
Abstract
To provide an optimal wound bed for epidermal regeneration, a viable dermis is needed. As the dermis is destroyed in full-thickness burns, dermal templates (DTs) are used to create a healthy dermis for grafting and other procedures. Neonatal foreskin has exclusively been used as the source of dermal fibroblasts in commercial DTs, as the tissue is readily available and because these cells are assumed to be more proliferative and capable of superior wound healing compared with adult fibroblasts. The goal of this study was to assess the function of adult fibroblasts compared with neonatal fibroblasts for DT construction and epidermal regeneration. Primary fibroblasts were isolated from neonatal or adult surgical discard tissue (n = 4 each). Expression of collagen type 1 A1 (COL1A1) and matrix metalloprotease 1 (MMP1), MMP3, and MMP9 was assessed for each cell strain, and proliferation was quantified in two-dimensional (2D) cultures and 3D DTs. Subsequently, DTs were constructed from each cell strain by inoculating fibroblasts onto electrospun collagen scaffolds. DT contraction, extracellular matrix remodeling, and cell viability were assessed over 7 days in culture, and the ability of the DTs to promote epidermal regeneration was assessed using primary adult keratinocytes. No differences in gene expression were observed in neonatal versus adult fibroblasts in 2D culture. Neonatal fibroblasts were significantly more proliferative at day 7 when cultured in 2D; however, fibroblast proliferation was independent of donor age in 3D culture. Neonatal DTs contracted significantly more than adult DTs (68.8% ± 6.2% vs. 91.7% ± 4.2% original wound area, respectively). Upon seeding with keratinocytes, a robust, stratified epidermis formed in all DT groups, with no statistically significant differences in dermal or epidermal thickness, basal keratinocyte proliferation, epidermal barrier function, or basement membrane deposition. Analysis of gene expression revealed modest differences in the expression of MMP1, COL1A1, and ACTA2 in neonatal versus adult engineered skin in vitro, which were not associated with any discernable histological differences. These results indicate that the fabrication of DTs with adult fibroblasts can promote epidermal regeneration equivalent to that of neonatal fibroblasts but with less in vitro contraction, which may enable the treatment of larger wound areas.
Impact Statement
A viable dermis is critical to the success of split-thickness autografts and other strategies to close full-thickness wounds. The current fibroblast-seeded dermal templates (DTs) utilize neonatal cells; however, the suitability of adult fibroblasts for DTs was previously unknown. The current study showed that adult dermal fibroblasts supported epidermal health and regeneration to an equivalent degree with less in vitro contraction.
Introduction
The prompt closure of wounds is critical to prevent infection and sepsis in patients with massive, full-thickness burn injuries.1,2 Limited donor tissue poses a significant challenge in achieving complete wound closure and thus delays patient recovery. Novel tissue engineering-based strategies have been developed to speed up wound closure and improve outcomes.3B4 -8 While many of these strategies, such as cultured epithelial autografts (CEA) and spray-on skin cells, focus on epidermal regeneration, a viable dermal component is required for successful engraftment and optimal function. 9 For example, prior preclinical studies in a full-thickness wound model demonstrated that engraftment rates were significantly improved by combining CEAs with a collagen-glycosaminoglycan (GAG) DT compared with CEA alone without a dermal component. 10
To prepare the wound bed for autograft or tissue-engineered graft application, acellular dermal matrices (ADMs), such as collagen-GAG matrices, can be applied to the wound bed.11B12 -17 ADMs allow cell infiltration into the matrix to promote dermal regeneration 18 and have shown improvements to wound contracture, mechanical properties, and aesthetic outcomes when used with split-thickness autografts.19B20 -22 Despite these benefits, ADMs are limited by their relatively long time (up to 6 weeks) to vascularize and become populated with cells.23,24 Dermal matrices seeded with allogeneic fibroblasts may reduce the time to grafting and were shown to significantly improve the rate of re-epithelialization compared with ADMs. 15
Commercially available allogeneic cellular dermal matrices predominantly utilize neonatal foreskin fibroblasts,25B26 -30 in part due to the availability of the tissue but also due to the perception that neonatal cells have an increased regenerative capacity compared with adult fibroblasts.15,31,32 As skin ages, dermal fibroblasts deposit less collagen while matrix metalloproteinase (MMP) production is increased,33B34 -36 contributing to the thinning of the epidermis due to reduced extracellular matrix (ECM) deposition and alterations in its composition. Neonatal fibroblasts deposit more hyaluronic acid, collagen I, and collagen III compared with adult fibroblasts, with enhanced ECM organization in younger ECM. 37 Young adult fibroblasts (18–29 years old) were reported to proliferate in vitro at much greater rates (∼8-fold) than fibroblasts isolated from older adults (80+ years old). 35 However, the results from comparisons of fibroblasts of different ages have been variable.31,38,39 While two-dimensional (2D) culture can provide insight into the phenotypic differences between neonatal and adult cells, the in vitro environment is significantly different than the complex, natural 3D matrix of the native dermis.35,38,40B41 -43
To study fibroblast behavior in a 3D environment, collagen hydrogels are commonly utilized, although these studies have mainly focused on contraction instead of proliferation.38,44B45 -48 Within collagen hydrogels, fibroblasts from younger donors displayed increased contraction when comparing fetal or neonatal fibroblasts with adult fibroblasts; this was observed for fibroblasts of both human and porcine origin.38,44 For example, one study compared fetal and adult human fibroblasts on anchored collagen gels and found that fetal gels contracted significantly more initially, but over time, adult gels exhibited greater contraction. 45 These studies suggest differences in ECM production and remodeling, in addition to proliferation, in fibroblasts from young versus old donors. However, the lack of cell donor demographic information in many publications, as well as contradicting reports and the lack of direct comparisons in well-controlled studies, complicates the interpretation of the literature to compare the potential wound healing responses of young versus old fibroblasts. 49
The current study addresses a gap in knowledge regarding the relationship between dermal fibroblast age and the paracrine regulation of keratinocyte function. Understanding how neonatal and adult fibroblasts alter keratinocyte function is essential to optimizing epidermal regeneration in wounds treated with DTs. Fibroblasts secrete growth factors and cytokines, such as basic fibroblast growth factor (bFGF), keratinocyte growth factor (KGF), vascular endothelial growth factor A (VEGF A), and interleukin-6 (IL-6), which stimulate keratinocyte proliferation. In response, keratinocytes secrete IL-1, VEGF, and platelet-derived growth factor, which induce the migration of surrounding cells and promote fibroblast proliferation and ECM production.50-52 With increasing age, changes in these paracrine interactions cause rete ridges, the interdigitations of the dermis and epidermis, to become blunted, thus reducing dermal–epidermal crosstalk and resulting in thin, fragile skin.53,54 As the goal of fibroblast-seeded DTs is to support optimal re-epithelialization and wound closure, understanding the role of fibroblast age in promoting epidermal development is critical. We hypothesized that neonatal fibroblasts in DTs would improve epidermal proliferation, leading to a thicker, more robust epidermis. To test this hypothesis, neonatal and adult fibroblasts were isolated from healthy donors and primary cultures established. Fibroblast proliferation, DT contraction, and expression of the myofibroblast marker α smooth muscle actin (α-SMA) were assessed. Subsequently, an engineered dermis was made from each strain and a bilayered engineered skin substitute formed by seeding these dermal constructs with primary human adult keratinocytes. Engineered skin anatomy, basement membrane deposition, epidermal differentiation and thickness, and ECM remodeling were assessed.
Materials and Methods
Primary human fibroblasts and keratinocytes
Primary human adult fibroblasts were isolated, as detailed elsewhere,
55
from deidentified surgical discard tissue from healthy adults undergoing elective plastic surgeries (2 male, 2 female, age, 21–42 years; see Supplementary Table S1) with approval of the University of Cincinnati (UC) Institutional Review Board (IRB) (Study # 2013-4582). Patient age, sex, race/ethnicity, and body site were the only demographic details collected for each skin sample. Primary neonatal fibroblasts were isolated from de-identified foreskin samples isolated from newborn males (0–3 months) at the University of Cincinnati Medical Center (UC IRB Study #85-09-17-01X). Fibroblasts were cultured in Dulbecco’s modified Eagle’s (DME) medium (ThermoFisher, Waltham, MA, USA) supplemented with 1% antibiotic-antimycotic (Gemini BioProducts, West Sacramento, CA, USA), 4% fetal bovine serum (FBS; GeminiBio), 5 µg/mL bovine insulin (Sigma, St. Louis, MO, USA), 0.1 mM
Collagen scaffold preparation
Type I collagen scaffolds were prepared as previously described.10,56-58 Briefly, a 10% wt./vol bovine collagen type I (DSM Biomedical, Exton, PA, USA) and hexafluoroisopropanol (Oakwood Chemical, Estill, SC, USA) solution was prepared and electrospun at a distance of 18 cm, a flow rate of 4.0–4.2 mL/h, and a voltage of 28–32 kV. Scaffolds were physically crosslinked in a vacuum at 140°C and then chemically crosslinked in 5 mM N-(3-dimethylaminopropyl)-N’-ethyl carbodiimide hydrochloride (EDC; Sigma) in pure ethanol for 24 h. Subsequently, scaffolds were disinfected by incubation in 70% ethanol, followed by serial rinsing in sterile phosphate-buffered saline (PBS; Sigma) and HEPES-buffered saline (Sigma) and incubation in cell culture medium for at least 30 min prior to cell seeding.
Fibroblast proliferation on 2D substrates and within 3D matrices
Fibroblasts were seeded at 5,000 cells/cm2 onto 24-well tissue culture plates and cultured in supplemented DME for a total of 7 days (6 wells per strain per time point, 4 strains per group). Proliferation was assessed at culture days 1, 2, 3, 5, and 7 using an MTT assay, as previously described.10,56-58 The MTT assay is used to quantify cellular metabolism, which is directly proportional to the cell number and therefore reflects cell proliferation. Briefly, 0.5 mL of sterile 0.5 mg/mL thiazolyl blue tetrazolium bromide (Sigma) in PBS solution was added to each well and the plates incubated for 2 h. Subsequently the MTT solution was removed, and 0.5 mL of methoxyethanol (ChemProducts, Tualatin, OR, USA) was added and agitated on a Belly Dancer (Stovall Life Science, Greensboro, NC, USA). After 2 h, absorbance was read at 590 nm (SpectraMax 190 Microplate Reader; Molecular Devices, San Jose, CA, USA). Fibroblast proliferation in 3D collagen scaffolds was assessed by inoculating a ∼3 × 3 cm scaffold with fibroblasts at 500,000 cells/cm2 (4 strains per group) to create DTs. On culture days 1, 2, 3, 5, and 7, six 4-mm biopsy punches for each cell strain were collected; each biopsy was placed in a well of a 24-well plate, and an MTT assay was run as above but with 3 h incubation and elution times. Readings were normalized by subtracting the average absorbance reading of empty wells. Average absorbance ± standard deviation is reported.
Dermal template contraction
DTs were created as detailed above by inoculating electrospun collagen scaffolds with 500,000 fibroblasts/cm2. After 1 day of culture, six 12-mm biopsy punches were taken from each DT and cultured for 7 days. Images of each DT biopsy were collected on days 1, 2, 3, 5, and 7, and the area was calculated using ImageJ v1.49. The DT area at each time point was normalized to the original area of a 12-mm biopsy punch. For each time point, the six replicates for each group were averaged to obtain a mean value for that strain, and the average for all strains within a group ± standard deviation is reported.
Fibroblast morphology and cytoskeletal organization
DTs were prepared as described above (n = 3 per cell strain) and cultured for 7 days for immunohistochemical assessment. Samples were collected on culture days 1, 4, and 7; fixed in 4% paraformaldehyde; and immunostained for α-SMA (1:250; Invitrogen, #14976082), as previously described. 58 Sections were counterstained with nuclear stain 4′,6-diamidino-2-phenylindole (DAPI; 1:200; ThermoFisher). At least three nonoverlapping images per sample were collected at 20x magnification using a confocal microscope (Leica Stellaris 5, Wetzlar, Germany). The intensity of α-SMA staining was qualitatively assessed with representative images for each group shown at each time point.
Fabrication of engineered skin
DTs were prepared as above. After 4 days of culture, the medium was changed to DME:F12 (Sigma) supplemented with 1% antibiotic-antimycotic (GeminiBio), 1% Insulin-Transferrin-Selenium (Sigma), 0.3% FBS (GeminiBio), 0.5 μL/mL hydrocortisone (Sigma), 1 mM strontium chloride (Sigma), 20 pM triiodothyronine (Sigma), 0.1 mM ascorbic acid-2-phosphate (Sigma), 1 ng/mL bFGF (PeproTech), 5 ng/mL KGF (PeproTech), and 10 μg/mL linoleic acid (ThermoFisher) in preparation for keratinocyte inoculation, as previously described. 56 Keratinocytes were seeded, 1,000,000 cells/cm2, onto each 3 × 3-cm DT (4 per strain, 4 strains per group) and lifted to an air–liquid interface the following day and were cultured for 14 days, with medium changes every 48 h.
Barrier function analysis
Barrier function was assessed by measuring surface electrical capacitance (SEC), which was performed using a NOVA dermal phase meter (DPM 9003; NOVA Technology, Portsmouth, NH, USA). Three measurements were collected from each piece of engineered skin on days 6, 10, and 14 after lifting to the air–liquid interface. Lower SEC values correspond to drier surfaces and thus increased barrier function.59,60 Average SEC ± standard deviation is reported.
Analysis of proliferation of engineered skin
The proliferation of cells within 3D engineered skin substitutes was assessed at 6, 10, and 14 days of lifted culture using an MTT assay, as described above, with 3 h incubation and elution times. Three 4-mm biopsy punches from the engineered skin of each cell strain (4 strains per group) were assessed, and the readings were normalized by subtracting the average absorbance reading of empty wells. Average absorbance ± standard deviation is reported.
Engineered skin anatomy: Skin organization and dermal and epidermal thickness
Three biopsies from each engineered skin group were taken 6, 10, and 14 days after lifting and embedded in OCT Mounting Medium (ThermoFisher). Samples were incubated in OCT at room temperature for 10 min before being placed in a −80°C freezer. Frozen samples were cryosectioned at a thickness of 7 μm and stored in a −20°C freezer. Sectioned slides were fixed in methanol (Macron Fine Chemicals, Centerville, PA, USA) and acetone (VWR chemicals, Radnor, PA, USA) prior to staining. Slides were stained with hematoxylin and eosin (H&E), with brightfield images taken at 2.5x, 5x, and 10x magnification with a light microscope (Leica DM2500 LED, Wetzlar, Germany). Using ImageJ, epidermal thickness was quantified by taking three measurements of three nonoverlapping images of each strain, measuring the distance from the basement membrane to the stratum corneum. Dermal thickness was quantified by taking three measurements of nine images of each strain, measuring the distance from the basement membrane to the bottom of the sample. The average thickness of each group ± standard deviation is reported.
Epidermal proliferation, viability, and basement membrane production
Engineered skin biopsies were collected and frozen in OCT Mounting Medium (ThermoFisher) and stored at −80°C until analyzed. Frozen samples were cryosectioned at a thickness of 7 μm and stored at −20°C until staining. The primary antibodies used for immunohistochemistry consisted of rabbit anti-ki67 (1:200; Abcam, #ab16667, Cambridge, MA, USA), mouse anti-laminin 332 (3:1000; Abcam, #ab78286), mouse anti-keratin15 (1:200; Abcam, #ab80522), rabbit anti-collagen VII (1:200; Abcam, #ab93350), and rabbit anti-collagen IV (1:200; Abcam, #ab6586). DAPI was used as a nuclear counterstain. AlexaFluor®-labeled secondary antibodies, including donkey 488 anti-mouse (1:250; Invitrogen, #A21202), donkey 594 anti-mouse (1:250; Invitrogen, #A21203), donkey 488 anti-rabbit (1:250; Invitrogen, #A21206), and donkey 594 anti-rabbit (1:250; Invitrogen, #A21207), were used to detect primary antibodies. Coverslips were mounted using Fluoromount (SouthernBiotech, Birmingham, AL, USA). Fluorescent images were taken using a confocal microscope (Leica Stellaris 5) at 20x. Two to three sections were analyzed for each of the four strains per group, with at least five nonoverlapping images analyzed per strain.
To quantify keratinocyte proliferation, the total number of Ki67+ cells was counted from nine unique 20× images per strain. The average total per group ± standard deviation is reported.
Collagen, MMP, and a-SMA expression in fibroblasts and engineered skin
Expression of the genes for α-SMA (ACTA2), collagen type 1 A1 (COL1A1), and MMP1, MMP3, and MMP9 was quantified in the source fibroblasts and in engineered skin fabricated using adult and neonatal fibroblasts by using quantitative real-time polymerase chain reaction (qRT-PCR), as previously described. 61 RNA was prepared using TRIzol (Invitrogen, Carlsbad, CA, USA), and the Direct-zol RNA Miniprep kit (Zymo Research, Irvine, CA, USA) was used to isolate the total RNA following the manufacturer’s instructions. The total amount and purity of the RNA were measured utilizing a NanoDrop2000 spectrophotometer (ThermoFisher). The RT2 First Strand Kit (330404, Qiagen, Venlo, Netherlands) was used to synthesize first-strand cDNA following the manufacturer’s instructions. qRT-PCR was performed using the TaqMan Universal PCR Master Mix (4304437, ThermoFisher) following the manufacturer’s protocol. The reaction was analyzed on QuantStudio 3 (Applied Biosystems, Foster City, CA, USA) using QuantStudio design and analysis software V.1.4.1. Gene expression profiles were normalized to the average of the RPL13a and GAPDH housekeeping genes. Expression was quantified relative to the average expression of adult cell strains by using the 2−ΔΔCt method. 62 Similarly, gene expression was quantified for neonatal and adult engineered skin, respectively (n = 4 per group), on culture days 6 and 14 following the procedures described above. Briefly, the tissue was homogenized, and the cells were lysed. Next, the total RNA was isolated and assessed, and the first-strand cDNA was synthesized. qRT-PCR was then performed using the TaqMan Universal PCR Master Mix (4304437, ThermoFisher) following the manufacturer’s protocol. Gene expression was normalized to the average of the GAPDH and RPL13a housekeeping genes and calculated relative to the average expression of the adult group at day 6. Average expression ± standard deviation is reported.
Statistical analysis
For all the experiments, technical replicates were performed in addition to biological replicates. For analysis, the values for technical replicates for each cell strain were averaged to provide a single mean value per strain; these values were then averaged to calculate a single mean value for each group (neonatal or adult), representing the mean of four biological replicates per group. Changes in the outcome with time were assessed using one-way ANOVA, with differences between the two groups at each time point or differences within one group at different time points assessed using Student’s t-test, with statistical significance achieved at p < 0.05. The Shapiro–Wilk normality test and the ROUT outlier test were used to assess the distribution of the datasets. A Mann–Whitney test was used to assess statistical significance in groups that did not follow a normal distribution.
Results
Fibroblast phenotype in 2D and 3D as a function of age
When cultured in 2D, both neonatal and adult fibroblasts significantly increased in number from day 1 to day 7 (p < 0.01, Fig. 1A). Although the mean proliferation values trended higher for neonatal versus adult fibroblasts, there was variability among donor strains, and the difference was only statistically significant on culture day 7 (p < 0.05, Fig. 1A). Similarly, neonatal (p < 0.05) and adult (p < 0.001) fibroblasts cultured in 3D on collagen scaffolds showed increased cell numbers throughout the culture period; however, no significant differences were observed between the groups (Fig. 1B).
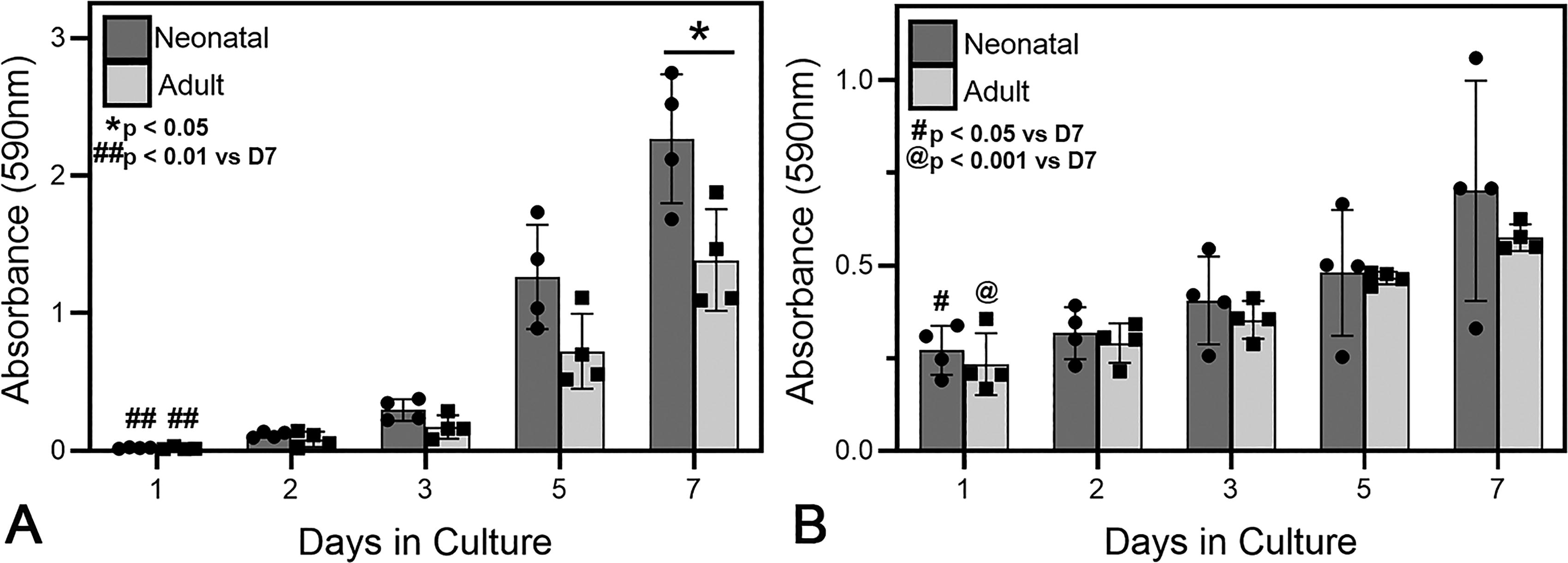
Adult and neonatal fibroblast proliferation as a function of time when cultured on tissue culture polystyrene
Neonatal DTs had a slight increase in size at day 1 and then contracted significantly with time (day 0 vs. day 7, p < 0.001), shrinking to 68.8% ± 6.2% of their original area. A significant reduction in area compared with adult DTs was observed on days 3–7 (Fig. 2A). Adult DTs exhibited modest but statistically significant contraction at day 7 compared with day 0 (p < 0.001). At day 7, adult DTs maintained 91.7% ± 4.2% of their original area, significantly greater than neonatal DTs (p < 0.001, Fig. 2A). Immunostaining for α-SMA showed no difference in the fibroblast size, shape, density, or organization between the groups (Fig. 2B).

Adult and neonatal fibroblast contraction as a function of time when cultured within an electrospun collagen scaffold
Analysis of MMP1, MMP3, MMP9, COL1A1, and ACTA2 mRNA expression in neonatal and adult fibroblasts grown in 2D culture showed no statistically significant differences between the groups (Fig. 3). ACTA2 expression was slightly higher in adult fibroblasts compared with neonatal cells (p = 0.065, Fig. 3). In both groups, there was substantial intragroup variability (Fig. 3).
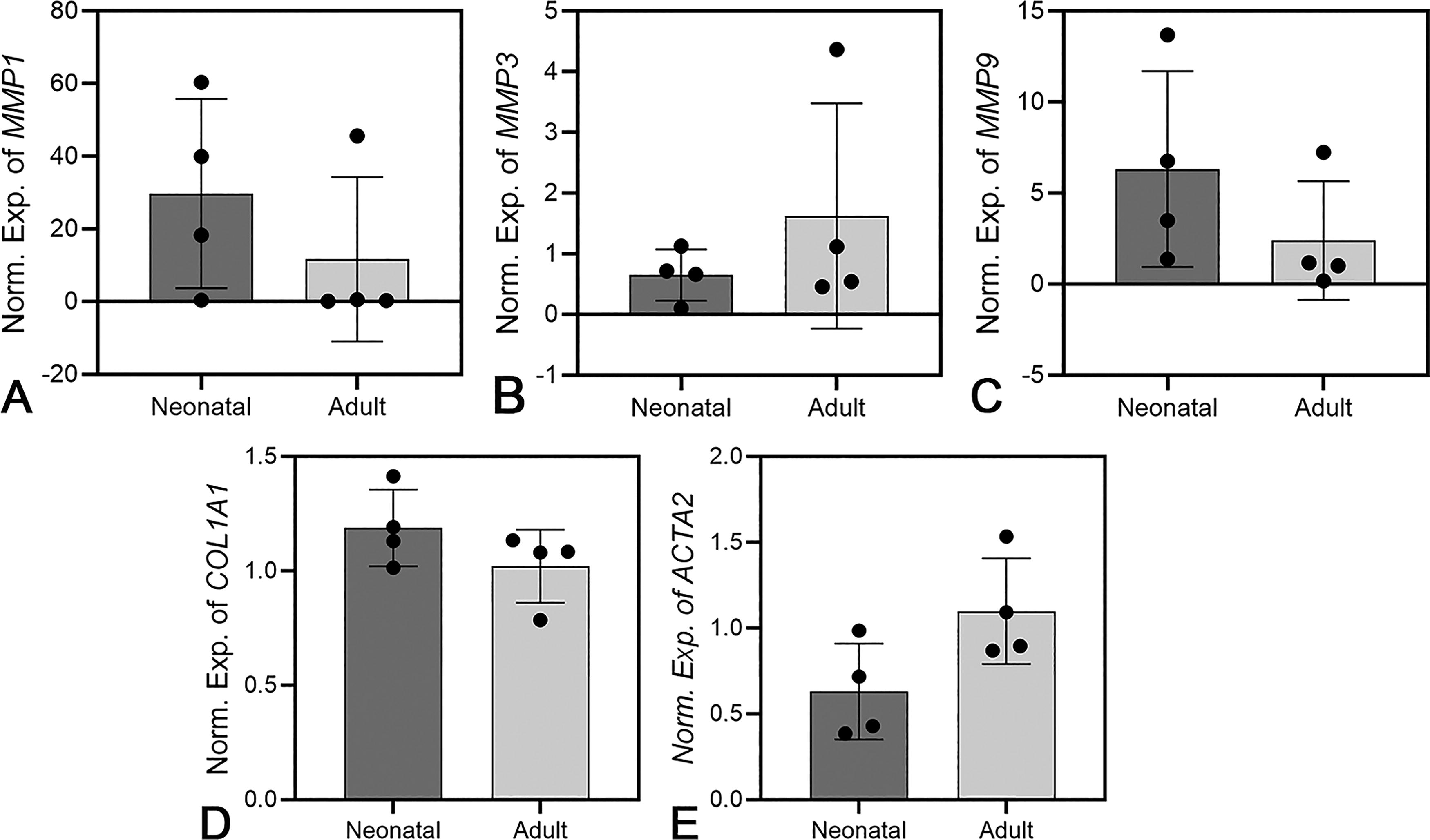
Expression of MMP1
Epidermal regeneration on dermal templates constructed with neonatal or adult fibroblasts
The ability of neonatal versus adult DTs to support epidermal regeneration was examined using the same strain of primary adult keratinocytes for all constructs. Following keratinocyte inoculation, a well-formed, stratified epidermis was observed in both groups (Fig. 4A), with the neonatal group significantly increasing in epidermal thickness from day 6 to day 14 (p < 0.05, Fig. 4B). Quantification of epidermal thickness showed almost equal average values in both groups at 10 and 14 days in culture (Fig. 4B). The average dermal thickness was lower in the adult group at days 10 and 14; however, this was not statistically significant (Fig. 4C). Two of the four adult strains had substantial decreases in thickness with time, whereas the other two strains did not. Both groups exhibited comparable proliferation levels on culture days 6 and 10. By day 14, the engineered skin prepared with neonatal fibroblasts displayed modest but statistically significantly increased proliferation compared with engineered skin prepared with adult fibroblasts (p < 0.01, Fig. 4D). Furthermore, neonatal fibroblasts exhibited a slight but statistically significant increase in proliferation from day 6 to day 14 (p < 0.001, Fig. 4D). Epidermal barrier function, as quantified by measuring SEC, was not affected by the biological age of the fibroblasts, with both groups increasing in surface dryness over 14 days of culture (reflected by lower SEC values) (Fig. 4E). The average adult SEC values significantly decreased from day 6 to day 14, representing enhanced barrier function over time (p < 0.05, Fig. 4E).

Epidermal regeneration on dermal templates constructed with neonatal or adult fibroblasts.
Assessment of keratinocyte proliferation via Ki67+ staining and image analysis revealed similar numbers of proliferating keratinocytes in both groups at culture day 6 that gradually declined over the culture period (Fig. 5A, B). Basement membrane deposition of laminin-332 (Fig. 5A), collagen VII (Fig. 6), and collagen IV (Fig. 7) showed consistent expression at the dermal–epidermal junction (DEJ), independent of the culture time or the biological age of fibroblasts. Keratin-15 showed similar localization patterns along the basal layer of the DEJ in both fibroblast groups (Fig. 6).
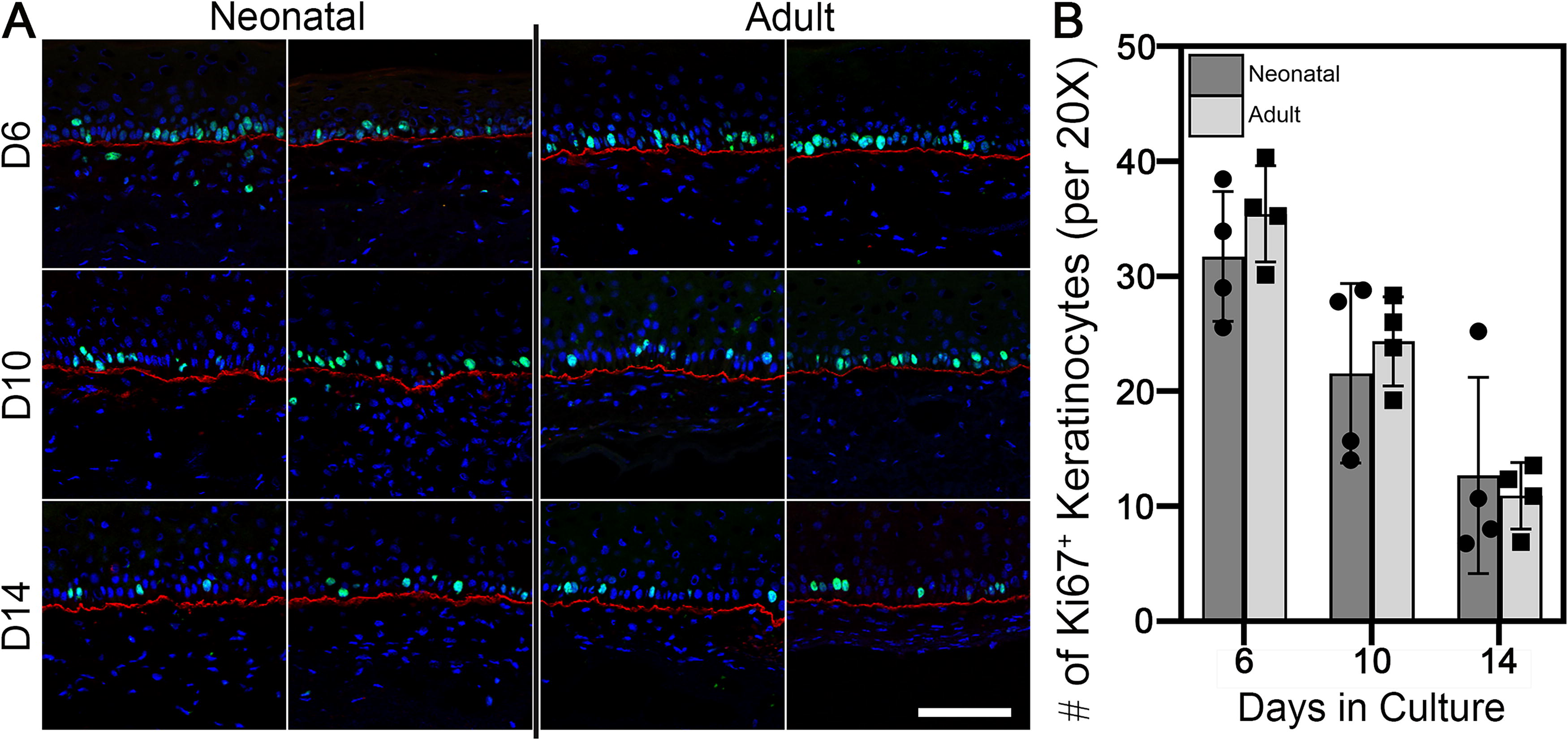
Keratinocyte proliferation and basement membrane formation as a function of time.

Immunohistochemical staining for Collagen VII (red), and Keratin-15 (green) on neonatal and adult engineered skin. Both groups showed similar expression over 14 days of culture. DAPI was used to stain nuclei in all sections. Four strains of adult and neonatal fibroblasts were tested. One representative image from two different cell strains is depicted per group. Scale bar = 100 μm.
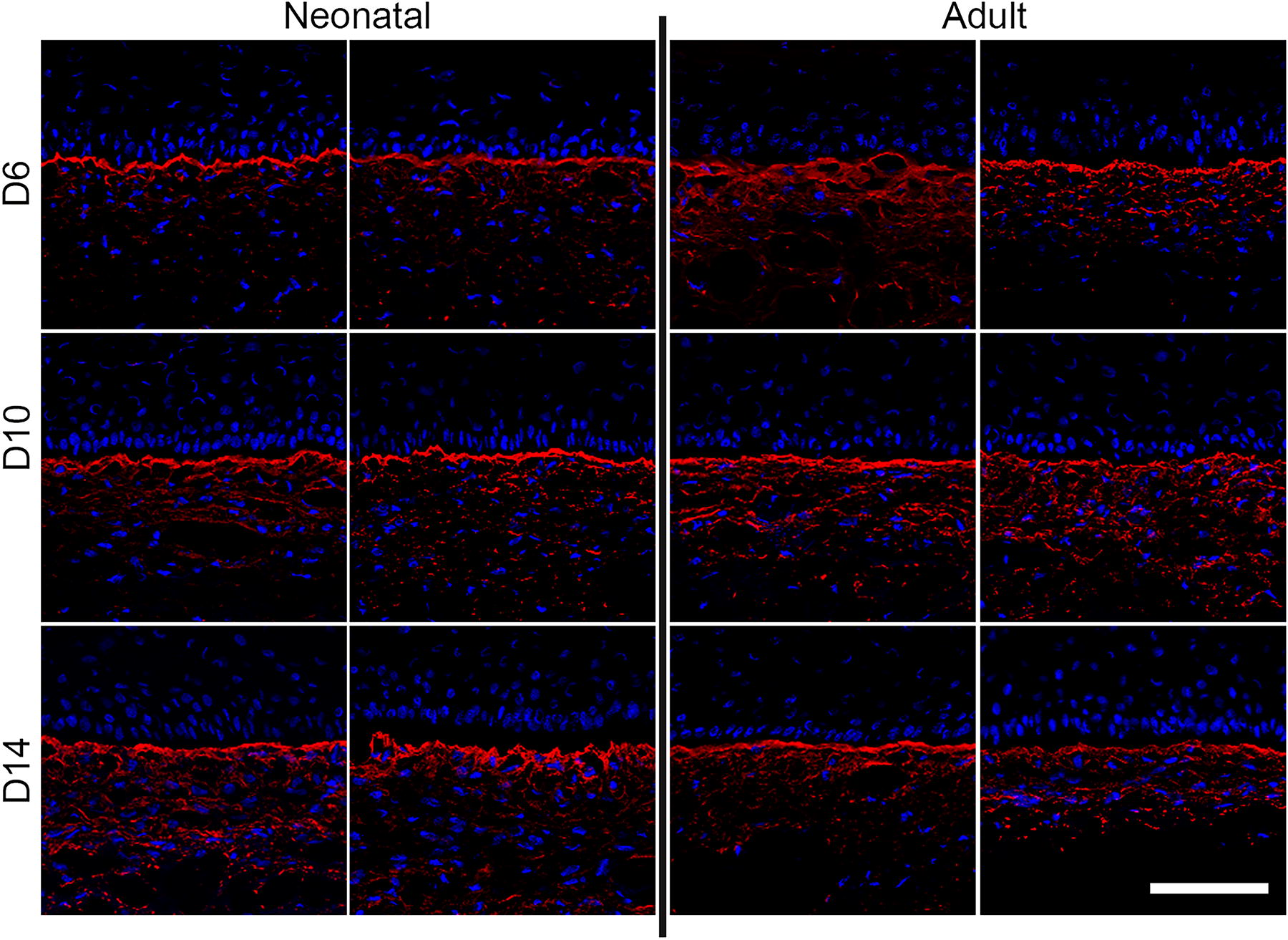
Immunohistochemical staining for Collagen IV (red) on neonatal and adult engineered skin. Both groups showed similar expression over 14 days of culture. DAPI was used to stain nuclei in all sections. Four strains of adult and neonatal fibroblasts were tested. One representative image from two different cell strains is depicted per group. Scale bar = 100 μm.
A comparison of MMP1, MMP3, MMP9, COL1A1, and ACTA2 expressions in engineered skin made with neonatal and adult DTs, inoculated with the same strain of primary human adult keratinocytes, showed no statistically significant differences between the two groups for MMP1, MMP3, MMP9, COL1A1, or ACTA2 at any time point (Fig. 8).
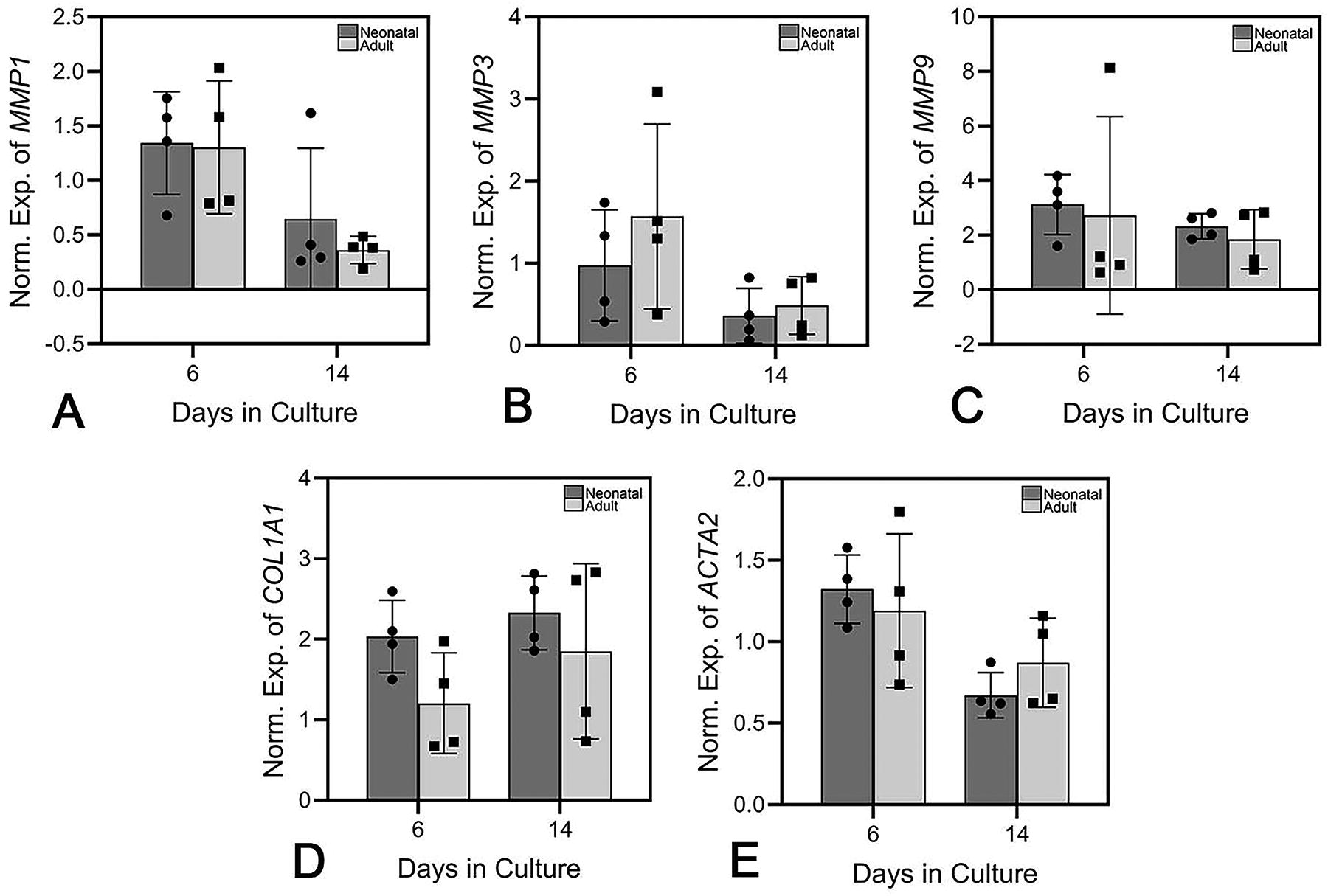
Expression of MMP1
Discussion
To develop allogeneic DTs with the greatest potential to support epidermal regeneration and wound healing, it is essential to understand the role of fibroblast populations in these processes. Neonatal fibroblasts historically have been the primary choice for DTs due, in part, to their availability but also due to reports showing increased proliferation, ECM deposition, and growth factor production compared with adult fibroblasts.25B26 -29,31 However, there are challenges to the general translation of these findings due to the non-native culture environment of 2D tissue culture on plastic, 42 variations in isolation and culture procedures, and differences in the age ranges studied. 35 In the current study, neonatal fibroblasts exhibited higher proliferative activity in a 2D environment, which was similar to a prior report. 35 In contrast, others found only a minimal difference in 2D proliferation between “young” and “old” fibroblasts.31,38,63 However, these studies differed in age demographics and substrates used for culture. Minimal differences in proliferation between young (15.3 ± 2.5 years) and old (83.3 ± 1.5 years) adult fibroblasts were observed in the early passages of 2D culture despite phenotypic differences ex vivo. 63 Similar results were observed in the neonatal and adult age ranges in human 31 and porcine 38 fibroblasts, respectively. In contrast, fibroblasts from donors aged 18–29 years were reported to be more proliferative than fibroblasts isolated from donors over the age of 80 years. 35 Although the differences in the fibroblast phenotype in 2D as a function of age can aid in our understanding of cellular aging, this environment is not similar to the native dermis, and studies should focus on culturing in 3D environments to more closely mimic skin in vivo.
In the current study, when fibroblasts of both age groups (neonatal and adult) were seeded in 3D collagen scaffolds, the DTs displayed comparable proliferative activity; however, neonatal DTs contracted in vitro significantly more than adult DTs. Similarly, a previous study reported that in 3D hydrogels, neonatal fibroblasts were shown to be more contractile than adult fibroblasts. 38 The enhanced contractility of wounds and engineered dermal tissues is the most commonly associated with increased myofibroblast presence and α-SMA production;46,64 however, no increase in α-SMA gene expression in neonatal fibroblasts, or protein production in our neonatal DTs, was observed in the current study. This is consistent with the previous report comparing the contraction of neonatal or adult porcine fibroblast-seeded hydrogels; the neonatal hydrogels were more contracted but did not have increased α-SMA expression (independent of the passage number). 38 The authors attributed the similar α-SMA gene expression among groups to substrate stiffness38,65 and concluded that there was no correlation between contraction and SMA expression.38,66 It is possible that small increases in MMP1 and MMP9 in neonatal fibroblasts may have caused enough degradation of the collagen scaffold to reduce its stiffness, making it easier to contract under similar levels of contractile forces.66-68 While the adult DTs contracted minimally in vitro compared with neonatal DTs, further studies are needed to determine if these trends persist in vivo.
Engineered skin constructs created using neonatal or adult fibroblasts developed similarly, with no significant differences observed in barrier function, epidermal thickness, differentiation potential, or basement membrane formation. These findings are supported by a prior report analyzing young (18–30 years) and older (65–75 years) in vivo skin biopsies, with no significant difference in epidermal thickness or genes encoding laminin-332, collagen IV alpha 1, and collagen VII alpha 1; however, that study did report a difference in the fluorescence intensity when immunostaining these basement membrane proteins directly. 69 When engineered skin, constructed using adult fibroblasts in a collagen gel, was seeded with keratinocytes isolated from either newborn, pediatric, or adult foreskin, similar architecture, barrier function, and lipid density were observed once the skin was brought to the air–liquid interface. 70
In native skin, age-related changes in viability and anatomical structure are well-documented.71-73 However, in the current study, the fibroblast biological age was not shown to have a strong effect on epidermal development. It is possible that in our study, the formulation of the culture medium provided sufficient growth factors to support epidermal development in the absence of any paracrine interactions between fibroblasts and keratinocytes, muting possible differences between the groups. Although the differences observed between neonatal and adult fibroblasts were relatively minor, it is possible that the differences in fibroblast subpopulations may have confounded the results. Previous studies have shown that the dermis contains at least four distinct fibroblast populations, and that functional differences among these populations can persist in culture. 74 These populations can differ in different body regions, 75 can change over time with aging, 76 and can affect interactions with keratinocytes that affect epidermal differentiation. 74 The current study did not seek to identify the subtypes of fibroblasts, but given the differences in the skin types utilized, it is possible that different fibroblast subpopulations were represented in the neonatal and adult cultures. Nevertheless, with minimal differences in 3D proliferation, gene expression, engineered skin architecture, basement membrane formation, keratinocyte stemness, and barrier function, adult fibroblasts support epidermal regeneration and skin development equivalently to neonatal fibroblasts. However, DTs prepared with adult fibroblasts underwent a third less contraction than DTs prepared with neonatal fibroblasts. Thus, adult fibroblasts are a suitable choice for the fabrication of cellular DTs and may enable the treatment of larger wound areas.
Footnotes
Authors’ Contributions
A.C.C. contributed to design, data acquisition, analysis and interpretation, original article draft (lead), and critical revisions. D.S. contributed to data acquisition, analysis and interpretation, and critical revisions of the article. B.N.B. contributed to design and the interpretation and critical revisions of the article. S.A.A. contributed to data acquisition and critical revisions of the article. D.M.S. contributed to conceptualization, interpretation, and critical revision. M.K. contributed to interpretation and critical revisions. H.M.P. guided conception, design, analysis, interpretation, original article draft, critical revisions of the article, and funding acquisition. All authors reviewed and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This project was supported by research grants from the National Institute of Arthritis and Musculoskeletal and Skin Diseases (NIAMS; 1R01AR080946-01A1) (H.M.P.), the National Heart, Lung, and Blood Institute (NHLBI; HL157453) (M.K.), the Shriners Hospitals for Children (SHC; 71003-OHI-22) (H.M.P.), and the Department of Defense (HT9425-23-1-0592) (H.M.P./D.M.S.).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.