
Abstract
Severe burn injuries pose significant clinical challenges, with high risks of infection, excessive inflammation, and impaired wound healing. Mesenchymal stem cells (MSCs) have shown regenerative and antimicrobial potential; however, their therapeutic efficacy is constrained by poor survival and engraftment. Here, we demonstrate that priming MSCs with insulin-secreting cells (ISCs) and encapsulating them in hydrogels (HEMI) enhances their regenerative function, leading to accelerated healing of full-thickness burns in a porcine model. By 5 weeks, 80% of HEMI-treated wounds achieved complete closure, compared with 50% of MSC-only wounds and 0% of standard-of-care control wounds. All groups exhibited partial wound closure over time, but no control wounds reached complete closure within the study period. Histological analysis revealed complete epidermal and dermal regeneration with minimal fibrosis in HEMI-treated wounds. Single-cell RNA sequencing and differentially abundant sequencing analysis identified distinct MSC subpopulations whose relative abundance and transcriptional profiles differed between insulin-primed and control MSCs, with insulin priming promoting pathways involved in extracellular matrix stabilization, immune modulation, and oxidative stress resistance. Our findings suggest that insulin priming enhances MSC-mediated tissue repair via paracrine mechanisms, providing a clinically translatable strategy for improving burn treatment and regenerative medicine applications.
Impact Statement
The clinical translation of mesenchymal stem cell therapies has been hindered by poor cell retention after transplantation and inconsistent regenerative efficacy. This study demonstrates that insulin priming and hydrogel encapsulation act synergistically to enhance MSC function and retention, thereby improving wound healing in a clinically relevant porcine burn model. Because these barriers are common across many forms of impaired skin repair, including burns, diabetic ulcers, and age-related chronic wounds, this platform has broad potential to advance regenerative medicine strategies for cutaneous tissue repair.
Introduction
Burn wounds remain a major global health burden, with high rates of morbidity, mortality, and healthcare costs. Despite advancements in wound management and skin grafting techniques, severe burns remain difficult to treat due to delayed healing, chronic inflammation, and increased risk of infection. More than 75% of burn-related deaths result from infection and systemic inflammation, rather than direct burn trauma.1,2 Antibiotic resistance further complicates treatment, underscoring the urgent need for alternative, infection-resistant therapies that accelerate wound healing.3,4
Mesenchymal stem cells (MSCs) offer a promising therapeutic strategy due to their ability to modulate immune responses, secrete growth factors, and enhance tissue regeneration.5–7 However, poor survival, low engraftment efficiency, and apoptosis in hypoxic wound environments limit their effectiveness.8,9 Our previous work has demonstrated that priming MSCs with insulin activates the PI3K-Akt signaling pathway, enhancing cell survival, immune modulation, and extracellular matrix (ECM) remodeling. 10 However, it remains unclear how insulin priming influences MSC functional states at a single-cell level and whether this approach can be translated into a clinically relevant wound-healing therapy.
Here, we investigate the therapeutic efficacy of insulin-primed MSCs encapsulated within hydrogels (hydrogel encapsulated MSCs and insulin-secreting cells [ISCs] = HEMI) using a clinically relevant porcine model of full-thickness burns. The hydrogel formulation used in this study is based on a previously established platform in which we demonstrated accelerated healing in a diabetic mouse model of delayed wound repair. 10 In both that study and the present work, polyethylene glycol diacrylate (PEGDA) encapsulation permits the use of the same encapsulated cell products across different animal model systems by providing immunoisolation,11–13 thereby eliminating the need to rederive species-matched cells for each model. Prior work from our group has characterized PEGDA-hydrogel swelling behavior, mesh size, protein diffusion, encapsulated cell viability, and sustained release of bioactive factors.14–16 PEGDA encapsulation has also been shown across multiple studies to support xenogeneic cell function while permitting diffusion of glucose, insulin, and other soluble therapeutic factors.10,11,14,17–19
We employ single-cell RNA sequencing (scRNA-seq) and differential abundance sequencing (DA-seq) analysis to characterize MSC transcriptional states associated with insulin priming. These analyses identify changes in MSC subpopulation abundance and gene expression programs linked to extracellular matrix remodeling, immune modulation, and oxidative stress resistance. Together, these findings support a paracrine mechanism by which insulin priming may enhance MSC-driven tissue repair.
Materials and Methods
Cell culture
Rat insulinoma beta (RIN-m) cells (ISCs) were purchased from the American Type Culture Collection (ATCC) and cultured in RPMI-1640 medium (ATCC) with 10% fetal bovine serum (FBS) and 1% penicillin-streptomycin in a tissue culture incubator. Human bone marrow-derived mesenchymal stem cells (hMSCs, 8013 L) were provided by the Texas A&M Health Science Center College of Medicine Institute for Regenerative Medicine at Scott & White through a grant from ORIP of the NIH. Cells were maintained in alpha-minimal essential medium supplemented with 10% FBS, 1% penicillin-streptomycin, 4 mM
Hydrogel fabrication and encapsulation of cells
Hydrogel sheets encapsulating ISCs and/or hMSCs were fabricated by photopolymerizing a precursor solution composed of 10 kDa PEGDA (10% w/v), 1.5% triethanolamine/4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES)-buffered saline (pH 7.4), 37 mM 1-vinyl-2-pyrrolidinone, and 0.1 mM eosin Y. Cells (1 × 104 cells/µL) were mixed into the precursor solution and pipetted into glass molds to create 400 µm thick 4 x 4 cm2 hydrogel sheets containing either hMSCs alone or both cell types at a density of 0.5 × 106 cells per cm2. This PEGDA formulation is based on prior work demonstrating support for encapsulated cell viability, diffusion of soluble factors, and sustained secretion of bioactive molecules.10–14 Accordingly, the hydrogel was treated as a well-characterized delivery platform rather than as an experimental variable. The hydrogels were photopolymerized under white light for 2 min and then incubated in complete culture medium until use. For animal experiments, hydrogels were shipped overnight from Irvine, CA, to Boston, MA, and incubated in a tissue culture incubator upon arrival before application to wounds.
Study design and animal allocation
Wounds were assigned to treatment groups based on spatial separation on the dorsum of each animal to reduce proximity effects, local diffusion of soluble factors, and potential crosstalk between treatment zones. As a result, wounds were not randomized. Treatment groups were distributed across anatomical regions when possible, and the personnel responsible for measuring wound closure were blinded to the treatment condition to mitigate measurement bias.
Sample size justification
Sample sizes were determined based on prior mouse studies evaluating wound-healing outcomes using hydrogel-based MSC therapies. Power analysis indicated that a minimum of n = 3 per group would be sufficient to detect statistically significant differences with 95% power. For the porcine studies, sample size was determined taking into account statistical power, ethical considerations, and logistical constraints associated with large-animal models.
Inclusion and exclusion criteria
Inclusion criteria required that wounds be free of infection at the time of treatment. Any wounds that showed signs of infection during the study were excluded from analysis to prevent confounding effects on wound healing.
Humane endpoints and animal monitoring
Animals were monitored daily for signs of distress or discomfort by trained veterinary staff. Humane endpoints included lethargy, lack of appetite, or persistent behavioral changes. If animals exhibited any of these criteria, veterinary intervention was initiated. No animals met the criteria for early euthanasia during the study.
Housing and husbandry
All porcine studies were conducted at Massachusetts General Hospital (MGH), which maintains animal housing in accordance with United States Department of Agriculture (USDA) guidelines. Minipigs were provided with enriched environments including appropriate bedding, toys, and access to social interaction as part of their standard care to promote well-being and reduce stress.
Protocol registration
The experimental protocol was prepared collaboratively across multiple institutions and finalized in advance of study initiation to meet institutional and funding compliance requirements. However, this protocol was not preregistered in a public repository.
Full-thickness burn model in swine
All animal procedures were conducted in accordance with a protocol approved by the MGH Institutional Animal Care and Use Committee (IACUC). A total of six MGH miniature swine (20–24 kg) were initially enrolled in the study. Due to institutional pandemic-related restrictions, one animal was sacrificed prior to the completion of the study, reducing the total number of animals to five. Additionally, another pig developed a nosocomial infection leading to the removal of its hydrogels after 1 week. The animal was treated with antibiotics and monitored. The wounds were excluded from wound rate calculations, further reducing the total number of animals to four for the remainder of the study.
All surgical procedures were conducted under general anesthesia and in accordance with approved institutional animal care protocols. Prior to surgery, swine received an intramuscular injection of Telazol (2 mg/kg) for sedation, followed by endotracheal intubation. Anesthesia was maintained with 2–3% inhaled isoflurane in oxygen throughout the procedure. The surgical site on the dorsum was aseptically prepared using sequential applications of 2% chlorhexidine acetate, 70% isopropyl alcohol, and 10% povidone-iodine. At the study endpoint, animals were euthanized via intravenous administration of a pentobarbital/phenytoin solution (100 mg/kg), in accordance with the American Veterinary Medical Association guidelines and institutional IACUC protocols.
Full-thickness burn wounds (4 × 4 cm2) were induced using a brass block heated to 200°C and applied with constant pressure for 30 s (Fig. 1). A total of eight burn wounds were created per animal (15% body surface area), evenly distributed on both sides of the thoracic paravertebral region (n = 32 burns total). Multiple wounds were created on each animal and treated as independent experimental units, consistent with standard practice in large-animal wound healing studies. Prior work has evaluated the potential interdependence of wounds within the same animal and found no correlation between wound healing outcomes, supporting this approach. 20 After the procedure, animals were allowed to recover under general anesthesia, with postoperative thermoregulation managed via heated blankets. Transdermal fentanyl patches (25 or 100 µg) were applied for 3 days to provide analgesia.

Full-thickness porcine burn model.
Wound treatment and monitoring
At day 5 postburn, an escharotomy was performed to remove necrotic tissue, exposing the wound bed. The same thickness of skin was removed from every wound to ensure comparable wounds. Hemostasis was achieved using monopolar cauterization and pressure. Hydrogel sheets containing either MSCs alone, MSCs with ISCs (HEMI), or a standard of care (SOC, control) treatment (Silver Sulfadiazine and Xeroform) were applied. Hydrogels were sealed with Tegaderm® sheets and covered with sterile gauze and Vetrap® bandaging tape. For control wounds, dressings were changed three times per week, while hydrogel-treated wounds were undisturbed but checked regularly to ensure the integrity of the hydrogels and absence of infection.
Two animals were treated with all three groups (Fig. 1D). To ensure there was no crosstalk between HEMI and MSC treatments, two animals received only HEMI and SOC. In total, there were 12 control wounds, 14 HEMI wounds, and 6 MSC wounds. During every dressing change, individual pictures of the wounds were taken. Control wounds were imaged directly. Wound area was measured using NIH ImageJ software, and percent closure was calculated as:
% Closure = 100 × [1−(wound area/initial wound area)]. Complete closure was defined as 100% closure, with no visible open wound area. Hydrogels were removed on day 15, and wound closure was monitored over a 5-week period.
Histological analysis
At week 2, biopsies were taken from the center and edges of one wound per group for histological evaluation, with the remaining wounds biopsied at week 5. Samples were fixed in 10% formalin for 24 h, transferred to 70% ethanol, and then sectioned and stained with hematoxylin and eosin for morphological analysis. A trained pathologist, blinded to treatment groups, assessed gross appearance, granulation tissue formation, vascularity, and signs of inflammation. The pathologist also measured the percentage of intact epidermis over the length of the section. Epidermal thickness was measured every 200 µm using ImageJ and averaged to estimate epidermal and dermal thickness across each wound. Biopsied wounds were excluded from further wound closure analyses.
Barcoding and library preparation for single-cell RNAseq
The Parse Biosciences Evercode Whole Transcriptome 100k kit was used to prepare single-cell libraries for scRNA-seq. One control sample (1320 cells/µL) and one treated with insulin (1930 cells/µL) were loaded into 10 separate wells (5 wells per sample) in a 48-well plate provided by Parse Biosciences for initial barcoding. Following reverse transcription and barcoding, all samples were pooled and incubated in the ligation mix for a second round of barcoding. The pooled cells were then redistributed to a 48-well plate. This step was repeated once more for a total of 3 barcoding cycles. The cells were then pooled again and counted to calculate the final concentration. The total number of cells in the control and insulin-treated samples were 12,308 and 19,366, respectively. For each sublibrary, cells were lysed, barcoded cDNA was amplified, fragmented, and a unique sublibrary index was added prior to sequencing. The two sublibraries were sequenced on a single NextSeq2000 (Illumina) sequencing run for an average of 51,296 reads/cell (control) and 50,868 reads/cell (insulin-treated).
Single-cell RNA sequencing
After 3 days of incubation with insulin, plated cells were detached using trypsin/ethylenediaminetetraacetic acid (EDTA) (0.05%) and counted using the automated cell counter Countess (Thermo). Cells were centrifuged at 200 ×g for 10 min, the supernatant was discarded, and cells were resuspended in 750 μL of Cell Prefixation Buffer (Parse Biosciences). Samples were then fixed using the Parse Biosciences Cell Fixation (v1 or v2) kit and stored at −80°C until the start of barcoding and library prep with the Parse Biosciences Evercode Whole Transcriptome 100k (v1) kit. Fixed cells were processed using a Parse Biosciences Evercode Whole Transcriptome 100k kit (EC-W01030) for scRNA-seq. Preprocessing of scRNA-seq data was performed using the Seurat package in R. Low-quality cells were filtered based on mitochondrial gene percentage (>10%), low feature counts (<200 genes), and high feature counts (>5000 genes) indicative of potential doublets. Data normalization was carried out using Seurat’s SCTransform function, followed by principal component analysis (PCA) for dimensionality reduction. Uniform manifold approximation and projection (UMAP) was applied to visualize clustering.
Differential abundance analysis
Following scRNA-seq, DA-seq was performed as previously described. 21 The DA-seq pipeline was implemented as follows: (1) Cells were projected into PCA space, and local cell density differences were computed between insulin-primed and control MSCs; (2) differentially abundant (DA) regions were identified using multiscale clustering, ensuring subpopulations were detected across different levels of granularity; (3) cells within DA regions were further analyzed for gene expression changes, focusing on pathways relevant to tissue regeneration, immune modulation, and extracellular matrix remodeling; and (4) enrichment analysis was performed using Gene Ontology (GO) terms to characterize functional pathways upregulated in insulin-primed MSCs. The DA-seq method provided an unbiased identification of subpopulations enriched in regenerative phenotypes, complementing traditional differential expression analysis. This approach enabled the detection of subtle but functionally significant MSC subpopulations that were not apparent in conventional clustering methods.
Statistical analysis
Statistical analyses were performed using Prism GraphPad and Microsoft Excel. Wound closure data were compared across groups using one-way analysis of variance (ANOVA), with individual wounds treated as experimental units for wound-level analyses. Assumptions of normality and equal variances were considered reasonable based on inspection of data distribution and comparable standard deviation sizes across groups. A p value of < 0.05 was considered statistically significant. Single-cell RNA-seq data were analyzed using the Seurat package for R, with clustering and dimensionality reduction techniques employed to identify distinct cell populations and their corresponding gene expression profiles via PCA and UMAP. Differential gene expression analysis was used to identify insulin-primed genes in MSCs, with functional annotation carried out through GO enrichment analysis using Metascape. 22
The four animals completing the full study protocol yielded a total of 32 wounds distributed across treatment groups (HEMI: 14 wounds; MSC: 6 wounds; control: 12 wounds). A post hoc power analysis based on the proportion of wounds achieving complete closure by week 5 (HEMI: 80%, MSC: 50%, SOC: 0%) and an assumed pooled standard deviation of 10% indicated that a minimum of 2–4 wounds per group would be required to detect statistically significant differences with 80% power at α = 0.05. This analysis was used to contextualize the observed effect size and was not used as a prospective sample size justification. The sample sizes used exceeded these thresholds and were sufficient to detect the large effect sizes observed in wound closure and histological outcomes.
Results
To evaluate the therapeutic potential of insulin-primed MSCs encapsulated in hydrogels (HEMIs), we employed a full-thickness burn model in MGH miniature swine, comparing three treatment groups: hydrogel-encapsulated MSCs alone (MSC group), hydrogel-encapsulated MSCs and ISCs (HEMI group), and SOC. Hydrogels were removed after 2 weeks, and wound closure was monitored over 5 weeks. HEMI-treated wounds exhibited accelerated healing over the 5-week study period. All wounds demonstrated progressive partial closure over time, including standard-of-care controls. By week 5, 80% of HEMI-treated wounds achieved complete closure, compared with 50% of MSC-only wounds and 0% of control wounds. In short, the reported 0% value for controls refers to the proportion of wounds reaching complete closure, not to an absence of wound healing. The most rapid wound closure was observed in the first 2 weeks while the hydrogel remained in place, suggesting that local continuous release of MSC-derived factors contributed to early tissue repair (Fig. 2). Cohen’s d effect sizes were estimated from the proportion of wounds achieving complete closure at week 5 (HEMI: 80%, MSC: 50%, control: 0%) and an assumed pooled standard deviation of 10%. The resulting effect sizes were d = 3.0 (HEMI vs. MSC), d = 8.0 (HEMI vs. SOC), and d = 5.0 (MSC vs. SOC), indicating large to extremely large treatment effects.

Wound closure over time.
Histological analysis further confirmed that HEMI-treated wounds regenerated skin layers, including epidermis, dermis, and hypodermis, with well-organized granulation tissue and minimal fibrosis. In contrast, MSC-treated wounds showed incomplete reepithelialization, while SOC-treated wounds showed partial but incomplete reepithelialization, indicating a significantly impaired healing response (Fig. 3).

Quantification of intact epidermis at wound edges and centers after 5 weeks. HEMI treatments showed fully intact epidermis across biopsies, indicating enhanced reepithelialization compared to MSC and control controls. Significance was found at the centers of wound for both the intact epidermis percent F(2,21) = 7.1, p = 0.004, and the epidermis thickness F(2,21) = 6.52, p = 0.006. Graphs show mean ± standard error.
The epidermal layer in HEMI-treated wounds was significantly thicker than in other groups, suggesting that insulin priming enhances epithelial proliferation, barrier restoration, and dermal-epidermal junction formation. Additionally, HEMI-treated wounds exhibited highly vascularized granulation tissue, correlating with the upregulation of angiogenic and extracellular matrix (ECM)-modulating genes in insulin-primed MSCs. These findings suggest that HEMI treatment promotes robust tissue remodeling while preventing excessive fibrosis, leading to enhanced wound architecture and functional recovery.
To identify transcriptional changes associated with insulin priming, we performed scRNA-seq on insulin-primed and unprimed MSCs (Fig. 4). While unsupervised clustering revealed similar cellular populations, DA-seq analysis identified four distinct MSC subpopulations that differed significantly in their abundance between insulin-primed and unprimed conditions. These subpopulations were distinguished by gene expression signatures associated with proliferation, extracellular matrix remodeling, adhesion, and contractile function.
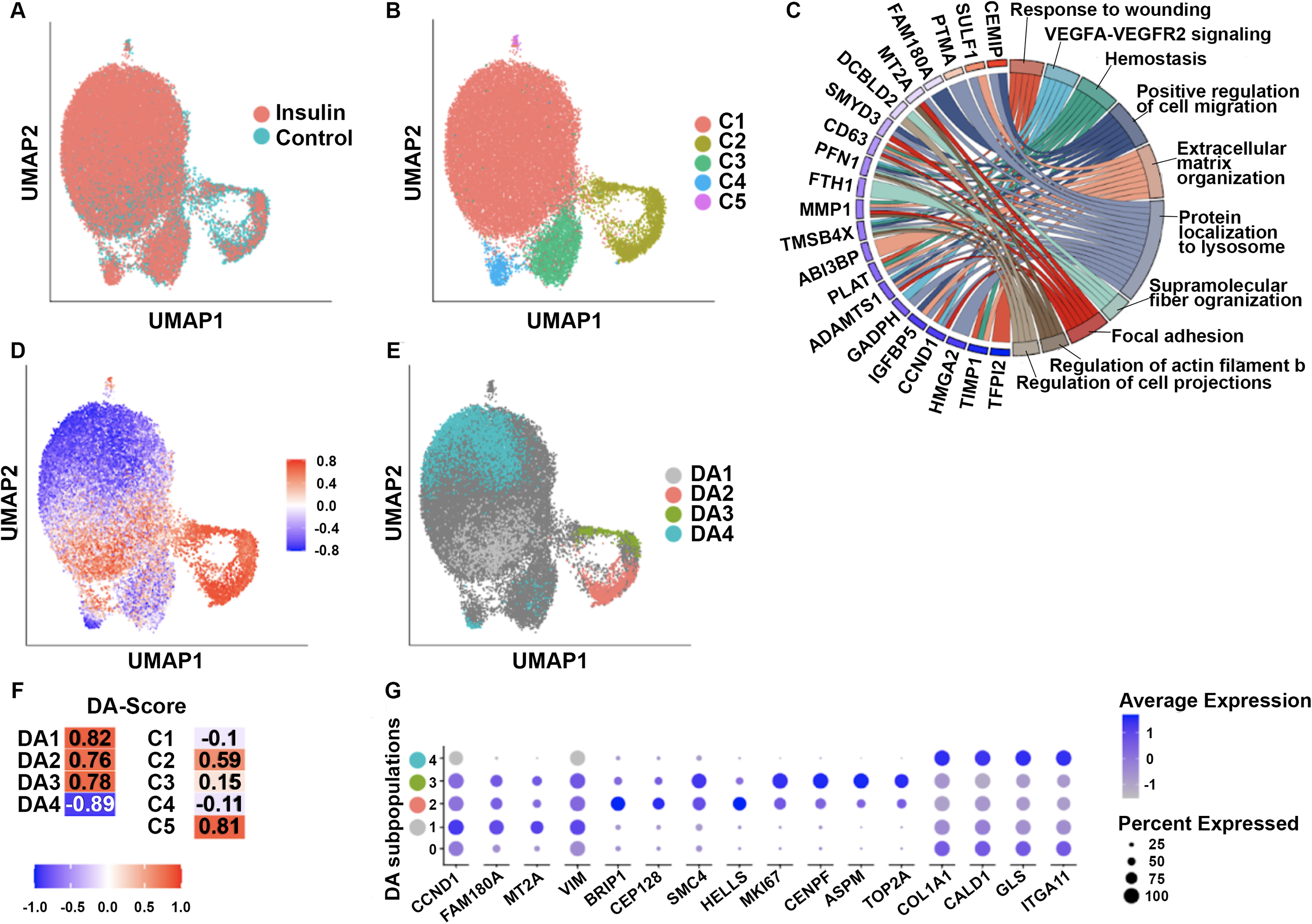
Single-cell RNA sequencing (scRNA-seq) and differential abundance analysis of MSCs primed with insulin or maintained in control media. Uniform manifold approximation and projection (UMAP) embedding of 31,674 MSCs was used to visualize transcriptional structure across insulin-primed and control conditions. Because samples were blinded during analysis, condition labels were initially reversed; after unblinding, negative scores correspond to features enriched in insulin-primed MSCs, while positive scores correspond to features reduced in insulin-primed MSCs.
DA Cluster 1 exhibited a myofibroblast-like transcriptional profile, characterized by high expression of contractile and adhesion-related genes, including collagen type I alpha 1 chain (COL1A1), caldesmon 1 (CALD1), glutaminase (GLS), and integrin subunit alpha 11 (ITGA11).23–26 These cells exhibited the lowest expression of proliferation markers among the DA clusters, consistent with an ECM-producing and contractile phenotype. This cluster may contribute to wound contraction and late-stage remodeling.
In DA Cluster 2, MSCs remained highly enriched in ECM-producing and adhesion-related genes, similar to Cluster 1, but exhibited mild downregulation of proliferation markers rather than a complete loss. These cells also displayed high expression of COL1A1, CALD1, GLS, and ITGA11, consistent with active extracellular matrix synthesis. This expression profile suggests that MSC populations in Cluster 2 may represent an ECM-remodeling MSC subpopulation with greater proliferative capacity than Cluster 1.
DA Cluster 3 contained MSCs with concurrent downregulation of cell cycle genes [structural maintenance of chromosomes 4 (SMC4), marker of proliferation Ki-67 (MKI67), centromere protein F (CENPF), abnormal spindle-like microcephaly-associated (ASPM), DNA topoisomerase II alpha (TOP2A)] 27 and upregulation of adhesion and ECM-related genes (COL1A1, ITGA11, CALD1, GLS). Compared with Clusters 1 and 2, these cells retained higher expression of select proliferation-related genes [cyclin D1 (CCND1), metallothionein 2A (MT2A), vimentin (VIM), and BRCA1 interacting protein C-terminal helicase 1 (BRIP1)],28–33 suggesting a transcriptional state that combines proliferative and ECM-remodeling features.
Finally, DA Cluster 4 represented a proliferative MSC population, characterized by strong upregulation of CCND1 and VIM, with low expression of ECM-related genes. These MSCs may function as a proliferative subpopulation within the hydrogel, providing a pool of cells with transcriptional features associated with expansion and regenerative potential.
Taken together, these results show that insulin priming is associated with altered abundance of MSC subpopulations with distinct proliferative (Cluster 4), ECM-remodeling, adhesion, and contractile gene expression signatures (Clusters 2 and 1). The increased abundance of ECM-producing (Cluster 3) and tissue-stabilizing (Clusters 2 and 1) subpopulations in insulin-primed MSCs is consistent with the enhanced wound closure and improved architecture observed in HEMI-treated wounds.
To further dissect the impact of insulin priming on MSC function, we performed differential gene expression analysis. Insulin-primed MSCs exhibited significant upregulation of key regenerative genes, including MT2A; discoidin, CUB and LCCL domain containing 2 (DCBLD2); insulin like growth factor binding protein 5 (IGFBP5); matrix metalloproteinase-1 (MMP1); profilin 1 (PFN1); and SMET and MYND domain containing 3 (SMYD3), which are associated with ECM remodeling, oxidative stress resistance, and immune modulation.32–37 Notably, insulin priming also downregulated fibrosis-associated genes, including cell migration-inducing protein (CEMIP), sulfatase 1 (SULF1), and prothymosin alpha (PTMA), which have been implicated in excessive scar formation and chronic inflammation.38–41 These transcriptional changes align with the histological findings of reduced fibrosis and enhanced tissue organization in HEMI-treated wounds.
Beyond ECM remodeling, insulin-primed MSCs exhibited a shift toward an immunosuppressive phenotype, as evidenced by upregulation of killer cell lectin-like receptor D1 (KLRD1) and sestrin 3 (SESN3), which are involved in immune suppression and oxidative stress resistance. 42 Conversely, unprimed MSCs showed higher expression of proinflammatory mediators such as C-X-C motif chemokine ligand 8 (CXCL8 or interleukin-8, IL-8), NLR family pyrin domain containing 11 (NLRP11), and serine protease inhibitor A3 (SERPINA3),43–45 indicating that insulin priming suppresses inflammation while fostering a regenerative microenvironment. These findings support the notion that HEMI treatment modulates the wound immune landscape, promoting faster inflammation resolution and optimized tissue repair.
Discussion
The findings of this study demonstrate that insulin priming enhances the regenerative potential of MSCs through paracrine-mediated effects rather than direct integration into the wound. By encapsulating MSCs in a hydrogel delivery system, continuous local release of bioactive factors was associated with accelerated healing, enhanced tissue organization, and reduced fibrosis. The observed improvement in wound healing supports the significance of insulin-mediated modulation of MSC function and its potential for clinical translation in regenerative medicine.
The rapid wound closure observed in the first 2 weeks of treatment suggests that insulin-primed MSCs accelerate the early phases of healing, likely through enhanced secretion of proregenerative factors that stimulate reepithelialization, fibroblast recruitment, and immune modulation. The histological findings support this notion, as HEMI-treated wounds exhibited fully regenerated epidermal, dermal, and hypodermal layers, with improved granulation tissue architecture and reduced fibrosis. In contrast, wounds treated with unprimed MSCs displayed incomplete reepithelialization, while control wounds exhibited partial but incomplete closure during the study period. No control wounds reached complete closure by week 5, highlighting the improved healing response observed in the HEMI-treated group.
The single-cell transcriptomic analysis revealed insulin-associated differences in MSC subpopulation abundance and gene expression state. Insulin-primed MSCs showed increased representation of subpopulations enriched for ECM-producing, adhesion-related, and tissue-stabilizing transcriptional signatures. Because trajectory analysis was not performed, these data should be interpreted as differences in population composition and transcriptional state rather than evidence of a defined lineage progression.
Beyond their role in ECM remodeling, insulin-primed MSCs exhibited significant transcriptional changes related to immune modulation, shifting toward an anti-inflammatory phenotype that likely contributed to faster inflammation resolution and reduced fibrosis. Upregulation of genes such as KLRD1 and SESN3, which are involved in immune suppression and oxidative stress resistance, suggests that insulin-primed MSCs create a proregenerative immune environment that optimizes tissue remodeling. Simultaneously, downregulation of fibrosis-associated genes, including CEMIP and SULF1, further supports the anti-scarring properties of insulin priming, which were reflected in the histological outcomes.
Several limitations should be noted. Wounds were not randomized because treatment assignment was based on spatial separation to reduce local crosstalk between treatment zones; therefore, residual spatial bias cannot be excluded. Multiple wounds were evaluated within each animal, which is common in large-animal wound studies but may introduce clustering effects. Animal attrition reduced the final number of animals completing the full protocol. Additional hydrogel-only, insulin-only, and immunohistochemical analyses would further isolate individual contributions to the observed healing response and are important areas for future work.
The use of human MSCs and rat ISCs in swine should also be interpreted in the context of the PEGDA encapsulation strategy. PEGDA hydrogels are designed to provide immunoisolation and have repeatedly been shown across multiple studies to permit xenogeneic cell function while allowing diffusion of soluble therapeutic factors.10–14,17–19 PEGDA-encapsulated pancreatic islets have restored normoglycemia in xenogeneic animal models,19,46,47 and PEGDA-encapsulated human islets have demonstrated function following implantation without evidence of immune rejection.48,49 In our prior work, this same PEGDA encapsulation system was used with ISCs and MSCs to significantly accelerate healing in a diabetic mouse model of delayed wound repair. 10 The present study extends that platform to a large-animal model, enabling evaluation of the identical cell-hydrogel system in a clinically relevant porcine burn model. Thus, encapsulated xenogeneic cells were used within a delivery system specifically selected to reduce immune-mediated clearance while preserving paracrine function.
Conclusions
These findings establish insulin priming as a promising tool to enhance MSC-mediated tissue repair by promoting transcriptional programs associated with ECM remodeling and immune modulation. The ability of insulin-primed MSCs to accelerate wound healing via sustained paracrine signaling, rather than direct engraftment, highlights the potential for hydrogel-encapsulated MSC therapies to overcome the limitations of traditional stem cell delivery. By leveraging DA-seq and scRNA-seq analyses, this study identifies insulin-associated MSC subpopulations and gene expression signatures that may contribute to improved regenerative outcomes.
From a clinical perspective, hydrogel-encapsulated, insulin-primed MSCs offer a localized and controlled delivery system that may reduce concerns related to poor cell retention and immune-mediated clearance. These findings suggest that strategic MSC priming could be tailored to optimize regenerative outcomes, with potential applications beyond burn treatment, including chronic wounds, musculoskeletal injuries, and fibrotic diseases.
Authors’ Contributions
R.O. conceived and designed the study, served as principal investigator, conducted data analysis, and prepared the article. C.L.C. contributed to study design, article preparation, and led the Massachusetts General Hospital (MGH) efforts. K.W. performed cell culture, hydrogel encapsulation, data analysis, and article writing. R.C. and A.T.D. assisted with data analysis. V.P., A.R.A., P.T., G.R., and A.G.L. conducted animal surgeries and husbandry, collected wound biopsies, contributed to data analysis, and participated in article preparation. All the authors reviewed the article.
Ethical Considerations
All animal experiments were approved by the Massachusetts General Hospital Institutional Animal Care and Use Committee (MGH IACUC) and were conducted in accordance with institutional guidelines and the United States National Institutes of Health Guide for the Care and Use of Laboratory Animals. The study included only male MGH miniature swine to reduce biological variability and align with prior wound healing protocols. While sex-specific analyses were not performed, future studies will aim to include both sexes to assess sex-based differences in wound healing. All experimental procedures were designed to ensure humane treatment of animals, minimize suffering, and use the minimum number of animals necessary to achieve statistically valid results. The research team supports diversity and inclusion in scientific collaboration and data interpretation. This study is reported in accordance with the ARRIVE guidelines (Animal Research: Reporting of In Vivo Experiments) 2.0 (https://arriveguidelines.org).
Footnotes
Data Availability
The datasets generated and/or analyzed during the current study have been deposited in Figshare (wound healing images) and in the public NCBI GEO repository (raw and processed transcriptomics data) and will be made publicly available upon publication. The data can be accessed at ![]() and GEO accession GSE307777, respectively.
and GEO accession GSE307777, respectively.
Disclosure Statement
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding Information
This material is based on the work supported by the National Science Foundation under grant no. CBET-2040657, the WiSTEM2D Scholar Award from Johnson & Johnson, and the Rutgers TechAdvance award no. AWD00004177.