
Abstract
Adipose tissue is a highly plastic organ whose remodeling dynamics are central to whole-body metabolic health. Expansion of white adipose tissue occurs through either hyperplasia, which preserves tissue function, or hypertrophy, which causes local hypoxia, inflammation, and pathological extracellular matrix (ECM) accumulation. Under hypertrophic conditions, the ECM stiffens and transitions from a supportive scaffold to a fibrotic barrier that limits expansion and perpetuates metabolic dysfunction. Understanding how mechanical cues regulate adipose tissue remodeling is, therefore, essential for identifying new therapeutic strategies. Two mechanosensitive cell populations, adipose stem cells (ASC) and mature adipocytes, are central to this process. ASC interpret ECM stiffness and compositional changes, which determine lineage outcomes. Soft and flexible matrices favor adipogenesis, whereas stiff matrices drive fibroblast-like activation and matrix deposition. Adipocytes, though differentiated, retain mechanosensitive signaling capabilities that shape their function. Under chronic mechanical stresses, cytoskeletal remodeling pathways lead to changes in gene expression and partial dedifferentiation toward a fibroblast-like phenotype. Reciprocal signaling between ASC and adipocytes amplifies these processes, establishing feedback loops that reinforce either healthy or pathological remodeling. Cell and tissue engineering approaches are essential for dissecting these processes, with hydrogel substrates, 3D scaffolds, compression assays, and atomic force microscopy offering physiologically relevant platforms to model progenitors and adipose tissue cellular mechanics. Emerging tools, including nanotopography and mechanical stimulation devices, have the capacity to further clarify how mechanical signals influence adipose remodeling. By positioning ASC and adipocytes as active regulators of ECM mechanics, we underscore the importance of mechanotransduction pathways in adipose tissue health and point to bioengineering strategies that may help discover ways to restore tissue flexibility and improve metabolic outcomes.
Impact Statement
Adipose tissue function is dependent on its ability to remodel in response to metabolic demands. Mechanical cues from the extracellular matrix (ECM) dictate whether remodeling under conditions of obesity proceeds via adaptive hyperplasia or maladaptive fibrosis. This review highlights how adipose stem cells and mature adipocytes integrate ECM mechanics through integrin signaling, cytoskeletal remodeling, and mechanosensitive transcriptional pathways. By connecting mechanobiology to adipose tissue health, we emphasize both the fundamental biology and engineering approaches of targeting mechanical signaling to restore adipose tissue plasticity and improve systemic metabolic health.
Keywords
Introduction
Adipose tissue is a dynamic organ, capable of extreme remodeling in response to nutrient availability and metabolic demands.1–4 This remodeling reflects its capacity to respond to changes in energy balance, which influence whole-body metabolic regulation and endocrine function.5,6 In states of caloric excess, white adipose tissue (WAT) expands through two primary modes: the differentiation of new adipocytes through hyperplasia or the enlargement of existing adipocytes via hypertrophy.7–9 Hyperplastic expansion distributes lipid storage demands across a larger cell pool, thereby preserving tissue function and metabolic health.10–13 In contrast, hypertrophic remodeling is associated with cellular hypoxia, local inflammation, and heightened mechanical stresses.7,14,15 These maladaptive changes alter tissue mechanics, leading to the further pathological remodeling of the extracellular matrix (ECM), ultimately culminating in tissue fibrosis.16–18 In this context, the ECM shifts from a permissive scaffold that supports adipose plasticity to a structural barrier that limits expansion and perpetuates metabolic dysfunction by altering how cells sense and transduce force19,20 (Fig. 1). This raises fundamental questions about how mechanical cues from and within the ECM regulate cell fate and adipose tissue function.
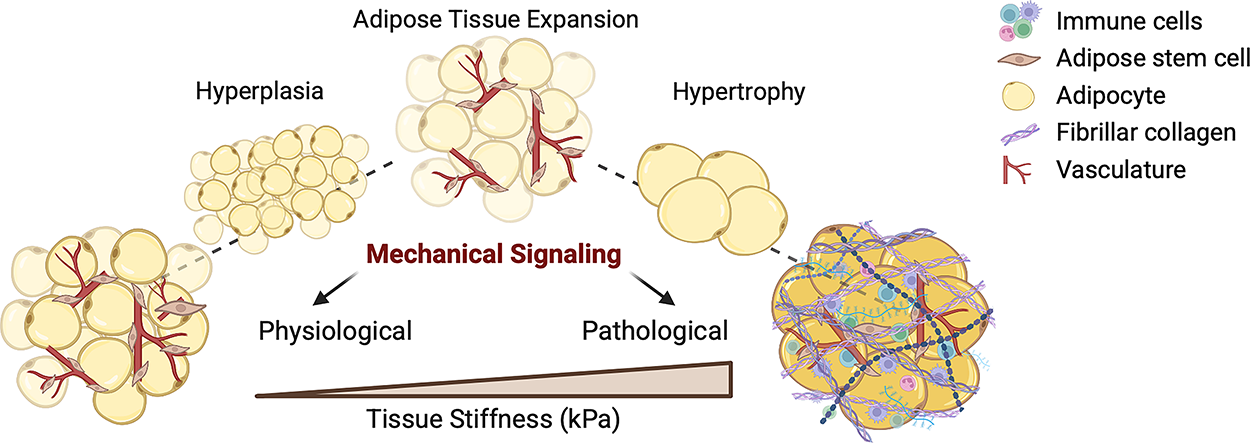
Adipose expansion governs tissue stiffness and fibrotic remodeling. During caloric excess, adipose tissue expands through either hyperplasia or hypertrophy. Hyperplasia, the differentiation of new adipocytes from progenitors, preserves tissue function and metabolic flexibility. In contrast, hypertrophy, the enlargement of existing adipocytes, drives hypoxia, inflammation, and ECM remodeling. Increased mechanical stress and tissue stiffening, resulting from hypertrophic expansion, promotes fibrotic tissue remodeling. Figure generated with BioRender. ECM, extracellular matrix.
Mesenchymal progenitors and adipocytes represent two mechanosensitive cell populations within adipose tissue that together regulate tissue remodeling by sensing and responding to ECM cues.21,22 Under physiological healthy ECM conditions, progenitors differentiate into new adipocytes, thereby maintaining tissue expandability and metabolic function. 23 By contrast, stiff or fibrotic environments trigger their activation into matrix-producing fibroblasts, exacerbating tissue rigidity and dysfunction.24,25 Adipocytes themselves also remodel their cytoskeleton and gene expression in response to matrix stiffening, undergoing partial dedifferentiation under chronic stress.26–28 These distinct responses to soft versus stiff ECM illustrate how matrix mechanics direct lineage specification. Compliant environments favor adipogenic programs, while rigid matrices enhance cytoskeletal tension, mechanotransducive signaling, and fibrogenic gene expression.29,30 In this way, progenitors and adipocytes act as integrators of mechanical and biochemical cues, reinforcing remodeling pathways. Mechanical inputs are conveyed through integrin-mediated signaling, integrating biochemical ligand engagement and matrix stiffness cues, and also via Piezo ion channels, which respond directly to membrane tension as a force-sensing mechanism.31,32 By positioning both cell types at the center of adipose tissue mechanobiology, it becomes evident that these cells are not passive responders but active regulators of adipose remodeling, determining whether expansion proceeds via hyperplasia and healthy lipid storage or becomes locked into a fibrotic state. The goal of this review is to highlight the mechanobiological mechanisms that govern adipose tissue remodeling, with a focus on progenitors and adipocytes. We will discuss how matrix composition and stiffness regulate cell fate and function, how these processes contribute to adipose tissue fibrosis, and where engineering approaches can help uncover ways to restore tissue flexibility and improve health.
Mechanical and Biochemical Signaling Governing Cell Fate and Phenotype
Cells sense mechanical cues through defined mechanotransduction pathways that link ECM mechanical and compositional properties to cell differentiation pathways and functional states 33 (Fig. 2). These signaling cascades begin at integrins and extend through focal adhesion complexes, cytoskeletal remodeling, and ultimately to mechanosensitive transcriptional regulators that determine cell fate. Integrins function primarily as biochemical and mechanical sensors, engaging ligands such as collagen and fibronectin while transmitting matrix-derived forces across the cell membrane.34,35 In adipose tissue, β1 integrins have been shown to regulate cell differentiation, while αv integrins control matrix stiffening and fibrosis.36,37
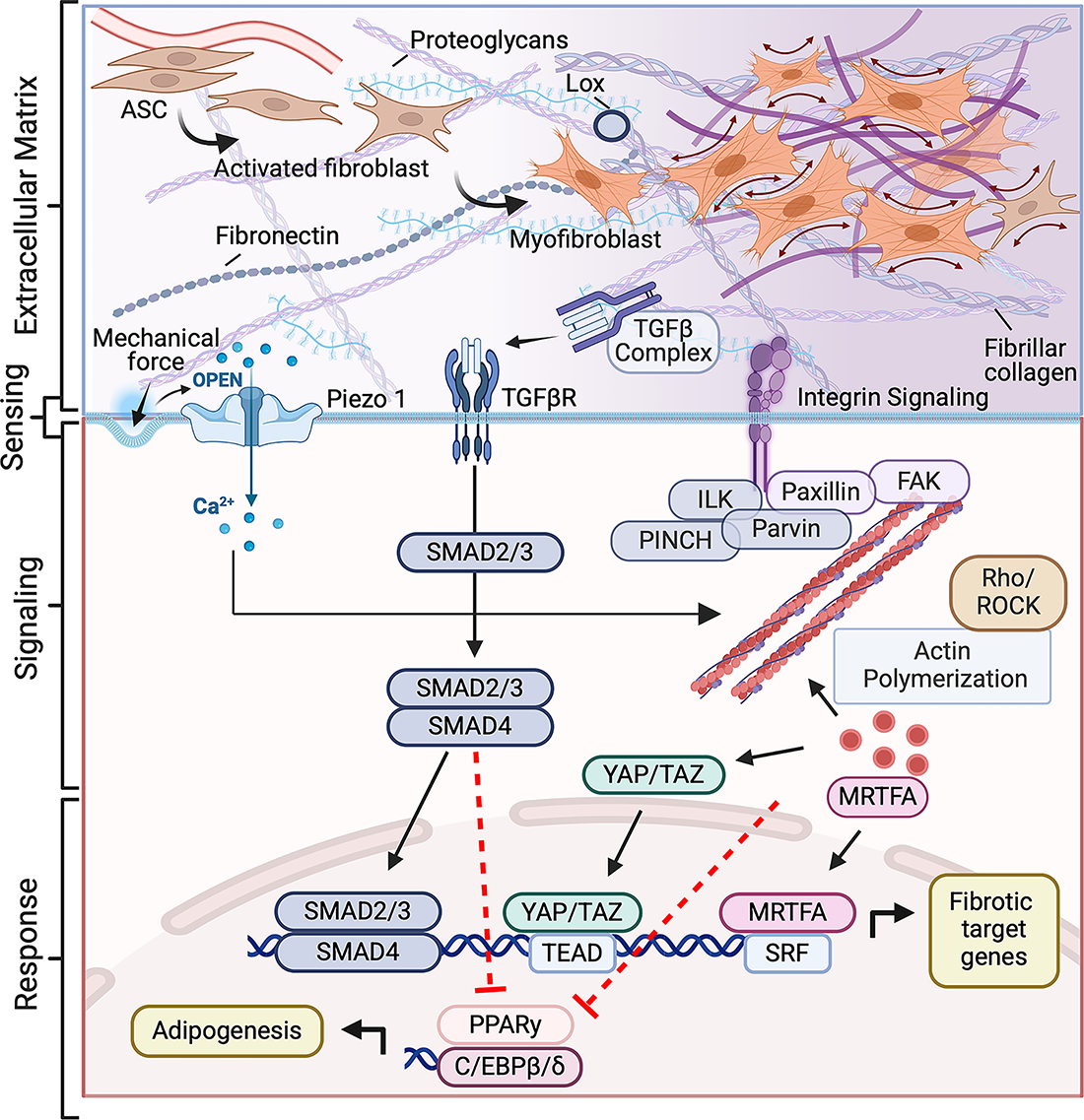
Mechanical and biochemical signaling converge to regulate fibroblast activation and adipogenic fate. Mechanical and biochemical cues transmitted from the extracellular matrix are sensed by integrins, mechanosensitive ion channels (Piezo1), and TGFβ receptors. These inputs activate focal adhesion complexes (FAK, Paxillin, ILK–PINCH–Parvin) and downstream Rho/ROCK-dependent actin polymerization, promoting nuclear translocation of mechanosensitive transcriptional regulators MRTFA and YAP/TAZ. In parallel, canonical TGFβ signaling activates SMAD2/3-SMAD4 complexes. Together, these pathways drive expression of fibrotic target genes while repressing adipogenic regulators PPARγ and C/EBPβ/δ, establishing a mechanochemical feedback loop that sustains fibroblast differentiation and ECM remodeling. Figure generated with BioRender. C/EBP, CCAAT/enhancer-binding protein; FAK, focal adhesion kinase; ILK, integrin-linked kinase; Lox, Lysyl Oxidase; MRTFA, myocardin-related transcription factor A; PINCH, Particularly Interesting Cysteine- and Histidine-rich protein; PPARγ, peroxisome proliferator-activated receptor; Rho, Ras homolog family member; ROCK, Rho-associated coiled-coil kinase; SMAD, Sma and Mad related proteins; SRF, Serum response factor; TAZ, Transcriptional coactivator with PDZ-binding motif; TEAD, TEA domain transcription factors; TGFβ, transforming growth factor beta; YAP, Yes-associated protein.
Upon ligand binding, integrin clustering initiates the formation of focal adhesions, dynamic signaling complexes that couple extracellular mechanical forces to the actin cytoskeleton. Key components include focal adhesion kinase (FAK), paxillin, integrin-linked kinase (ILK), and Particularly Interesting Cysteine- and Histidine-rich protein (PINCH), which together form mechanosensitive complexes that translate substrate stiffness into intracellular tension.34,38–40 ILK acts as a central mechanosensitive linker within the integrin-PINCH-parvin complex, connecting β-integrins to actin-regulatory pathways. Its activity enhances actomyosin contractility and focal adhesion maturation, promoting progenitors into a fibroblast-like state in stiff or fibrotic ECM through mechanosensitive transcription factors. Inhibition of ILK restores adipogenic differentiation and reduces cytoskeletal tension.41,42 These findings suggest that integrin–ILK signaling operates as an early mechanical checkpoint in fibroblast activation, converting external matrix cues into sustained cytoskeletal tension that favors fibrosis over adipogenesis. In earlier studies, integrin-mediated adhesion and cell spreading were shown to bias stem cell/progenitor differentiation; rounded cells on compliant substrates underwent adipogenesis, whereas spread cells on stiff matrices activated cytoskeletal tension and fibrogenic programs.43,44 These principles extend to both adipose progenitors and adipocytes, where matrix stiffening promotes focal adhesion maturation, fibroblast-like activation, and altered cell function.26,45,46
Downstream of focal adhesion complexes, the Ras homolog family member (Rho)/Rho-associated coiled-coil kinase (ROCK) axis serves as a central mechanical-to-biochemical pathway that governs actin polymerization and cellular contractility. Increased cytoskeletal tension releases the transcriptional coactivator myocardin-related transcription factor A (MRTF-A) from globular actin (G-actin), enabling nuclear translocation and activation of serum response factor (SRF)-dependent genes. 47 In progenitors, this pathway promotes expression of α-smooth muscle actin (α-SMA) and other profibrotic markers while repressing peroxisome proliferator-activated receptor (PPARγ)-driven adipogenesis. 48 MRTF-A also regulates alternative adipocyte phenotypes, including beige adipocyte conversion, underscoring the flexibility of actin-dependent transcriptional control. 49
In parallel, Yes-associated protein (YAP) and Transcriptional coactivator with PDZ-binding motif (TAZ) transduce mechanical signals into nuclear transcriptional outputs. Activated large tumor suppressor inase 1 and 2 (LATS1/2) repress YAP/TAZ by phosphorylation and by targeting them for degradation by the ubiquitin proteasome system. 50 In response to mechanical stimuli, YAP/TAZ are activated by FAK and ILK in the focal adhesion complex by canonical LATS1/2 inhibition and noncanonical direct phosphorylation of YAP/TAZ. 50 More recently, YAP/TAZ have been shown to be activated in response to mechanical stimuli by PIEZO signaling.51,52 Because of these mechanisms, YAP/TAZ activation is strongly correlated with substrate stiffness and cytoskeletal tension, and in adipose contexts, YAP activity represses adipogenesis and induces fibrosis.53,54 Recent work showed that the Hippo/YAP pathway integrates adipocyte plasticity with fibrotic remodeling, identifying it as a central regulator of adipose tissue remodeling. 55 Together, MRTF-A and YAP/TAZ exemplify how biophysical signals are converted into transcriptional outcomes that determine whether progenitors generate new adipocytes or contribute to matrix deposition.
Mechanosensitive ion channels provide an additional route for force detection. Piezo1, a stretch-activated nonselective cation channel, converts changes in membrane tension into Ca2+ influx that reprograms cellular behavior and tissue phenotypes. 56 Piezo1 activation suppresses adipogenic differentiation and promotes cytoskeletal remodeling across multiple contexts, including perivascular adipose progenitors and stiffness-dependent lineage switching in mesenchymal and osteosarcoma cells.57,58 In adipose systems, this signaling axis shapes cell–cell communication and tissue state, modulating paracrine signaling, inflammation, and metabolic regulation.59,60 Recent findings further identify a complementary mechanosensory role for Piezo2 in adipose sensory innervation, linking tissue mechanics to neural feedback and metabolic control. 61 Functionally, Piezo1-driven Ca2+ signals operate alongside integrin/FAK–Rho circuits and transforming growth factor beta (TGFβ) activation, providing an additional pathway by which mechanical cues are integrated into fate and phenotype decisions. Collectively, these studies identified Piezo channels as critical regulators of adipose tissue mechanotransduction, functioning in parallel with integrin signaling to fine-tune how cells interpret mechanical stiffness versus membrane tension.
TGFβ signaling further amplifies these effects. Latent TGFβ complexes tethered to the ECM are mechanically activated by integrin-mediated traction, making their bioavailability directly dependent on matrix stiffness and cellular contractility.62–64 Once active, TGFβ/SMAD signaling suppresses adipogenic differentiation and reinforces fibrotic remodeling.65–68 Crosstalk with other mechanosensitive pathways reinforces this response. YAP/TAZ and MRTF-A intersect with SMADs in the nucleus to produce synergistic transcriptional programs that stabilize fibroblast-like activation, while integrin/FAK and Rho/ROCK signaling remodels the cytoskeleton to support nuclear translocation of these transcription regulators.34,69–71 Together, these pathways form reinforcing regulatory loops in which mechanical stress remodels the ECM, cells sense these changes, and mechanotransduction drives a shift away from adipogenesis toward fibrosis.72–74
Mechanobiology of Adipose Stem Cells
Adipose stem cells (ASC) are a fibroblast-like, multipotent cell population within the stromal vascular fraction of WAT.10,11 Throughout the literature, this population is variably termed ASC, mesenchymal progenitors, adipose progenitors, and perivascular stromal cells, potentially reflecting differences in isolation, marker sets, and functional definitions across depots and species; here, we use ASC for clarity. These cells are abundant, heterogeneous, and retain a plastic phenotype that enables them to differentiate into mature adipocytes or transition toward a profibrotic fate depending on environmental cues.23,24 Lineage-tracing studies have shown that platelet-derived growth factor receptor (PDGFR)α+ and PDGFRβ+ stromal perivascular cells give rise to most new adipocytes during high-fat feeding and cold exposure, while fibro-inflammatory subpopulations emerge under metabolic stress to reinforce ECM deposition.12,75 Single-cell analyses in both mice and humans reveal depot-specific and disease-associated biases, with some subpopulations primed for adipogenesis and others for fibrosis.9,76,77
A defining feature of ASC is its responsiveness to ECM mechanics. In compliant environments that approximate healthy adipose stiffness (∼0.5–2 kPa), ASC typically favor adipogenic differentiation.43,78 By contrast, stiffened or fibrotic ECM drives fibroblast-like activation characterized by actin stress fiber formation, focal adhesion maturation, and expression of profibrotic genes.79,80 These opposing outcomes reflect the dual sensitivity of ASC to both biochemical ligand composition and physical stiffness of the ECM, integrating integrin-mediated adhesion signaling with mechanotransducive force sensing. Pioneering work with polyacrylamide and polyethylene glycol-based hydrogels revealed that healthy adipose stiffness promoted lipid accumulation and PPARγ-driven adipogenesis, whereas stiffer matrices (>10 kPa) induced cytoskeletal tension, nuclear translocation of MRTF-A and YAP/TAZ, and expression of α-SMA and collagen.26,45 Complementary micropatterning and cell-shape studies confirmed that rounded ASC on soft or constrained substrates adopt adipogenic fates, whereas tension-bearing cells resist differentiation and instead activate fibrotic programs.44,81 Atomic force microscopy (AFM) of diabetic human adipose tissue further validates these concepts in vivo, showing that ASC become more contractile and stiffer in disease states. 82 This in vivo validation underscores that mechanical adaptation of ASC is not an artifact of culture models but a defining feature of metabolic disease, linking tissue stiffness directly to impaired adipogenic capacity.
Matrix composition provides equally potent cues. Collagen VI, abundant in the adipose tissue ECM, restricts tissue expandability, and its loss improves metabolic flexibility in mice. 83 Lysyl oxidase-mediated crosslinking stiffens collagen networks and amplifies profibrotic signaling. 16 Fibronectin fragments engage integrins and promote focal adhesion signaling, biasing ASC toward fibroblast-like activation.34,84 In addition to structural components, there are ECM-associated proteins that can act as active modulators of ASC fate. For example, aortic carboxypeptidase-like protein (ACLP), strongly induced in fibrotic WAT, enhances ASC differentiation into myofibroblasts and represses adipogenesis. 85 Additional proteins, such as periostin and thrombospondins, further sensitize ASC to mechanical stress and reinforce matrix deposition.86,87 Further research is needed to identify relevant ECM ligand-receptor interactions that govern ASC fate in fibrotic tissues. Integrating ECM proteomics with single-cell transcriptomics offers a powerful strategy to map these signaling axes and pinpoint therapeutic targets that could restore adipogenic capacity in fibrotic tissues.
Together, these studies demonstrate that ASC are highly responsive to mechanical signals that translate the cues from their microenvironment into lineage outcomes. Through integrin engagement, cytoskeletal remodeling, and activation of mechanosensitive transcription factors, they determine whether adipose tissue expansion proceeds through healthy adipogenesis or becomes constrained by fibrosis. For tissue engineering, this highlights the need to consider not only the availability of the ASC populations but also the mechanical and molecular context in which they reside, as subtle shifts in ECM architecture can tip the balance between regeneration and pathological remodeling.
Mechanobiology of Adipocytes
During adipogenesis, ASC undergo dramatic morphological changes, cytoskeletal reorganization, inactivation of focal adhesion factors, and repression of mechanoresponsive proteins such as MRTF-A.88,89 Despite these changes during differentiation, mature adipocytes retain mechanosensitive activity, continuing to detect both biochemical ligand engagement and physical mechanical load. This mechanical responsiveness is crucial for their survival and may mediate pathological phenotypes during adipose tissue fibrosis and chronic obesity.
Mechanical sensitivity is important for adipocyte survival and function. In adipocytes, active β1 integrins cooperate with insulin receptor signaling to regulate insulin sensitivity. Adipose-specific loss of β1 integrin activity impairs insulin action and drives systemic insulin resistance. 90 Stretching of the adipocyte plasma membrane and subsequent FAK activation induces Glut4 translocation, which mediates glucose import. 91 In fact, adipocyte-specific loss of FAK reduces adipocyte survival, which underscores the importance of FAK and glucose import for adipocyte function.92,93
Similar phenotypes have been observed in adipocyte-specific knock-out (KO) of other focal adhesion and mechanosensitive proteins. Adipocyte-specific KO of kindlin-2, which is a focal adhesion protein, results in lipodystrophy and adipocyte apoptosis. 94 Similarly, adipocyte-specific KO of the mechanosensitive ion channel, Piezo1, results in reduced adipose fat mass and an increase in compensatory hepatic lipid accumulation, demonstrating how loss of tension sensing alters systemic lipid distribution. 59 Interestingly, KO of PINCH, a focal adhesion adaptor protein, in adipocytes results in adipocyte death but improved metabolic outcomes in mice fed a high-fat diet. 39 Similar phenotypes have been reported with adipocyte-specific KO of ILK, which reduced adipose size and improved systemic glucose tolerance on a high-fat diet.40,95 These findings suggest that while ILK supports adipocyte structural integrity and survival through focal adhesion maintenance, its chronic activation in obesity may drive insulin resistance and fibrosis via persistent Rho/ROCK and YAP/MRTF-A signaling. Consistent with this, recent reviews highlight that focal adhesion proteins such as β1 integrin, FAK, Kindlin-2, ILK, and PINCH collectively coordinate adipocyte differentiation, survival, and insulin sensitivity, and that elevated ILK expression in visceral fat of obese humans correlates with insulin resistance and fibrosis. 96 While KO of all these focal adhesion and mechanosensitive proteins resulted in adipocyte death or reduced the capacity of the adipose tissue to store lipid, more research is needed to uncover the mechanisms behind the disparate metabolic outcomes resulting from perturbation of these pathways.
Although mechanosensitivity is important for adipocyte survival, hyperactivation of mechanosensitive pathways can cause loss of the adipocyte differentiated phenotype and pathological remodeling. For example, prolonged mechanical stress induces de-differentiation of mature adipocytes, characterized by the extrusion of the lipid droplets and morphological reversion to a fibroblastic-like state.97,98 This phenomenon has been observed in vitro with ceiling culture systems, which inflict superphysiological mechanical stresses on adipocytes through prolonged contact with stiff cell culture plastic.97,98 Similar de-differentiation events have been induced in vivo by genetic constitutive activation of YAP/TAZ in mature adipocytes, underscoring the sufficiency of mechanotransducive signaling in adipocyte de-differentiation.27,55
During chronic obesity in mice, complete de-differentiation of adipocytes in adipose tissue has not been observed; however, partial phenotypic reversions of adipocytes toward an ASC-like state have been reported.99,100 Adipocytes from obese mice compared with adipocytes from lean mice exhibit reduced expression of adipocyte marker genes, increased expression of fibroblastic marker genes, and increased cortical actin stress fiber formation.99,101 These phenotypic reversions have been attributed to increased TGFβ activity in obese adipose tissue, but in vitro evidence additionally supports direct mechanical signaling for these changes. 102 Adipocytes suspended in 3D gel systems have increased fibrotic and inflammatory gene expression in response to stiffer matrices or direct compression of the gel scaffolds.26,103 Furthermore, RhoA inhibition in adipocytes of obese mice has been shown to improve adipokine expression and reduce inflammatory signaling, suggesting that modulating cytoskeletal tension can restore aspects of normal function. 104
Consistent with these findings, hypertrophic adipocytes display increased integrin expression and load-dependent activation, linking ECM stiffness to cytoskeletal tension and altered gene expression.105,106 These in vitro and in vivo studies suggest a paradigm whereby extracellular mechanical stressors activate adipocyte focal adhesion complexes, causing filamentous actin (F-actin) formation and subsequent transcriptional reprogramming. Transcription factors regulated by mechanical activation, such as MRTF-A and YAP/TAZ, may play a role in gene expression changes resulting from mechanical activation. In fact, YAP/TAZ are activated in adipocytes during obesity, mediate the de-differentiation response to ceiling culture conditions, and inhibit PPARγ activity in adipocytes.16,55,107 Likewise, MRTF-A is activated in adipocytes in response to cortical F-actin formation. 108 Further research is needed to understand how mechanical stress directly affects adipocyte function and cell-state, especially in clinically relevant mechanical ranges representative of fibrotic or hypertrophic WAT.
Mechano-Crosstalk Between ASC and Adipocytes
ASC and adipocytes transmit reciprocal mechanical signals to orchestrate gene expression patterns and cell fate of proximal cells. Both ASC and adipocytes respond to biochemical and mechanical cues by secreting collagen and other ECM components, which stiffens their proximal ECM and may establish a positive feedback loop of ECM deposition between neighboring ASC and adipocytes as they respond to the mechanical niche.75,109 Through these processes, mechanical and compositional remodeling reinforce one another, amplifying local stiffness and altering mechanosensitive signaling dynamics.
Furthermore, enhanced plasma membrane stiffness in hypertrophic adipocytes or adipocytes with increased cortical F-actin can create a stiffer niche for adjacent ASC or adipocytes.110–113 It has been computationally modeled that plasma membrane stiffness of adipocytes is potentiated by the volume of neighboring adipocytes, indicating that hypertrophic adipocytes can mechanically influence their neighbors by transmitting lateral stress across shared ECM networks. 114 These findings highlight how mechanical load is not distributed uniformly in adipose tissue but propagates through intercellular and matrix connections, coupling adipocyte hypertrophy to stromal remodeling.
Mechanical stimulation of adipocytes has also been shown to direct the cell fate of ASC; the mechanical stimulation of Piezo1 in mature adipocytes is reported to stimulate the secretion of fibroblast growth factor 1 (FGF1), which promotes the differentiation of nearby ASC to adipocytes.58,60 This mechanism demonstrates how tension sensors, such as Piezo channels, translate mechanical strain into additional paracrine biochemical cues that act on other cells. Together, these observations suggest that adipocytes are not passive responders to matrix stiffness but active participants in shaping the mechanical landscape sensed by ASC.
Engineering Perspectives
There have been many advancements in culture methods and mechanical assays to study the mechanobiology of adipocytes and ASC. These cells are highly mechanosensitive, meaning conventional growth on rigid polystyrene (∼1 GPa) can artifactually increase cytoskeletal tension and mechanotransduction, skewing fate and function compared with the soft adipose niche (∼0.5–2 kPa). 115 This mechanical mismatch biases lineage decisions in mesenchymal cells and alters adipocyte gene programs. Consequently, in vitro systems that better approximate tissue-level stiffness, viscoelasticity, and 3D architecture are preferred to capture physiological behavior (Fig. 3). These culture techniques can be coupled with biomechanical assays to study cell fate decisions and transcriptional responses to ASC and adipocytes under mechanical stimulation.
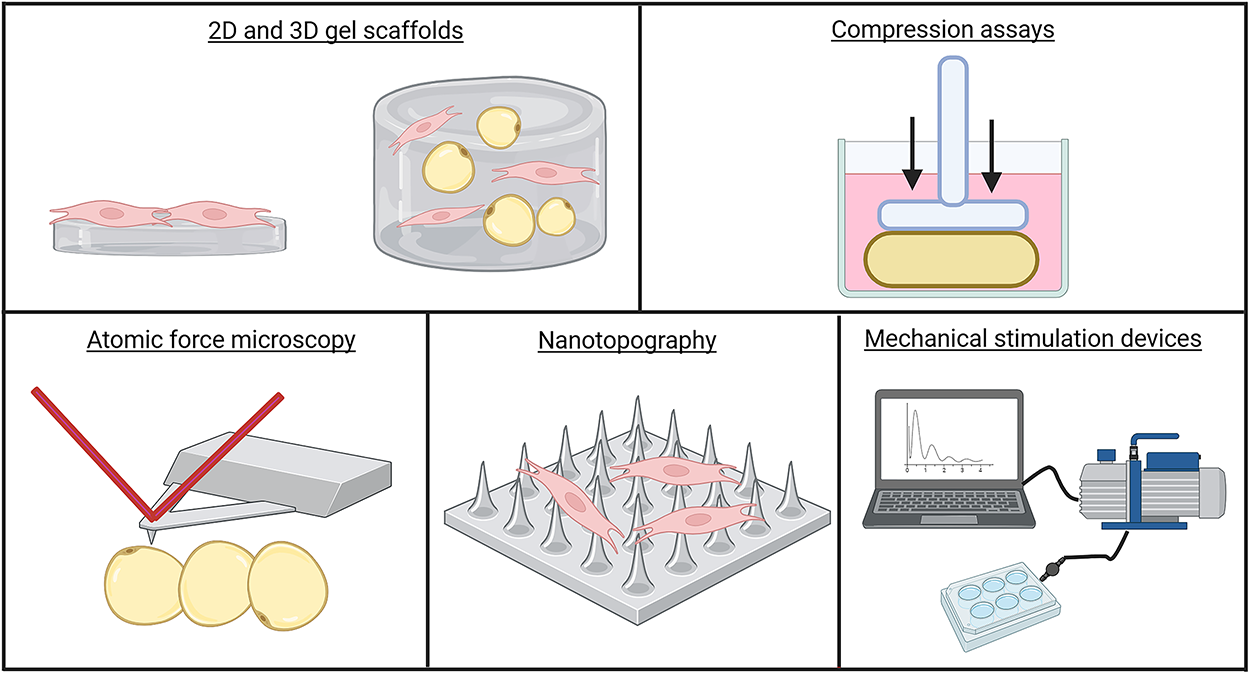
Engineering methods to study adipose tissue remodeling. ASC and adipocyte mechanics have been explored using a variety of engineering techniques. Biomimetic surfaces, including 2D and 3D hydrogels and nanotopography, have been used to simulate adipose-like mechanical properties. Precise compressive and tensile forces have been imposed on intact tissue, adipocytes, and ASC using weight-loading compression assays and mechanical stimulation devices. To quantify stiffness changes in tissues and cells in response to mechanical stimuli, atomic force microscopy remains a staple technique in the field. Figure generated with BioRender. ASC, adipose stem cells.
Adipocyte ceiling culture
It is often challenging to culture primary adipocytes, in part due to their buoyancy in culture media. One culture method developed is the ceiling culture system, which leverages the buoyancy of adipocytes to induce their adherence to the ceiling of plastic culture flasks. 98 This culture method proved ineffective at culturing adipocytes because the culture plastic-induced superphysiological mechanical stress on the adipocytes, leading to loss of the lipid droplets and reversion to a fibroblastic morphology. 98 While not useful for stabilizing adipocytes in culture, this technique has been used to study the de-differentiation of adipocytes in vitro.97,98,116,117 This technique was used to find that adipocyte de-differentiation was dependent on mechanical stress and associated with YAP expression. 116 Kim et al. improved this technique in a ceiling culture chip system to study the mechanical response of adipocytes, while facilitating live-imaging and preventing cell loss. 97 Using this technique, they demonstrated that mature adipocyte de-differentitation is mechanically dependent and driven by actin remodeling, Hippo suppression, and hedgehog signaling. 97 This technique may be combined with 2D gel matrices to simulate physiological stiffness and investigate partial phenotypic reversion observed in adipocytes of obese individuals.
2D gels and 3D gel scaffolds
2D gels have been used to study the response of ASC to mechanical signals. Most commonly, 2D gels are polyacrylamide hydrogels with the ability to tune stiffness by adjusting acrylamide and bis-acrylamide concentrations. 29 The surface is made cell-adhesive by covalently coupling ECM proteins, such as collagen and fibronectin, using crosslinkers such as sulfo-SANPAH or polydopamine. 118 These gels can also be prepared as stiffness gradients in a single well (∼0.5–8 kPa) to assess cell durotaxis. 119 To study the mechanical sensitivity of mature adipocytes, 3D gel scaffolds have been developed and optimized to encapsulate buoyant adipocytes.26,103,108 The stiffness of these gels can be tuned similarly to 2D gels by changing the gel composition. 26 By tuning mechanical 3D gel stiffness, Caprio et al. demonstrated that gel mechanical cues drive actin polymerization and collagen expression in mature adipocytes. 26 This technique was advanced by integrating decellularized matrix proteins from human adipose tissue into 3D collagen gels to mimic physiological mechanical cues. Using this approach, it was found that obese human matrices increased the secretion of pro-inflammatory cytokines, reduced adiponectin expression, and impaired lipolysis in mature primary human adipocytes. 103 It was also demonstrated that human matrices drove Yap/Yaz transcriptional activity through β1 integrin signaling in mature adipocytes. 103 Additionally, 3D gels can be mechanically compressed, stretched, or otherwise deformed by external interventions to induce specific mechanical signals to adipocytes. 103 These 3D gel scaffolds have also been used to coculture adipocytes with other cell types. 120 While this approach has mostly been explored with tumor cells, ASC and immune cells could also be cocultured with adipocytes in 3D gels, enabling the investigation of mechanical crosstalk between various adipose tissue cell types.
Uniaxial compression assays
Several groups have developed uniaxial compression assays to model external mechanical stressors on intact adipose tissue and 3D culture gels.103,121 Adipose tissue can be uniaxially compressed by adding weights atop tissue in culture wells. 121 This technique was used to measure tissue fragility but may be adapted to model the physiological compression of adipose tissue in obese subjects. Pellegrinelli et al. performed uniaxial compression on adipocytes cultured in a 3D gel using a similar method. Using this technique, they demonstrated compression mimics the mechanical stress induced by obese adipose tissue matrices and increases pro-inflammatory cytokine secretion in human adipocytes. 103 Applications of this technique can be further explored to study phenotypic changes in ASC and adipocytes ex vivo.
Atomic force microscopy
AFM can precisely measure surface stiffness of fragile biomaterials by indenting with a calibrated cantilever, recording force/distance curves, and fitting contact models to extract a Young’s modulus. 122 In adipose research, AFM has been applied directly to native tissue using semi-infinite or finite thickness models to avoid substrate effects that bias modulus estimates. 123 For example, human visceral adipose from individuals with diabetes was approximately twice as stiff as nondiabetic tissue measured by AFM nanoindentation (∼11.5 kPa vs ∼4.5 kPa), consistent with fibrosis-associated remodeling.82,124 Beyond tissue-level measurements, AFM has also been used to study the surface stiffness of ASC as they differentiate to adipocytes. AFM resolves subcellular stiffness changes as preadipocytes accumulate lipid. 125 While in mature adipocytes, AFM has captured stiffness changes with hypertrophy and maturation and linked them to cytoskeletal contractility. 113 AFM will continue to be leveraged as a tool to study cell surface stiffness in response to mechanical perturbations in adipose tissue.
Nanotopography
Nanotopography is the fabrication of contact surface features at the scale of cell surface receptors to alter cell shape and fate. 126 Nanotopography has been used to study the differentiation of stem cells to adipocytes in response to distinct surface architecture. Using laser or capillary force lithography, highly precise patterns can be constructed to mimic tissue surface architectures in vitro.127,128 In fact, nanotopography has been utilized to mimic adipocyte surface patterns, which have been shown to enhance stem cell adipogenesis. 128 This technique can be further explored to model how tissue matrix or cell–cell contacts regulate ASC fate.
Mechanical stimulation devices
Mechanical stimulation devices enable precise and standardized application of compressive or tensile forces on 2D and 3D culture systems to study responses to mechanical stimuli. 129 Mechanical stimulation devices have been leveraged by some groups to apply standardized and controlled compression or tensile forces to 2D culture systems of ASC. 80 Early systems have been used to investigate the effect of periodic stretch on the differentiation of stem cells to adipocytes. 80 More advanced systems can now apply precise tensile or compression forces to 2D and 3D culture systems, enabling their utility to study adipocytes or complex adipose cell culture systems in 3D gel scaffolds. 130 Mechanical stimulation devices have been underutilized in mechanical studies of 3D adipose culture systems with precise parameters.
Outlook
Adipose tissue remodeling is a tightly regulated process in which progenitors and adipocytes act as key mechanosensitive cell populations. Integrin signaling, PIEZO signaling, cytoskeletal remodeling, MRTF-A and YAP/TAZ transcriptional programs, and TGFβ activation converge to determine whether adipose tissue expands through healthy adipogenesis or becomes constrained by fibrosis. ASC interpret changes in ECM stiffness and composition to guide lineage outcomes, while adipocytes, though differentiated, retain mechanosensitive signaling that influences their survival and plasticity. Crosstalk between these populations amplifies remodeling decisions, creating feedback loops that perpetuate either healthy adaptation or pathological remodeling.
Key questions remain regarding how adipocytes alter their function under pathological mechanical stress, and how ASC and other stromal and immune cell populations integrate and remodel the mechanical niche. Addressing these unknowns requires advanced bioengineering approaches that move beyond traditional rigid culture systems. Polyacrylamide hydrogels allow precise control over stiffness and ligand presentation, while 3D scaffolds and mechanical stimulation devices can replicate the buoyancy, compression, and dynamic loading that adipose tissue experiences in vivo. Tools such as AFM and traction force microscopy provide quantitative insight into how cells perceive and generate mechanical forces, and emerging technologies, including nanotopographic patterning and microfluidic coculture systems, offer new ways to model the complex crosstalk between adipocytes, ASC, and immune cells. Together, these approaches provide a path toward unraveling how mechanical cues shape remodeling outcomes. By integrating mechanobiology with bioengineering, the field is well-positioned to identify strategies that restore adipose tissue flexibility, interrupt fibrotic positive feedback loops, and ultimately improve metabolic health.
Authors’ Contributions
C.F.: Writing—original draft; Writing—review & editing. S.C.: Writing—original draft; Writing—review & editing. M.L.: Supervision; Writing—review & editing.
Footnotes
Acknowledgments
The authors would like to thank the Center for Multiscale & Translational Mechanobiology (CMTM) at Boston University for numerous helpful discussions, fostering collaborations, and providing trainees and faculty support. The authors would also like to thank current and past members of the Layne laboratory for discussions of these topics.
Funding Information
This work was supported in part by National Institutes of Health grants R01DK132080 and R01DK134534 (to M.D.L.) and F31DK139746 (to C.L.F.).
Disclosure Statement
No competing financial interests exist.