
Abstract
The mechanical properties of the extracellular matrix (ECM) play a critical role in regulating cellular behavior and fate. In the design and application of tissue engineering materials, previous studies have primarily focused on the role of material stiffness (elastic modulus) in modulating cellular events. However, biological tissues and the ECM exhibit more complex mechanical behaviors, such as viscoelasticity, highlighting the importance of considering viscoelasticity as a design parameter for biomaterials. Current biomimetic strategies might place less emphasis on the dynamic mechanical microenvironment of viscoelastic ECMs. Emerging evidence suggests that independently tuning the viscoelasticity of matrices can influence cellular biological processes and enhance tissue regeneration outcomes. This review highlights the emerging focus on independently tunable viscoelastic hydrogels and their potential applications in tissue engineering. In this article, we review the design of hydrogels with adjustable viscoelasticity aimed at guiding cellular and tissue behavior, advancing the development of in vitro cell culture models and in vivo regenerative therapies. This review introduces the concept of viscoelasticity, elaborates on the viscoelastic properties of biological tissues, and summarizes commonly used evaluation metrics and characterization techniques for viscoelasticity. Next, it highlights the strategies for constructing hydrogels with tunable viscoelasticity and discusses the regulatory effects of viscoelasticity on cellular behaviors, along with the associated mechanobiological mechanisms and signaling pathways. Finally, the review provides an overview of the current applications of viscoelastic hydrogels in tissue engineering and offers perspectives on future research directions.
Impact Statement
Viscoelasticity is an essential but often overlooked mechanical property that governs cellular behaviors and tissue remodeling. Recent advances reveal that cells actively sense and respond to viscoelastic cues, influencing adhesion, migration, differentiation, and proliferation. By examining emerging hydrogel designs with independently tunable viscoelasticity, we highlight their potential to enhance cell-instructive biomaterials, improve organoid models, and enable personalized regenerative therapies. This review provides a comprehensive perspective on viscoelasticity-driven cell regulation and offers insights into future directions for designing biomaterials that better mimic native tissue mechanics.
Introduction
Mechanical properties fundamentally determine the suitability of scaffolds for biomedical applications. The elastic modulus is commonly used to characterize material stiffness.1–4 However, biological tissues exhibit viscoelasticity rather than pure elasticity.5–7 The extracellular matrix (ECM) exhibits viscoelastic behavior across multiple length scales, from molecular-level rearrangements to tissue-scale deformations.8,9 Increasing evidence shows that cells sense and respond to ECM viscoelasticity. Controlling matrix viscoelasticity alone offers a method of pure mechanical regulation without the involvement of growth factors, which can serve as a reference for the differentiation of specific cell lineages in tissue engineering.10,11 This challenges the traditional viewpoint of cell–matrix mechanotransduction that is centered on stiffness.11–13 In addition, matrix viscoelasticity plays a crucial role in tissue morphogenesis, providing a potential strategy for guiding tissue development in regenerative medicine. 14
Elasticity and stiffness both refer to the ability of materials to resist elastic deformation. 15 Most materials exhibit approximate elastic behavior under specific conditions but may display viscoelasticity or other complex mechanical behavior under different conditions. 16 Viscoelastic materials display both time-dependent elastic deformation and viscous flow, characterized by phenomena such as stress relaxation, creep, and hysteresis (Fig. 1A).45,46
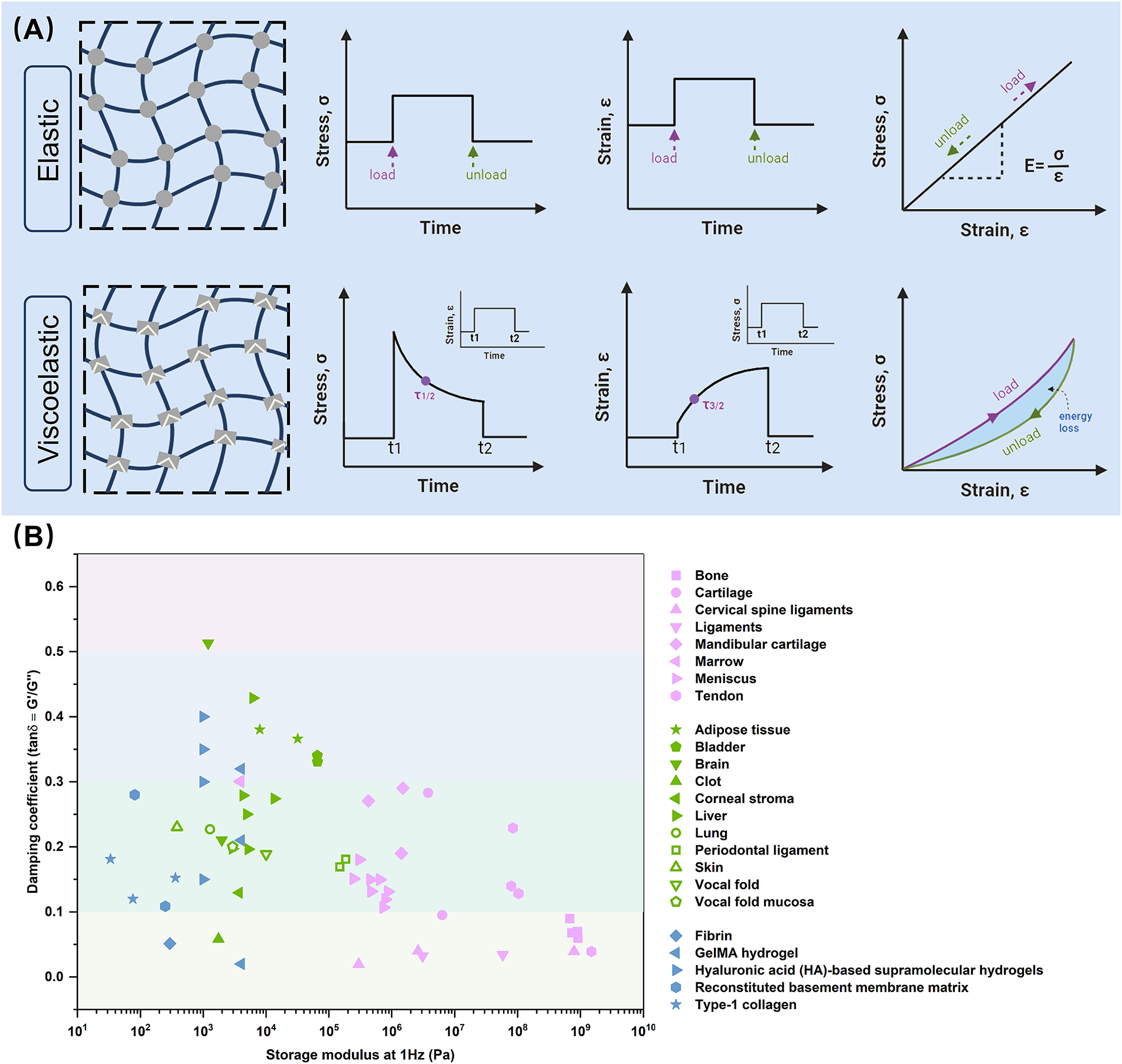
Mechanical behavior and characterization of materials and biological tissues.
From macroscopic tissues and organs to microscopic ECMs, viscoelasticity is a common property.12,47,48 For instance, the brain, 17 liver, 18 muscle, 19 and adipose tissue 20 exhibit higher viscoelasticity, while bone, 21 ligament, 22 tendon, 23 and skin 24 have lower viscoelasticity. The viscoelasticity of cartilage25,26 and vocal cord 27 falls in between (Fig. 1B). At the microscale, ECM viscoelasticity arises from polymer entanglements, structural protein folding, and reversible noncovalent interactions.45,49,50
Evaluation Indicators and Characterization Methods for Viscoelasticity
Viscoelasticity of biological tissues
The viscoelasticity of biological tissues plays a vital role in maintaining physiological activities. For example, the energy dissipation in viscoelastic connective tissue protects against continuous or periodic high stress or strain. Blood vessels endure millions of periodic mechanical loads, and viscoelasticity prolongs their fatigue life.51,52 Similarly, the viscoelasticity of the periodontal ligament dissipates energy during mastication, protecting tooth roots and alveolar bone from high masticatory forces.53,54 Bones are also viscoelastic, capable of withstanding long-term physiological loads. Furthermore, viscoelasticity changes often occur in specific pathological processes, such as lung and liver fibrosis 55 or tumor tissue formation.9,56
At the molecular level, the viscoelastic behavior arises from the dynamic and hierarchical organization of ECM components. 57 Fibrillar collagens contribute to ECM viscoelasticity through their hierarchical architecture. Time-dependent behaviors, such as stress relaxation and creep, result from fiber sliding and reversible dissociation of intrafibrillar and enzymatic cross-links.58,59 Glycosaminoglycans (GAGs), rich in anionic groups such as carboxylates and sulfates, attract and retain water, forming highly hydrated gel-like environments. This hydration not only provides compressive resistance but also affects ECM topology by influencing collagen spacing and network organization. 60 Proteoglycans, consisting of core proteins and GAG chains, form supramolecular assemblies that regulate ECM poroelasticity and viscoelasticity. Hydration-driven swelling, reversible GAG slippage, and core protein unfolding contribute to energy dissipation and mechanical adaptability.59,61 Collectively, these components form a dynamic, multiscale network that enables the ECM to store and dissipate mechanical energy, remodel in response to force, and transmit mechanical cues to resident cells.
Evaluation indicators
The commonly used indicators for viscoelasticity can be divided into three categories, corresponding to stress relaxation,62–64 creep,65–67 and energy dissipation68,69 of materials. First, the relaxation time τ1/2 or τ1/e represents the time required for stress to decay to 1/2 or 1/e under constant strain. 70 Similarly, the creep time (τ3/2) is the time required for a material to reach 150% initial strain under constant stress.65,71 The third category describes the stress–strain behavior under periodic stress. The storage modulus (G′) represents the energy stored in the material, while the loss modulus (G″) represents the energy dissipation related to its viscous behavior. The loss tangent (tanδ = G″/G′) expresses material viscoelasticity.69,72 A higher loss tangent indicates greater viscous behavior. The loss tangent of biological tissues typically ranges between 10% and 30%. 68 The half-life of stress relaxation in biological tissues varies from 10 to 1000 s. 70
Characterization methods
At present, various techniques are available to characterize material viscoelasticity. These include static and dynamic tests at macro- and microscales.73–75 Static mechanical tests are conducted by applying and measuring tensile, compressive, and shear stresses and strains, primarily through stress relaxation and creep tests.76–81 Dynamic mechanical tests include frequency-dependent rheological tests and cyclic loading tests.82–84 Recently, particle-based microrheological measurement methods have also emerged, enabling in situ detection of materials at micron or smaller length scales.85–87 Thus, it is possible to measure viscoelasticity within a material rather than only on its surface.85,88 In addition, elastography technologies,89–92 such as magnetic resonance elastography,90,93–95 shear-wave dispersion ultrasound vibrometry, 96 and shear-wave elastography, 97 have enabled completely noninvasive characterization of in vivo tissue viscoelasticity by detecting shear wave velocity in biological tissues (Fig. 2). 98
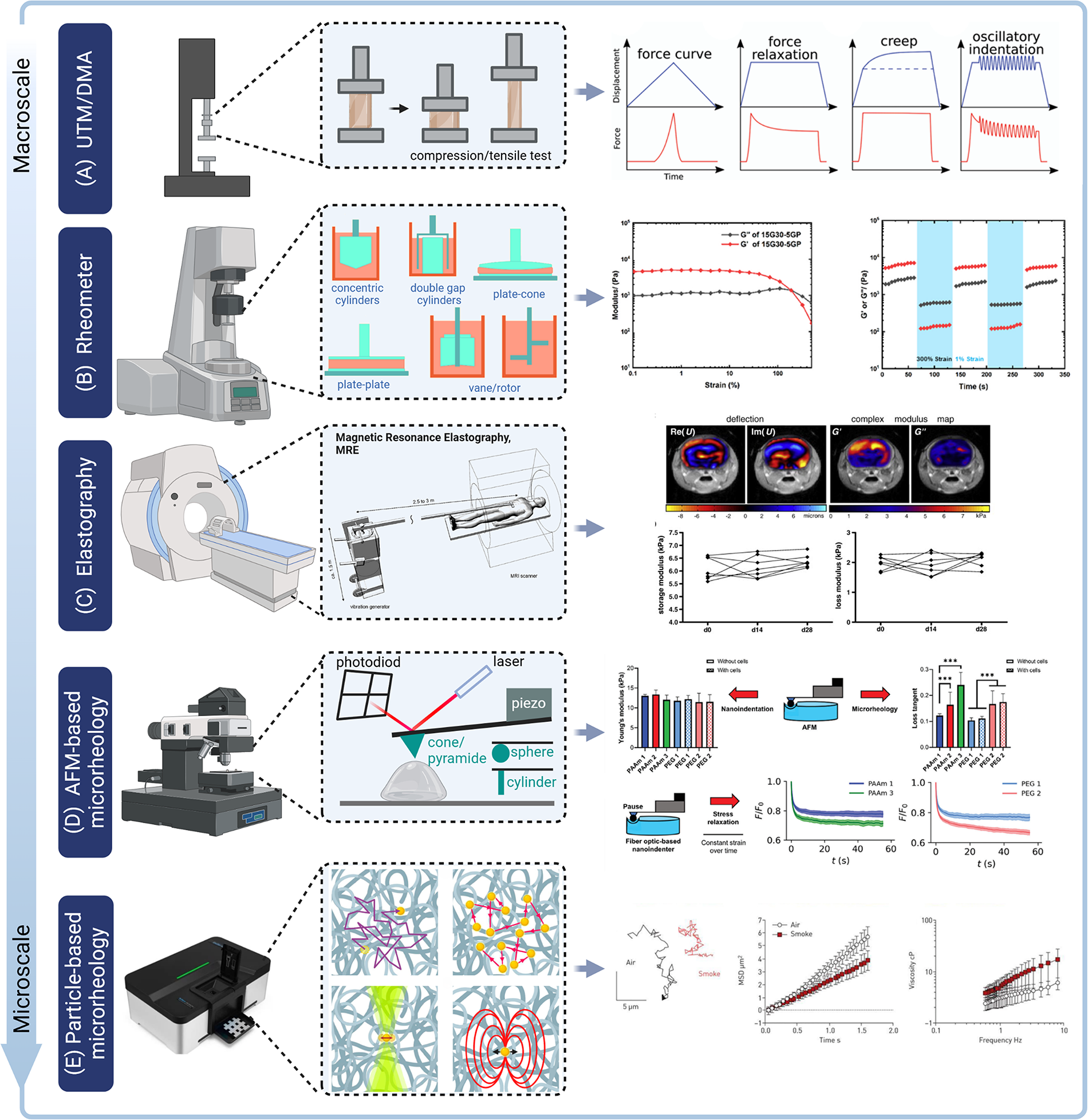
Multiscale mechanical characterization of biological tissues and materials.
Preparation Strategy of Viscoelasticity Adjustable Hydrogel
Preparation strategy of viscoelastic hydrogels
Hydrogels are hydrophilic polymer networks capable of absorbing and retaining water. 99 Common cross-linking methods include chemical and physical cross-linking. 100 Chemically cross-linking hydrogels typically have stable and permanent binding points. In contrast, physically cross-linked hydrogels contain reversible noncovalent interactions, forming dynamic networks with notable viscoelasticity.101–103 Reversible covalent bonds dynamically break and reform under certain conditions, imparting self-adaptability and viscoelastic properties. 50 Examples of reversible dynamic covalent bonds include borate ester bonds, 82 disulfide bonds, imine bonds, and acylhydrazone bonds.104,105
Meanwhile, viscoelasticity can also emerge from higher order structural features such as sliding cross-links, supramolecular assemblies, or interpenetrating networks, where energy dissipation results from topological rearrangements or multicomponent coupling rather than bond exchange alone. These strategies are orthogonal but often synergistic, and their integration provides versatile platforms for tailoring viscoelasticity in biomaterial design (Table 1).
Comparative Strategies for Introducing Viscoelasticity into Hydrogels

sGAG, sulfated glycosaminoglycan; UPy, ureidopyrimidinone; CD–ADA, cyclodextrin–adamantane; G′, storage modulus; G″, loss modulus.
Strategies for controlling viscoelasticity of hydrogels
Previously, researchers used various strategies to control the viscoelastic properties of hydrogels (Fig. 3).120–124 It is important to note that some methods only affect the viscoelastic parameters, while others may alter additional mechanical properties (such as elastic modulus) or even the three-dimensional (3D) structure (e.g., pore size, porosity).104,110,125 Therefore, it is crucial to select appropriate methods for specific applications.126–129
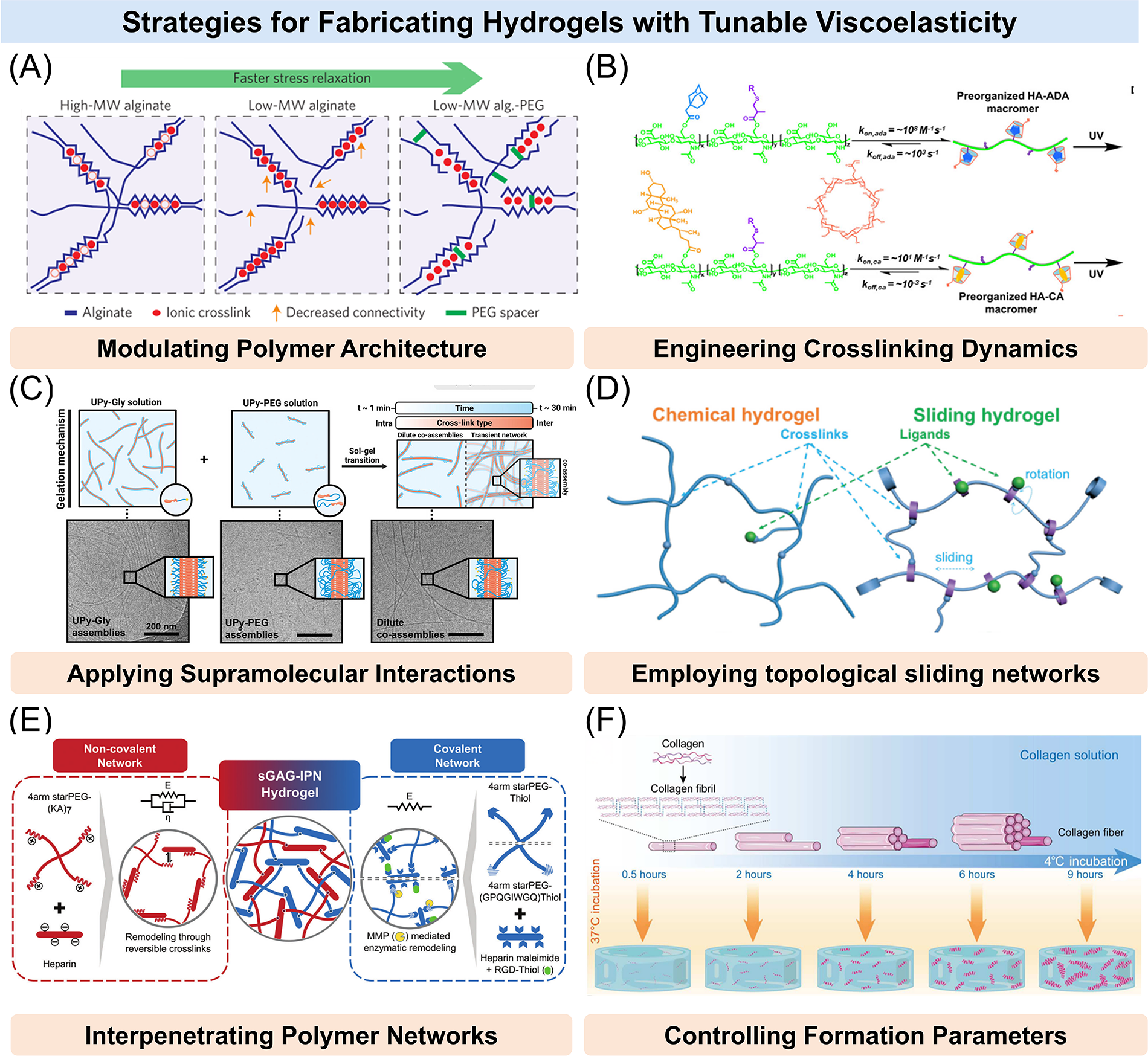
Strategies for fabricating hydrogels with tunable viscoelasticity.
Controlling viscoelasticity by changing polymer structure
Viscoelastic properties of hydrogels can be modulated by tailoring the molecular structure of the polymer building blocks. Specifically, molecular weight, chain flexibility, and block architecture significantly influence stress relaxation behavior and energy dissipation capacity. 130 Chaudhuri et al. applied different molecular weight alginates with varying cross-linking densities of calcium to modulate the stress relaxation properties of the hydrogels by altering connectivity and chain mobility in the network. A decrease in the initial elastic modulus resulting from decreased polymer molecular weight could be compensated for by increased cross-linking. Also, the covalent coupling of short polyethylene glycol (PEG) spacers to the alginate provided a steric hindrance to cross-linking of alginate chains, enhancing stress relaxation in the gel (Fig. 3A).63,131 In a related approach, Walker et al. produced PEG-maleimide (MAL) hydrogels with similar initial elastic moduli but different viscoelasticities by varying the molecular weight of PEG. 69 Furthermore, Jung et al. tuned the viscoelasticity of PEO-based ABA triblock copolymer hydrogels by varying hydrophobic end-block length, with longer blocks increasing relaxation time. 112 Notably, Hafeez et al. independently tuned hydrogel viscoelasticity by altering the hydrophobic length of end-groups (Fig. 3B). 132 Using a distinct strategy, Peng et al. used programmable DNA sequences with varying lengths to adjust hydrogel stress relaxation by altering DNA chain overlap. 111 These studies demonstrate that polymer chain architecture and hydrophobic segment design can offer predictable and tunable control over hydrogel viscoelasticity.
Controlling viscoelasticity by adjusting cross-linking method or density
Beyond polymer architecture, the viscoelastic behavior of hydrogels can be finely tuned by altering the mode and density of cross-linking.133,134 Chaudhuri et al. ionically cross-linked Arg-Gly-Asp peptide (RGD)-alginate with Ca2+ to obtain a soft viscoelastic hydrogel and, in parallel, covalently cross-linked RGD-alginate with acyl-hydrazide cross-linking to yield an elastic hydrogel. By adjusting cross-linker concentrations, both gels shared the same initial stiffness. 135 Similarly, Narita et al. found that polyvinyl alcohol hydrogels prepared using different cross-linking methods exhibited variations in viscoelasticity (Fig. 3C). 136 Regarding dynamic cross-linking, McKinnon et al. prepared multiarm PEG hydrogels based on reversible dynamic acylhydrazone bond cross-linking. Hydrogels with similar initial elastic moduli but different viscoelastic properties were developed by adjusting the proportion of two acylhydrazone bonds. 113 Likewise, Yang et al. precisely controlled the viscoelasticity of hyaluronic acid (HA) hydrogels by adjusting the proportions of reversible supramolecular cross-links with different kinetic constants in the cross-linking bond (Fig. 3D). 118 Meanwhile, Chrisnandy et al. developed a dynamically cross-linked hydrogel system with reversible cytosine–cytosine triple hydrogen bonding. By tuning the proportion of physical cross-links, they precisely controlled the percentage of stress relaxation. 137 Together, these findings highlight that both the chemistry and kinetics of cross-link formation can determine the viscoelastic response of hydrogels.
Controlling viscoelasticity via supramolecular interactions
Specific hydrogel systems harness noncovalent supramolecular motifs, such as self-complementary hydrogen bonding, 117 host–guest interactions, 118 π–π stacking, or metal coordination, to form physically cross-linked and dynamically reconfigurable networks. For instance, ureidopyrimidinone (UPy)-based supramolecular hydrogels self-assemble into nanofibrous networks through quadruple hydrogen bonding, and their viscoelastic behavior can be finely tuned by adjusting the ratio of mono- and bifunctional UPy monomers, as well as the density of RGD-functionalized UPy motifs. This approach offers a versatile platform for dynamically controlling matrix mechanics and mechanotransduction in bioengineered microenvironments. 117
Controlling viscoelasticity by topological sliding networks
Sliding hydrogels leverage topologically interlocked architectures, such as polyrotaxanes composed of cyclic molecules (e.g., α-cyclodextrins) threaded onto linear polymer chains (e.g., PEG), to introduce locally mobile yet topologically fixed cross-linking points. These movable cross-links and ligands endow the hydrogel with dynamic mechanical adaptability while maintaining overall structural integrity. For example, Tong and Yang et al. developed a sliding hydrogel based on polyrotaxane structures in which α-cyclodextrins were threaded onto PEG chains and capped at both ends to prevent unthreading. 115 The cross-linkers and RGD ligands attached to the α-cyclodextrins were thus free to move along the polymer backbone. Rheological characterization revealed that increased mobility of the sliding units significantly enhanced stress relaxation without altering the initial elastic modulus.
Controlling viscoelasticity using interpenetrating polymer networks
Interpenetrating polymer networks (IPNs), formed by combining covalent and dynamic or degradable networks, offer a strategy to modulate hydrogel viscoelasticity. By adjusting the composition or cross-linking of each network, IPNs enable independent control of stress relaxation without altering initial stiffness. By constructing a semi-interpenetrating polymer network (sGAG-IPN) hydrogel composed of a covalently cross-linked PEG-MAL network and a noncovalently assembled matrix metalloproteinase (MMP)-degradable sGAG-like network, the overall viscoelastic properties of the hydrogel could be precisely tuned by adjusting the molar ratio between the covalent and noncovalent components. 116 By combining covalent and supramolecular cross-links, Loebel et al. designed a double-network hydrogel comprising a covalent PEG-fibrinogen network and a β-cyclodextrin adamantane supramolecular network, which exhibited independent control of viscosity and elasticity. 138
Controlling viscoelasticity by changing the conditions of hydrogel formation
Environmental factors during hydrogel formation, such as temperature, pH, and reaction time, can also influence polymer assembly and network configuration, thereby affecting viscoelasticity. Huang et al. extended low-temperature incubation of the collagen solution, which resulted in more weak interactions within the collagen fiber network, and increased diameter of collagen fibers. Self-assembly changes of collagen fibers led to different stress relaxation behaviors (Fig. 3E). 67 In addition, Kaewprasit et al. found that the addition of monohydric alcohol enhanced the formation of self-assembled β-fold structures by increasing hydrophobic interactions with silk fibroin chains, significantly improving the mechanical properties and viscoelasticity of the hydrogels. 139 Under pH-responsive conditions, Jay et al. developed a reversible covalent hydrogel using dynamic cross-linking between boronic acid and diol groups for vaginal applications. The hydrogel rapidly reformed at vaginal pH (4.5–5), leading to a viscoelastic response but stabilized at pH >5.5 (Fig. 3F). 114 Moheimani et al. regulated the viscoelasticity of alginate hydrogels through repeated high-pressure sterilization. With increasing sterilization cycles, the molecular weight of the alginate decreased significantly, thereby modulating the hydrogel’s mechanical properties. 140 These studies underscore that even without modifying polymer composition or cross-linking chemistry, process conditions alone can be leveraged to dynamically tune viscoelasticity.
Effect of Viscoelastic Matrix on Cell Behavior
The ECM is not only a structural network but also provides 3D biochemical and biophysical cues that regulate cellular behavior in development, physiology, and pathophysiology.141–144 Increasing evidence has shown that biophysical cues significantly affect the behavior and fate of cells.145,146 Previous studies have focused primarily on ECM stiffness.11,147,148 Recent research has found that the mechanotransduction response of cells to their surrounding environment may play a key role in determining the fate of stem cells. 128 Controlling the viscoelasticity can guide the differentiation of specific cell pedigrees in tissue engineering,149–152 as well as regulate other cell behaviors (Fig. 4).153–155
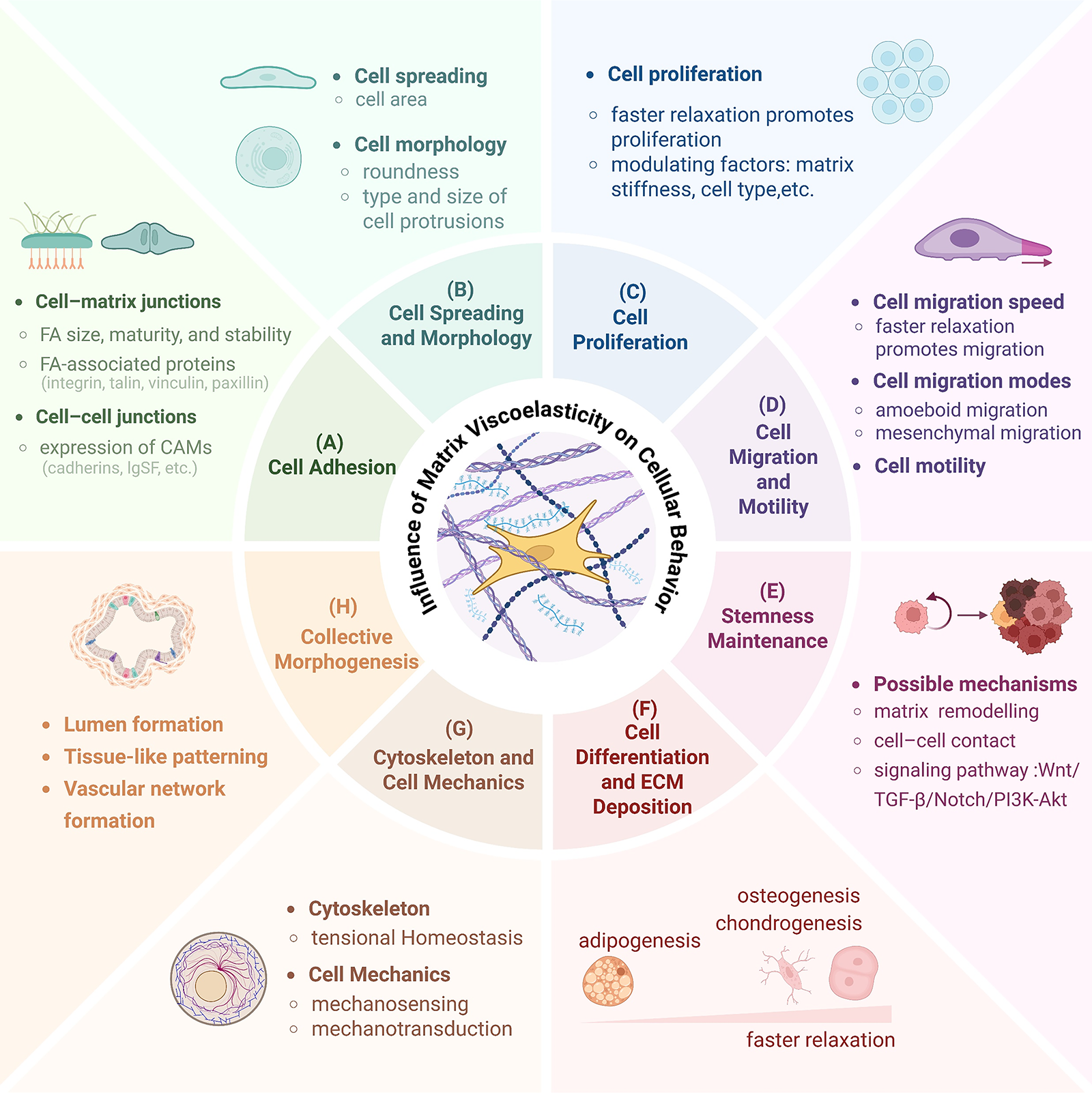
Matrix viscoelasticity regulates a wide array of cellular functions.
Cell adhesion
Cell adhesion, encompassing both cell–matrix and cell–cell interactions, plays a vital role in regulating stem cell behavior, including spreading, migration, and differentiation.156–158 Focal adhesions (FAs) act as hubs of mechanical and biological signals, essential for cell-ECM mechanical sensitivity, while cadherin-mediated adhesions regulate intercellular cohesion and tissue organization (Fig. 5A, B).160,161 In type-I collagen hydrogels with fast stress relaxation, the FAs of mesenchymal stem cells (MSCs) displayed a more mature structure. 150 Similar results were observed in a 3D culture of human MSCs (hMSCs). 119 In contrast, FA size and number differed significantly when hMSCs were cultured on polyacrylamide hydrogels. Specifically, FA maturation was reduced on substrates with higher tanδ, while cell aggregation was enhanced in 3D PEG-MAL hydrogels under the same conditions, promoting mesenchymal condensation and chondrogenic differentiation (Fig. 5C). 69 Cameron et al. reported similar findings. 65 Adebowale et al. also concluded that cell migration was associated with fewer and weaker paxillin adhesions on matrices with fast relaxation. 70
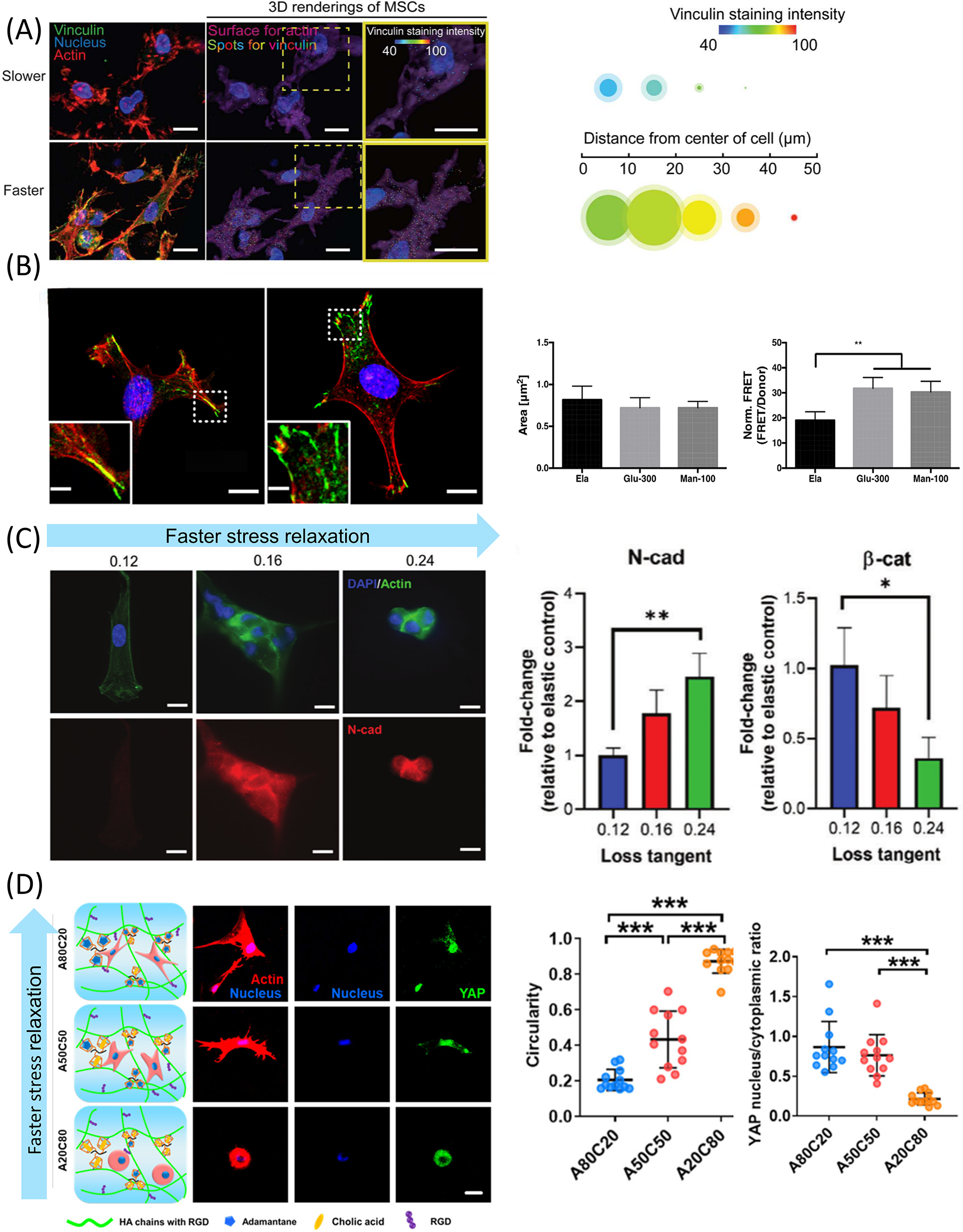
Impact of matrix viscoelasticity on cell–matrix adhesion, cell–cell adhesion, cytoskeletal organization, and cell spreading and morphology.
Cell spreading and cell morphology
Most previous studies have shown that matrices with high stiffness can enhance cell traction forces and promote cell spreading to maintain tension balance. 162 However, the effect of viscoelasticity on cell spreading and morphology remains uncertain. Cameron et al. found that a high loss modulus in the matrix increased the spreading area of hMSCs. 65 Chaudhuri et al. observed that U2OS cells and 3T3 mouse fibroblasts exhibited larger spreading areas and more stress fiber formation on substrates with stress relaxation. 135 A similar phenomenon was observed in mouse MSCs. 63 Lou et al. identified a linear relationship between matrix relaxation rate and cell roundness, with cells in viscoelastic hydrogels being less circular and developing longer protrusions. 119 However, Mandal et al. reported smaller spreading areas in primary human hepatocytes (PHHs) and fibroblasts, but larger areas and faster spreading rates in hepatocellular carcinoma cells (Huh7) on viscoelastic substrates. The opposite responses of PHHs and Huh7 cells to viscoelasticity may be explained by differences in the characteristic lifetimes of substrate binding between liver cancer and normal cells, which could lead to altered cell spreading and motility within diseased tissue. 163 In addition, Gong et al. found that viscosity promoted cell spreading at low long-term stiffness—defined as the residual equilibrium modulus after stress relaxation—with maximum spreading achieved at medium viscosity (Fig. 5D). 164
Based on the motor-clutch model combined with a standard linear viscoelastic substrate, the differential cellular responses to substrate viscoelasticity are primarily determined by whether the cell possesses clutch reinforcement mechanisms, as well as its traction force generation, clutch density and binding kinetics, sensitivity to mechanical feedback, and FA lifetime. Cells with strong traction forces, stable FAs, and reinforcement capacity, such as fibroblasts, exhibit saturated spreading on stiff substrates and show limited sensitivity to viscosity. In contrast, cells lacking reinforcement mechanisms, with weaker traction forces or short-lived FAs—such as neurons—rely more heavily on substrate viscoelastic matching to sustain effective spreading.11,164,165
Cell proliferation
Cell proliferation is influenced by microenvironmental physical properties.166,167 Alginate hydrogels with fast stress relaxation promote proliferation of C2C12 myoblasts and fibroblasts.62,63 Liu et al. found that the proliferation and migration of human bone marrow MSCs (hBMSCs) were enhanced as the loss modulus increased. 68 Lee et al. found that fast stress-relaxing hydrogels promoted chondrocyte proliferation while inhibiting cell death. 106 Similarly, viscoelastic scaffolds supported 1.67-fold higher chondrocyte proliferation compared with unmodified scaffolds after 3 days. 168 Indana et al. revealed that a higher RGD density and faster stress relaxation promoted human induced pluripotent stem cell (hiPSC) activity, proliferation, and lumen formation (Fig. 6C). 107
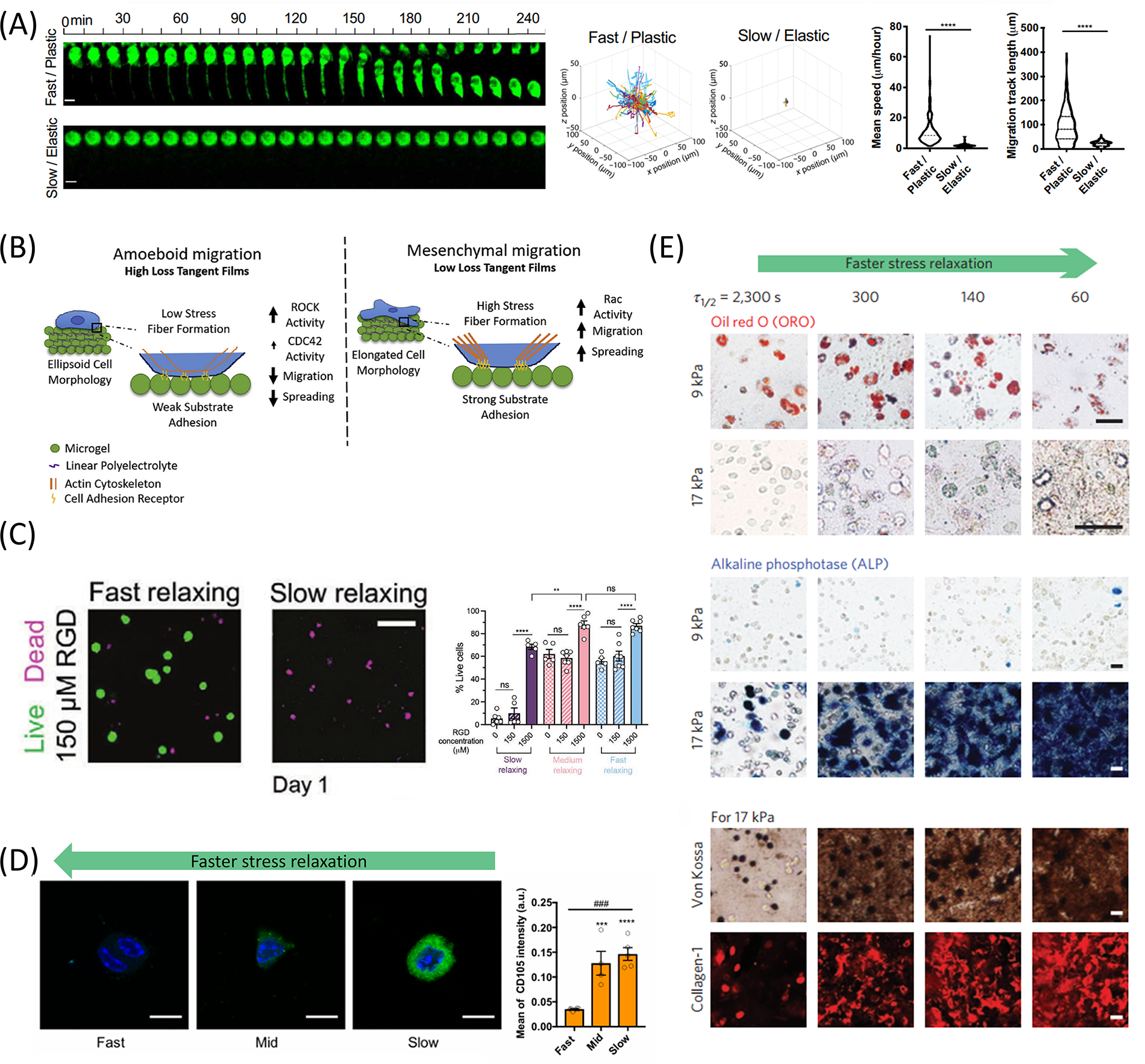
Impact of matrix viscoelasticity on cell migration, proliferation, stemness maintenance, and differentiation.
Cell migration
The cell–matrix connection and the formation of cell movement-related structures are two major factors determining cell migration. 172 ECM stress relaxation regulates cell migration speed and mode (Fig. 6A).173,174 Darnell et al. observed nearly doubled migration distance over a week on fast-relaxing alginate hydrogels compared with slow-relaxing gels. 64 Adebowale et al. showed minimal cell migration on 2 kPa substrates with slow relaxation but significantly increased migration on substrates with faster relaxation. 70 Matrix mechanical properties also influence migration modes. On soft viscoelastic substrates with fast stress relaxation, cells adopted a rounded morphology and migrated using filopodia-like protrusions, forming only nascent peripheral adhesions. In contrast, on purely elastic substrates of similar stiffness, cells spread more extensively, exhibited lamellipodia-driven migration, and developed larger, more stable FAs. 70 Chester et al. demonstrated that viscoelasticity influences migration mode: a high loss tangent promotes amoeboid migration, while a low loss tangent supports mesenchymal migration. Signaling molecules such as Rho-associated kinase (ROCK) and Ras-related C3 botulinum toxin substrate (Rac) play central roles in this process—ROCK enhances actomyosin contractility and cortical tension, supporting rounded, weakly adherent amoeboid migration, whereas Rac facilitates lamellipodia formation, FA maturation, and polarization, enabling mesenchymal migration with elongated morphology and directional motility (Fig. 6B). 170
Stemness maintenance
Stemness maintenance is also a critical feature of stem cells in regenerative medicine applications.175–177 Appropriate mechanical signals can promote stem cell self-renewal, while excessive mechanical stress may induce differentiation.178–180 Madl et al. found that inhibiting physical remodeling by adding covalent cross-linking to alginate materials reduced neural progenitor cell stemness. 178 However, Lee et al. found that MSCs exhibited a higher expression level of CD105 in hydrogels with slow relaxation rates (Fig. 6D). 171 Lin et al. found that slow stress relaxation in the hydrogel could induce MSCs to enter a reversible resting G0 state, suggesting that MSC quiescence may help preserve stemness when exposed to low-deformation matrices. 181
Cell differentiation and ECM deposition
The process of cell differentiation and ECM deposition is closely dependent on the microenvironment (Fig. 6E).182–186 Increasing matrix loss modulus has been shown to enhance osteogenic and chondrogenic differentiation while suppressing adipogenesis. 68 ,106,171 In contrast, hydrogels exhibiting slower stress relaxation fail to sustain early chondrogenic advantages over time, whereas those with faster relaxation kinetics promote long-term cartilage matrix deposition by MSCs. 67 In addition, the effectiveness of chondrogenic induction media appears contingent upon the mechanical context: while high-elasticity environments boost cartilage matrix deposition, this effect diminishes in high tanδ hydrogels, suggesting a saturable cellular response governed by mechanical cues alone. 69
Multicellular aggregates and collective morphogenesis
In addition to studies focused on single-cell behaviors, an emerging body of work investigates how viscoelasticity regulates cell behavior at the level of multicellular aggregates, bridging the gap between single-cell mechanobiology and complex organoid systems. These models exhibit collective behaviors such as aggregation, lumen formation, and tissue-like organization, and they undergo coordinated morphogenetic processes that often mimic early developmental events.
In a viscoelastic and degradable IPN hydrogel system, high viscoelasticity combined with proteolytic degradability most effectively promoted lumen formation, growth expansion, and maintenance of pluripotency in hiPSC-derived cysts, accompanied by increased YAP nuclear localization and enhanced secretion of ECM proteins. 116 In a two-phase system combining hypoxic preconditioning and viscoelastic hydrogel encapsulation, hypoxia first induced endothelial progenitor cell (EPC) aggregation into clusters, and the subsequent exposure to high-viscoelasticity matrices specifically promoted vasculogenesis by enhancing sprouting, branching, and network formation. 187 In 3D alginate hydrogels with independently tunable viscoelasticity and RGD ligand density, fast stress relaxation combined with high RGD density promoted hiPSC viability, proliferation, apicobasal polarization, and lumen formation, whereas slow relaxation at low RGD density led to apoptosis and impaired morphogenesis, highlighting matrix viscoelasticity as a key regulator of collective morphogenesis. 107
Mechanism of Viscoelastic Mechanotransduction
Mechanobiology involves (1) mechanical sensing, the ability of cells to perceive biomechanical cues from the surrounding microenvironment, and (2) mechanotransduction, the ability of cells to convert external forces into biochemical signals that trigger specific cellular functions.188–190 Current research on viscoelastic mechanical signal sensing focuses on two main categories of membrane proteins: (1) Integrin pathway: integrin is a transmembrane protein that interacts with various proteins, including FA kinase (FAK), within the FA complex. The FA complex connects with the actin cytoskeleton to form a mechanical link and transmits external physical signals into the cell; (2) Mechanosensitive ion channel-mediated pathways: mechanosensitive ion channels108,191–194 respond to external mechanical forces by opening or closing, thereby regulating the flow of ions inside and outside the cell, which subsequently triggers a series of downstream signaling pathways (Fig. 7A).195–198
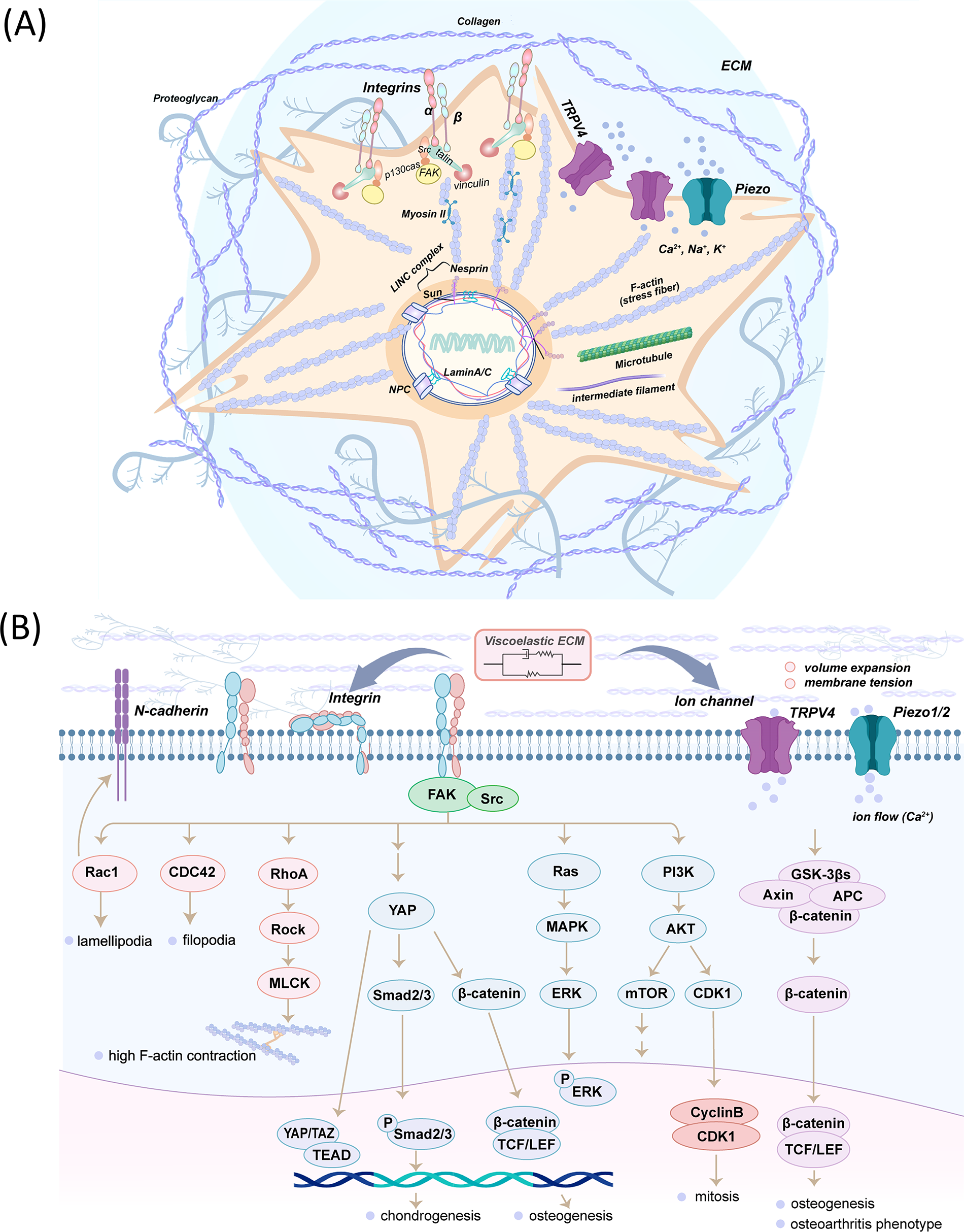
Schematic of viscoelastic mechanotransduction: Cellular structures and signaling pathways.
Integrin-mediated mechanotransduction
During mechanotransduction, mechanical stimulation can alter the function of mechanosensitive proteins within the cell adhesion structure, triggering biochemical signals that regulate both the rapid mechanical response of cells and long-term changes in gene expression.199,200 These pathways can be broadly divided into two categories: (1) Cytoskeletal regulators, including the FAK-Src complex and Rho-family small GTPases such as RhoA/ROCK, Rac1, and CDC42, control actin polymerization, contractility, and FA dynamics, thereby modulating cell tension, shape, and migration; and (2) transcriptional regulators, including YAP/TAZ, β-catenin, Smad2/3, ERK, and CyclinB/CDK1, are activated downstream of mechanosensitive signaling networks such as integrin–FAK–Src, Ras–MAPK, or PI3K–Akt pathways. These molecules translocate into the nucleus where they regulate gene expression governing cell fate.133,172,201–207
Integrin signaling facilitates YAP nuclear translocation, along with Smad2/3 and β-catenin, thereby promoting chondrogenesis and osteogenesis. 68 Furthermore, integrin–FAK-mediated PI3K–Akt–CDK1 signaling in fast-relaxing hydrogels upregulates MSC proliferation. 181 Meanwhile, small GTPases play an important role in integrin-mediated mechanotransduction. Integrin β1 activates FAK–RhoA–ROCK–MLCK signaling, enhancing actomyosin contractility and promoting MSC spreading on stress-relaxing substrates. 135 High-creep hydrogels increase Rac1 activity in hMSCs and upregulate N-cadherin expression, thereby promoting smooth muscle cell differentiation. 71 In contrast, in mouse MSCs, high matrix viscosity suppresses ROCK and Rac signaling, reduces β1 integrin and β-catenin activation, while enhancing N-cadherin-mediated adhesion, suggesting cell-type-dependent and context-specific roles of viscoelasticity. 69
Ion channel protein-mediated mechanotransduction
Mechanosensitive ion channels are directly activated by stress exerted on lipid bilayers, converting physical stimuli into electrical signals and responding to mechanical stress. 197 Cell volume expansion can occur in biodegradable hydrogels or hydrogels with fast stress relaxation, but it is limited in more restrictive elastic hydrogels. 171 Therefore, cell volume regulation may mediate viscoelastic mechanotransduction of the ECM.172,208,209 Among mechanosensitive ion channels, TRPV4 channels have emerged as key sensors of cell volume changes.208,209 Feedback between TRPV4 activation and volume expansion has been shown to regulate the nuclear localization of RUNX2, thereby promoting osteogenic differentiation. 171 In pathological conditions such as osteoarthritis, dysfunction of the TRPV4–GSK3β signaling axis impairs the cell’s ability to sense and respond to viscoelastic cues from the surrounding matrix, resulting in persistently elevated intracellular calcium levels (Fig. 7B). 191
Computational Biology Research on Viscoelasticity-Mediated Cell Behavior
Computational biology research can complement, analyze, and interpret experimental data in the field of cell mechanics. In addition, it can provide valuable insights for designing mechanical parameters of viscoelastic materials. Researchers have proposed numerous models, including the power-law model,210,211 fractional model, 212 and the viscoelastic and active viscoelastic model (Fig. 8A).214,215 To study cell adhesion and migration, mechanical models70,135,164,216,217 were developed to understand molecular-level cellular responses to viscoelastic microenvironments. Furthermore, finite element analysis (FEA) can study the complex cell-ECM mechanical systems.150,181
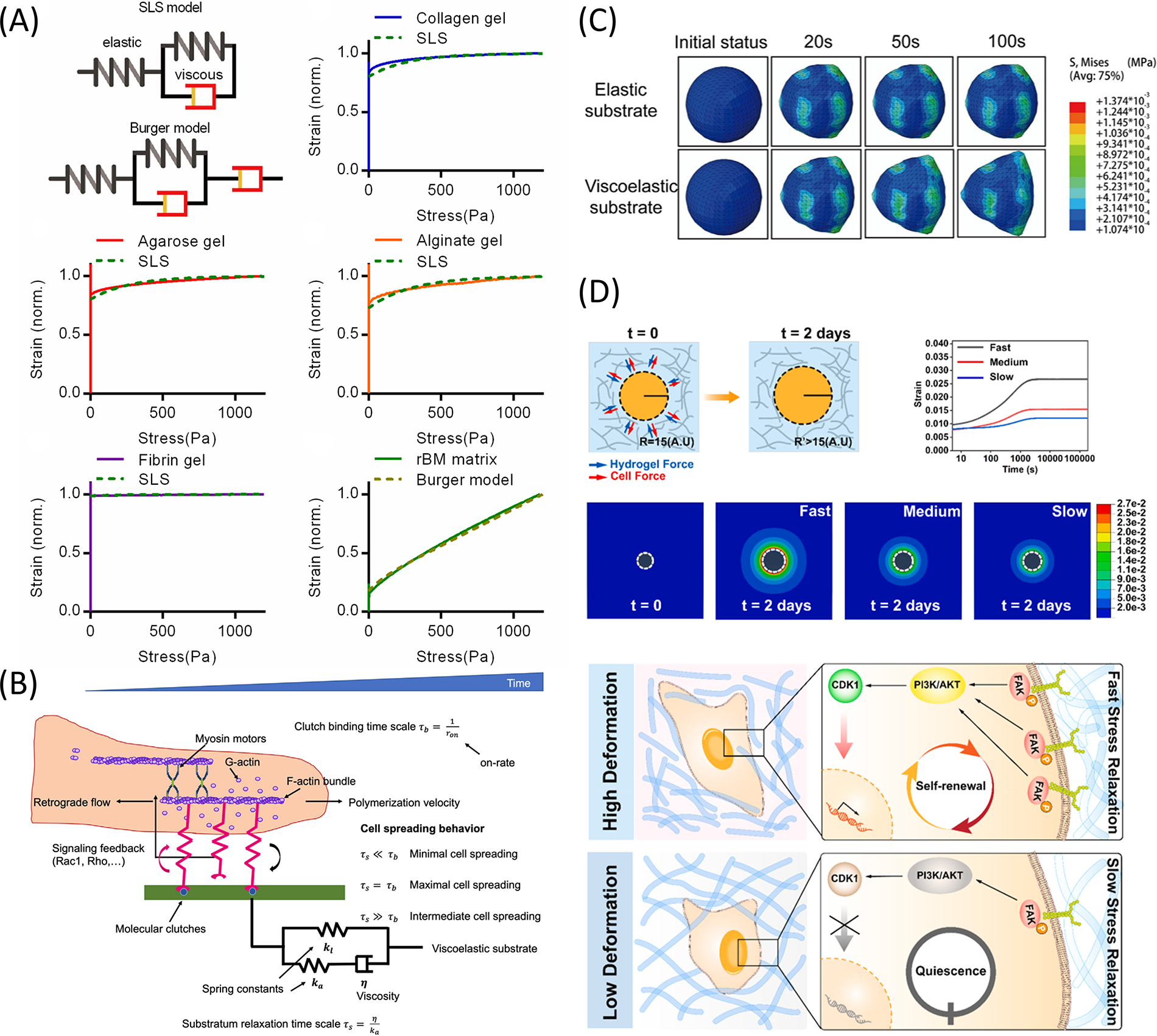
Computational biology insights into motor-clutch models and finite element analysis of matrix viscoelasticity in cell behavior.
Mechanical model of cell adhesion
Chan and Odde introduced the motor-clutch model of cell adhesion. Cells can detect and respond to the biophysical properties of the extracellular environment through integrin-based adhesion points (Fig. 8B). These adhesion points connect the ECM to the F-actin cytoskeleton, transmitting the mechanical forces generated by actin retrograde flow and myosin II to the ECM.45,218–220 Adebowale et al. conducted two simulations using the motor-clutch model. The results of both were consistent with experimental data, demonstrating how stress relaxation and matrix stiffness affect cell behaviors. 70 Using the motor-clutch and linear viscoelastic models, Gong et al. found that medium viscosity maximized proliferation on soft matrices but had minimal effect on stiff ones. 164
3D finite element model analysis
FEA can simulate and analyze the behavior of complex physical systems.221–223 It can be used in cell adhesion research, cell migration simulation, and tissue engineering design and optimization.224,225 Huang et al. used FEA to predict the response of cells to external stress. In the viscoelastic matrix, stress distribution and cell deformation evolve to adapt to matrix deformation, promoting the dynamic expansion of cells (Fig. 8C). 150 Lin et al. showed that fast matrix relaxation induced significant deformation around MSCs, activating PI3K/Akt-CDK1 pathways to accelerate cell cycle progression (Fig. 8D). 181
Application of Viscoelastic Hydrogels in Regenerative Medicine
Viscoelastic biomaterials have long been used in regenerative medicine.226–230 These materials have been shown to promote the regeneration of various tissues, including cartilage and bone.231–235 The most commonly used recombinant basement membrane materials for organoid culture are also viscoelastic. 236 However, only a few studies have been conducted on hydrogels with adjustable viscoelasticity that control viscoelasticity as an independent variable (Table 2).
Independent Tuning of Viscoelasticity in Various Hydrogel Systems and Its Effects on Cell Behavior
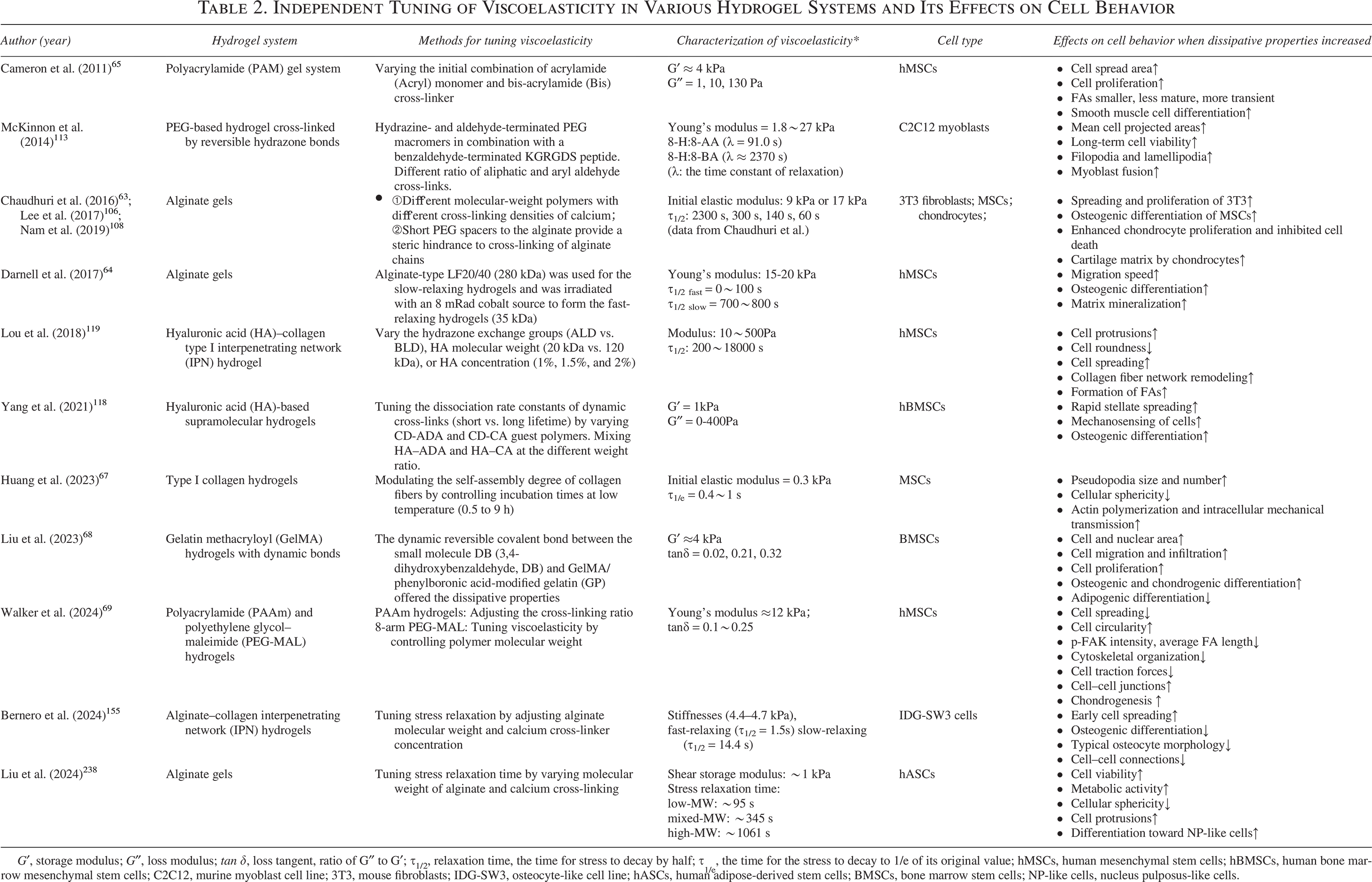
G′, storage modulus; G″, loss modulus; tan δ, loss tangent, ratio of G″ to G′; τ1/2, relaxation time, the time for stress to decay by half; τ1/e, the time for the stress to decay to 1/e of its original value; hMSCs, human mesenchymal stem cells; hBMSCs, human bone marrow mesenchymal stem cells; C2C12, murine myoblast cell line; 3T3, mouse fibroblasts; IDG-SW3, osteocyte-like cell line; hASCs, human adipose-derived stem cells; BMSCs, bone marrow stem cells; NP-like cells, nucleus pulposus-like cells.
Application on tissue engineering scaffolds
Currently, research on the effect of viscoelastic matrices on cell behavior mainly focuses on 2D or 3D in vitro cell cultures or organoid models.14,107,109 Few studies have explored the application of hydrogel materials with adjustable viscoelasticity for drug delivery, cell delivery and transplantation, or tissue engineering.46,150,167,237 Hydrogels with independently adjustable viscoelasticity have been used to regulate adipose-derived MSCs (ASCs) for nucleus pulposus regeneration, showing that fast stress-relaxing hydrogels significantly increased the long-term survival rate of ASCs, as well as the secretion of nucleus pulposus (NP)-like ECM proteoglycan and type II collagen. However, this study was limited to 3D cell culture in vitro. 238 In vivo studies have demonstrated that fast stress-relaxing alginate hydrogels loaded with hMSCs can significantly enhance new bone formation in calvarial defect models. 64 Similarly, GelMA-based hydrogels with tunable viscoelasticity have been applied to femoral trochlear defects, where higher viscous dissipation correlated with improved cartilage and bone regeneration. 68 Lin et al. found that a hydrogel with the fastest stress relaxation delivered the largest area and volume of new periodontal ligament. 239 Zhang et al. demonstrated that the viscoelasticity of Alg-PBA/Spd hydrogels could cooperate with biochemical factors to achieve optimal periodontal ligament repair (Fig. 9). 240
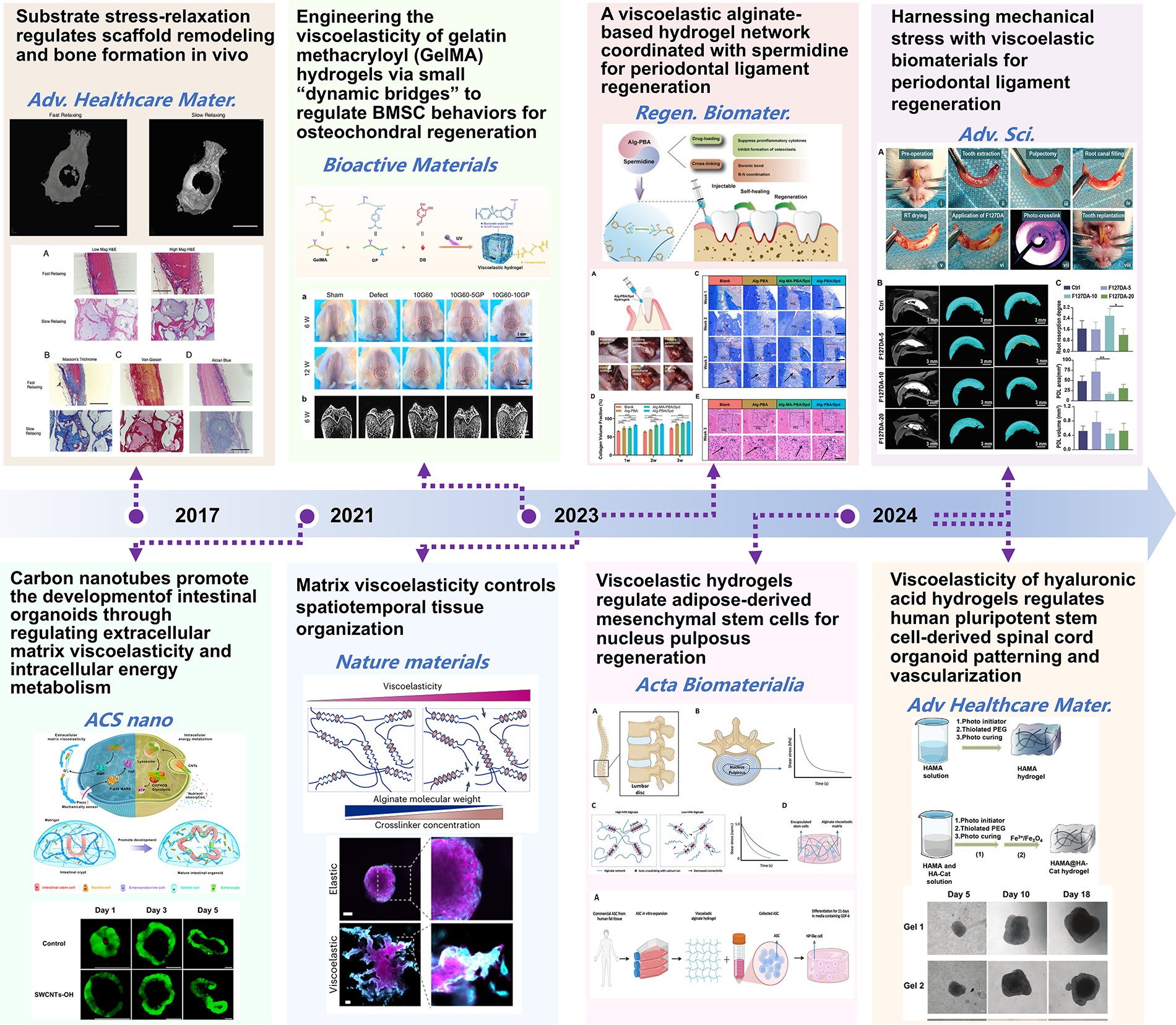
Chronological representation of key studies illustrating the application of independently tunable viscoelastic hydrogels in regenerative medicine. These studies highlight the evolution of viscoelastic materials in both tissue engineering and organoid culture. These findings emphasize the role of tuning viscoelastic properties to regulate cell behavior, tissue formation, and matrix remodeling, driving innovation in regenerative medicine across various applications.14,64,68,109,238–241
Application on organoid culture system
Recent research about organoids has expanded rapidly.242,243 Elosegui-Artola et al. found that matrix viscoelasticity could induce morphological instability in cell spheroids, leading to symmetry disruption, the formation of finger-like protrusions, and epithelial–mesenchymal transition. 244 In addition, the incorporation of carbon nanotubes promoted the proliferation and differentiation of intestinal organoids by modulating the viscoelasticity of the ECM and activating the Piezo-p38-YAP signaling pathway. 109 Studies also indicate that the viscoelasticity of HA hydrogel played a key role in the patterning and differentiation of stem cell-derived organoids. Viscoelasticity regulated cell–matrix mechanical signal transduction more effectively and promoted the expression of dorsal and interneuron markers more efficiently than traditional elastic properties. It also exhibited significant advantages in the vascularization of organoids, particularly in promoting the formation of the blood–spinal cord barrier. 241
Chrisnandy et al. demonstrated that their stress-relaxing synthetic hydrogel supports degradation-independent growth and morphogenesis of intestinal organoids. Compared with nondegradable elastic gels, this matrix significantly enhanced symmetry breaking and crypt-like structure formation, promoting spatial differentiation of stem/progenitor cells and Paneth cells. 137 van Sprang et al. developed an injectable UPy-based supramolecular hydrogel to encapsulate hiPSC-derived kidney organoids. The hydrogel’s tunable viscoelasticity and cell-adhesive properties contributed to modulating nephron patterning during the S-shaped body stage, leading to an increased proportion of WT1+ glomerular lineage cells and upregulated vascular endothelial growth factor (VEGF) expression. 117
Current Challenges and Future Directions
Currently, the importance of viscoelasticity as a design parameter in tissue engineering biomaterials has been increasingly recognized. Despite these advances, several challenges remain that may hinder clinical translation:
Trade-offs between viscoelastic tunability and biocompatibility: Covalent cross-linking often requires initiators or cross-linkers with potential cytotoxicity; degradation products of synthetic polymers may elicit adverse cellular responses.65,68 In addition, commonly used inert materials lack cell-adhesive motifs, necessitating further biochemical functionalization.107,113,115,117 Mechanical limitations: Although physically cross-linked systems exhibit favorable viscoelastic profiles, their inherently weak mechanical strength restricts their use in mechanically demanding scenarios.110,111,117 Environmental constraints: Viscoelasticity tuning via dynamic covalent bonds is often highly dependent on specific physicochemical conditions.113,114 For example, hydrazone bonds exhibit dynamic behavior predominantly under mildly acidic environments (pH 4.0–6.0), whereas under physiological pH, bond exchange is retarded.114,130 Complexity and scalability of advanced network designs: Cutting-edge strategies such as topological sliding networks
115
or supramolecular assemblies116–118
offer exquisite control over stress relaxation behavior, yet they often require complex molecular design, multistep synthesis, and sophisticated fabrication processes.
Future research may focus on the following directions (Fig. 10):
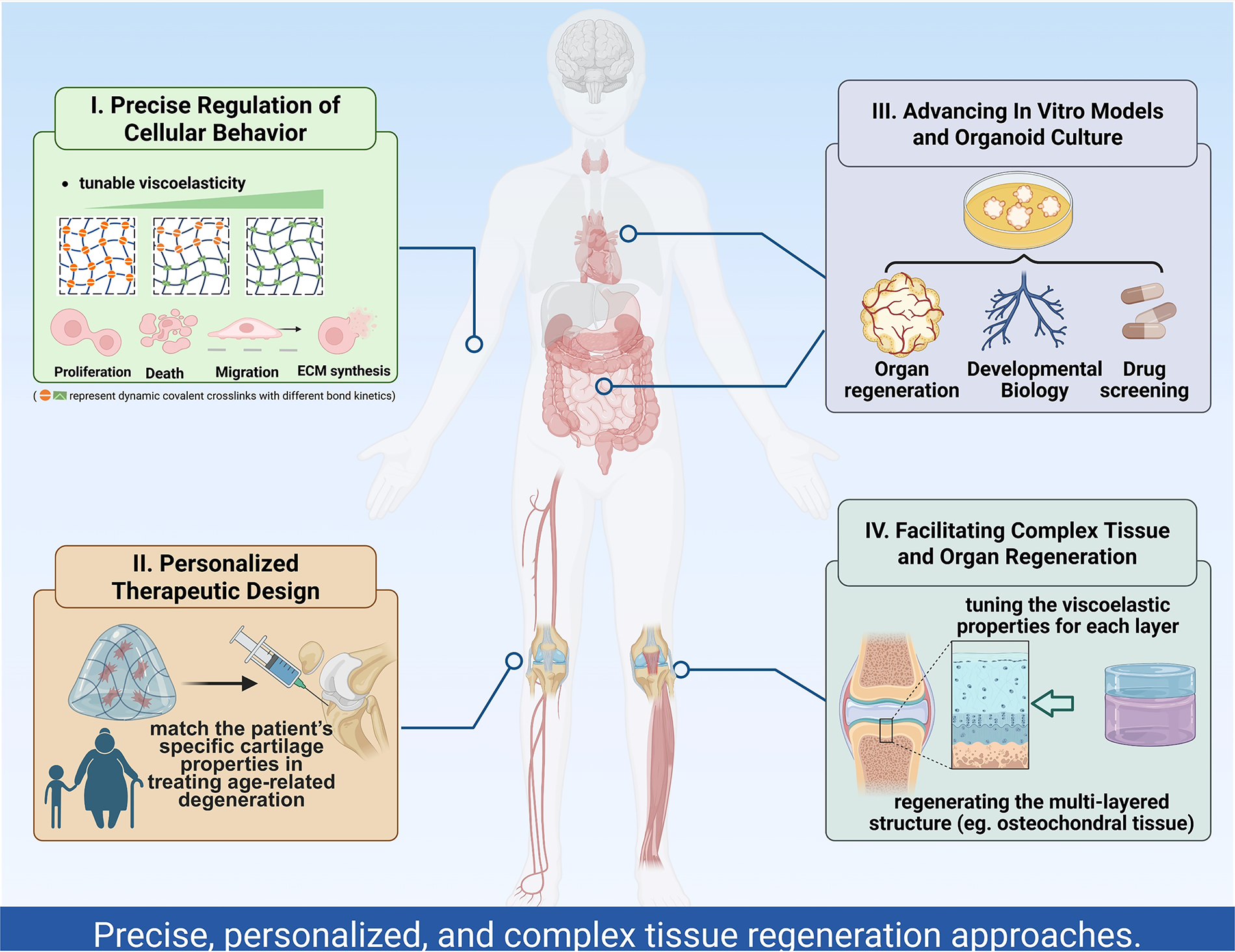
Schematic representation of the prospective applications of viscoelastic independently tunable hydrogels in regenerative medicine.
Precise regulation of cell behavior: By independently adjusting viscoelasticity, researchers can fine-tune cell proliferation, migration, and differentiation, thus providing an optimal microenvironment for specific cells in regenerative medicine.
Personalized treatment schemes: Viscoelastic hydrogels with independent regulation can be customized according to the physiological conditions of individual patients. In particular, for tissue repair or organ regeneration, this allows for personalized material design tailored to specific diseases or injuries, further advancing the field of personalized regenerative medicine.245,246
Improvement of organ models and organoid culture in vitro: Applying hydrogels with independently adjustable viscoelasticity in organ and organoid cultures in vitro can better simulate the in vivo microenvironment, enhancing the physiological relevance of the models and the accuracy of experimental results.109,117,241,244
Regeneration of complex tissues and organs: These materials may provide a foundation for the regeneration of complex tissues or organs with multiple layers and components. More research is needed to gradually explore the appropriate viscoelastic parameters of materials for various tissue engineering applications.247–249
Overall, viscoelastic independently adjustable hydrogels are expected to drive the development of regenerative medicine toward more refined, personalized, and complex system regeneration.
Authors’ Contributions
Q.Q.: Investigation, visualization, validation, and writing—original draft. Y.S.: Methodology and data curation. J.W.: Visualization and methodology. X.L.: Conceptualization. L.Z.: Conceptualization. H.Y.: Conceptualization. N.Z.: Conceptualization. K.Z.: Supervision. Z.Z.: Investigation, methodology, and funding acquisition. Y.B.: Writing—review and editing, project administration, funding acquisition, and conceptualization.
Footnotes
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant Nos. 82071144 and 82301117), the Innovation Research Team Project of Beijing Stomatological Hospital, Capital Medical University (Grant No. CXTD202203), the Beijing Stomatological Hospital, Capital Medical University Young Scientist Program (Grant No. YSP202010), and the Beijing Hospitals Authority Clinical Medicine Development Special Funding Support (Grant No. ZLRK202330).
Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.