
Abstract
The poor prognosis constitutes a significant difficulty for spinal cord injury (SCI) individuals. Although mesenchymal stem cells (MSCs) hold promises as advanced therapy medicinal products (ATMPs) for SCI patients, challenges such as Good Manufacturing Practice-compliant manufacturing, cellular senescence, and limited therapeutic efficacy continue to hinder their clinical translation. Recent advances have identified botanical nanovesicles (BNs) as potent bioactive mediators capable of “priming” MSCs to self-rejuvenate, augment paracrine effect, and establish inflammatory tolerance. In this review, we introduce the physicochemical properties of BNs and systematically explore their synergistic relationship with MSCs in regenerative medicine. By integrating BNs with MSC, BNs-empowered MSCs (Be-MSCs) represent next-generation ATMPs. This innovative strategy addresses the limitations of conventional MSC therapies and offers a scalable, nonimmunogenic solution with significant potential for clinical application in SCI.
Impact Statement
Despite the potential of mesenchymal stem cells (MSCs) as regenerative therapies in spinal cord injury (SCI), challenges such as manufacturing standardization, cellular senescence, suboptimal efficacy, and low survival rate in the injury site impede their clinical translation. Botanical nanovesicles (BNs) serve as potent bioactive mediators, priming MSCs to self-rejuvenate, augment paracrine effect, and bolster inflammatory tolerance. In this review, BN-empowered MSCs (Be-MSCs), a next-generation therapeutic product, was thoroughly discussed in SCI, aiming to bring new hope for SCI patients.
Introduction
Spinal cord injury (SCI) disrupts the transmission of nerve signals and stands as one of the most severe conditions in neurology. The following injury frequently results in lasting deficits in sensory and motor functions as well as dysfunction of the autonomic nervous system, significantly exacerbating both the physiological and psychological burdens patients face. 1 Current research has predominantly concentrated on mesenchymal stem cell (MSC) therapies, which demonstrate therapeutic potential by mitigating inflammatory processes and supporting neuronal survival, among other mechanisms. 2 The success of MSC therapy is contingent upon the biological performance of MSCs. However, the hostile environment in injury sites frequently jeopardizes MSCs and adversely impacts therapeutic outcomes.
In the 1960s, Halperin’s team first documented the phenomenon of multivesicular bodies fusing with the plasma membrane in carrot suspension cultures and releasing exosome-like vesicles into the cell wall region. 3 For decades afterward, botanical nanovesicle (BN) hypotheses persisted without experimental validation until Regente et al. achieved a critical breakthrough. Through pioneering the tissue-infiltration-centrifugation technique, they successfully isolated exosome-like vesicles from apoplastic wash fluids of sunflower (Helianthus annuus) seeds. 4 In 2013, the landmark study by Ju et al. established grape exosome-like nanoparticles as cross-kingdom bioactive agents capable of modulating mammalian intestinal cell activity. 5 This breakthrough ignited exponential research expansion, establishing BNs as a transformative frontier in nanomedicine. 6 Complementing this potential, their excellent biocompatibility and drug-delivery capabilities further solidify them as a significant research hot spot.7,8 Unlike animal-derived exosomes, it can transport not only proteins, microRNA, and lipids but also active pharmaceutical ingredients. They infiltrate mammalian cells and act as interspecies messengers, potentially exhibiting anticancer, antioxidant, and anti-inflammatory properties. 9 As naturally derived nanocarriers, BNs are increasingly recognized as a transformative tool in drug delivery and biotechnology, offering a promising platform for innovative therapeutic strategies. 10
The application of BNs to augment the therapeutic functionality of MSCs represents a compelling and rapidly evolving area of scientific exploration. This review comprehensively synthesizes current research, highlighting the potential of BNs to address key limitations in MSC-based therapies, such as MSC senescence, poor cell survival rate, and suboptimal paracrine activity. By leveraging the unique bioactive properties of BNs, this innovative approach holds promise for overcoming critical barriers in regenerative medicine (Fig. 1). Ultimately, BN-empowered MSCs (Be-MSCs) may serve as a transformative strategy to unlock novel therapeutic solutions for complex conditions such as SCI.
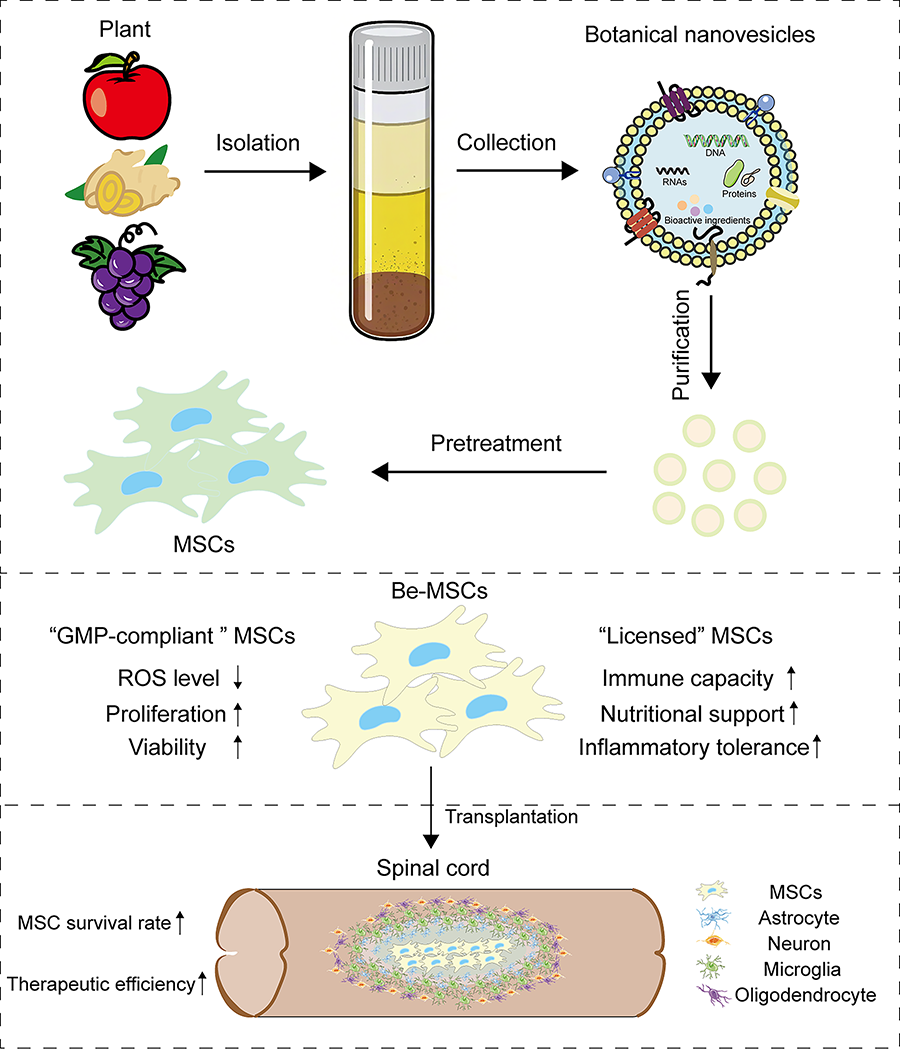
The acquisition of Good Manufacturing Practice (GMP)-compliant MSCs and licensed MSCs through BNs facilitates the clinical translation of therapeutic interventions for SCI. MSCs, mesenchymal stem cells; BNs, botanical nanovesicles; SCI, spinal cord injury.
Prospects for MSC-Mediated Repair in SCI
The secondary injury phase after SCI determines the prognosis of patients. This phase typically unfolds over hours, days, or even weeks after the initial trauma and is characterized by a cascade of complex biological events that exacerbate the initial injury. Key mechanisms driving secondary injury include sustained inflammation, oxidative stress, excitotoxicity, and apoptotic cell death, contributing to further neuronal loss and demyelination. These pathological processes exacerbate tissue damage and create a hostile microenvironment that impedes endogenous repair mechanisms and functional recovery. 11
To date, MSCs have been classified as advanced therapy medicinal products (ATMPs) owing to their demonstrated therapeutic potential in regenerative medicine applications. They are a distinct class of multipotent stem cells recognized for their remarkable capacity for self-renewal and differentiation into various cell lineages, including adipocytes, chondrocytes, and osteoblasts. In addition to their multipotent differentiation capacity, MSCs exhibit immunoprivileged properties characterized by negligible expression of major histocompatibility complex class I and II molecules, a feature that supports their potential application as “universal donor” cells in clinical therapies. 12 Significantly, MSCs exhibit immunomodulatory functions and secrete trophic factors to the injured microenvironment, facilitating tissue regeneration (Fig. 2).13,14
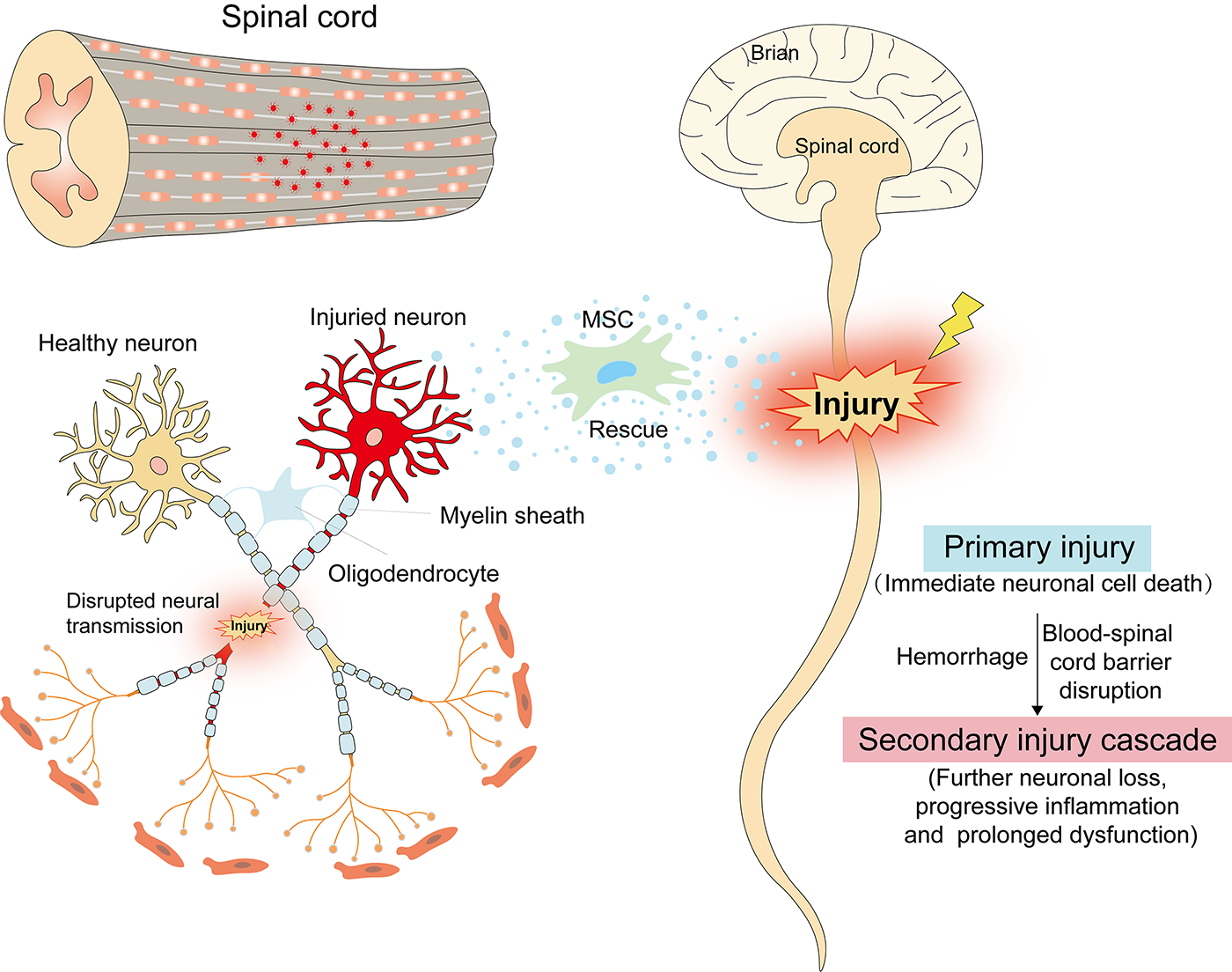
MSC-mediated regenerative strategy in traumatic SCI repair. MSC, mesenchymal stem cell; SCI, spinal cord injury.
Following SCI, the disruption of the blood–spinal barrier triggers a robust immune response. Microglia, the resident immune cells of the central nervous system, were activated into M1 inflammatory microglia. Transplanted MSCs exhibit targeted migration to lesion sites, modulating the immune microenvironment through direct cell–cell interactions and paracrine-mediated signaling pathways. 15 Specifically, exosomes released by MSCs are internalized by microglia, driving their polarization from pro-inflammatory M1 to anti-inflammatory M2 phenotypes. This shift suppresses neuroinflammation, improves tissue retention, and creates a regenerative microenvironment conducive to axon regrowth and neuronal recovery after SCI. 16 In addition, MSC-derived cytokines contribute to neuroprotection and neuroregeneration by modulating immune responses and enhancing neuronal survival. 17 These findings highlight the multifaceted paracrine actions of MSCs, which collectively promote functional recovery post-SCI. MSCs also exert therapeutic effects in SCI through mitochondrial quality control mechanisms. MSC-mediated mitochondrial transfer, which restores the neuronal mitochondrial pool through intercellular tunneling nanotubes or gap junctions, may represent another key reason.18,19 Incorporating exogenous mitochondria into damaged neurons aids in maintaining adenosine triphosphate homeostasis and protects against excitotoxicity and calcium overload in the central nervous system.20,21 Importantly, the efficacy of MSC therapy in SCI is not attributed to a single mechanism, but arises from the coordinated regulation of multiple pathways, including mitochondrial repair, anti-inflammatory responses, and neuroprotection. These multifaceted actions underscore the potential of MSCs as a promising therapeutic strategy for SCI.
Critical Barriers Hindering MSC Efficacy in SCI
MSC therapy, while widely researched and recognized as a promising approach for disease, faces significant challenges in clinical translation. Most clinical-stage MSC therapies have been unable to meet primary efficacy end points.22,23 Despite decades of study, only one U.S. Food and Drug Administration (FDA)-approved MSC therapy existed as of December 2024, highlighting the scarcity of robust clinical evidence supporting its efficacy and safety in humans. 22 Current clinical trials remain in early exploratory phases, with limited large-scale validation of their therapeutic outcome. Furthermore, inconsistencies in quality control standards (e.g., variations in MSC sourcing, expansion protocols, and functional characterization) and heterogeneous therapeutic effects across trials have hindered reliable clinical application. The application of MSC in SCI is complicated, primarily due to the hostile postinjury microenvironment that severely compromises transplanted cell survival and therapeutic efficacy. Following injury, processes such as inflammation, glial scar formation, and oxidative stress create a milieu that compromises MSC survival, engraftment, and functional integration. This environment may diminish the cells’ ability to exert neuroprotective or regenerative effects. We will delve into these challenges from the following points.
Manufacturing Challenges: Achieving Good Manufacturing Practice-compliant MSC Production
Maintaining optimal cellular activity and quality is critical to the efficacy of cell-based therapies. The production of Good Manufacturing Practice (GMP)-compliant MSCs is an intricate and highly regulated process encompassing various stages of cell handling, characterization, and rigorous quality control. 24 To safeguard the quality, safety, and efficacy of such innovative cell and gene therapies, dedicated regulatory frameworks were established. While initial European Union medicines legislation (Directive 2003/63/EC, Regulation 726/2004/EC) addressed the potential of cell therapies as medicines, the specific ATMP framework arrived later with Regulation 1394/2007/EC in 2007. Subsequent directives have further defined scientific and technical requirements for ATMPs, notably Directive 2009/120/EC and the EU GMP-ATMP guidelines. 25 The FDA also uses a risk-based approach, categorizing cell-based products into two groups as follows: minimally manipulated (cryopreservation, thawing, density gradient isolation, washing, and dilution) and more than minimally manipulated (expansion and genetic modification). 25 Consequently, MSC cultures derived from diverse tissue sources must adhere to GMP standards, with rigorous monitoring to ensure sterility, safety, potency, and compliance with predefined specifications.
A central challenge arises from the tension between clinical demand and manufacturing constraints. Achieving therapeutic doses (typically 1–2 million cells per kilogram of body weight) necessitates extensive passaging during in vitro expansion. 26 However, prolonged passaging risks compromising cell quality and functionality, directly conflicting with GMP requirements for maintaining batch consistency and safety. This scalability-efficacy paradox represents a primary barrier to the widespread clinical adoption of MSC therapies.
Cellular senescence: Functional decline of aged MSCs
Cellular senescence, a stress-triggered irreversible proliferative arrest, manifests through persistent metabolic activity and senescence-associated secretory phenotype (SASP)-mediated hypersecretion of pro-inflammatory/proteolytic factors.27–29 As soon as the number of cell divisions reaches Hayflick’s limit, cellular senescence promotes the aging and dysfunction of organisms. 30 These senescence-associated processes also substantially impair the regenerative capacity and therapeutic efficacy of MSCs.
Autologous MSC transplantation has emerged as a clinically recommended therapeutic strategy, offering distinct advantages in personalized regenerative medicine for sufficient quantities directly from patients. However, the problem we may face is that the quality and lifespan of MSCs are influenced strongly by the chronological age of a donor. 31 In donors, intrinsic aging (also termed in vivo aging) directly triggers profound functional impairments in MSCs, evidenced by decreased cell population, compromised migratory potential, and attenuated differentiation capacity.32–35 Particularly concerning is the emergence of SASP, which transforms these cells into pro-inflammatory mediators through paracrine signaling (Fig. 3). 36 The impaired functionality of MSCs derived from elderly donors may notably contribute to the marked disparities in clinical therapeutic outcomes.
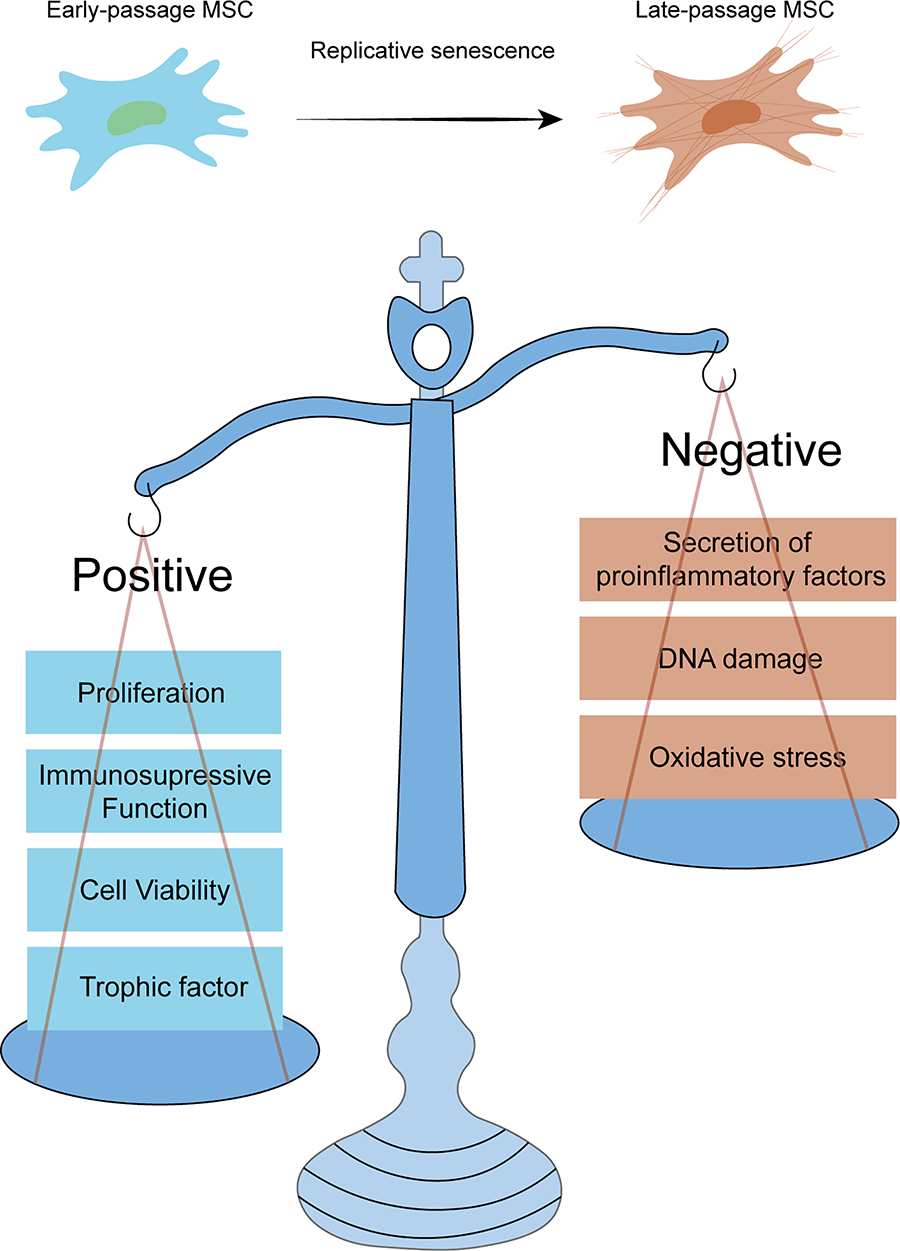
Alterations in cellular functions during MSC senescence. MSC, mesenchymal stem cell.
Umbilical cord mesenchymal stem cells (UC-MSCs) provided an alternative strategy due to their derivation from young donors, circumventing age-related cellular senescence inherent in adult-derived stem cells. However, their clinical application is limited by the substantial costs associated with long-term cell banking for maintaining cell inventories. Critically, the challenge of replicative senescence during extended in vitro culture periods remains a persistent concern in cellular applications.37,38 Researchers demonstrated that even cells with initially robust proliferative potential eventually undergo senescence due to telomere shortening and accumulated DNA damage during repeated divisions. 39 In vivo aging pertains to the chronological age of MSC donors, which directly impacts cellular longevity, whereas in vitro aging manifests as the progressive loss of stem cell properties due to senescence-associated phenotypic alterations during culture expansion. 40 Analogous to in vivo aging, in vitro aging induces marked molecular divergence that profoundly impacts cellular architecture, mitochondrial dynamics, and age-related stress responses, accompanied by characteristic cytomorphological transformations, including nuclear enlargement and chromatin structural reorganization. 41 The distinction lies in that in vitro aging of MSCs appears to result in more severe consequences, including a complete loss of their progenitor cell signature. 42 Long-term in vitro culture may affect MSCs more than the natural aging process of their host. 43 The artificial environment fails to replicate native niche conditions, with suboptimal nutrient gradients and mechanical stimuli precipitating premature cell cycle arrest (G1 phase accumulation) and senescence-associated β-galactosidase overexpression. Some external exposures, such as reactive oxygen species (ROS), significantly accelerate the functional degradation of MSCs. 44 These physical and chemical cues indicate that senescence resulting from in vitro replicative expansion definitively leads to the functional deterioration of MSCs.
The senescence of MSCs, referred to as in vitro or in vivo, compromises their functional capacity and poses a significant barrier to clinical translation. Future efforts focused on combating the MSC senescence will be essential in the advancement of ATMPs.
Suboptimal treatment responses: A consequence of physiologically limited regeneration
Although MSC-based therapies have demonstrated therapeutic efficacy in SCI, current treatment regimens remain insufficient to reverse the pathological sequelae of neural damage fully. MSC-mediated therapeutic outcomes for SCI stem fundamentally from intrinsic biological constraints on their regenerative potential. Understanding the intrinsic biological determinants governing MSCs constitutes a pivotal prerequisite for advancing their therapeutic applications.
The intrinsic biological abilities of MSCs directly correlate with their therapeutic efficacy across pathological conditions. MSCs exhibit distinct regenerative capacities dictated by their embryonic germ layer origins and distribution position.45,46 For instance, ectodermal mesenchymal stem cells (EMSCs) exhibit fundamental functional divergence from their mesoderm-derived counterparts, particularly in neuroregenerative applications. Originating from neuroectodermal progenitors sharing ontogenetic lineage with neural progenitor cells, they demonstrate enhanced neurogenic differentiation capacity and superior neurotrophic factor secretion efficacy. 47 Other studies highlighted that UC-MSCs exhibit superior immune regulatory capabilities compared with other stem cells.48–50 Despite the advantages of MSCs derived from various tissue sources in specific fields, their current efficacy remains insufficient to reverse SCI completely. Achieving complete functional recovery necessitates a multifaceted enhancement of MSC performance (referred to as MSC v2.0) across various therapeutic dimensions.
Hostile microenvironment: Poor MSC survival post-transplantation
The mechanism underlying the efficacy of MSCs in treating diseases remains controversial within the scientific community. Researchers have proposed divergent mechanisms to explain the therapeutic effects of MSCs: some emphasize that the paracrine activity of surviving MSCs at the injury site plays a predominant role. In contrast, others contend that the regenerative function is mediated through MSC apoptosis in vivo.51–53 Notably, post-transplantation apoptosis patterns alone cannot fully account for the observed clinical efficacy variations. A plausible hypothesis is that the survival rate of MSCs determines the duration of their paracrine effects, thereby influencing the overall recovery outcome. 54
The engraftment efficiency of transplanted MSCs at lesion sites is critically modulated by two distinct yet interconnected phases as follows: (i) preimplantation parameters encompassing administration protocol design (including delivery route optimization and dosage calibration), cellular preparation techniques (involving cell source selection, viability assessment, and tissue engineering), and (2) postimplantation dynamics governed by the pathophysiological microenvironment (characterized by inflammatory cytokine profiles and hypoxic stress gradients). The postimplantation phase of MSCs constitutes a critical survival rate determinant, in which dynamic microenvironmental interactions mediate cellular survival kinetics. This pivotal biological process warrants in-depth discussion due to its fundamental role in modulating engraftment success rates and long-term tissue regeneration outcomes.
SCI triggers a self-perpetuating pathological cascade that catastrophically disrupts the homeostatic equilibrium of the spinal cord microenvironment, establishing a pro-inflammatory microenvironment. The pathological overproduction of ROS at the lesion epicenter has been identified as a principal pathogenic driver, which amplifies neuroinflammatory cascades and dysregulates multiple regulated cell death pathways. 55 In the hostile microenvironment of the injury site, transplanted MSCs must initially confront intrinsic viability challenges before executing their therapeutic potential. Chronic ROS accumulation and sustained pro-inflammatory signaling at the injury site accelerate MSC senescence and/or programmed necrosis. 56 Meanwhile, these cells are subjected to macrophage-mediated phagocytosis, further contributing to MSC loss.57,58 The ultimate result is that only a minimal fraction of the transplanted MSCs can be detected at the injury site post-transplantation.59–61 The strategic modulation of MSCs to resist inflammatory damage from hostile microenvironments may bring new hope for enhancing MSC viability and functional persistence post-transplantation.
BNs as a Multifunctional Platform for MSC Optimization
Developing optimization protocols to improve MSC functionality is poised to be a critical focus in next-generation cell-based therapeutics. Current MSC optimization strategies primarily encompass three principal approaches as follows: genetic engineering, pharmacological preconditioning, and hypoxic conditioning.62–64 Small molecule-based preconditioning has emerged as the predominant research focus of these modalities, owing to its exceptional controllability.
BNs are safe, natural carriers that rarely trigger immune responses. 65 They maintain stable properties and can be produced affordably at large scale-key advantages over many artificial nanocarriers, which often require complex and costly manufacturing. 66 Researchers could choose specific isolation methods based on the plant type, extraction process, and final use to obtain high yields of pure extracellular vesicles efficiently. Predominant techniques include differential ultracentrifugation, density gradient centrifugation, and size-exclusion chromatography. 67 Critically, BNs can be readily engineered using modification or postproduction functionalization to enhance targeting or drug-loading properties. 68 This functionalization capability, coupled with their intrinsic lipid composition and bilayer membrane substructure, makes BNs exceptionally effective carriers for insoluble drugs and facilitates their highly efficient cellular uptake mechanism,69,70 outperforming many conventional delivery systems in bioavailability. It represents a groundbreaking strategy for addressing the limitations commonly encountered in MSCs-based therapies (Fig. 4). These challenges encompass MSC cellular senescence, compromised regenerative capacity, and poor post-transplantation survival. By leveraging BNs to enhance the resilience, functionality, and therapeutic potency of MSCs, this strategy holds the potential to significantly improve the efficacy and reliability of MSC-based treatments in a wide range of medical conditions.
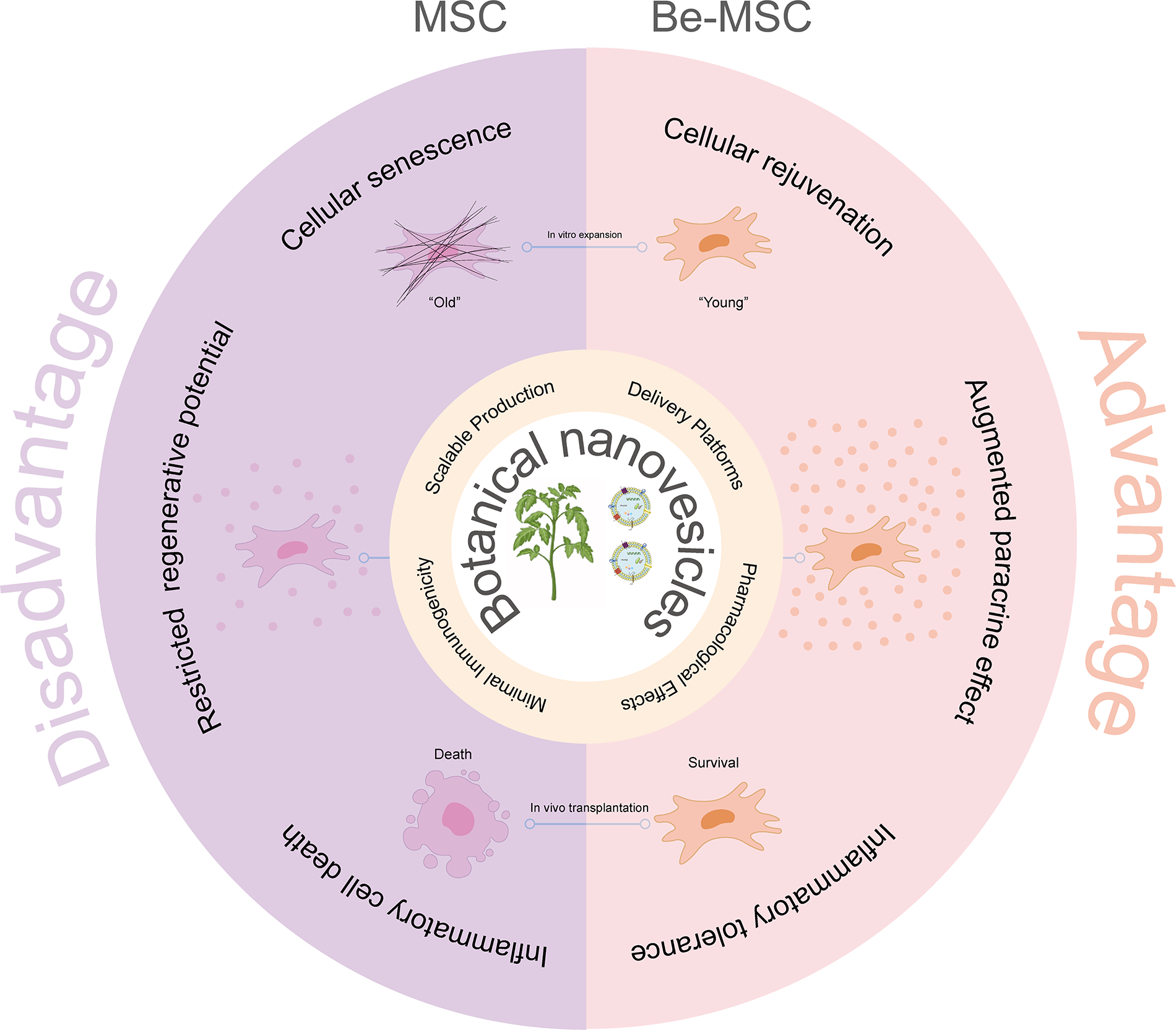
Schematic illustration of BNs as a multifunctional platform for optimizing MSC function. BNs, botanical nanovesicles; MSC, mesenchymal stem cell.
Rejuvenating MSCs: BNs counteract MSC senescence
MSC senescence is a cellular response to intrinsic and extrinsic pressures, constituting a significant barrier to the GMP-compliant MSCs and markedly diminishing their therapeutic potential. 71 Contemporary research reveals that the aging state of MSCs could be reversible through targeted external interventions. 72 BNs, which contain intrinsic drug compositions, lipids, proteins, and mRNAs, act as carriers for bioactive molecules. 73 They play a crucial role in advancing therapeutic applications and enhancing biological interactions by facilitating intercellular communication. Mechanistically, these BNs are readily taken up and integrated by target cells, leading to significant biological effects (Fig. 5).74,75
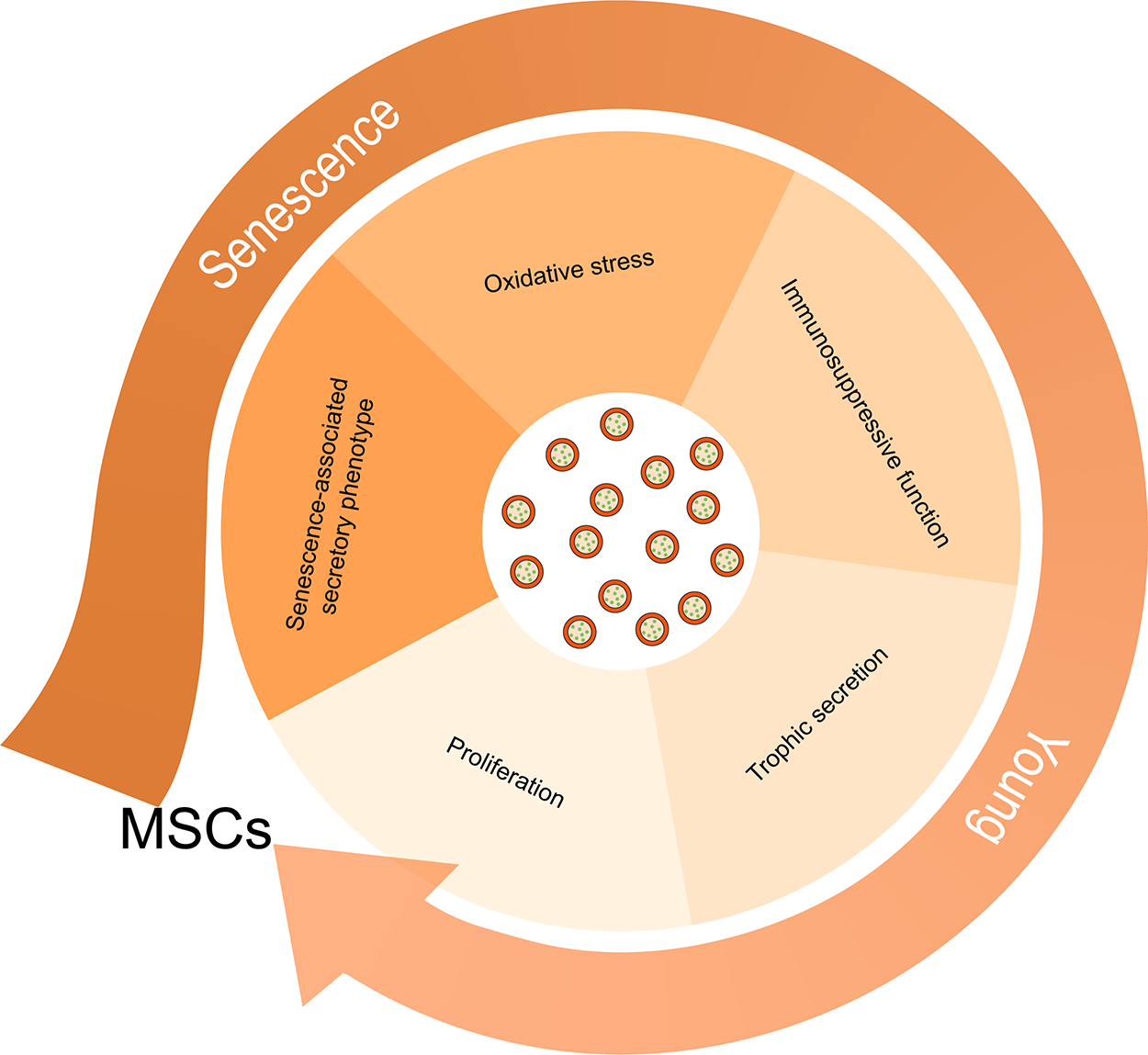
BN-mediated cellular rejuvenation strategy for aging MSCs. BN, botanical nanovesicle; MSCs, mesenchymal stem cells.
Oxidative damage to cellular components has gained widespread recognition as a key pathophysiological event that contributes to the onset and progression of various diseases as well as the aging process. 76 It was also recognized as a major driver of MSC senescence. 77 Plant fruits are one of the most popular and widely consumed healthy diet sources today for antiaging purposes.78,79 Their popularity is primarily due to their high concentration of vitamin C. However, vitamin C has a short half-life, requiring continuous infusion to maintain sufficient in vivo levels. Its pharmacokinetics are complex and influenced by numerous environmental and lifestyle factors, complicating its effective delivery. 80 Recent study has highlighted the ability of BNs to improve the bioavailability and efficacy of vitamin C, thereby augmenting its antioxidant properties. 81 When combined with MSCs, these plant-derived compounds may help prevent or reverse oxidative damage, thereby delaying the onset of MSC senescence and preserving their regenerative capacity.82–84 Another potential reason BNs regulate the youthful state of MSCs is in the drug components they encapsulate. The active ingredients extracted from natural plants have shown superior antiaging properties, involving polysaccharide, polyphenol, saponin, and so on.85–88 These pharmacological effects also apply to the regulation of MSC function. For instance, fucoxanthin derived from brown seaweed could mitigate the age-related decline in vitality and function as well as the detrimental effects of prolonged in vitro culture by modulating the PI3K/Akt/Nrf-2 signaling pathway. 89 The natural bioactive lipid named oleoylethanolamide (OEA) also has been demonstrated to enhance the activity of antioxidant enzymes, including superoxide dismutase and glutathione peroxidase (GSH-px), in MSCs. Pretreatment with OEA improves MSC viability and overall bioperformance. 90 Recent research demonstrated that curcumin liposomes alleviate senescence of bone marrow mesenchymal stem cells by activating mitophagy, which provides new evidence for the antiaging effect of drugs on MSCs. 91 Our recent findings also demonstrated that Lycium barbarum oligosaccharides (LBOs) promote GSH synthesis in EMSCs, reshaping EMSC functional condition via facilitating the removal of ROS. 92
The large-scale production of MSCs is inherently stringent and lengthy. While current methods enhance MSC quality through direct addition of antioxidants like vitamin C and bioactive compounds such as fucoxanthin, these exogenous additives face critical limitations: characteristically short half-lives, unpredictable metabolic kinetics, and inherently low water solubility—all of which drastically constrain bioavailability and consistent efficacy. These fundamental challenges of transient activity and solubility-driven delivery inefficiency underscore the compelling advantages of BNs in MSC production. BNs provide an ideal nano-platform for efficient encapsulation, protection, and targeted delivery of diverse actives. This capability significantly extends compound bioavailability and presents a transformative solution to core biomanufacturing challenges.
MSC functional improvement: BNs augment MSC therapeutic efficiency in SCI
The active ingredients and efficacy profiles of different drugs are distinct, resulting in marked differences in their biological effects. The pathogenesis and progression of certain diseases often guide the selection of BN-based strategies for MSCs, which can be pivotal in influencing cell-based therapies. The severe local pathological cascade and limited nerve regeneration following SCI are primary obstacles to effective treatment. 84 Thereby, the key challenge in MSC therapy lies in enhancing MSC immune regulation and nerve regeneration.
In our previous work, we reported that LBO confers an inflammatory license to EMSCs, enhancing their paracrine effects and ultimately augmenting the therapeutic efficacy of EMSCs in treating SCI. 93 Recently, we have successfully isolated BNs derived from L. barbarum. They serve as an efficient natural carrier, contain various active ingredients, including LBO, and have demonstrated a therapeutic role. 94 Collective clues provide valuable insights for future research to explore the promotive effects of L. barbarum-derived BNs on MSCs for treating SCI. Due to their superior aqueous solubility, polysaccharides can rapidly confer an inflammatory “license” on MSCs. This pharmacological characteristic has motivated our thinking about whether insoluble therapeutic agents could similarly augment the regenerative potential of MSCs.
Numerous insoluble drugs exist, with curcumin being the primary focus of this discussion. Curcumin, a polyphenolic compound, is renowned for its broad spectrum of biological activities, including anti-inflammatory and neuroprotective properties. 95 Curcumin-based therapeutic strategies have achieved significant success in treating SCI.96,97 More intriguingly, some scholars have proposed the application of curcumin to “domesticate” MSCs, thereby equipping them with enhanced immune-regulatory effects. In their study, they evaluated the impact of curcumin on a peripheral blood mononuclear cell/MSC coculture model. They found that low concentrations of curcumin reduced the levels of inflammatory cytokines (IL-17, IL-6, IL-1β, and IFN-γ) while increasing the levels of anti-inflammatory cytokines (TGF-β and IL-10). 98 In addition, other studies have demonstrated that curcumin enhances the immunomodulatory function of MSCs by upregulating the expression of Indoleamine 2,3-dioxygenase 1, which in turn inhibits allogeneic T cell responses.99,100 The above clues also highlight that the effects of curcumin on MSCs are markedly concentration dependent. High concentrations of curcumin promote apoptosis and are detrimental to MSCs, whereas low concentrations inhibit apoptosis, promote cell proliferation, and facilitate the acquisition of an inflammatory “license.” This suggests that the process of “domesticating” MSCs with curcumin is a gradual, concentration-dependent process. However, applying these strategies generally requires methods to enhance the solubility and stability of curcumin to overcome its inherent limitations in these aspects.101–103 Natural BNs exhibit unique advantages for addressing drug delivery challenges without undesirable side effects caused by chemical or biological carriers. 104 Liu et al. isolated and purified nanovesicles from turmeric by ultracentrifugation and sucrose gradient centrifugation. They identified bioactive constituents termed curcumin in nanovesicles with the support of mass spectrometry and HPLC analysis. 105 This discovery positions BNs as efficient natural carriers for curcumin, suggesting a novel delivery paradigm for insoluble pharmaceuticals. Even more promising is the possibility that these BNs, combined with MSCs, could bring novel breakthroughs. BNs may solve this challenge in future research, as they facilitate a durable and more controlled endocytosis process for cellular uptake. 106 As mentioned above, concentration gradients of curcumin have diametrically opposed effects: high concentrations induce MSC apoptosis, whereas low doses promote proliferation and confer inflammatory licensing capacity. This biphasic response underscores the need for precise control in the “domestication” of MSCs, highlighting it as a delicate and finely tuned process.
The preceding examples highlight the superior mechanistic profile of BN-mediated curcumin delivery systems for targeting MSC conditioning relative to conventional strategies. BNs offer dual advantages: (1) overcoming the critical solubility/stability barrier via a safer, more biocompatible solution than synthetic carriers, and (2) circumventing curcumin’s detrimental concentration dependence through controlled intracellular release and sustained low-dose exposure in MSCs. BN-mediated preconditioning licenses MSCs for enhanced therapeutic efficacy in SCI by strategically modulating their paracrine secretion profile. Critically, this functional improvement occurs without compromising cellular viability, thereby amplifying MSC-mediated repair mechanisms.
Prolong MSC survival status: BN-mediated biological effect
While aiming to improve therapeutic outcomes, the arbitrary escalation of dosage or frequency in MSC-based therapies may compromise therapeutic rationality due to the elevated risks of severe clinical complications (e.g., pulmonary embolism) and the disproportionate increase in treatment costs relative to efficacy gains. Optimizing the potency of MSCs to transform them into enhanced ATMPs capable of thriving in hostile biological environments is a top priority.
To date, the low survival rate of transplanted cells remains a significant factor affecting treatment efficacy, especially in SCI, where cell survival is often limited. This highlights the need to explore combined strategies to enhance cell survival. 107 These strategies are tailored to the mechanism of injury and focus on protecting MSCs against detrimental microenvironments characterized by hypoxia, oxidative stress, inflammatory cytokines, and metabolic disturbances.
Long-term severe hypoxia exposure induces metabolic dysregulation and cellular stress responses that significantly impair the survival of transplanted MSCs, as hypoxic microenvironments disrupt their physiological homeostasis and regenerative functions. 108 Concurrently, oxidative stress exacerbates this damage by promoting ROS accumulation, directly disrupting redox balance and triggering inflammatory cascades. 109 In SCI, ROS imbalance perpetuates neuronal damage and correlates with poor clinical outcomes, suggesting that oxidative stress management is critical for modulating post-SCI tissue repair processes.110,111
Enhancing MSC survival under oxidative stress at injury sites involves three primary strategies as follows: tissue engineering, gene modification, and pharmaceutical interventions. (1) Tissue Engineering: This approach requires meticulous optimization of biocompatibility, material toxicity, and MSC mobility within scaffolds to ensure functional integration and regenerative efficacy. However, the complexity of balancing these factors often limits its practicality. (2) Gene modification: While genetically modified MSCs show promise for regenerative therapies, their engineering processes carry risks such as off-target effects and unintended mutations, potentially compromising genomic stability.112,113 These technical and safety challenges collectively underscore the necessity for stringent regulatory oversight in the clinical use of genetically modified MSCs. (3) Pharmaceutical Empowerment: A more streamlined strategy involves pretreating MSCs with exogenous pharmacological agents. Controlled drug treatment could improve the ability of MSCs to resist excessive oxidative stress following transplantation.89,114,115 Several studies have created cellular oxidative stress damage models by exposing MSCs to hydrogen peroxide (H2O2). The results from these studies consistently suggest that pretreating MSCs with specific drugs, such as antioxidants or other protective agents, can significantly enhance their survival under oxidative stress conditions.116,117 This strategy reduces cellular damage caused by ROS and preserves MSC functionality, improving their ability to proliferate, migrate, and differentiate. This pharmaceutical approach demonstrates superior operational precision and process controllability in MSC improvement compared with conventional genetic or physical modification techniques, establishing a paradigm-shifting methodology in cellular stress management.
Studies demonstrate that sustained low-dose exposure to lipopolysaccharide (LPS), a bacterial exotoxin, induces immune tolerance in mice and confers protection against subsequent severe inflammatory diseases.118,119 Extending these discoveries, scholars exploring LPS stimulation of MSCs at varying concentrations and durations confirmed that prolonged, low-dose LPS exposure effectively promotes an inflammatory tolerance state, significantly enhancing MSC survival. 120 This success has spurred broader interest in MSC preconditioning strategies, particularly within plant-derived therapeutics. Although established as a potent antitumor agent inducing dose-dependent cancer cell apoptosis and oxidative stress, andrographolide demonstrates unexplored potential in MSC research. 121 A recent study revealed that andrographolide preconditioning equips MSCs to withstand environmental stressors such as H2O2 and oxygen-glucose deprivation. 122 A key divergence lies in the effective concentration: andrographolide necessitates 10–30 μM for tumor applications, but triggers effective MSC preconditioning at just 1–10 μM. To leverage this difference, concentrations must be carefully controlled, enabling MSCs to acquire inflammatory tolerance while avoiding cytotoxicity. Consequently, we speculate that all antitumor natural compounds that kill tumors via oxidative stress could potentially enhance MSC tolerance to inflammatory damage. Novel drug delivery systems such as BNs, with their capacity for controlled, gentle release, emerge as compelling candidates for future MSC “training” protocols to induce functional tolerance.
Enhancing the inflammatory tolerance threshold of MSCs constitutes a slow, gentle, long-term process, making sustained drug release and precise concentration control essential for generating high-tolerance MSCs. BNs address this need through a controlled, sustained drug delivery approach that circumvents cellular damage from high drug concentrations and leverages gradual release kinetics to elevate the tolerance threshold efficiently. From this perspective, BNs offer distinct advantages for enhancing MSC survival in inflammatory environments.
Conclusions and Future Direction
The central challenge in MSC therapy revolves around two critical axes as follows: establishing robust quality control protocols pretransplantation and ensuring sustained cellular high performance postdelivery. Notably, the pathological microenvironment at SCI sites, characterized by oxidative stress and inflammatory cascades, severely compromises MSC viability, thereby diminishing therapeutic outcomes. These technical barriers have propelled the emergence of MSC2.0 therapeutics, ushering in accelerated progress in regenerative medicine. Current research endeavors are strategically focused on developing tailored MSC subpopulations with enhanced therapeutic profiles through advanced approaches.
BNs have garnered significant scientific attention as a transformative optimization platform for next-generation MSC therapies, which is mainly due to the following reasons: (1) Superior drug delivery efficiency; (2) an inherent lipid bilayer configuration enabling enhanced cellular uptake; and (3) industrial-scale manufacturability that positions them as pivotal components for GMP-compliant MSC v2.0 therapeutic development.
BNs exhibit intercellular signaling capabilities such as animal-derived exosomal counterparts, through bioactive cargo transfer. While functional similarities have been established, the precise compositional profile of BNs remains a critical knowledge gap demanding comprehensive characterization. It is essential to understand this drug’s loading capacity and release efficiency in BNs. Recent advances in nanovesicle modification, exemplified by the development of ROS-responsive hybrid vesicles for rapamycin delivery, suggest that strategically engineered BNs could revolutionize MSC functional improvement. 123 An engaging and thought-provoking question is whether artificial BNs can be used as an effective platform to control the release of active drugs, thereby influencing the future functions of MSCs. Finding the influence targets of MSCs and designing the drug expression patterns of BNs may be an interesting direction for future research.
Authors’ Contributions
Z.W.: Wrote the article and designed figures. W.D.: Participated in the editing of the article. Q.Y.: Provided formulation or evolution of overarching research goals and aims. X.X. and J.Y.: Participated in the management and coordination responsibility for the research activity planning and execution. All authors read and approved the final article.
Footnotes
Funding Information
This work was supported by the National Natural Science Foundation of China (82173785, 82373820, 81720108030, 81803475) and the National Key Research and Development Project of China (2018YFE0208600).
Availability of Data and Materials
Data availability is not applicable to this article as no new data were created or analyzed in this study.
Disclosure Statement
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.