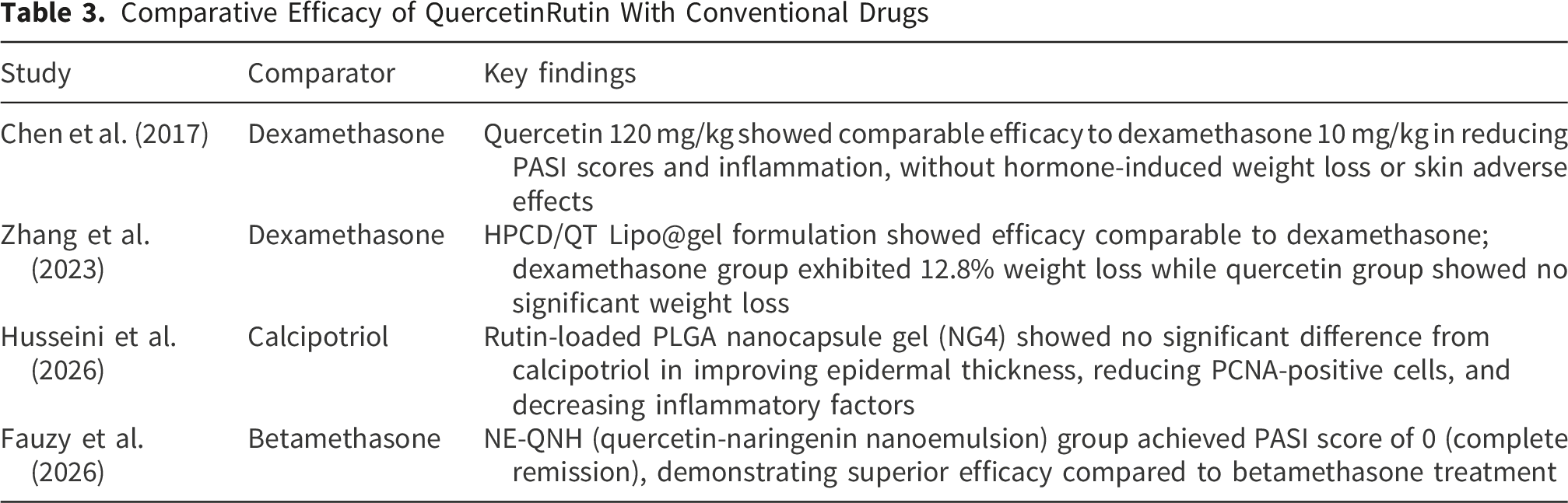
Comparative Efficacy of QuercetinRutin With Conventional Drugs
Discussion
Psoriasis is a chronic inflammatory skin disease driven by genetic, environmental, and immune factors, characterized by hyperproliferation and abnormal differentiation of keratinocytes, along with persistent activation of immune cells. In recent years, with the in-depth understanding of the IL-23/Th17 axis, biologics targeting this pathway have become important therapeutic options for moderate to severe psoriasis. However, these agents are associated with high costs, uncertain long-term safety, and drug resistance in some patients, prompting researchers to turn their attention to natural compounds with multi-target regulatory properties. Quercetin, a flavonoid widely present in vegetables, fruits, and various medicinal herbs, has attracted increasing attention in psoriasis research due to its significant anti-inflammatory, antioxidant, immunomodulatory, and anti-proliferative activities.
This systematic review comprehensively analyzes 22 studies on quercetin and its derivatives (including rutin and taxifolin) in psoriasis treatment, systematically elucidating their multi-layered pharmacological mechanisms. As shown in the Results section, quercetin constructs a multi-target network centered on four interconnected core modules—inflammation, oxidative stress, proliferation, and metabolism—regulating multiple pathological aspects of psoriasis and demonstrating a typical “multi-target, multi-pathway” action profile. This characteristic highly aligns with the complex pathophysiology of psoriasis itself—psoriasis is not driven by a single molecular defect but involves interactions among multiple cell types (immune cells, keratinocytes, endothelial cells) and multiple pathological processes (inflammation, oxidative stress, metabolic dysregulation). 35 Therefore, treatment strategies targeting a single pathway often fail to achieve long-term stable disease control, whereas natural multi-target compounds like quercetin may offer a more comprehensive therapeutic approach.
From a pathophysiological perspective, the development and progression of psoriasis can be viewed as the establishment and perpetuation of an “inflammation-oxidative stress-proliferation” vicious cycle. Initial immune activation (e.g., activation of dendritic cells and T cells) leads to IL-23/Th17 axis activation and massive release of pro-inflammatory factors such as IL-17A and TNF-α. 12 These inflammatory factors, on one hand, directly stimulate keratinocyte hyperproliferation, and on the other hand, induce intracellular ROS production. Excessive ROS not only causes cell membrane lipid peroxidation, DNA damage, and mitochondrial dysfunction but also further activates inflammatory pathways such as NF-κB, forming a positive feedback loop of “oxidative stress-inflammation”. 19 Simultaneously, hyperproliferating keratinocytes have increased energy demands, with upregulated glycolysis and increased expression of the glucose transporter Glut1, and this metabolic reprogramming further supports abnormal cell proliferation. Quercetin and its derivatives intervene at multiple nodes within this vicious cycle: interrupting inflammatory signaling by inhibiting NF-κB and JAK/STAT pathways17,19; enhancing antioxidant defenses and scavenging excessive ROS by activating the Nrf2/Keap1 pathway; and inhibiting metabolic support and hyperproliferation of keratinocytes by downregulating the PI3K/AKT/Glut1 signaling axis. This multi-level intervention strategy theoretically can more effectively break the pathological cycle of psoriasis. 36
Notably, the role of mitochondrial dysfunction and mitochondrial ROS (mtROS) in the pathogenesis of psoriasis has received increasing attention in recent years. The review by Ahmad Jamil and Abdul Karim systematically elaborates the central role of mtROS in psoriasis: mtROS,.37,38 Concurrently, mtROS-mediated oxidative stress can lead to abnormal proliferation and differentiation of keratinocytes, promoting epidermal hyperplasia and plaque formation. More importantly, inflammatory factors such as IL-17A and TNF-α can, in turn, stimulate immune cells and keratinocytes to produce more mtROS, forming a self-amplifying vicious cycle. 39 In the studies included in this review, quercetin and its derivatives effectively broke this cycle by activating the Nrf2 pathway, upregulating antioxidant enzyme activities (SOD, CAT), and reducing MDA and ROS levels. 40 In particular, the study by Hu et al. 20 using Nrf2 knockout mice directly demonstrated that Nrf2 is a key target for rutin’s antioxidant effects, providing strong experimental evidence for treatment strategies targeting mtROS.
From the perspective of metabolic regulation, psoriasis can be regarded as an immunometabolic disease. The activation of immune cells and the hyperproliferation of keratinocytes are both accompanied by metabolic reprogramming, shifting from oxidative phosphorylation to glycolysis (i.e., the Warburg effect). 41 The review by Theodosis-Nobelos et al. elaborates the potential of various natural antioxidants, delivered via nanocarriers, to regulate immunometabolism. 42 In the present review, the study by Wang et al. first reported that quercetin, by activating AMPK, regulates metabolic reprogramming in dendritic cells—inhibiting glycolysis (reducing HK1) and promoting fatty acid oxidation (increasing CPT1A)—thereby suppressing dendritic cell maturation and antigen presentation function. 27 This discovery links cellular metabolism to immune function, providing a novel perspective for understanding the immunomodulatory mechanisms of quercetin. AMPK, as a cellular energy sensor, can simultaneously regulate inflammation, oxidative stress, and metabolism upon activation, making it a key molecule connecting multiple pathological aspects of psoriasis. 43 Quercetin’s activation of AMPK positions it as an ideal candidate for regulating immunometabolism.
T cell differentiation imbalance is a core aspect of psoriasis immunopathology. Th17 cells are the main source of IL-17 and play a central role in psoriasis pathogenesis. The study by Yuan et al. 34 found that taxifolin selectively inhibits pro-inflammatory Th1 and Th17 cells without affecting the proportions of total T cells or CD4+ T cells, demonstrating good selectivity. Further mechanistic studies confirmed that this effect is associated with inhibition of the Notch1 and JAK2/STAT3 pathways and downregulation of the transcription factors T-bet, GATA3, and RORγt. Notably, the differentiation and functional maintenance of Th17 cells also depend on glycolytic metabolism, with Glut1-mediated glucose uptake being a critical step for Th17 cell activation. 44 By downregulating Glut1 expression, quercetin may not only affect the energy metabolism of keratinocytes but also directly inhibit the metabolic support of Th17 cells, thereby reducing IL-17 production at its source. 12 This dual regulation of immune cells and structural cells may be an important feature distinguishing quercetin from traditional immunosuppressants.
Keratinocyte hyperproliferation is the most typical histopathological feature of psoriasis, and this process is closely related to cellular energy metabolism. Studies by Chen et al. 12 and Ma et al. 13 collectively reveal the key role of the PI3K/AKT/Glut1 axis. Glut1, as a glucose transporter, is upregulated in hyperproliferating cells to meet their increased energy demands. 45 Quercetin, by inhibiting the PI3K/AKT pathway, downregulates Glut1 expression, thereby blocking glycolysis, limiting the energy supply to keratinocytes, and inducing apoptosis. This mechanism parallels the concept of cancer metabolic therapy—inhibiting tumor growth by cutting off energy supply. Keratinocytes in psoriatic lesions exhibit metabolic features similar to tumor cells to some extent, and targeting Glut1 may be a novel strategy for psoriasis treatment. 46 The study by Zhang et al. 33 confirmed that inhibiting Glut1 significantly reduced lesion severity in an animal model of psoriasis, further supporting this concept.
From the perspective of formulation innovation, multiple studies included in this review provide solutions to the clinical challenges of quercetin application. The poor water solubility (0.2 μg/mL), low bioavailability, and poor skin permeability of quercetin are major obstacles to its clinical translation. Zhang et al. 33 developed a hydroxypropyl-β-cyclodextrin (HPCD)-modified liposome-in-gel system, where HPCD forms a coating on the liposomal surface via hydrogen bonding with phospholipid head groups, improving stability. The HPCD/QT Lipo@gel showed approximately 6-fold higher transdermal penetration and 5.8-fold higher skin retention compared to free QT@gel, achieving a 56.7% reduction in epidermal thickness in a psoriasis mouse model. Husseini et al. 21 used PLGA nanocapsules to encapsulate rutin, achieving a 1.86-fold increase in transdermal penetration and reducing PASI scores from 8.00 to 3.67. Fauzy et al. 22 developed a quercetin-naringenin nanoemulsion (NE-QNH) that achieved a 240% improvement in transdermal penetration and 290% enhancement in skin retention, with the treatment group achieving a PASI score of 0 (complete remission). These formulation advances indicate that the clinical potential of quercetin is gradually transitioning from theory to reality through modern drug delivery technologies. Notably, many antioxidants mentioned in the review by Ahmad Jamil and Abdul Karim (e.g., curcumin, resveratrol, EGCG) also face low bioavailability challenges, and nanodelivery systems may be an effective strategy to address the common issues of such natural products. 38
From a comparative efficacy perspective, studies included in this review show that optimized quercetin/rutin formulations are comparable or even superior to conventional therapeutic agents in multiple outcome measures. Chen et al. 12 found that quercetin 120 mg/kg showed comparable efficacy to dexamethasone 10 mg/kg, without hormone-induced weight loss or skin adverse effects. In the study by Zhang et al, 33 the dexamethasone group exhibited 12.8% weight loss, while the quercetin group showed no significant weight loss. Husseini et al. confirmed that the rutin-loaded PLGA nanocapsule gel showed no significant difference from calcipotriol in improving epidermal thickness, reducing PCNA-positive cells, and decreasing inflammatory factors. Fauzy et al. 22 demonstrated that NE-QNH achieved superior efficacy compared to betamethasone (PASI score 0 vs. incomplete remission). These comparative results have important clinical implications, as glucocorticoids and vitamin D3 derivatives are first-line topical treatments for psoriasis, but long-term use is associated with side effects such as skin atrophy, telangiectasia, and rebound. 47 Quercetin, as a natural product, exhibits a better safety profile while achieving comparable efficacy, providing strong support for its use as an alternative or adjunctive therapy.
Based on the above analysis, we can propose an integrated pathophysiological model: The development of psoriasis begins with immune activation in genetically susceptible individuals triggered by environmental factors. Activated dendritic cells secrete IL-23, inducing Th17 cell differentiation and IL-17A production. IL-17A acts on keratinocytes, directly stimulating their hyperproliferation while also inducing intracellular ROS production and mitochondrial dysfunction. 48 Excessive mtROS activates inflammatory pathways such as NF-κB, promoting further release of inflammatory factors, forming a positive feedback loop. Simultaneously, hyperproliferating keratinocytes undergo metabolic reprogramming, with upregulated Glut1 expression and enhanced glycolysis to support their abnormal proliferation. 49 Quercetin and its derivatives intervene at multiple nodes within this model: inhibiting dendritic cell maturation (via the AMPK pathway), suppressing Th17 cell differentiation (via the Notch/JAK/STAT pathway), scavenging excessive ROS (via the Nrf2 pathway), and inhibiting keratinocyte proliferation (via the PI3K/AKT/Glut1 pathway). This multi-target intervention strategy theoretically can more effectively break the pathological cycle of psoriasis and restore skin homeostasis.
From a clinical translation perspective, future research should focus on the following directions: First, conducting rigorously designed randomized controlled trials to validate the clinical efficacy and safety of quercetin and its derivatives, particularly their value as first-line or adjunctive therapy in patients with mild to moderate psoriasis. Second, deeply investigating the synergistic mechanisms of quercetin with other natural compounds and optimizing combination regimens, such as the quercetin-naringenin and quercetin-astilbin-resveratrol combinations reported by Fauzy et al. 22 and Wu et al. 29 in this review. Third, further optimizing formulation technologies to improve the bioavailability and targeting of quercetin, particularly developing stable formulations suitable for long-term topical use. Fourth, exploring the potential therapeutic value of quercetin for psoriasis comorbidities (e.g., psoriatic arthritis, cardiovascular disease, metabolic syndrome). The review by Ahmad Jamil and Abdul Karim 38 emphasizes the role of oxidative stress in systemic comorbidities of psoriasis, and the systemic antioxidant and anti-inflammatory effects of quercetin may also benefit these comorbidities. Fifth, combining modern omics technologies and bioinformatics methods to deeply dissect the multi-target network of quercetin, identify key targets and biomarkers, and provide a basis for precision medicine.
Considerations Regarding Bioinformatics and Molecular Docking Studies
Several included studies (e.g., Zhou et al., 2025; Zhai et al., 2025; Shen et al., 2025) employed bioinformatics analyses and molecular docking to predict potential targets of quercetin. While these computational approaches provide valuable hypotheses, they do not substitute for experimental validation. The binding affinities reported (e.g., -9.6 to -6.2 kcal/mol for TOP2A) are theoretical predictions, and the actual in vivo interactions may differ. Readers should interpret these findings as exploratory, and future experimental studies are needed to confirm the predicted targets.
Limitations
Several limitations of this review should be acknowledged. First, most included studies (21 of 22) were preclinical (in vitro or animal models), with only a limited number of clinical studies, which restricts the direct generalizability of findings to human patients. 50 This limitation suggests that the promising preclinical efficacy may not fully translate to clinical settings, and future well-designed randomized controlled trials are needed to validate these findings. Second, the small sample sizes of animal studies (typically 6-10 mice per group) may increase the risk of type I and type II errors, potentially inflating effect size estimates; future animal studies should follow ARRIVE guidelines with rigorous sample size calculations. 51 Third, heterogeneity in intervention protocols and outcome measures precluded meta-analysis, limiting quantitative synthesis; standardized protocols would facilitate cross-study comparisons. Fourth, publication bias may exist as studies with positive findings are more likely to be published than those reporting null or negative results, potentially overestimating therapeutic effects. 52 Fifth, only English and Chinese publications were included due to language constraints, potentially introducing language bias; however, previous meta-epidemiological studies suggest that excluding non-English studies may have minimal impact on overall conclusions. Sixth, none of the included RCTs reported the fragility index, limiting assessment of statistical robustness; the fragility index quantifies the minimum number of event reversals required to change statistical significance and should be reported alongside conventional p-values in future trials. 53 Seventh, several studies lacked detailed documentation of data integrity (e.g., complete chemical validation parameters, cell line authentication), and the quality of some studies may be overestimated due to incomplete reporting; future studies should adhere to minimum reporting standards such as ARRIVE guidelines and provide STR authentication for human cell lines. 54 Despite these limitations, the 22 included studies collectively suggest that quercetin and its derivatives may exert multi-target therapeutic effects in psoriasis treatment. However, as noted above and in Table 1, the incomplete reporting of validation parameters and cell line authentication in several studies limits the strength of this evidence. Therefore, while the mechanistic consistency across studies is encouraging, the current evidence base should be regarded as preliminary, and confirmatory studies with comprehensive data integrity documentation are warranted.
Implications for Future Research: ALCOA+ Principles in Preclinical Natural Product Studies
The findings of this systematic review highlight an important methodological concern in the current body of literature on natural products for psoriasis: the incomplete reporting of analytical validation parameters and biological resource authentication. As shown in Table 1, while most included studies reported compound verification by HPLC or HPLC-MS, critical validation parameters such as calibration curves, recovery rates, limits of detection and quantification, and intra-day/inter-day precision were often omitted. Similarly, several studies did not provide STR profiling or authentication documentation for human cell lines (e.g., HaCaT keratinocytes), which is essential for ensuring the reproducibility and reliability of cell-based experiments [54].
To address these issues and strengthen the translational potential of quercetin-based therapies, future studies should adhere to the ALCOA+ principles for data integrity. In the context of preclinical natural product research, these principles would require: (1)
By implementing these standards, future research can provide a more robust evidence base for the clinical translation of quercetin and its derivatives in psoriasis treatment, addressing the reproducibility concerns raised in the present review.
Conclusions
In conclusion, this review of 22 studies suggests that quercetin and its derivatives (rutin and taxifolin) may exert multi-target therapeutic effects in psoriasis treatment through coordinated regulation of inflammatory (NF-κB, JAK/STAT), antioxidant (Nrf2), anti-proliferative (PI3K/AKT/Glut1), and immunometabolic (AMPK) pathways. Advanced nanoformulation technologies have shown promise in enhancing the bioavailability and skin permeation of these compounds. However, given the incomplete reporting of analytical validation parameters and cell line authentication in several included studies, these findings should be interpreted as preliminary. Future large-scale, rigorously designed clinical trials, along with preclinical studies conducted in accordance with ALCOA+ principles to ensure data integrity and reproducibility, are warranted to validate these findings and establish quercetin-based therapies as viable alternative or adjunctive treatment options for patients with psoriasis.
Supplemental Material
Supplemental Material - The Potential of Quercetin and Its Derivatives in the Treatment of Psoriasis: A Systematic Review
Supplemental Material for The Potential of Quercetin and Its Derivatives in the Treatment of Psoriasis: A Systematic Review by Yuqiao Pan, Mingqiu Mei, Yafang Liu in Natural Product Communications
Supplemental Material
Supplemental Material - The Potential of Quercetin and Its Derivatives in the Treatment of Psoriasis: A Systematic Review
Supplemental Material for The Potential of Quercetin and Its Derivatives in the Treatment of Psoriasis: A Systematic Review by Yuqiao Pan, Mingqiu Mei, Yafang Liu in Natural Product Communications
Footnotes
Ethical Considerations
The authors affirm that this manuscript represents original work and has not been published elsewhere, nor is it under consideration by any other journal. All sources of information are accurately cited and referenced, and any use of third-party material is properly acknowledged. In accordance with the journal’s policies, the authors have no undisclosed competing interests to declare, as stated in the Competing Interests section. The authors are responsible for the accuracy and originality of the content, and any potential issues regarding plagiarism, data fabrication, or inappropriate authorship are strictly disclaimed. The authors affirm their commitment to upholding the integrity of the academic record and will cooperate fully with the editorial office to address any post-publication issues should they arise.
Authors’ contributions
Y.P, and F.L. wrote the main manuscript text and prepared figures. Y.P, F.L., and M.M. reviewed the manuscript. Y.P, F.L., and M.M. approved the final version of manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
This study is a systematic review. All the data sourced from the articles listed in the tables within the manuscript.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.