
Abstract
In tendons, ligaments, and menisci, collagen fibers running the length of the tissue are the primary source of strength and function. Cells assemble these fibers hierarchically from nanometer-wide fibrils into larger fibers and fascicles, increasing in size throughout development and with mechanical loading. These fibers largely do not regenerate after injury or with repair, limiting recovery options. Engineered replacements are a promising treatment option; however, it remains a challenge to produce the hierarchical collagen fibers essential to tissue strength, limiting their applications. To better repair, regenerate, and engineer these tissues, we must better understand how cells regulate hierarchical fiber formation and maintenance. It is well established that mechanical cues are critical for cell-driven hierarchical fiber formation, which cells sense through several mechanisms, such as integrin-mediated adhesions, cell-to-cell connections, mechanosensitive ion channels, primary cilia, and caveolae. These mechanisms of mechanosensation have been well studied at the fibril scale of collagen organization, but as tissues mature, the loading environment becomes more complex, with cells experiencing increasing secondary shear and compressive loads generated by the developing hierarchical structure. Mechanical cues in this environment are likely sensed through several pathways, each likely playing a role in tissue maturation and injury. There remains a clear gap in our understanding of the later stages of hierarchical fiber formation, which are crucial to better understand since large hierarchical fibers dominate the human musculoskeletal system. Here, we review the role of mechanobiology in hierarchical fiber development and maintenance, and highlight what still needs further research to better regenerate fibers in engineered replacements or in vivo after injury. A better understanding of the mechanisms by which cells form hierarchically organized collagen fibers could help to overcome the limitations of current tissue engineering techniques and help to create functional repairs and replacements.
Impact Statement
Here we review the various mechanisms by which cells within tendons, ligaments, and menisci sense mechanical stimuli and the role that each mechanosensor has been shown to play in hierarchical fiber formation. Furthermore, we highlight potential future research directions to better understand the role of mechanobiology in hierarchical fiber formation. Understanding the mechanisms by which hierarchical fibers are formed, and by which mature tissues are maintained, is crucial to regenerating fibers after injury or in tissue-engineered replacements, and will help to inform the pathophysiological effects of unloading and overloading.
Introduction
Collagen fibers are the primary source of strength in tendons, ligaments, and menisci. In tendons and ligaments, these fibers translate load from muscle-to-bone and bone-to-bone, respectively.1,2 In menisci, these fibers run circumferentially around the tissue, translating compressive loads into horizontal tensile hoop stresses and distributing loads across the knee.3–5 In all of these tissues, cells organize these fibers hierarchically, assembling collagen molecules into aligned nanometer-wide fibrils, which they bundle into larger micron-wide fibers and millimeter-to-centimeter-wide fascicles, maturing with time and loading (Fig. 1).1,4,6–19 Cells mature fibrils and fibers simultaneously throughout development,1,10,20–22 with each hierarchical level contributing to the overall strength of the tissues.10–13,23
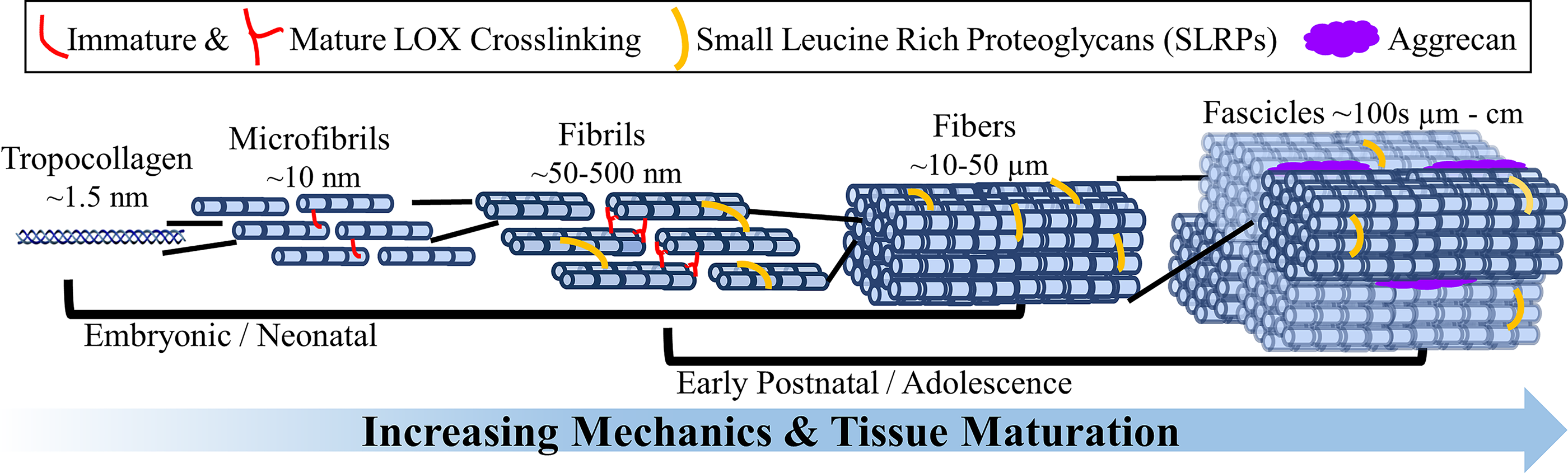
Depiction of hierarchical fiber development and maturation, with relative diameters and localization of lysyl oxidase (LOX)-induced crosslinks and proteoglycans. Cells produce tropocollagen and assemble it into aligned fibrils, which they bundle together into larger fibers and fascicles. The exact timing of when these structures emerge and become mature is still largely unknown and will likely vary between species and tissues. However, current evidence suggests cells continue to mature fibrils and fibers simultaneously throughout early development, with each hierarchical level contributing to the overall strength of the tissue. The fibers are further reinforced by the accumulation of LOX crosslinks, SLRPs, and aggrecan throughout development. SLRPs, small leucine-rich proteoglycans.
Injuries that sever these fibers result in loss of function, pain, and decreased mobility.3,5,24 These injuries have little-to-no healing, ultimately resulting in over 1,230,000 surgeries a year in the United States, including 850,000 meniscus,3,25 130,000 anterior cruciate ligament,26,27 and 250,000 rotator cuff 28 repairs annually. Small tears are often treated via rehabilitation, which typically produces unorganized scar tissue with inferior mechanics.29,30 More severe tears are replaced by autograft or allograft transplants, which have limited availability, risk of immune response, donor-site morbidity, and high rerupture rates.2,3,24,31,32 Collectively, current treatments for torn tendon, ligament, or meniscus, whether repaired or replaced, often fail to restore the native structure and function.2,3,24,30,32 It remains a challenge to regenerate the large hierarchical fibers that dominate these tissues both in vivo and in engineered tissues, limiting repair options. 30
To regenerate hierarchical fibers, we must better understand how cells regulate their formation and maintenance.10,33–35 The mechanisms by which cells mediate hierarchical formation are numerous and not thoroughly understood.34,35 Integrin-mediated actin contractility, mechanosensitive ion channels, cell–cell connections, primary cilia, and caveolae are all known to have differential yet crucial roles.34,35 Here we review the role of mechanobiology in hierarchical fiber development and maintenance, and highlight what still needs further research to better regenerate fibers in engineered replacements or in vivo after injury.
Hierarchical Fiber Development and Maintenance
Collagen fibrillogenesis begins during embryonic development when molecular signaling and cell–cell connections dominate.1,22,36 While embryonic movements contribute to early fibril formation,18,37–43 hierarchical fibers primarily mature during postnatal development and adolescence, when cell–matrix interactions and mechanical cues dominate.9,13,29,43–51 During development, cells deposit aligned collagen fibrils as nanometer-wide bundles between cells, which they grow longitudinally and then expand laterally, increasing individual fibril and bundle diameters to form mature fibrils, fibers, and fascicles.1,10,13–15,20–22,30,36,52,53 How collagen fibrils form and assemble into hierarchical formations is still largely debated, 10 particularly at fascicle length scales, and will likely vary between species and tissues. However, current evidence suggests fibrils and fibers mature throughout development simultaneously,1,10,20–22,54 and with continued maturation, there is increased tissue strength (Fig. 1).10,13,19,30,55
While fiber diameters tend to correlate with mechanical properties, 19 it is not the sole determining factor of tissue strength.10,13 Proteoglycans, enzymatic crosslinks, and different types of collagen play significant and complex roles in fiber maturation. Specifically, small leucine-rich proteoglycans (SLRPs), including decorin, biglycan, fibromodulin, and lumican, and larger proteoglycans, such as aggrecan, accumulate throughout development, with SLRPs influencing fibril length, fibril spacing, and fiber diameter, altering tissue mechanics through changes in collagen structure.1,13,33,48,52,56 Larger proteoglycans, like aggrecan, influence fiber spacing and load distribution between fibers and fascicles in hierarchical fibers (Fig. 1).33,57 However, larger proteoglycans are at higher concentrations in the entheses of tendons, ligaments, and menisci and the inner region of menisci, where hierarchical fibers are less dominant. In these regions, the swelling properties of proteoglycans play a substantial role in the compressive properties.33,35,49,58–62 Enzymatic collagen crosslinks, induced by cellular expression of lysyl oxidase (LOX), also increase throughout development. Specifically, LOX reacts with lysine in collagen to form immature divalent crosslinks that spontaneously condense into mature trivalent crosslinks.63–66 It has been suggested that mature crosslinks provide the primary strength in fibers.38,63,65,67,68 Furthermore, while type I collagen is the primary collagen in fibers, smaller amounts of minor collagens, including types III, V, XI, XII, and XIV, and structural proteins such as fibronectin also contribute, helping to regulate fiber diameter, spacing, length, and fibrillogenesis.69–82 While it is largely unknown how cells regulate proteoglycans, LOX, or collagen types to develop fibers, inhibition of any results in altered organization and reduced tissue mechanics.1,38,56,66,69–74,76,78,79,82–84
Additionally, fibers and fascicles exhibit wave-like morphologies called crimp. Crimp is fundamental in force transduction by responding first to load, forming the toe region of the stress–strain curve.20,85,86 Crimp appears during early development20,87 and increases in wavelength with time and load.20,88–90 How crimp forms is largely unknown; however, it is suggested to be highly regulated by the mechanical environment.87–89,91
Mechanical Regulation of Hierarchical Fiber Formation, Homeostasis, and Repair
Loading is essential for tendon, ligament, and meniscus development, homeostasis, and repair, but how loading influences cellular regulation of these tissues is poorly understood.29,35,43,92 During development, cells convert mechanical stimuli into biochemical signals via mechanotransduction to develop hierarchical fibers. These mechanical cues are transduced to the cell nucleus via multiple signaling pathways, such as Hippo signaling through YAP/TAZ, Rho/ROCK signaling, and the LINC complex, resulting in nuclear and chromatin deformations to alter gene expression (these pathways are outside the scope of the article, but refer the reader to the following cited reviews for further information).93–95 Generally, as loads increase throughout development, cells produce larger fibers and fascicles, increase crosslinking, and ultimately produce stronger tissues; however, the extent of hierarchical development varies between tissues.15,17,29,45,96 If mechanical loads are absent during development, tissues fail to mature, resulting in underdeveloped fibers and impaired expression of SLRPs, LOX, and collagens.9,13,18,29,35,43,45,51,75 Once fibers have formed, exercise can increase fiber maturation, producing stronger tissues, but too much exercise/load can lead to degeneration and injury.9,29,35,48
The mechanisms by which cells sense mechanical cues are complex and likely change throughout hierarchical fiber formation (Fig. 2). It is established that cells initially form aligned fibrils largely through cell–cell connections and cellular contraction forces, highly regulated by integrins, focal adhesion kinase (FAK), and the actomyosin network,20,34,36,91,97–101 with cells remodeling the matrix to achieve a tensional homeostasis.34,98,102,103 Tensional homeostasis is a set point of tensile strain that cells attempt to maintain.98,103 When this is lost, cells respond with an injurious, catabolic response.88,98,103 However, stimulation that challenges tensional homeostasis can promote an anabolic response, increasing tissue maturation.29,45,96,104 In fact, tensile loads, including slow growth and cyclic muscle activity, are thought to drive development by applying uniaxial stretch to fibers, which is translated to cells via focal adhesions (FAs), altering their tensional homeostasis and stimulating further development.9,29,43,45,51,105,106 However, while cells predominantly receive tensile strains transferred via cell–cell connections or aligned fibrils in early postnatal development, as tissues mature, this environment becomes more complex, with cells increasingly experiencing higher degrees of secondary shear and compressive loads generated by the developing hierarchical structure (Fig. 2).44,107 Throughout development, cells organize themselves in rows, joined on either end by cell–cell adhesions, and form mature fibers around themselves.100,108 Primary cilia align with tension, and cells alter the expression of mechanosensors such as mechanically gated ion channels and FA-related proteins.104,109,110 Mechanical cues are sensed through several pathways, each likely playing a role in cellular response to load during fiber development and injury.
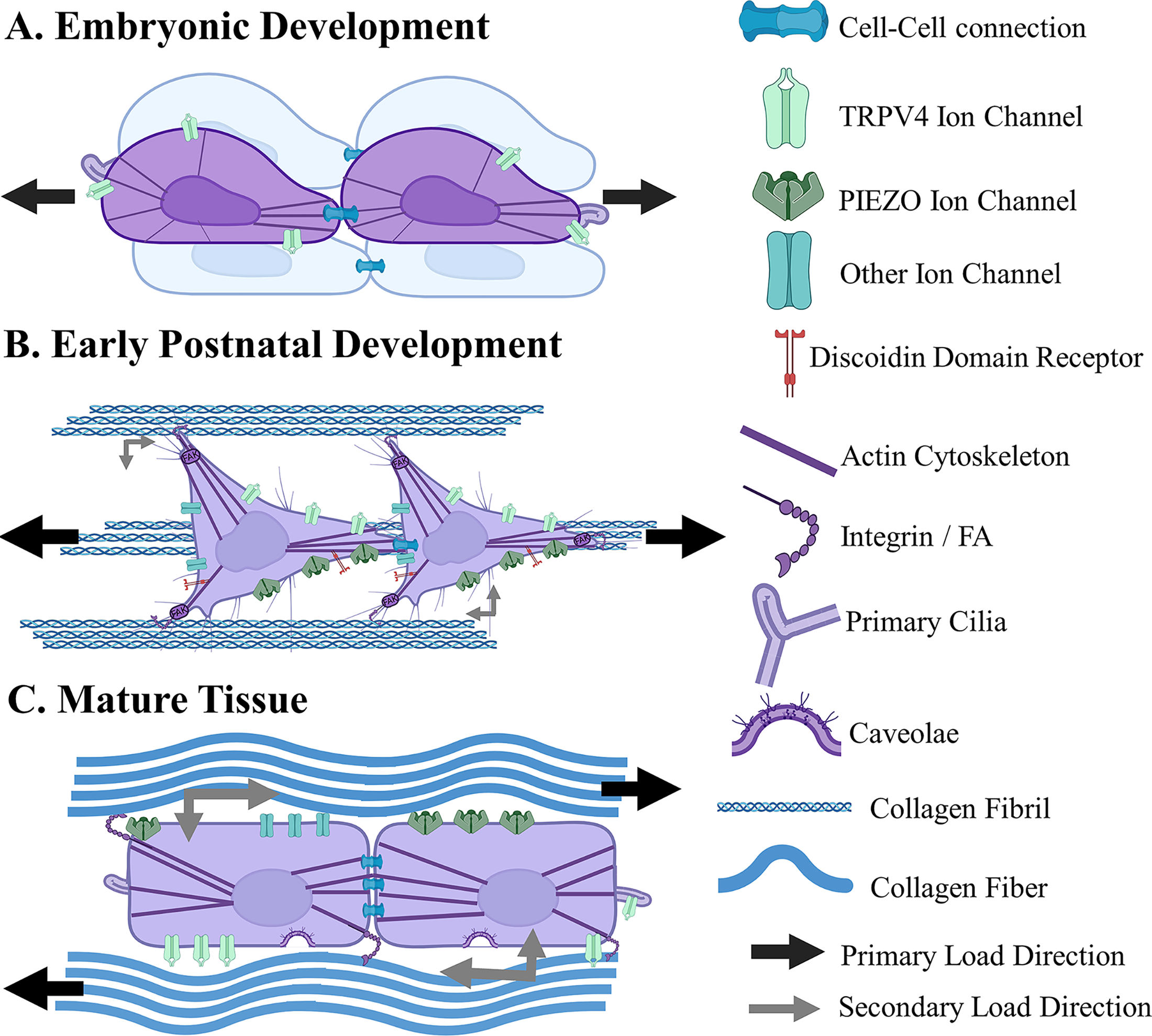
Depiction of major cell mechanosensation pathways and the evolving mechanical environment throughout hierarchical fiber development.
Effects of Tensional Homeostasis/Cytoskeleton Disruption on Collagen Degradation, Fibril Formation, and Tissue Maturation
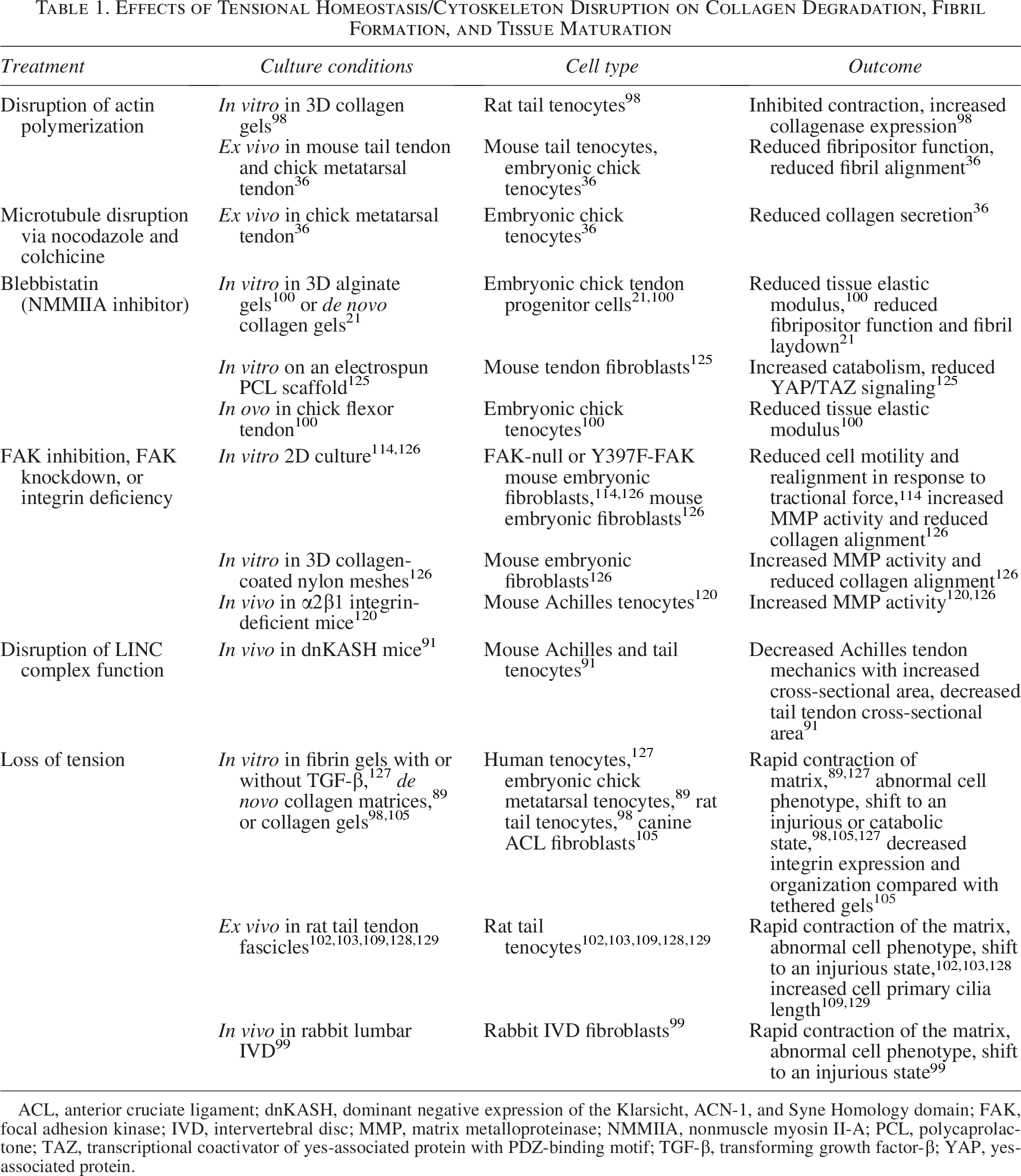
ACL, anterior cruciate ligament; dnKASH, dominant negative expression of the Klarsicht, ACN-1, and Syne Homology domain; FAK, focal adhesion kinase; IVD, intervertebral disc; MMP, matrix metalloproteinase; NMMIIA, nonmuscle myosin II-A; PCL, polycaprolactone; TAZ, transcriptional coactivator of yes-associated protein with PDZ-binding motif; TGF-β, transforming growth factor-β; YAP, yes-associated protein.
Actin-Cytoskeletal Tension and Integrin-Mediated Adhesions
While collagen remodeling is regulated by numerous signals, none are as well studied as actin-mediated mechanotransduction. Cells reach tensional homeostasis via regulation of the actin-cytoskeleton, largely through actin-to-extracellular matrix (ECM) connections called FAs.103,111–113 FAs are theorized to be one of the primary mechanisms by which cells in hierarchically organized tissues sense forces, though much of this work has been in 2D or ex vivo systems.103,105,111,114–120 These mechanosensitive macromolecular structures are composed of hundreds of proteins and act as a direct signaling pathway between the ECM and the nucleus.111,121,122
FAs are primarily mediated by integrins, FAK, and the actin-cytoskeleton.112,116,118,123 FAs first form when integrins bind to ECM.105,111 The cytoplasmic tail of integrins initiates a cascade of protein binding, including FAK, and the resulting complex of proteins attaches to the actin-cytoskeleton, providing a direct mechanical signal from ECM to nucleus.111,112,124 Several components of this connection are known to have mediatory roles in fibril production and alignment (Table 1). 36 For example, nonmuscle myosin II-A (NMMIIA) of the actin-cytoskeleton contributes to collagen synthesis by forming cell extensions called fibripositors.21,36 Fibripositors are extensions of the cell membrane that align with the axis of tension to lay down fibrils during embryonic development.21,36 Disruption of these actin filaments with cytochalasin D (CytoD), 36 inhibition of NMMIIA via blebbistatin, 100 and inhibition of the LINC complex 91 during embryonic development all result in reduced collagen organization and loss of tendon strength.
Cytoskeletal tension also plays a significant role in mature tissues. It is established that ex vivo untethered tendons, or tethered tendons treated with CytoD to disrupt actin polymerization, result in loss of cytoskeletal tension (Table 1).89,100,103,125,130–132 In both cases, cells lose their tensional homeostasis and respond injuriously, increasing catabolic gene expression and matrix degradation,98,99,103,127,128,130 resulting in reduced collagen secretion, alignment, and tissue strength.36,103,125,130–132 Treatment with blebbistatin also reduces YAP/TAZ activation, suggesting that actin-cytoskeletal tension may activate anabolic genes via this pathway. 125 This work demonstrates that the actin-cytoskeleton provides a mechanical cue to cells in the form of prestress to the ECM, but also a mechanism by which cells sense dynamic cues from the environment.133,134
Harnessing actin-cytoskeletal tension in engineered tissues
The role of actin-cytoskeletal contraction in fibril formation is so critical that efforts to produce aligned collagen fibers in engineered tissues have primarily focused on harnessing cellular contraction. In these culture systems, cells are seeded into collagen or fibrin hydrogels, and boundary conditions are applied, often using pins or clamps to mimic the natural attachment points of these tissues (Table 2).40,105,133,134,136–148,150,151 As cells contract the gel through FAs to develop tensional homeostasis, they sense higher tension between the boundary constraints, prompting formation of aligned fibrils along the primary axis of stress.105,133,140,141,149,152 This actin-mediated ECM alignment and remodeling is notably similar to embryonic fibrillogenesis,40,105,136,137 with colocalization of collagen types I, III, XII, and XIV, a parallel fibronectin network, and fibripositors. 40 The boundary conditions allow cells to return to their tensile homeostatic set point. 103 However, these systems have primarily consisted of low-density collagen or fibrin gels (1–5 mg/mL), which have not been shown to support larger fibers and fascicle development. We have found that when using higher concentrations of collagen (10–20 mg/mL), cells form native-sized fibers and early fascicles.68,144,145,149 As collagen concentration increases, gross cellular contraction of the gel decreases, and tensile moduli significantly increase.144,153,154 We hypothesize these attributes of high-density collagen gels provide the time and mechanical cues needed to support cellular development of larger hierarchical fibers.
Effects of Static Boundary Conditions on Collagen Fiber Formation and Maintenance In Vitro

MMP1, matrix metalloproteinase 1.
Increasing maturation with dynamic stimulation in engineered tissues
As discussed previously, dynamic stimulation is critical to fiber development in vivo and similarly in engineered tissues. A great deal of research has investigated the application of dynamic loading to drive fiber formation in vitro (Table 3).105,106,150,155,156,161,163–165,168–172 The most studied mechanical cues are those that mimic in situ cues, namely cyclic strain similar to what tendons, ligaments, and menisci undergo when walking or running. Cyclic strain upregulates integrin expression and cellular alignment, implying it is sensed through the integrin/FAK pathway.105,157,171,173 This change in integrin expression corresponds to changes in collagen organization at the micro- and macroscopic scale, 105 with cyclic load often driving collagen turnover and matrix maturation,105,155,156,163,164 resulting in increased fibril, fiber,136,161,164,165 and crimp formation.164,165 Mechanotransduction has been less explored in the meniscus, and since the meniscus primarily experiences compressive loads that are translated into tensile hoop stresses, mechanosensors may have different roles in meniscal fibrochondrocytes. However, it has been shown in engineered menisci that dual compressive/tensile loading regimes, which convert compressive loads into tensile hoop stresses, further enhance fiber formation and zonal matrix formation over compressive loading alone, demonstrating the critical role of tensile cues.154,168,169,174
Effects of Dynamic Load on Hierarchical Fiber Formation in Engineered Tissues and Fiber Maintenance in Mature Tissues
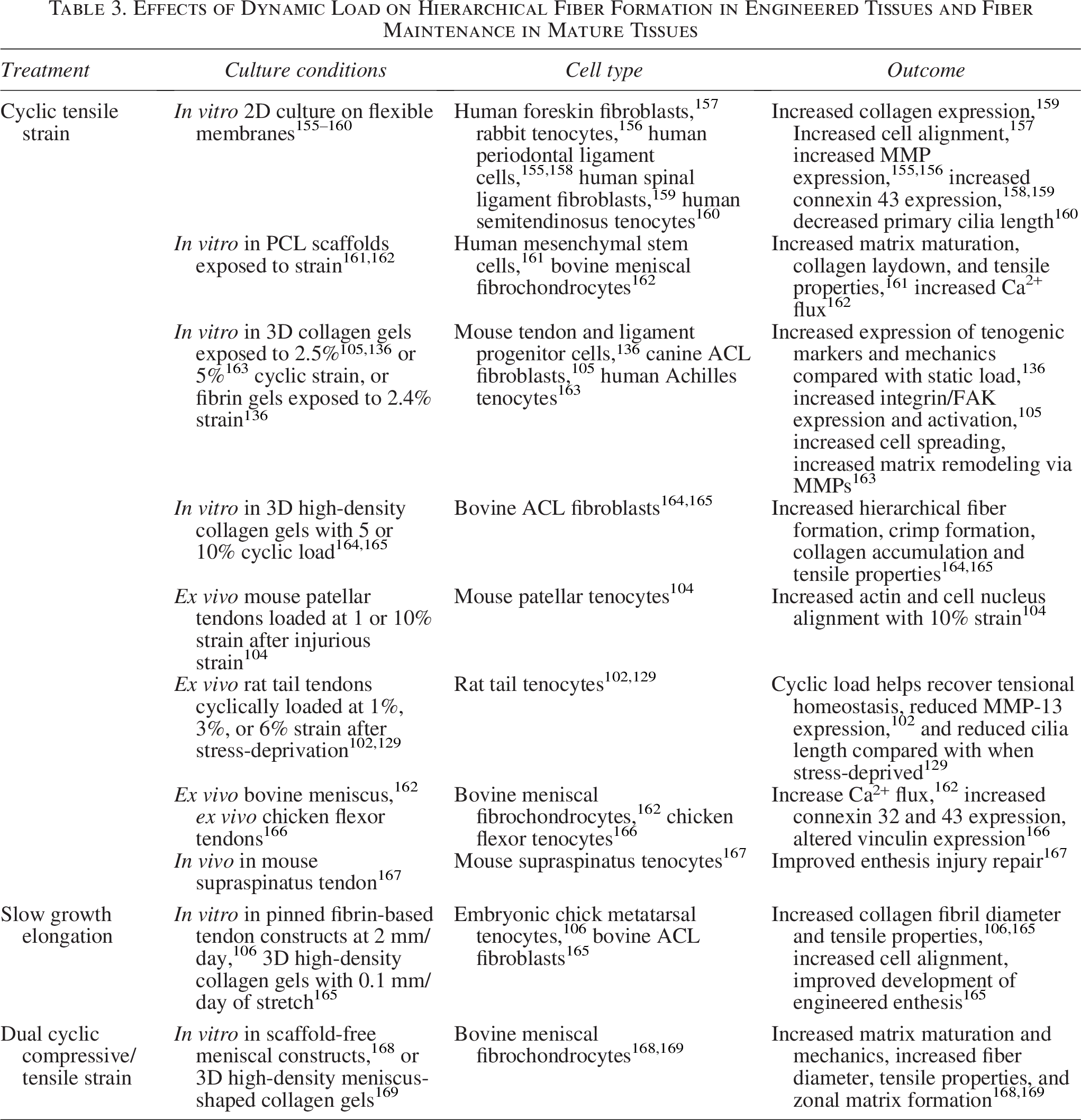
As discussed previously, while cells may predominantly receive tensile strains transferred via aligned collagen fibril bundles or cell–cell connections in early development, as tissues mature, this environment becomes more complex, with cells likely experiencing higher degrees of secondary shear and compressive loads generated by the developing hierarchical structure (Fig. 2).150,175,176 Furthermore, as cells align collagen fibrils, they stiffen the tissue, activating further integrins and other mechanical signaling pathways such as mechanically gated ion channels.126,142,162 In fact, in conditional FAK knockout mouse models, FAK inhibition during postnatal development did not abolish collagen organization in tendons and ligaments, and instead only reduced collagen alignment by 20% and collagen density by 40% in the periodontal ligament, 126 and reduced size and fibril diameters in flexor, Achilles, and tail tendons, but tensile properties were largely maintained. 119 This demonstrates that while FAK- and integrin-mediated mechanotransduction play a significant role in collagen synthesis and alignment, other mechanisms beyond integrin-based contraction are needed during later stages of hierarchical fiber formation.
Cell–Cell Adhesions
Cell–cell adhesions are highly expressed in tendon, ligament, and meniscus cells and are crucial to tissue development and tensional homeostasis.158,177–184 In embryonic development, cells are tightly packed and communicate via cell–cell junctions mediated by cadherins and connexin-mediated gap junctions (Fig. 2).100,108,182,185 As tissues mature, cells are arranged in long rows joined together by cell–cell adhesions.100,108,166,182
Gap junctions are crucial to fibril synthesis and alignment, with inhibition often increasing fibril dispersion and upregulating inflammatory genes.173,183,186,187 In particular, connexin (Cx)-32 and -43 are established to help regulate collagen secretion and to be mechanosensitive, but with distinct roles (Table 4).158,159,173,180 For example, in chick flexor tenocytes, signaling through Cx32 stimulates collagen secretion in 2D, while Cx43 signaling inhibits collagen secretion, both in static and dynamically loaded cultures. 180 Furthermore, while Cx32 and Cx43 are upregulated in response to load, strains beyond physiological levels (>8% strain) have been shown to suppress gap junction signaling, hampering intercellular communication.135,188,189
Role of Cell–Cell Adhesions in Response to Load and in Collagen Maturation
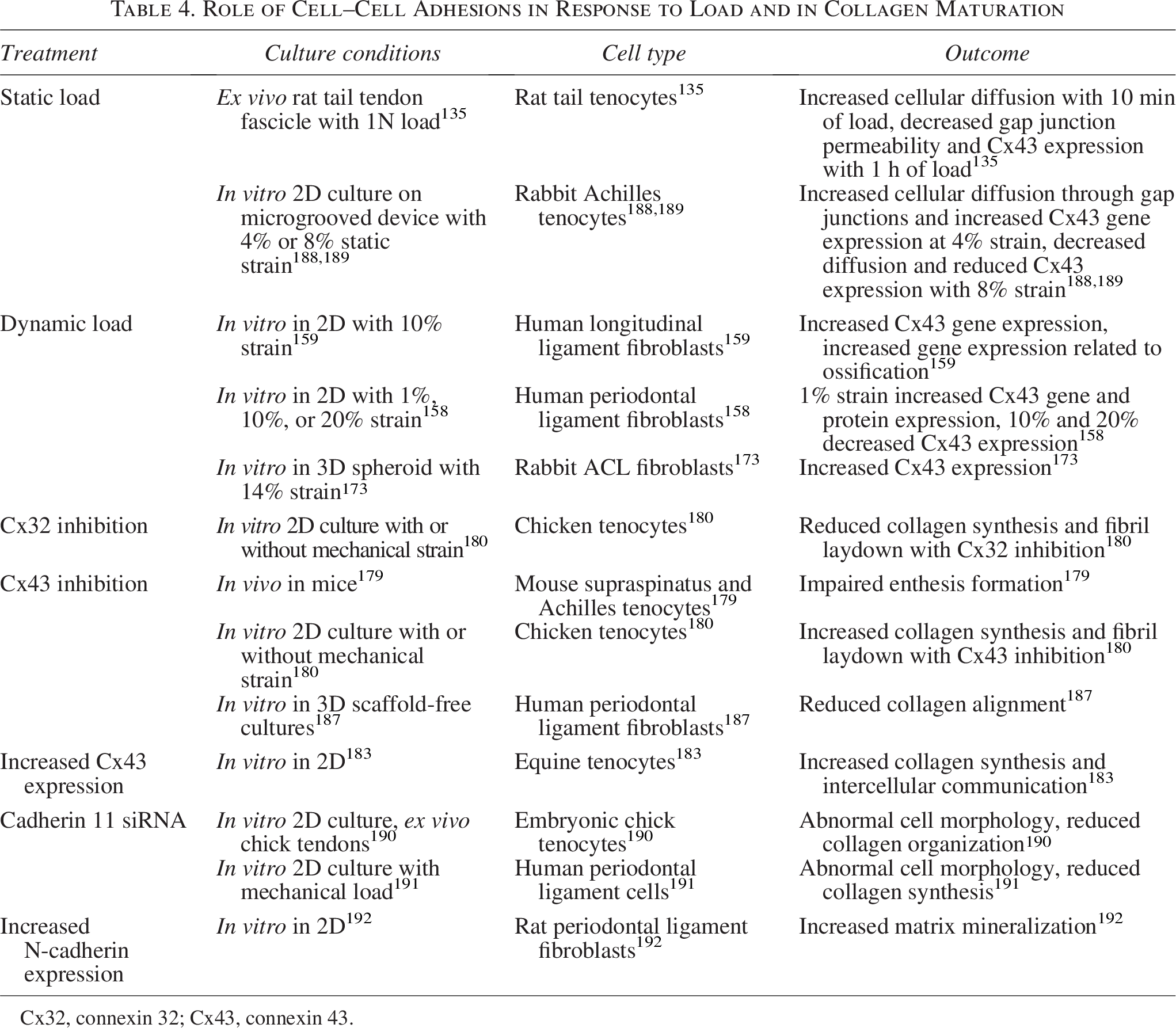
Cx32, connexin 32; Cx43, connexin 43.
Gap junctions are also critical in enthesis formation. The enthesis is the fibrocartilaginous transition region in tendons, ligaments, and menisci that translates loads between elastic ligamentous tissue and stiff bone. 179 In a murine model, deletion of Gja1, which encodes for Cx43, results in reduced cell alignment and mineralization in the enthesis compared with wildtype (WT) controls. 179 This correlates well with studies in periodontal ligament cells (PDLCs) and other ligament fibroblasts, where Cx43 was shown to have a significant role in ossification.158,159,193
Another mechanism of force transduction in cell–cell adhesions is cadherins, transmembrane proteins that form cell–cell junctions. These proteins are highly expressed in musculoskeletal tissues, mechanosensitive, and have a role in regulating collagen secretion.190,191 In particular, cadherin 11 is expressed in tendon and ligament fibroblasts and suppressed by load.190,191 Furthermore, knockdown of cadherin 11 in these tissues reduces collagen synthesis 191 and collagen organization. 190 N-cadherins have also been shown to be expressed in PDLCs and are more highly expressed when differentiated to form mineralized nodules, again suggesting that cell–cell connections have a role in ossification. 192
Mechanosensitive Ion Channels
Due to the large array of cellular functions regulated by calcium, stretch-activated calcium-permeable channels in the cell membrane are suggested to be significant regulators of musculoskeletal development.162,194 Traditionally, these mechanically activated ion channels were thought to be primarily stimulated by external loads; however, recent work has reported that these channels have significant roles in regulating cellular contraction forces as well. 195 Cellular contraction or variations in matrix stiffness can activate ion channels, increasing intracellular calcium and altering cellular mechanosensitivity.194–197 This allows cells to respond to a continuum of mechanical stimuli, often stimulating downstream events distinct from FAK-actin contractility.198–200
While there are many ion channels, transient receptor potential vanilloid 4 (TRPV4), Piezo1 and 2, and voltage-gated channels are reported to have significant and independent roles in the development of many connective tissues194,198,201,202 and are found to be highly expressed in fibrous tissues, including tendons, ligaments, and menisci.175,198,203–206 These mechanosensitive channels respond to a wide range of mechanical cues, including passive stimulation from cellular contraction and dynamic stimulation from forces such as stretch, compression, shear flow, and osmotic stress.175,195–197,199,200,207,208 Furthermore, while all respond to a wide range of mechanical cues, they appear to differ in the strength of load needed for activation and often stimulate distinct downstream signaling (Table 5).198–201,207
Role of Ion Channels in Cell Signaling, Collagen Fiber Formation, and Tissue Maturation
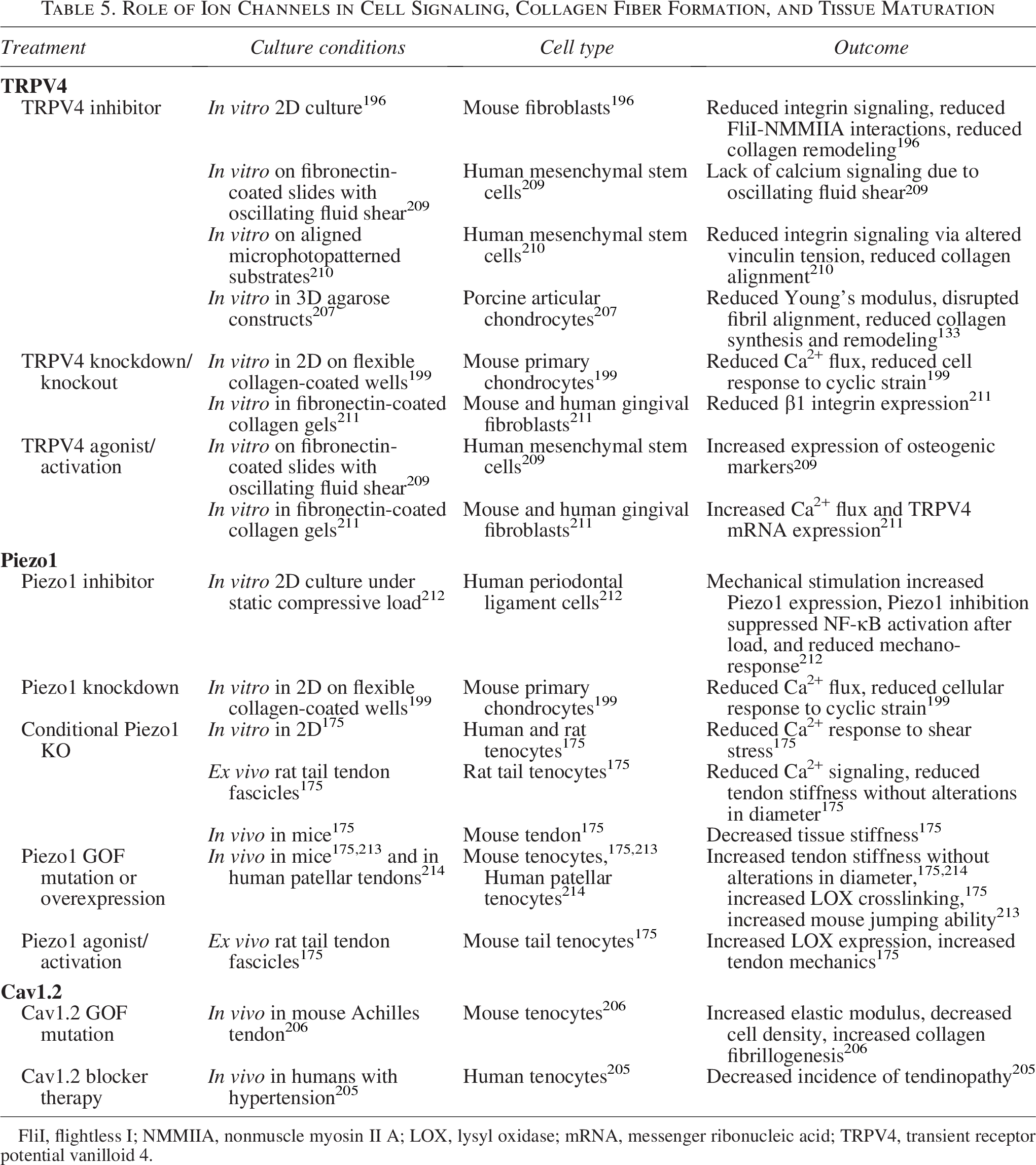
FliI, flightless I; NMMIIA, nonmuscle myosin II A; LOX, lysyl oxidase; mRNA, messenger ribonucleic acid; TRPV4, transient receptor potential vanilloid 4.
TRPV4
Transient receptor potential (TRP) channels are a group of ion channels that are crucial in cell signal transduction, sensing various stimuli such as chemical, temperature, and mechanical cues. 202 Studies on 2D surfaces have found TRPV4, in comparison with several TRP family members, mediates the Ca2+ influx needed to develop cellular contraction and is required for collagen remodeling in fibroblasts and human mesenchymal stem cells (hMSCs).194,196,210 Furthermore, TRPV4 has garnered interest since it has been shown to be highly expressed in musculoskeletal tissues,199,204,210,215 to play a significant role in fibril formation, and to respond to mechanical cues that tendons, ligaments, and menisci experience in situ (3–8% strain).199,201,207,216,217
TRPV4 activation has been shown to regulate fibril laydown and collagen remodeling in several cell types (Table 5).194,196,210,211,218,219 More specifically, Ca2+ transfer through TRPV4 assists in a molecular chaperone function during protein synthesis, 220 regulating early alignment of fibrils and the creation of fibripositors in dermal fibroblasts and stem cells.196,210 In a microphotopatterned system, TRPV4 was critical during the initial stages of collagen matrix assembly in hMSCs, contributing to fibril synthesis and alignment. 210 However, its activity significantly decreased once aligned fibrils were produced, 210 suggesting its activity once fibrils are formed may be detrimental and it has less of a role during fiber and fascicle formation.
While TRPV4 mechanotransduction has a role in fibril formation independent of integrin-FAK-cytoskeletal signaling, it also helps regulate actin-cytoskeleton function, activating various proteins in the integrin signaling cascade.198,221–224 In particular, calcium flux through TRPV4 affects NMMIIA in cell extensions194,196 and modulates tension across vinculin, significantly impacting FA formation and collagen remodeling. 210 Collectively, TRPV4 plays a significant role in FA function and cell–ECM interactions, including matrix synthesis, cellular contractility, MMP-1 activity, and cytokine production194,218,225; however, this work has primarily been in 2D, and its role in organization beyond fibril alignment is largely unknown.
Piezo
Another family of ion channels that has received significant attention is the Piezo family, specifically Piezo1 and Piezo2, with Piezo1 suggested to be a critical regulator of LOX expression in mature tendons. 175 Similar to TRPV4, Piezo1 is activated by cellular contraction, with Piezo1 activation increasing as contraction forces increase,195,197,200,226 suggesting it is a higher-load sensor than TRPV4. Furthermore, Piezo channels respond to a wide range of mechanical cues, such as osmotic stress, compression, tension, and shear stress.197,227–231 A consensus on the mechanism of Piezo activation has yet to be reached, though it is well established that Piezo channels are activated by stretch and shear stress and are highly expressed in bone, cartilage, tendons, and ligaments (Table 5).198,212,232–235 In cartilage, Piezo1 and Piezo2 are believed to activate at different tensile strains, with Piezo2 activating at higher strains than Piezo1,199,200 and have been shown to have synergistic effects. 200 While this work was in chondrocytes, the same is likely true for cells in other musculoskeletal tissues where Piezo1 and 2 are both expressed.
In mature tendons, Piezo1 is reported to be a critical sensor of shear stress, responding to the level of stress generated by fiber sliding during physiological strains. 175 Recent work in mouse and human tenocytes has found that Piezo1 is associated with increases in tendon strength and LOX expression, without changes in tendon or fibril diameter, suggesting Piezo1 activation increases LOX crosslinking.175,214,236 Furthermore, mice with a Piezo1 tendon-specific knock-in were found to have stiffer Achilles tendons, resulting in increased storage modulus, increased jumping ability, and faster running speeds compared with WT mice. 213 Humans carrying the Piezo1 GOF mutation, E756del, also had no differences in tendon size compared with healthy controls but had significantly increased jumping ability 213 and patellar tendon stiffness. 214
As already discussed, mechanosensitive ion channels are activated by external mechanical cues, with both TRPV4 and Piezo1 responding to stretch, compression, shear, and osmotic stress.175,195–197,199,200,207,208 In fact, mechanical stimulation of tendon, ligament, and meniscus significantly upregulates calcium signaling,162,175 suggesting a role of these calcium stretch channels. Furthermore, TRPV4 and Piezo1 are both upregulated in response to load but respond to different ranges of load with differential gene expression.142,198,199,201 Specifically, in chondrocytes, it has been shown that TRPV4 responds to low strain, while Piezo1 responds to high strain, allowing cells to respond to a continuum of mechanical cues.199,200,235
Voltage-gated channels
Voltage-gated calcium channels can also be activated as part of the cell’s response to mechanical stimulation. CaV1.2 is expressed in tendons and activates downstream of Piezo, as a result of the rapid influx of cations through Piezo1 and 2. 206 It has been suggested that CaV1.2 helps regulate collagen fibrillogenesis and tendon mechanics, since mice with CaV1.2 GOF mutation have tendon hypertrophy, with increased tendon stiffness, ultimate tensile strength, and increased expression of ECM proteins (Table 5).205,206
While the work to understand the role of mechanosensitive ion channels in tendons, ligaments, and menisci is promising, the scope of work is limited. There is still much to understand about the roles of these mechanosensitive ion channels in hierarchical fiber formation and maturation.
Other Mechanosensitive Cell Membrane Sensors
Primary cilia
The primary cilia of cells are microtubule-based organelles that sense force and transduce signals via the hedgehog pathway (Table 6). They are believed to be involved in the regulation of bone and cartilage245,246 and are expressed in tenocytes, ligament fibroblasts, and meniscal fibrochondrocytes.184,238,247–251 Furthermore, they are found primarily in the outer region of the meniscus where hierarchical fibers dominate. 184 Primary cilia orient themselves parallel to the length of the collagen fibers and the direction of primary load, 110 lengthening with reduced load to increase mechanosensation and shortening with cyclic loading.109,129,160 Cilia length is also affected by hypoxia, shortening in hypoxic conditions, leading to a decreased response to mechanical strain. 237 This suggests primary cilia have a smaller role in adult tissues, where tendons, ligaments, and menisci are in a more hypoxic environment with relatively constant cyclic load.237,252 However, cilia may have a key role in wound healing and tissue generation, as stress-deprived tenocytes have been shown to lengthen their cilia as they attempt to re-establish tensional homeostasis both in 2D flexible membranes and ex vivo rat tail tendons.109,129,160
Role of Mechanosensitive Cell Membrane Sensors in Cell Signaling, Collagen Fiber Formation, and Tissue Maturation
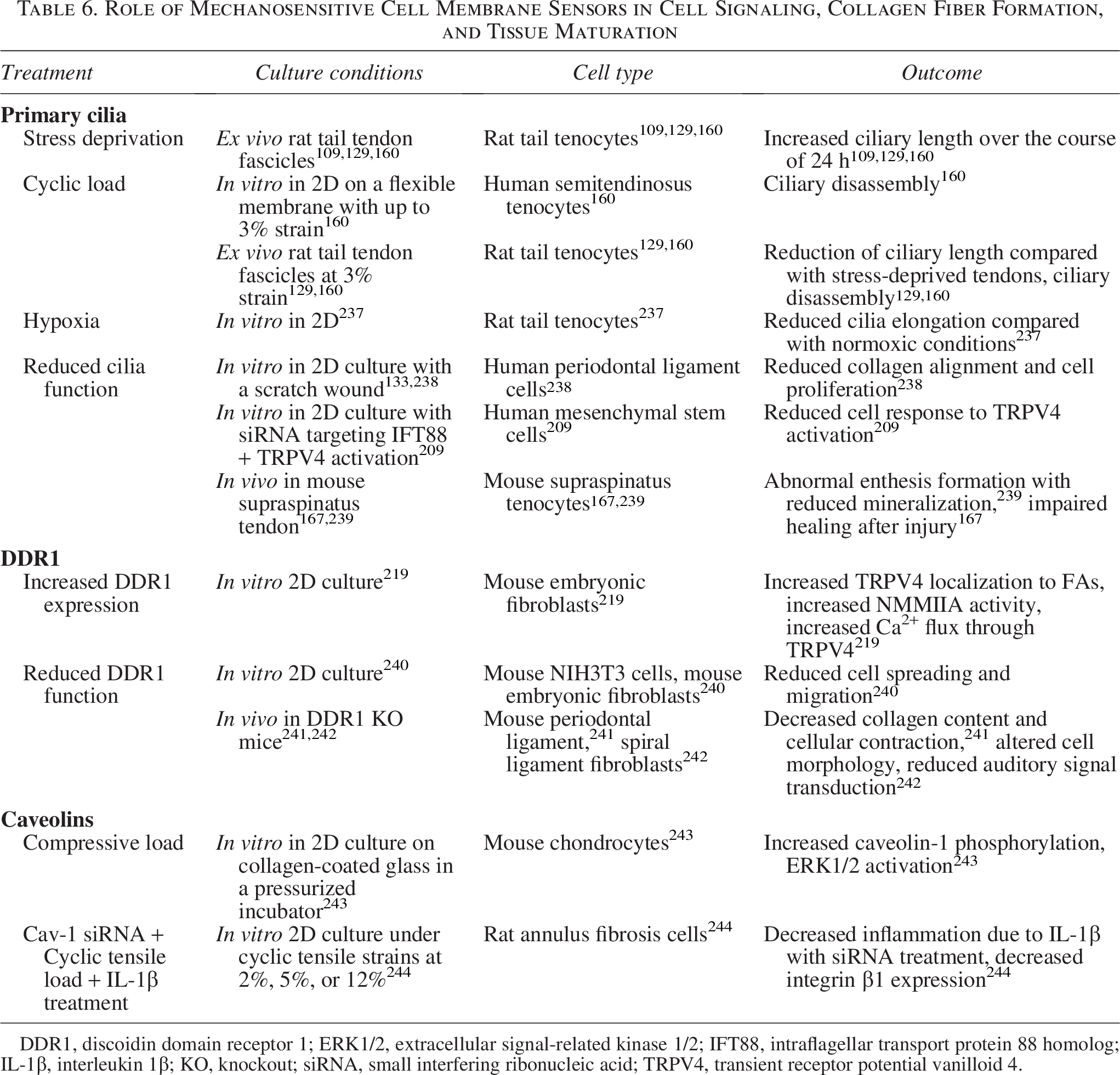
DDR1, discoidin domain receptor 1; ERK1/2, extracellular signal-related kinase 1/2; IFT88, intraflagellar transport protein 88 homolog; IL-1β, interleukin 1β; KO, knockout; siRNA, small interfering ribonucleic acid; TRPV4, transient receptor potential vanilloid 4.
Primary cilia also play a major role in the enthesis.167,238,239 Cilia are present in more cells in the tendon enthesis than the midsection and coincide with hedgehog signaling and enthesis mineralization.167,238,239 Furthermore, primary cilia are critical to enthesis mechano-responsiveness in developing and adult tissue, with loading driving assembly and disassembly of cilia and removal of cilia causing significant structural and compositional deficits in the enthesis. 239
Interestingly, primary cilia may also have crosstalk with TRPV4. It has been reported that TRPV4 localizes to the primary cilium of MSCs, and the effects of TRPV4 activation were reduced without functional primary cilia. 209 Collectively, while explored in the enthesis, the role of primary cilia in hierarchical fiber formation is less established. More work is necessary to understand how it directly, or in combination with other mechanosensors, influences hierarchical fiber formation.
Discoidin domain receptors
Discoidin domain receptors (DDRs) are mechanosensitive collagen-binding tyrosine kinases with a possible role in fibril alignment (Table 6). 253 DDR1 is a cell surface receptor that associates with NMMIIA upon binding to collagen, promoting cell migration and inhibiting spreading. 240 This suggests interplay between DDR binding and actin-cytoskeletal pathways, aligning with research that indicates both have a role in collagen fibril alignment.36,219,241 DDR1 also colocalizes with TRPV4 in mouse embryonic fibroblasts, and DDR1-collagen binding can induce TRPV4-mediated Ca2+ signaling. 219 This results in enhanced collagen binding and remodeling mediated by DDR1, which is lost if TRPV4 is inhibited. 219 The mechanosensitivity of DDR1 has been shown in the spiral ligament of the ear canal, where deletion of the DDR1 gene in mice altered outer hair cell shape. 242 The altered cell morphology was observed in the areas where DDR1 and NMMIIA colocalized in healthy mice, suggesting that altered DDR1 function results in alterations in actomyosin-mediated tensional homeostasis. 242
Caveolae
Caveolae are mechanosensitive indentations in the cell membrane of several cell types (Table 6).254,255 These indentations associate with actin and contain proteins such as cavin-1 and EH-containing domain 2 (EHD2), which are distributed into the cytoplasm and localize to the nucleus when caveolae are flattened due to stretching of the cell membrane.172,254 Caveolae have been implicated in altering actin-cytoskeletal dynamics, as well as activation of downstream signaling pathways such as YAP/TAZ and RhoA. 255 Caveolae are expressed in meniscal fibrochondrocytes and are again more numerous in the outer region of the meniscus where hierarchical fibers dominate, suggesting a role in responding to tension and regulation of collagen fibers. 256 While the role of caveolae in meniscal mechanobiology has not been thoroughly explored, similar tissues have shown that caveolins such as Cav1 respond to compressive and cyclic tensile load and may contribute to cellular tensional homeostasis.243,244 Despite the known association of caveolae and the actin-cytoskeleton, little is still known about their role in hierarchical fiber development or in tendons and ligaments, making them an enticing target for future research.
Conclusion
Our understanding of cell-driven hierarchical fiber formation is still rapidly evolving. There remain several research gaps and unanswered questions that are crucial to further our understanding, such as investigating how mechanical forces contribute to later stages of hierarchical fiber development, which mechanotransduction pathways contribute to tissue maturation via increased synthesis of different types of collagen, SLRPs, and LOX, and how various mechanosensation pathways interact with and alter one another. Most work has focused on the roles of actin-cytoskeletal mechanosensation in fiber development. While it is clear that actin-cytoskeletal-based mechanotransduction is a key mediator of fibril synthesis and tissue growth,119,126,157,257 it is becoming clearer that other mechanosensors are crucial to fiber maturation. For example, ion channels are now well established to play a role in modulating collagen synthesis and LOX crosslinking, both key to fiber maturation.196,199,210,213,214,219,258 Yet other mechanosensors such as primary cilia, DDRs, and caveolae are just beginning to be explored. All of these mechanosensation pathways, how they interact with each other, and how they influence downstream signaling pathways are exciting future research directions. Furthermore, as hierarchical fibers form, from embryonic development through adolescence, the loading environment, including the type, rate, and magnitude of load to which cells are exposed, will shift. We suspect this shifts the activity, role, or quantity of these mechanosensors and may suggest a need for adaptive stimulation or stage-targeted therapies to fully regenerate hierarchical fibers. In particular, we suspect that while actin-cytoskeletal sensing is critical to initial fibril and fiber formation, this sensing pathway will be less critical as fibers and fascicles mature and the cell experiences more secondary shear and compression. Here we may find a more dominant role of ion channels or other mechanotransduction pathways yet to be discovered.
Paramount to furthering our understanding of hierarchical fiber formation is understanding the mechanisms by which cells organize larger fibers and fascicles, which dominate the human musculoskeletal system. The majority of studies evaluating fiber formation are in 2D or rodent models. While mouse transgenic models are extremely powerful for exploring mechanisms, rodents largely lack the larger hierarchical organization of human musculoskeletal tissues,17,49,259 limiting our ability to understand how cells regulate fascicle formation. Thus, there is a need to use larger animal models for in vivo and ex vivo analysis of fiber regulation and to develop engineered systems that can reliably reproduce cellular environments within these tissues to better understand the role of mechanotransduction in fascicle formation. Little is understood about the mechanisms by which cells mediate later stages of hierarchical development, and cells largely do not regenerate these fibers after injury or in engineered replacements. Understanding the mechanisms by which hierarchical fiber formation is mediated and maintained is crucial to regenerating fibers after injury or in engineered replacements, and will help to inform the pathophysiological effects of unloading and overloading, and will help inform the benefits of controlled loading during rehabilitation.9,29,34
Authors’ Contributions
All authors have made substantial contributions to drafting and critical revision of the article and have read and approved the final submitted article.
Footnotes
Author Disclosure Statement
The authors declare they have no known competing financial interests or personal relationships that influenced the work reported in this article.
Funding Information
This work was supported in part by the National Institute of Arthritis and Musculoskeletal and Skin Disease of the NIH (