
Abstract
Volumetric muscle loss (VML) injuries result in an irrecoverable loss of muscle mass and function. VML injury causes loss of both contractile tissue and associated neuromuscular junctions (NMJs). Biosponge (BSG) scaffolds, composed of gelatin, collagen, and laminin-111, have improved recovery following VML. However, improvements in NMJ quantity were not observed. Glial cell line-derived neurotrophic factor (GDNF) is known to promote motor unit survival and stimulate neurite outgrowth. In this work, BSG scaffolds were electrostatically coupled with GDNF via gelatin nanoparticles (GNPs) to support myofiber regeneration and preserve NMJs post-VML in a rodent model. In vitro determination of release kinetics revealed an initial burst release of surface-bound GDNF with almost an equivalent amount of electrostatically bound GDNF retained within the BSG post 1 week of incubation at 37°C in phosphate-buffered saline. To create the VML injury in male Lewis rats (10–12 weeks old), ∼20% of the muscle mass was removed from the tibialis anterior (TA) muscle of both hindlimbs. Relative to BSG + GNP alone, treatment with BSG + GNP + GDNF showed a significant increase (∼25%) in peak isometric torque at 6 weeks postinjury. Qualitative and quantitative histological analysis of NMJs revealed an enhanced overlap between pre and postsynaptic structures in the BSG + GNP + GDNF group. Additionally, the incorporation of GDNF slowed BSG remodeling and degradation. Overall, these results suggest that BSG-mediated GDNF delivery is an effective strategy for mitigating NMJ loss and enhancing muscle recovery following VML.
Impact Statement
Volumetric muscle loss triggers progressive neuromuscular degeneration. Our findings demonstrate that treatment with biosponge scaffolds delivering glial cell-derived neurotrophic factor significantly enhances neuromuscular junction density and torque production. This approach represents a significant step forward in developing therapies that improve functional recovery and quality of life for extremity trauma patients.
Graphical abstract
Introduction
Skeletal muscle tissue shows minimal regeneration after volumetric muscle loss (VML), defined as the traumatic or surgical loss of skeletal muscle with resultant functional impairment. 1 Preclinical animal models of VML injury indicate that a relatively small VML injury (10–20% loss of muscle) can disproportionately reduce function (30–90% peak isometric torque deficit).2,3 Therefore, VML injury results in a loss of muscle function that exceeds the physical volume of tissue removed. This disproportionate impairment in function is driven by the simultaneous destruction of both myofibers and their innervating neurons. VML directly reduces the number of myofibers and causes axotomy of nerves, resulting in reduced muscle function and neuromuscular junctions (NMJs).4–7 In a recent study, a ∼20% VML defect resulted in chronic axotomy of ∼69% of the motor neurons innervating that muscle. 8 The reinnervation of tissue-engineered skeletal muscle (or denervated muscle) is a slow and extensive process that may require > 3 months for maximal recovery,9,10 suggesting that functional recovery can remain incomplete for months after the initial physical trauma. Together, these observations highlight that the functional deficit post-VML reflects both the removed and the denervated contractile tissue.
Recent work has also shown a continuous increase in denervation of VML-injured muscles. 11 Within 3 days of injury, denervation increased by 10%, but further escalated to 22% and 32% on days 21 and 48 postinjury, respectively. These changes were accompanied by a progressive increase in irregular morphological characteristics for NMJs (e.g., polyinnervation, axon sprouting, and endplate fragmentation). A similar progression of worsening pathology has been reported in both aged 12 and dystrophic 13 skeletal muscles.
In previous studies, biosponge (BSG) scaffolds composed of gelatin, collagen, and laminin (LM)-111, have improved muscle mass, function, cross-sectional area, and myofiber number following VML.14–17 However, improvements in NMJ quantity were not observed. Glial cell line-derived neurotrophic factor (GDNF) is a growth factor that enhances motor unit survival and neurite outgrowth.18–23 This study aimed to evaluate how incorporating GDNF into the structure of a BSG scaffold would improve the quantity and morphology of NMJs, as well as muscle function post-VML. We postulated that electrostatic conjugation of GDNF to a biomaterial carrier would protect and stabilize the growth factor while enabling sustained release through controlled dissociation. GDNF plays a crucial role in maintaining NMJ health through multiple mechanisms, 19 including regulating motor neuron survival, protecting motor neurons from chronic degeneration, and rescuing them from axotomy-induced cell death. While GDNF expression is induced and upregulated in denervation models up to 48 h postinjury, this endogenous response is not sustained enough for long-term or complete recovery. 24
We hypothesized that BSG scaffolds encapsulating GDNF-decorated gelatin nanoparticles (GNPs) 25 would provide a stable and targeted delivery system for GDNF, thereby preserving and increasing the quantity of NMJs following VML. Specifically, the electrostatic coating of GDNF on the GNPs and their subsequent encapsulation within the BSGs will improve the bioavailability of the growth factor by increasing the amount that can be carried and released at the target site and by ensuring the stability and protection of the electrostatically bound growth factor. We further propose that exogenous GDNF can play crucial roles during different phases of injury. In the acute phase, it can augment existing levels and counteract degradation caused by inflammation.26,27 In the chronic phase, it can provide sustained support for reinnervation and recovery, particularly when endogenous GDNF levels decline. 24
Materials and Methods
Detailed methods are provided as Supplementary Data.
Biosponge preparation
Sterile deionized (DI) water was heated to 50°C to make a 3 wt% Type A porcine skin gelatin (G2500-100G, Sigma Aldrich) solution. After complete dissolution, 20 mM of 1-ethyl-3-(3-dimethyl aminopropyl) carbodiimide (EDC; 22980, ThermoFisher) and 8 mM of n-hydroxysuccinimide (NHS; 130672-100 G, Sigma Aldrich) were added to the gelatin solution. Rat tail collagen I (9.50 mg/mL; 354249, BD Corning) was diluted in 1X phosphate-buffered saline (PBS; 10010-023, Gibco) to a concentration of 3 mg/mL. In a 6-well plate, 3.5 mL of the gelatin solution was combined with 1.5 mL of the collagen solution and 42 μL of LM-111 solution (6 mg/mL; 3446-005-01, R&D Systems). The final concentrations of the components were 21 mg/mL of gelatin, 0.9 mg/mL of collagen, 50 μg/mL of LM-111, 14.6 mM of EDC, and 5.6 mM of NHS. The solution was polymerized in the refrigerator (4°C) for 1 hour and was frozen overnight at −8°C. The well plate was then moved to −80°C for at least 72 h, then lyophilized for 12–14 h. After lyophilization, the BSGs were frozen at −20°C until needed.
Gelatin nanoparticle synthesis
To synthesize negatively charged GNPs, a previously described desolvation method was followed. 25 Type B bovine skin gelatin solution (G9382-100G, Sigma) at pH 10.0 was added dropwise using a syringe with a 21 G needle to a solution containing 42% acetone and 30% poloxamer-188 (P556, Sigma; 6230 A, Mirus) under mechanical agitation. Diisopropylcarbodimiide (DIC; A19292.30, ThermoFisher Scientific) was added at a final concentration of 9.2 mg/mL to the gelatin solution. This solution was vortexed and then allowed to stir at room temperature for 24 h to complete crosslinking. The alkaline Type B GNPs were then isolated by centrifugation at 5000 rpm for 10 min. This resulted in a phase separation, allowing GNPs to be collected from the top layer. The collected GNPs were then frozen, lyophilized, and stored at −20°C until needed. Please refer to the supplemental methods section for DLS and Fourier Transform Infrared Spectroscopy (FTIR) characterization.
GDNF encapsulation within biosponges
Alkaline type B GNPs were suspended at a concentration of 5.0 mg/mL in nuclease-free water. This solution was combined with murine GDNF (30 kDa; 450-40-50UG, PeproTech) to achieve final concentrations of 1.0 mg/mL GNP and 1 µg/mL GDNF, respectively. This GNP-:GDNF solution was allowed to incubate at room temperature for 10 min for electrostatic interactions to occur. BSG scaffolds were disinfected with 70% ethanol for 10 mins and then biopsied to a diameter of 6 mm to fit into a 24-well plate. The BSGs were then sterilized in the EcoVs 59S UVC LED sterilizing chamber for two cycles of 3 min and then rinsed in 1× PBS twice for 5 mins each. The BSG scaffolds were allowed to dry for 48 h in a sealed desiccator at 4°C. The BSG scaffolds (6 mm disk) were then rehydrated individually in a GNP−: GDNF solution (100 µL) in the 24-well plate for 2 h at 4°C. The release of GDNF was measured using the Mouse GDNF Enzyme Linked Immunosorbent Assay (ELISA) Kit (EEL117, Invitrogen) and Western blotting.
Release of GNP−:GDNF conjugates and their temporal effects on NSC-34 neurite growth
Previously developed BSGs were disinfected by rinsing in 70% ethanol and then biopsied to a diameter of 6 mm to fit into a 24-well Transwell® plate (Costar, REF 3413) with a 0.4 µm pore polycarbonate membrane insert. The BSGs were then sterilized in the EcoVs 59S UVC LED sterilizing chamber for two cycles of 3 min and then rinsed in 1x PBS twice for 5 mins each. They were then stored and allowed to dry for 96 h at 4°C. Mouse Motor Neuron-Like Hybrid Cell Line (Neural Stem Cell [NSC-34]; RRID:CVCL_D356; CLU140, Cedarlane Cellutions Biosystems Inc.) were seeded in the bottom chamber of a 24-well Transwell® plate at a density of 104 cells per well (n = 4/group) in complete proliferation media (DMEM-F12 + 10% Fetal Bovine Serum (FBS) + 1% P/S). The cells were imaged after 24 h and the media was replaced with complete differentiation media containing DMEM-F12, 1% FBS, 1% P/S, 1% Minimum Essential Medium (MEM)-Non-essential amino acid, and 1 µM retinoic acid.
After 24 h of culture, dry BSGs were separated into four groups: blank, GNP−-only, GDNF-only, and GNP−:GDNF and rehydrated as follows for 1 hour at 4°C. The blank group BSGs were each rehydrated with 100 µL of sterile nuclease-free water. The GNP−—only BSGs were rehydrated with 100 µL of 5 mg/mL GNP− suspension. GDNF-only BSGs were rehydrated with 100 µL of 1 µg/mL GDNF suspension in sterile nuclease-free water. The GNP−:GDNF BSGs were rehydrated with 100 µL of a solution that included 5 mg/mL GNP with 1 µg/mL GDNF, which had been incubated at room temperature for 10 min to facilitate electrostatic conjugation. BSGs were placed in transwell inserts and placed on top of the wells containing NSC-34 cells. On days 3, 5, and 7, transwell inserts were removed from the wells and stored under aseptic conditions in the biosafety cabinet, and the cells were reimaged. Following imaging, differentiation media was replaced, and the transwell inserts were returned to their original wells. An 2,3-bis-(2-methoxy-4-nitro-5-sulfophenyl)-2H-tetrazolium-5-carboxanilide (Tetrazolium salt) (XTT) assay was performed after imaging on day 7. Images showed NSC-34 neurite extensions, and their length was manually quantified using ImageJ (or Fiji; National Institutes of Health).
Animal experiments
All animal work was conducted in compliance with the Animal Welfare Act, the implemented Animal Welfare Regulations, and in accordance with the principles of the Guide for the Care and Use of Laboratory. All animal procedures were approved by Saint Louis University’s Institutional Animal Care and Use Committee (Animal Protocol # 2645).
Male Lewis rats (10–12 weeks old; Charles River Laboratories) were housed in a vivarium accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International, which provided water and food ad libitum. A total of 12 rats were randomly assigned to experimental groups (n = 4 per group) and an age-matched cage control group (n = 4). A full-thickness ∼20% VML defect was created in the TA muscle with a 6 mm biopsy punch. The injury was treated with 6 mm disks of either blank GNP BSG or with GNP + GDNF-loaded BSG (n = 4/group). The surgery was performed bilaterally, with both legs receiving the same treatment (i.e., n = 8 muscles/group). The metal spatula was removed after the BSG (6-mm diameter) was placed in the defect. The fascia was pulled over the BSG and sutured together with 6-0 Prolene sutures (Ethicon). The skin incision was closed using simple interrupted Prolene (5-0; Ethicon) sutures. Investigators were blinded to group allocation during data collection, and animals were identified by a numerical code.
After 6 weeks of recovery, all animals underwent peak isometric torque measurement followed by euthanasia. In vivo function testing of the anterior crural muscles was performed 6 weeks postinjury, as described previously.4,15,17,28–30 Animals were euthanized via thoracotomy and exsanguination. The TA muscle was then harvested, weighed, and processed for histological analysis.
Histological analysis
Frozen Optimal Cutting Temperature (Compound) (OCT) molds embedded with TA muscles were cryosectioned using a Leica CM1850 cryostat (RRID: SCR_025772) to obtain longitudinal cross-sections (60 µm). Sections were mounted on SuperfrostTM Plus Gold Slides (22–035813, Fisher Brand). Longitudinal sections (60 µm) taken at a depth of 720–960 µm from the muscle’s surface were stained and analyzed. Antibodies for alpha bungarotoxin (α-BTX), synaptophysin (Syn), and neurofilament were utilized to label the following structures, i.e., postsynaptic endplate, presynaptic endplate, and axons, respectively. 4′,6-diamidino-2-phenylindole (DAPI) staining was used to label nuclei.
NMJ analysis
A total of 1536 NMJs were initially imaged across all experimental groups. Of these, 1055 NMJs met the criteria for en face orientation and were utilized for analysis. Data are presented as the average per individual muscle sample in each experimental group. We analyzed a substantial number of en face NMJs per muscle (Control: 62 ± 30; VML + GNP: 29 ± 14; VML + GNP + GDNF: 41 ± 13). The higher variability in the control group may be attributed to the larger anatomical volume and more expansive motor endplate zone available for sampling compared to the injured groups. Imaging was performed on the Andor Dragonfly 200 spinning disk confocal (Oxford Instruments), mounted on a Leica DMi8 base, equipped with a HC Fluotar 25x/0.95 W Visible Infrared (VISIR) water-immersion lens.
Qualitative analysis of NMJs in confocal images was performed manually. NMJs were classified by the extent of overlap between the presynaptic nerve terminal stained with Syn + NF and the postsynaptic acetylcholine receptors stained with α-BTX. 31 Complete overlap between pre and postsynaptic terminals was defined as >75% colocalization, partial overlap as 25–75% colocalization, and no overlap as <25% colocalization between α-BTX+ and Syn + NF+ structures. Quantitative analysis was performed by two blinded investigators using a modified version of the NMJ-Morph methodology 32 to quantify the following: presynaptic area, postsynaptic area, colocalization area, and number of fragments. An innervated endplate was defined by the spatial colocalization of the presynaptic and postsynaptic structures. A denervated endplate was defined as the postsynaptic Acetylcholine Receptor ACh: Acetylcholine (AchR) clusters lacking associated presynaptic labeling.
Atomic force microscopy analysis
A high-performance MFP-3D-Bio atomic force microscope (AFM; Oxford Instruments) was used to analyze the mechanical properties of muscle tissues as described previously. 33
Statistical analysis
Data are presented as a mean ± standard error of the mean. Unpaired t-tests were used for two-group comparisons of GDNF retained vs. released, biopsy mass, and BSG area. One-way analysis of variance (ANOVA) was used to investigate differences in zeta potential, TA muscle mass, peak isometric torque, and pre- and postsynaptic area. For multifactor datasets, two-way ANOVA was used to determine main effects and interactions for neurite length and size distribution, Young’s modulus using atomic force microscopy (AFM), endplate fragmentation, and synaptic structural analysis (area, overlap, and colocalization). The normality of data sets was assessed using the Shapiro-Wilk test. For datasets that deviated significantly from a normal distribution, specifically fragmentation and colocalization, the nonparametric Kruskal-Wallis test was employed. GraphPad Prism 10 for Windows was used to perform statistical analyses and graph data. A Fisher’s Least Significant Difference (LSD) posthoc comparison was utilized to identify the source of significance with p < 0.05 unless otherwise specified in figure captions.
Results
GDNF release kinetics
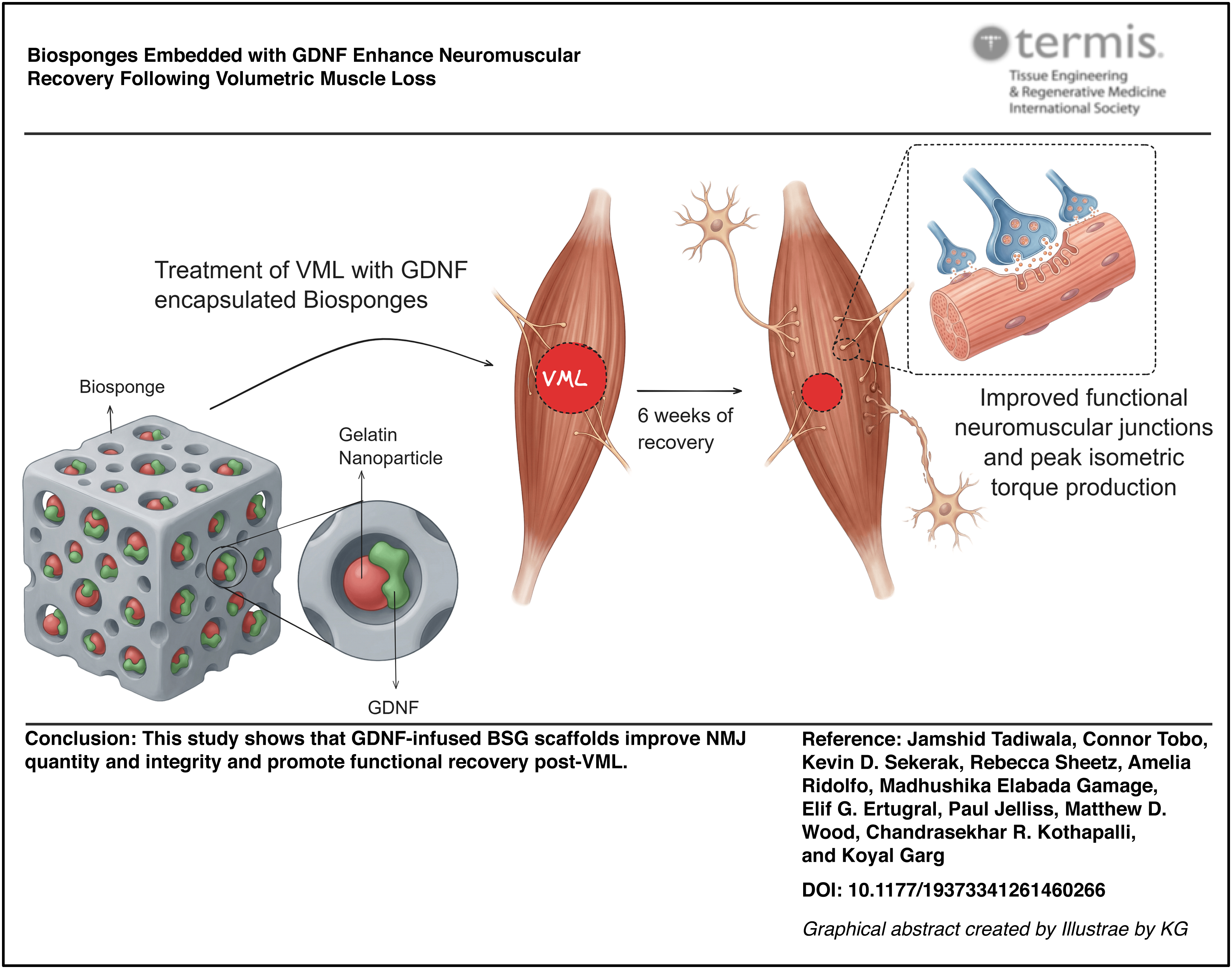
Zeta potential measurements confirmed that alkaline Type B GNPs were negatively charged (−12 mV) and Type A BSGs were positively charged (11.8 mV). GDNF-coated GNPs showed a reduced charge (−8.6 mV) likely due to the coating of GDNF on the GNP surface (Fig. 1A). However, all three groups were statistically different from each other (one-way ANOVA, p < 0.0001). The GDNF release study demonstrated a gradual, consistent decline in concentration over 6 days in vitro (Fig. 1B). A third-order (cubic) polynomial fit was used for the release data (R2 = 0.3334). Western blot results indicated that GDNF remained within the BSG structure even after 6 days of release (Fig. 1C). GDNF release from the BSG scaffold was modeled using the Korsmeyer-Peppas power law model (Mt/M∞ = Kt^n), yielding a release rate constant K of 0.77 and a diffusional exponent n of 0.79 (R2 = 0.95). The n value falls within the anomalous transport range (0.45 < n < 0.89), indicating that GDNF release is governed by a combination of diffusion and scaffold erosion rather than simple Fickian diffusion. Cumulative GDNF release reached approximately 3% of total loaded content over 6 days, consistent with the sustained retention of GDNF within the BSG scaffold and suggesting prolonged local bioavailability at the injury site (Supplementary Fig. S4).
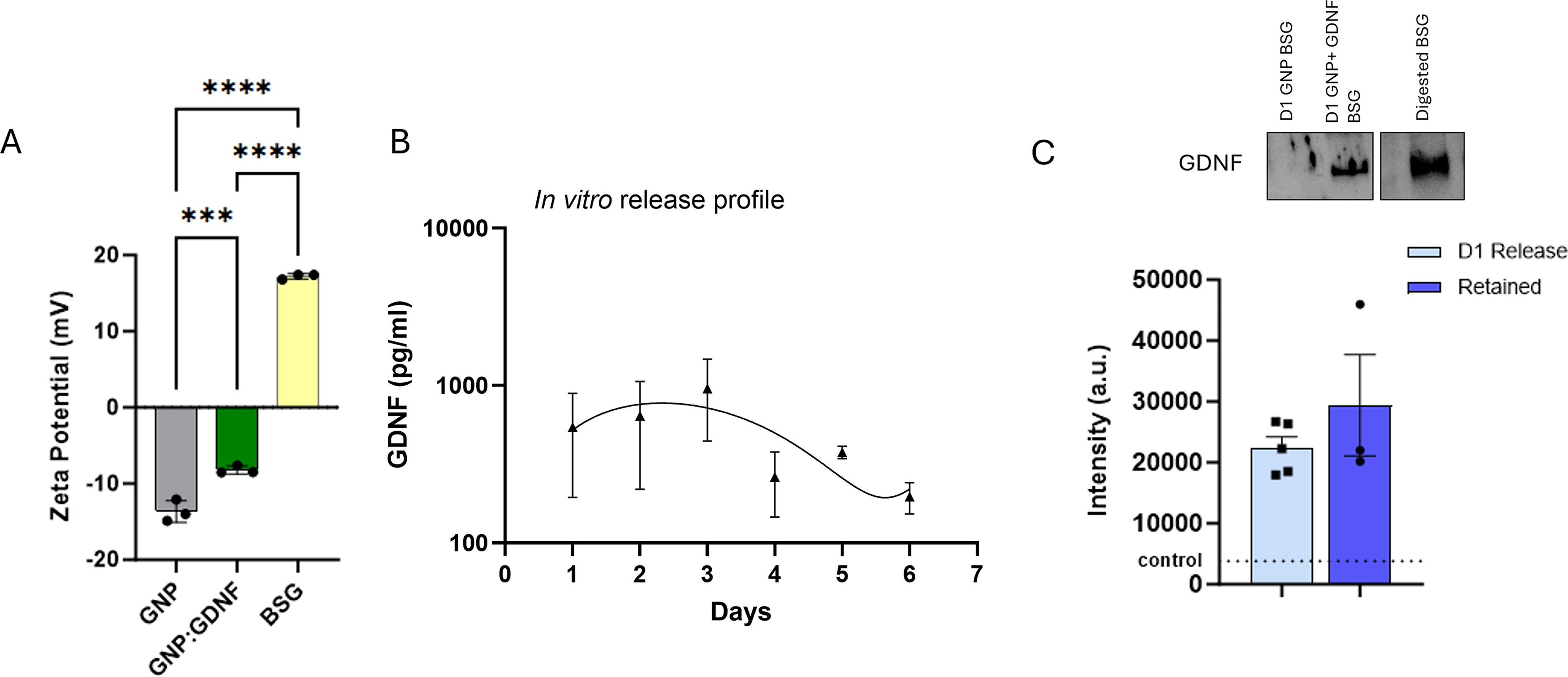
FTIR spectra of both acidic (Fig. 2A) and alkaline GNPs (Fig. 2C) demonstrate increases in transmittance upon conjugation with GDNF (Fig. 2B–D). These changes occur primarily around the Amide I (1600–1700 cm−1) and Amide II (1510–1580 cm−1) bands. Sharper amide bands typically suggest a reduction in conformational flexibility.34–38 These changes indicate that the interaction of GDNF with GNP has induced a more ordered or specific structural arrangement within the conjugate. The structure of GDNF is known to feature both positive and negative residues on its surface, 39 so differences in spectral intensity between GDNF conjugated to positively and negatively charged GNPs are expected.
The FTIR spectra for
To characterize the endogenous neurotrophic environment following VML injury, we quantified transcript abundance for a panel of neurogenic factors using bulk RNA sequencing (Supplementary Fig. S5) on days 7 and 14 postinjury. Differential expression analysis confirmed the presence of several neurotrophic transcripts within the injury site. Members of the GDNF family exhibited near-baseline expression levels throughout the recovery period. Specifically, GDNF was undetectable in control samples and remained at trace levels postinjury (<0.5 Transcripts Per Million [TPM]). Artemin, Neurturin, and Persephin similarly demonstrated low endogenous titers, consistently remaining below 2.0 TPM. Similarly, brain-derived neurotrophic factor and ciliary neurotrophic factor remained below 0.2 TPM and 2.1 TPM, respectively. While nerve growth factor and neurotrophin-3 were detectable, their expression never exceeded 4.5 TPM. In contrast, the regenerative response was dominated by robust expression of insulin-like growth factor (IGF1), IGF2, and Vascular Endothelial Growth Factor A (VEGFA), with higher transcript levels. These results highlight that the endogenous expression of neurogenic signaling molecules is orders of magnitude lower than that of myogenic and angiogenic growth factors in the VML injury microenvironment. Collectively, these results highlight a deficit in neurotrophic signaling required for reinnervation and NMJ stability.
Bioactivity of GDNF on NSC-34 cells
NSC-34 cellular morphology on days 1, 3, 5, and 7 of culture is shown in Figure 3A. Neurite extension quantification (Fig. 3B) on days 3 and 5 showed significant main effects for both treatment and time, with no significant interaction between the two (2-way ANOVA, interaction p = 0.7525, time effect, p = 0.0347, treatment effect, p < 0.0001). Posthoc analysis across all time-points demonstrated that groups containing GDNF exhibited significantly longer neurite extensions compared to groups without GDNF. Quantifiable neurites were not observed on day 1, and cellular confluency obscured measurements on day 7. Therefore, neurite extensions were not quantified on days 1 and 7. A size distribution analysis of neurite length was also performed. A highly significant interaction between treatment and neurite size was observed (Fig. 4A–B
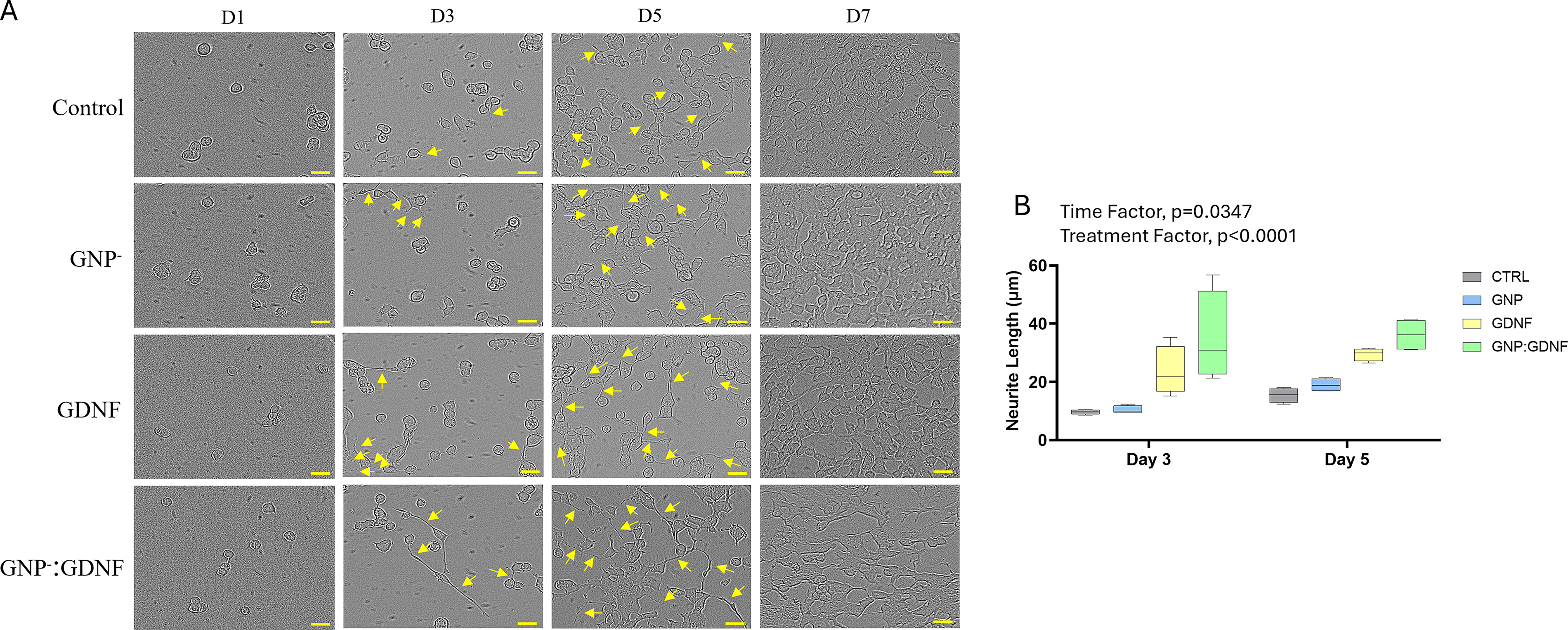
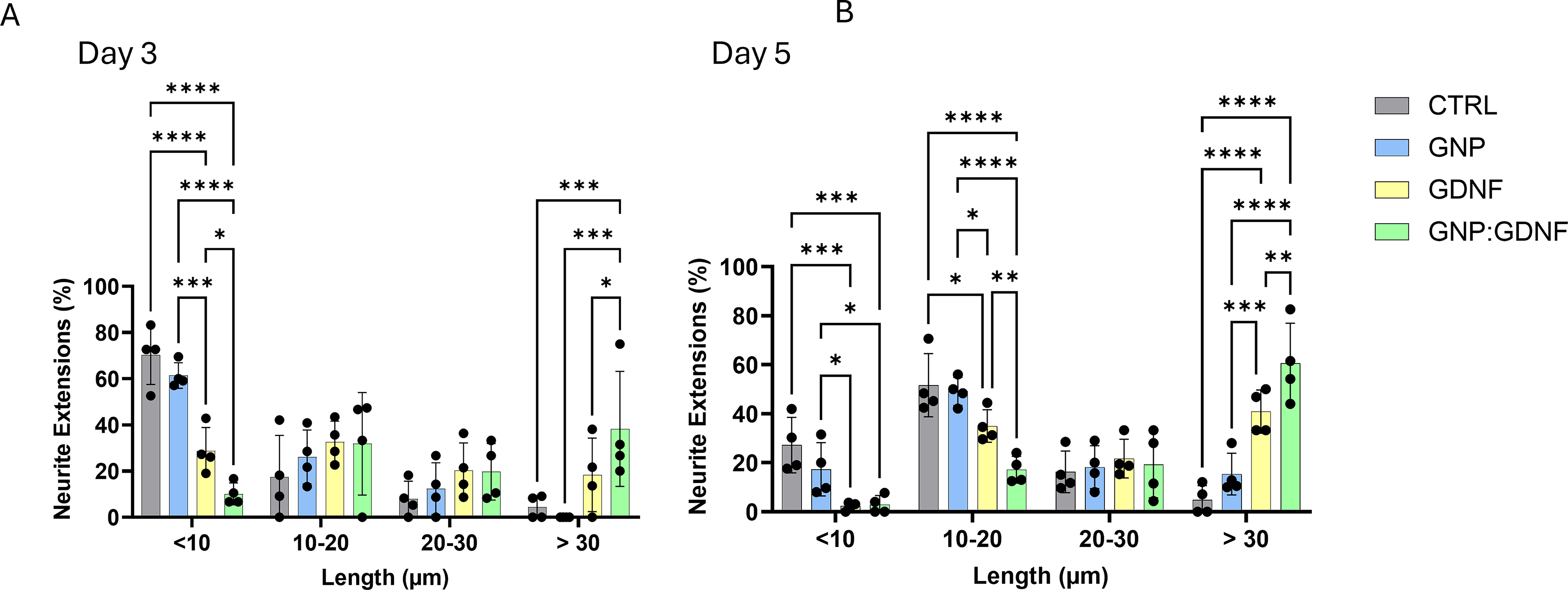
The size distribution analysis of neurite extensions from NSC-34 cells showed that GNP + GDNF containing biosponges resulted in significantly longer neurite extensions in the >30 μm size range on days 3
Muscle mass and function
The biopsy mass removed to create the VML injury was consistent between the experimental groups (Fig. 5A) (t-test, p = 0.9074). The TA muscle mass was measured at 6 weeks postinjury. Postnormalization of muscle mass to body weight of the animal, the results showed ∼33.5% (GNP) and ∼26.5% (GNP + GDNF) deficits relative to the uninjured muscle. However, no differences between treatment groups were observed (Fig. 5B) (one-way ANOVA, p < 0.0001; Uninjured vs. GNP, p < 0.0001; Uninjured vs. GNP + GDNF, p = 0.0002).
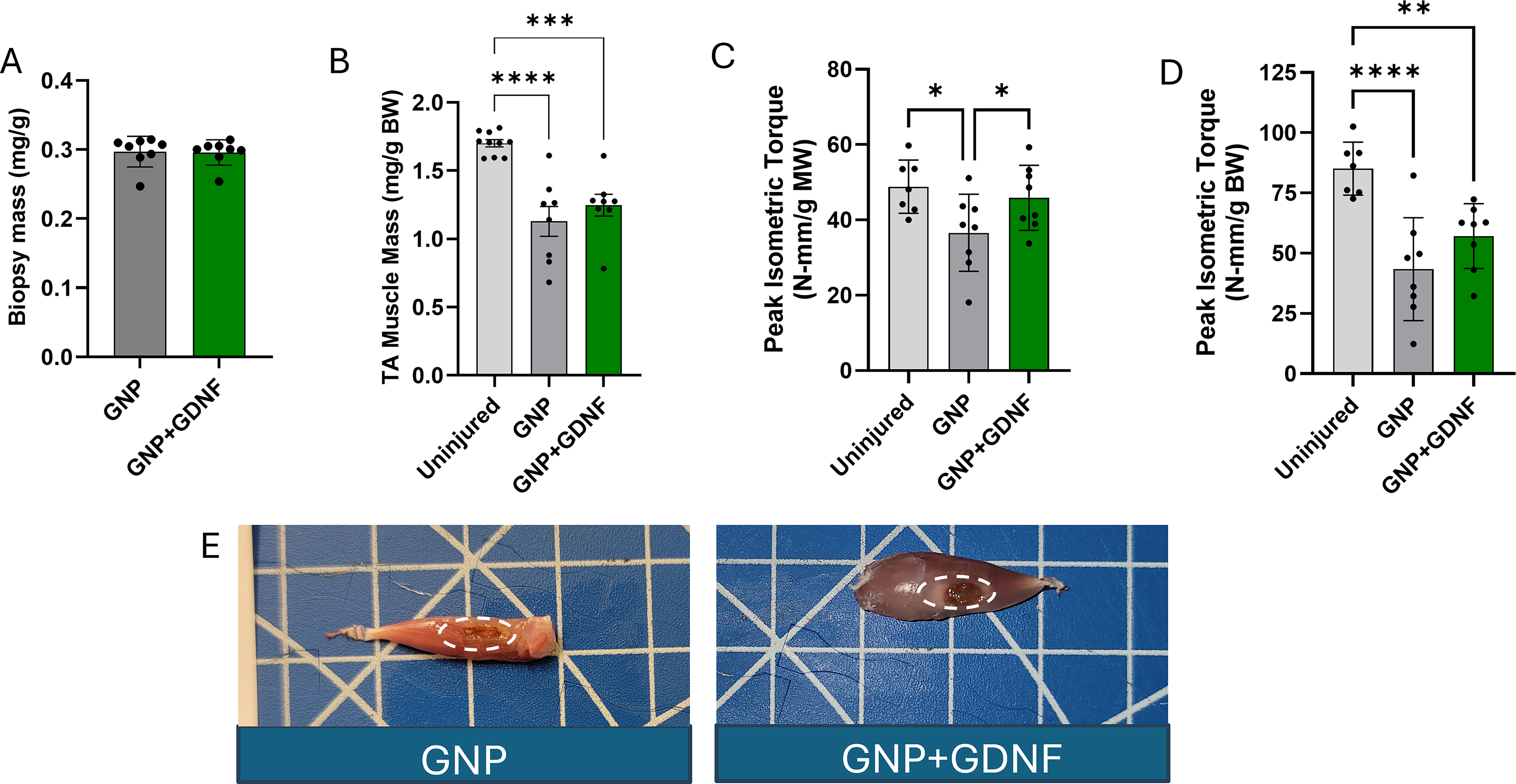
Peak isometric torque was measured at 6 weeks postinjury and showed a significant increase (∼25%) with GNP + GDNF treatment relative to the GNP-only group when normalized to the TA muscle mass (Fig. 5C) (one-way ANOVA, p = 0.0337). However, no differences were found when comparing the peak isometric torque normalized to the body weight of the animal between GNP only and GNP + GDNF (Fig. 5D; One-way ANOVA, p = 0.0003). The BSG could be visually identified in the excised TA muscles in both groups, as shown in the photographs (Fig. 5E). The raw data for TA muscle mass and peak isometric torque are shown in Supplementary Fig. S3 A–B.
Biosponge remodeling and AFM analysis
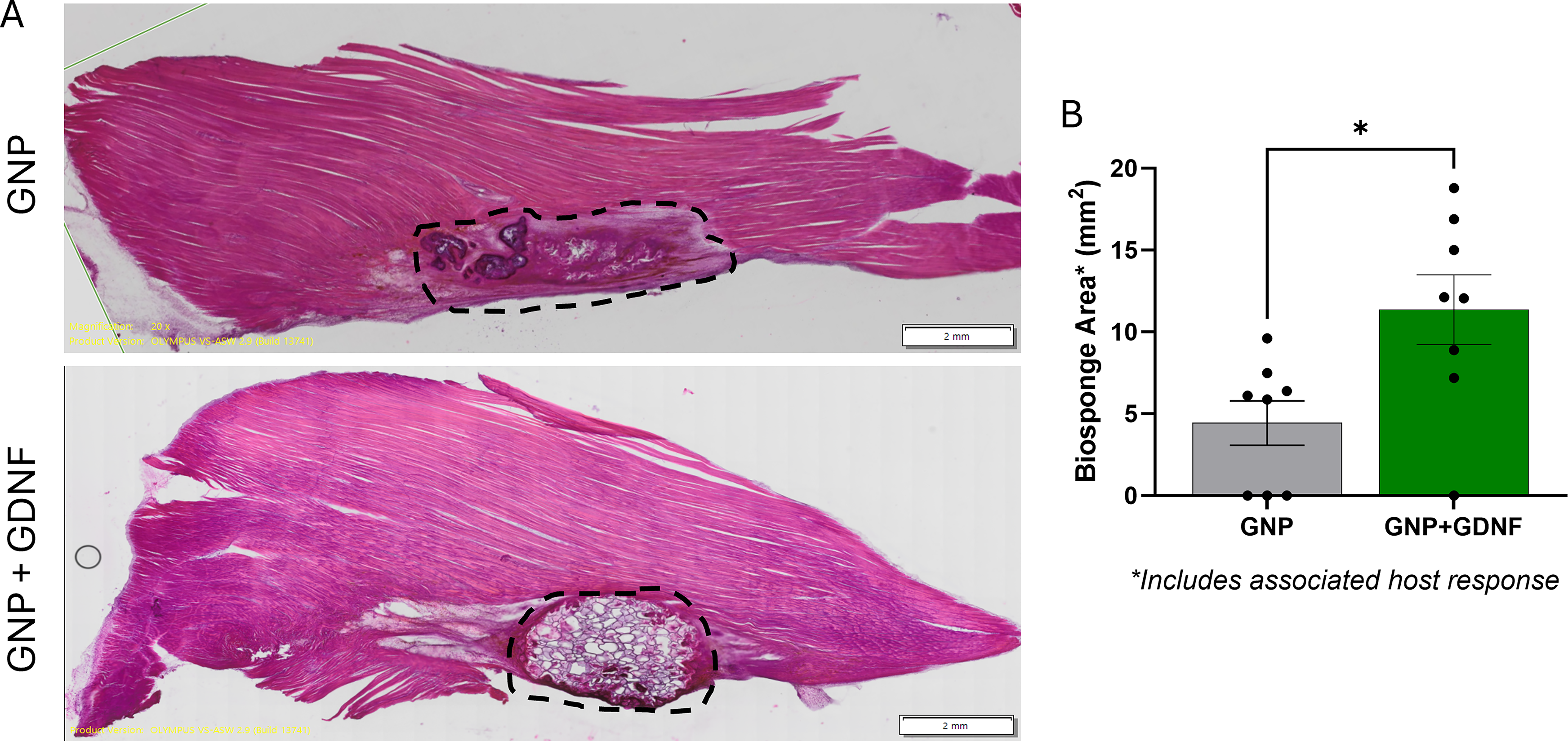
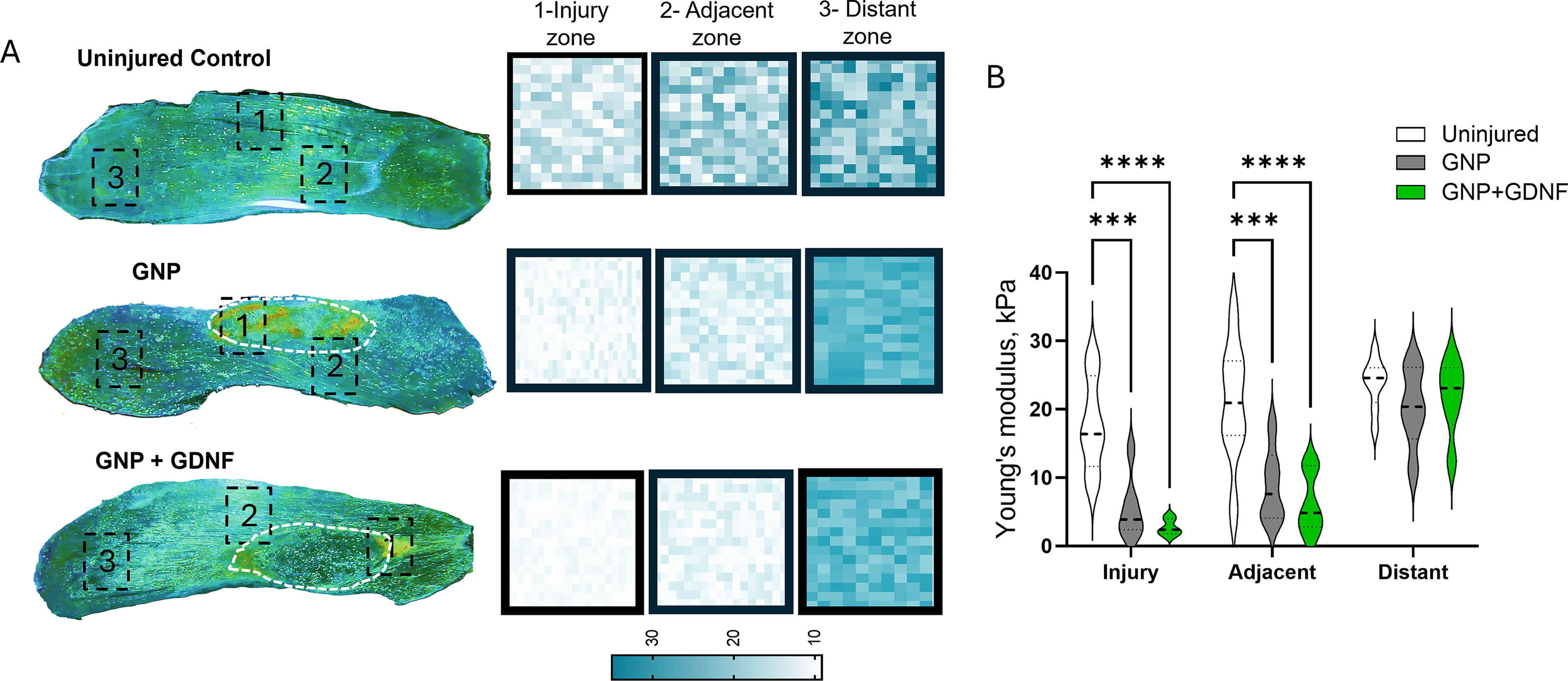
The BSG area quantified from H&E-stained muscle histological sections is shown in Figure 6A. The GNP + GDNF group showed a higher area of BSG relative to that of the GNP-only group, suggesting slower remodeling and degradation in the presence of GDNF (Fig. 6B; t-test, p = 0.0157. AFM analysis revealed significant main effects for both region and treatment, with a significant interaction between the two factors (Fig. 7A–B; Two-way ANOVA, Interaction, p = 0.0193, regional factor, p < 0.0001, treatment factor, p < 0.0001). Posthoc analysis revealed that the Young’s modulus was significantly reduced in both GNP (p = 0.0002) and GNP + GDNF (p < 0.0001) muscles compared to uninjured controls within the injury and adjacent zones. This mechanical deficit was localized to the VML defect, as no significant differences in Young’s modulus were observed between any groups in the distant zone (p > 0.05). Neither GNP nor GNP + GDNF treatment significantly restored tissue stiffness in the affected zones to the uninjured levels.
Neuromuscular junction analysis
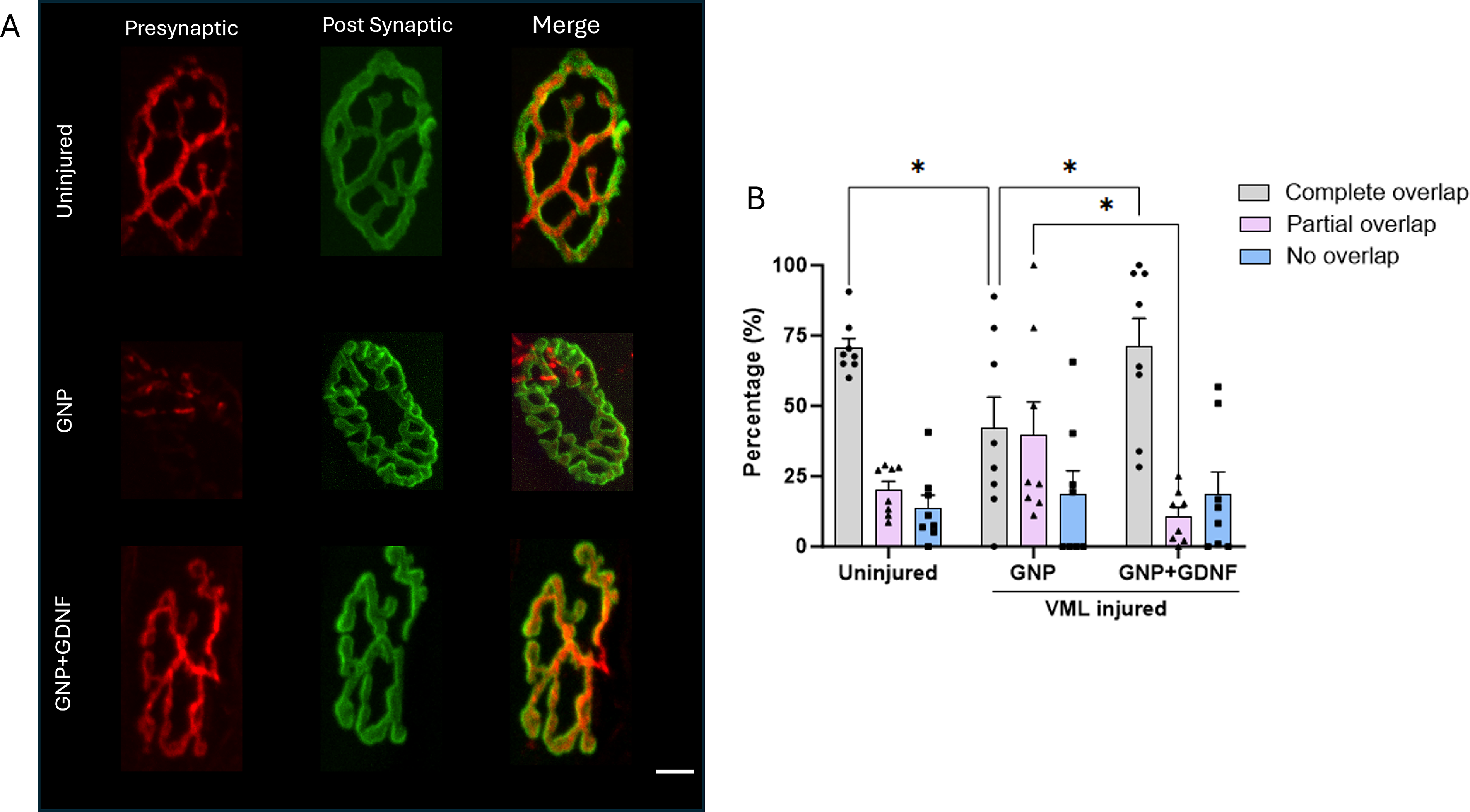
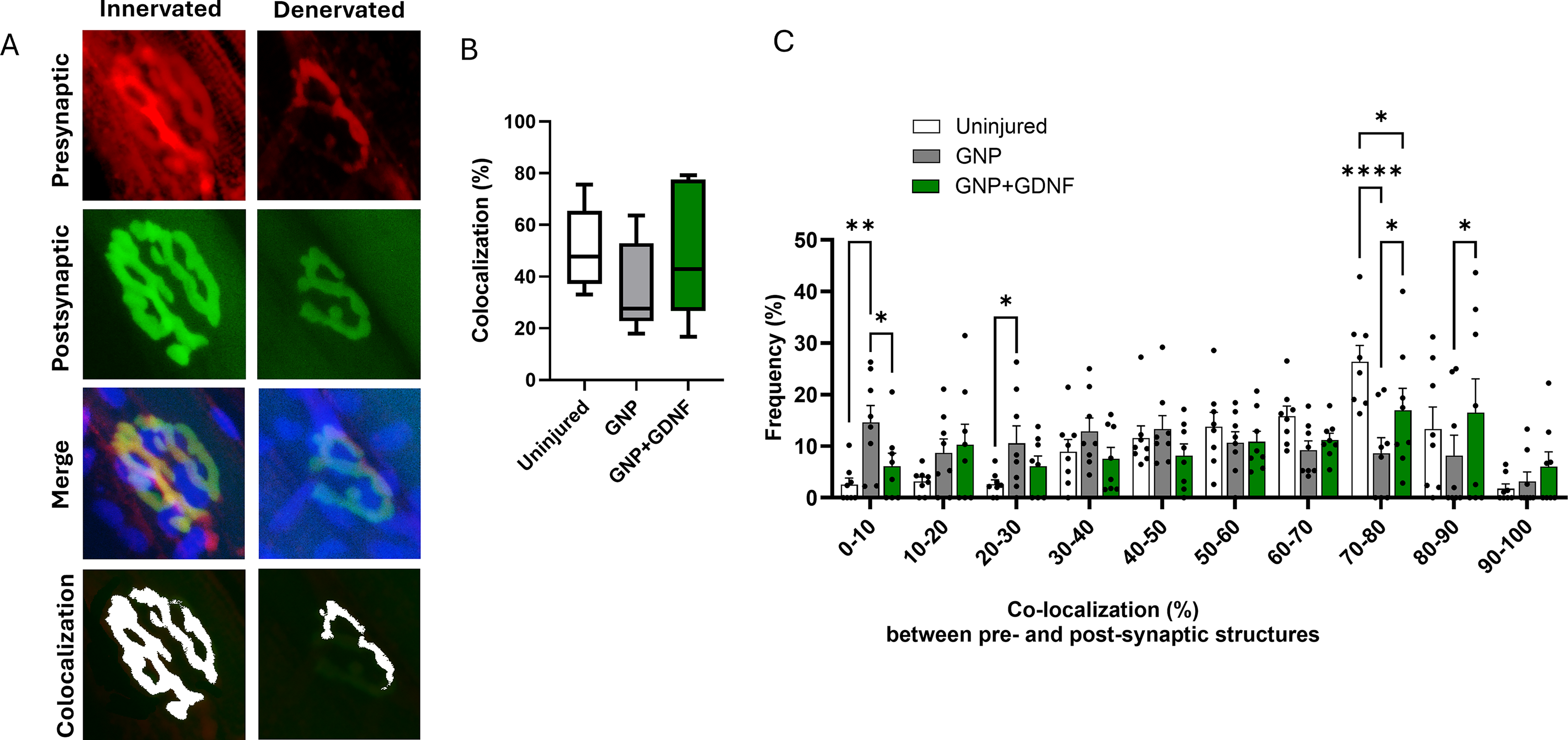
Representative confocal images for each experimental group showing the pre and postsynaptic structures labeled with NF + SYN and α-BTX are presented in Figure 8A. Semiquantitative assessment of NMJs in the confocal images showed an average of ∼71% complete, 20% partial, and 14% no overlap in the uninjured TA muscles. The GNP + GDNF group exhibited 71% complete, 11% partial, and 18% no overlap between pre- and postsynaptic structures, compared to 42% complete, 40% partial, and 18% no overlap in the GNP group. A significant interaction was observed between treatment and synaptic overlap (Fig. 8B; Two-way ANOVA Interaction p = 0.0054, Treatment Factor p = 0.9643, Overlap Factor p < 0.0001). Posthoc analysis revealed a significantly higher percentage of complete overlap in the GNP + GDNF group compared to the GNP only group (p = 0.0113). Conversely, the GNP group exhibited a higher proportion of partial overlap compared to the GNP + GDNF group (p = 0.0112). These results suggest a higher quantity of innervated endplates in the GNP + GDNF group. Representative images in Figure 9A depict both individual and merged, presynaptic (red) and postsynaptic (green) terminals, including the overlapping region (colocalization; white) for a fully innervated and a partially denervated endplate. A quantitative assessment of presynaptic and postsynaptic terminal colocalization revealed no significant difference in the overall mean percentage between the treatment groups. The mean percentage of colocalization was found to be ∼51% in the uninjured group, ∼34% in the GNP-treated group, and ∼49% in the GNP + GDNF-treated group (Fig. 9B
The postsynaptic area was measured to be 287 µm2 in the uninjured group, 388 µm2 in the GNP group, and 320 µm2 in the GNP + GDNF group, showing a 35% and 11.5% increase relative to the uninjured group, respectively (Fig. 10A). Significant differences were observed between groups (one-way ANOVA, p = 0.0131). The GNP group showed a larger endplate area relative to the uninjured (p = 0.0041) and GNP + GDNF (p = 0.0440). Distribution analysis also revealed a shift towards larger postsynaptic terminals, primarily in the GNP alone group (Fig. 10B). A significant interaction between treatment and endplate size was observed (2-way ANOVA; Interaction, p = 0.0074; Size factor, p < 0.0001, Treatment factor, p > 0.9999). Posthoc analysis revealed that within the largest size category (>700 µm2), the GNP-only group exhibited a significantly higher percentage of endplates compared to the GNP + GDNF group (p = 0.0392).
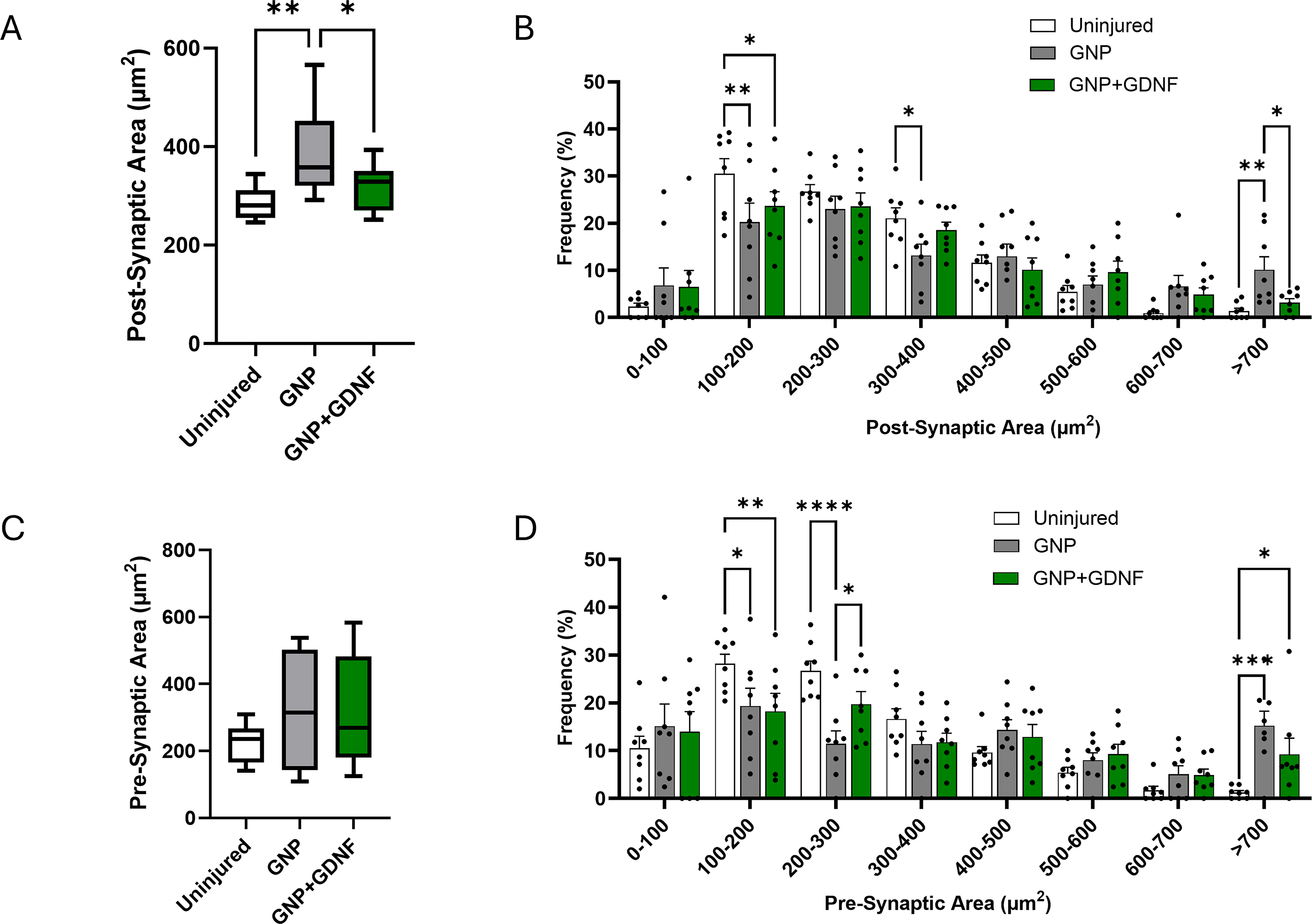
The presynaptic area was measured to be 225 µm2 in the uninjured group, 320 µm2 in the GNP group, and 314 µm2 in the GNP + GDNF group, revealing a 42% and 40% increase relative to the uninjured group, respectively (Fig. 10C). However, statistical analysis showed no significant differences between the groups (one-way ANOVA, p = 0.3427). However, distribution analysis revealed a shift toward larger presynaptic terminals in both the GNP and GNP + GDNF groups (Fig. 10D). A significant interaction between treatment and presynaptic terminal size was observed (2-way ANOVA; Interaction, p < 0.0001; Size factor, p < 0.0001, Treatment factor, p > 0.9999). Posthoc analysis revealed that within the largest size category (>700 µm2), the uninjured control group exhibited a significantly lower percentage of presynaptic terminals than the GNP (p = 0.0002) and GNP + GDNF (p = 0.0285) groups.
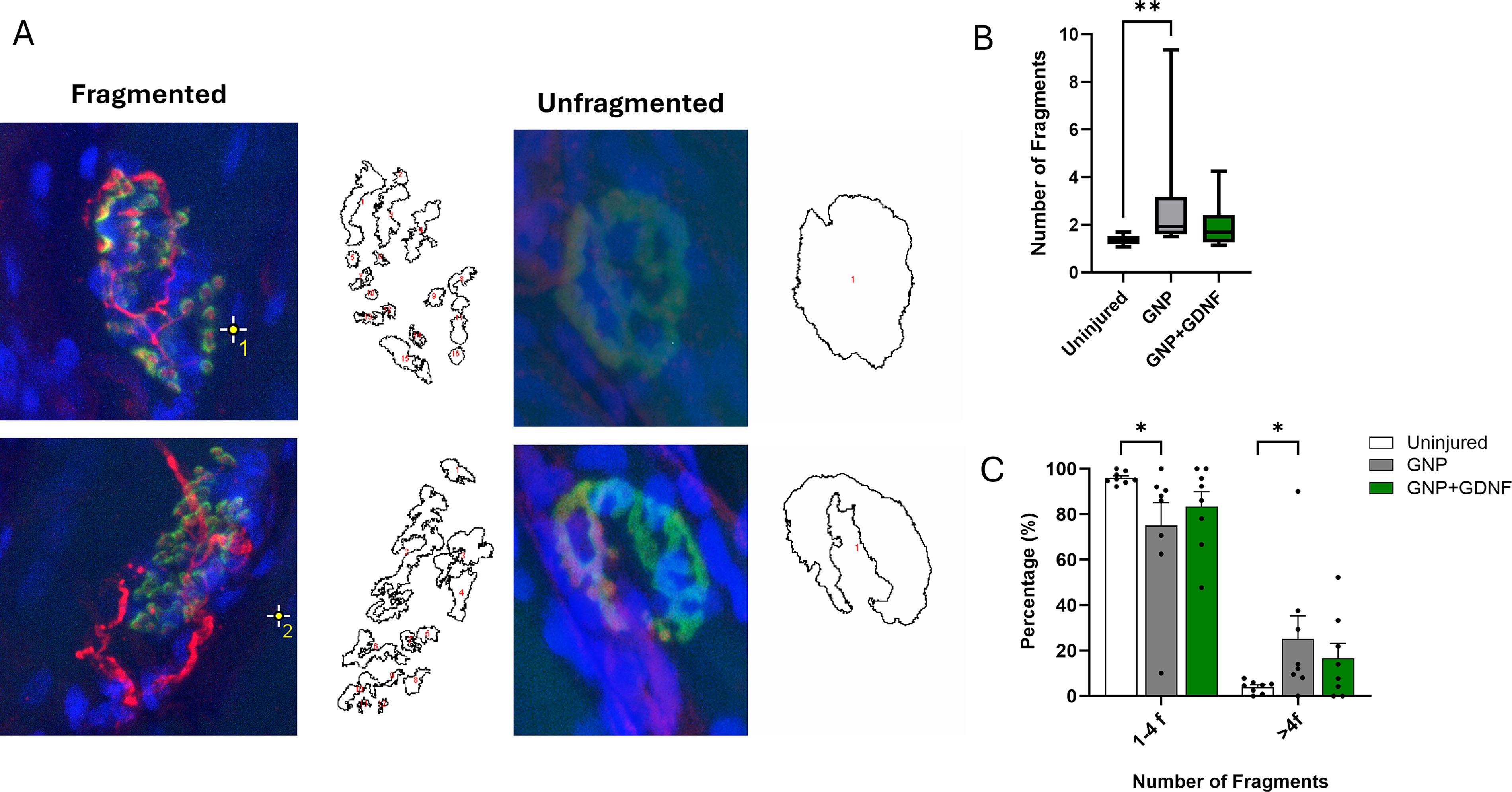
Representative images of fragmentation in motor endplates are depicted in Figure 11A. Treatment of VML with BSG containing GNP alone led to an increase in postsynaptic fragmentation (Fig. 11B; Kruskal–Wallis test, p = 0.0229). However, when the BSG was supplemented with GNP + GDNF, the extent of fragmentation was comparable to that of uninjured control muscles. Analysis of endplate morphology revealed a significant interaction between treatment and fragmentation state (Fig. 11C; Two-way ANOVA, Interaction p = 0.0159, Fragmentation factor p < 0.0001, Treatment Factor p > 0.9999). Posthoc analysis revealed an increase in the percentage of endplates with greater than four fragments in the GNP alone group relative to the uninjured muscles (p = 0.0395), suggesting structural degradation of the motor endplate. Notably, the GNP + GDNF group did not differ significantly from uninjured controls (p > 0.05).
Discussion
The most salient finding of this study is that treatment with a BSG scaffold encapsulating GDNF improved the number of innervated endplates as well as peak isometric torque production in VML-injured muscles compared to scaffold treatment alone. These results suggest that GDNF loading augments the therapeutic efficacy of BSG scaffolds for VML treatment.
In a previous study, Sorensen et al. 11 observed a leftward shift in the presynaptic terminal size distribution, suggesting a decrease in size, coupled with a rightward shift in the postsynaptic terminal size distribution, indicating an increase in size following VML injury. An increase in postsynaptic endplate area following VML injury has also been reported by Hoffman et al. 40 A shrinking of the presynaptic terminal would suggest that Acetylcholine (ACh) molecules are not being consistently released. The enlargement of postsynaptic terminals suggests a compensatory adaptation in which the AChRs attempt to maximize their chances of capturing ACh. Additionally, enhanced fragmentation of the postsynaptic terminal was also observed by Sorensen et al. 11 A similar pathology has also been observed in dystrophic mice.13,41
We observed, however, that both the presynaptic and postsynaptic structures of the NMJ were enlarged in the BSG + GNP-treated VML-injured muscles. This adaptation likely enables the NMJ to store increased amounts of ACh in synaptic vesicles and upregulate the expression of AChR receptors on the myofibers. In support, previous studies have shown that the amount of ACh released at a given terminal region is highly correlated with the local abundance of synaptic vesicles at the same site and that a relatively constant fraction (∼30%) of the total vesicle pool is released per stimulus. 42 These changes may help delay the onset of neuromuscular transmission failure, thereby mitigating the associated muscular fatigue. These findings suggest a protective effect and a positive overall impact of the BSG + GNP on both muscle and nerve repair. However, the extent of postsynaptic terminal fragmentation was increased despite BSG treatment, suggesting a lack of AChR organization, which will inevitably limit neuromuscular transmission and prevent functional recovery.
The BSG + GNP + GDNF group showed a different pattern, with the presynaptic terminals exhibiting a rightward shift, indicating an increase in size, while the postsynaptic terminal size remained comparable to controls. Another important finding was the reduction in postsynaptic fragmentation with this treatment. The BSG + GNP + GDNF treatment strategy also resulted in increased colocalization between pre- and postsynaptic structures, as well as increased peak isometric torque production. Taken together, these findings suggest that the postsynaptic structures or AChRs were better organized and positioned to form a stable, accurate contact with the presynaptic terminal. A larger presynaptic terminal would suggest ample ACh release for effective signal transmission and torque production. Therefore, the improvement in NMJ quantity and function, driven by GDNF, provides the underlying mechanism for increased torque production in this group.
We speculate that an ECM-based BSG scaffold provides structural support and guidance cues for regenerating axons and myofibers while simultaneously modulating the inflammatory response, creating a conducive microenvironment for both muscle and nerve survival and growth.14–17,28,29 However, this treatment alone was insufficient to enhance the percentage of innervated endplates. In other studies, improvements in NMJ quantity were not observed when BSG scaffolds were combined with eccentric exercise 28 or placental stem cells. 4 Nakayama et al. implanted aligned nanofibrillar collagen scaffolds loaded with IGF-1 into a mouse VML model. However, the NMJ quantity was increased only when scaffolds were combined with exercise. 43 In another study by Quarta et al., decellularized bioconstructs containing muscle stem cells increased NMJ number only when combined with exercise. 7 These observations suggest that incorporating potent neurotrophic or synaptogenic factors within scaffolds is necessary to enhance reinnervation and functional recovery.
Other groups have utilized biomaterials loaded with various biomolecules to both study and improve neuromuscular regeneration following VML. For instance, Mihaly et al., 44 and Scott et al., 45 used agrin-tethered fibrin scaffolds to enhance AChR clustering on skeletal muscle. Ziemkiewicz et al., showed increased NMJ quantity and functional recovery in fibrin hydrogels supplemented with LM-111. 6 The success of these strategies further suggests that localized delivery of neurogenic signaling molecules is necessary for myofiber reinnervation. A key limitation of all the studies mentioned above is reliance on transverse 10–15 µm sections for quantitative analysis of NMJs. This method often yields inaccurate estimates of NMJ quantity and morphology. However, the preferred method of using longitudinal sections (>50 µm thick) for accurate NMJ analysis 46 is precluded in these experimental designs because multiple other measurements and analyses must be performed on the same tissue samples to determine the extent of tissue repair and regeneration.
The current study has several limitations. Because the harvested muscle tissue was allocated exclusively for longitudinal histological analysis, gene or protein expression analysis of GDNF levels could not be performed after treatment. Although in vivo release kinetics were not measured, the enhanced recovery outcomes observed at 6 weeks postinjury in the BSG + GNP + GDNF group suggest prolonged GDNF availability. This sustained release is mechanistically supported by the slow degradation of the BSG, which was histologically identifiable even at 6 weeks. In vitro data corroborates this, showing an initial burst release of loosely bound GDNF within the first 3 days, followed by a slower, sustained release of electrostatically bound, deeply embedded GDNF within the BSG pores. Additionally, a single time-point (i.e., 6 weeks postinjury) was used for analysis. This prevents the assessment of the kinetic healing process. Furthermore, the need for thick longitudinal tissue sections for high-fidelity NMJ analysis, while beneficial for quantifying total NMJ count, precludes detailed analysis of muscle regeneration, specifically the quantification of myofiber CSA and fiber type distribution. Future studies should examine NMJ numbers at multiple time points and include transverse tissue sections to provide a more detailed analysis, thereby establishing a comprehensive, kinetic understanding of both muscle and nerve regeneration. Another limitation of this work is the reliance on a male-only cohort. Although this design controlled for physiological fluctuations, it highlights the need for subsequent investigations. These data provide the preliminary groundwork for exploring sex-specific recovery mechanisms in female populations. Another limitation of this study is the absence of an untreated VML control group.
Despite these limitations, the study demonstrates the efficacy of GDNF-augmented BSG scaffolds as a promising approach to preserve NMJ numbers and enhance functional recovery following VML. These findings provide a foundational framework for the development of future scaffold-based therapies to restore neuromuscular function following traumatic injuries.
Authors’ Contributions
K.G., P.J., M.D.W., and C.R.K. designed the study. J.T., C.T., A.R., K.D.S., R.S., M.E.G., E.G.E., and K.G. performed experiments, collected data, performed statistical analysis, and prepared figures. J.T. and K.G. drafted the article. All authors read and approved the final version for publication.
Footnotes
Acknowledgments
The authors also thank Caroline Murphy at the SLU histological core for assistance and training with confocal microscopy. We would like to express our gratitude to the Genomics Core at Cleveland Clinic for their invaluable assistance with library preparation and sequencing.
Author Disclosure Statement
GenAssist, Inc. is developing products related to the research described in this article. KG has an equity interest in GenAssist, Inc. and serves on the company’s scientific advisory board. The terms of this arrangement have been reviewed and approved by Saint Louis University, in accordance with its conflict-of-interest policies. The authors declare no other competing financial or nonfinancial interests.
Funding Information
This work was supported by a pilot seed grant from the Institute of Translational Neuroscience (ITN) at Saint Louis University, awarded to KG. This work was also supported by NIGMS
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.