
Abstract
Diabetes mellitus is a global public health problem, and impaired wound healing is a complication that significantly reduces patients’ quality of life. Dysregulation of mitochondrial homeostasis is a key pathological feature contributing to impaired wound healing in diabetes. This dysregulation increases oxidative stress, resulting in impaired energy metabolism, endothelial dysfunction, and prolonged inflammatory responses. Photobiomodulation (PBM) is a noninvasive therapy that has been successfully used to promote diabetic wound healing by modulating mitochondrial homeostasis via multiple mechanisms. In this review, we have systematically summarized the following roles of PBM in restoring mitochondrial homeostasis to accelerate diabetic wound healing: improving mitochondrial dysfunction and oxidative stress through cytochrome C oxidase in the electron transport chain, thereby enhancing oxidative phosphorylation and adenosine triphosphate production; modifying mitochondrial dynamics by inhibiting the expression of dynamin-related protein 1 and promoting mitofusin-2 expression to restore mitochondrial morphology and function; reducing inflammation and promoting macrophage polarization from the M1 to M2 phenotype; activating signaling pathways (e.g., VEGF, PI3K/AKT/mTOR/GSK3-β, AMPK, RAS/MAPK, JAK/STAT, NF-κB, TGF-β/Smad) to enhance cell proliferation and angiogenesis and resolve inflammation. Beyond monotherapy, this review synthesizes the burgeoning field of PBM in combination with advanced therapeutic strategies, such as hydrogels, nanomaterials, small-molecule drugs, adipose-derived stem cells, and extracellular vesicles, highlighting their synergistic potential for enhanced efficacy. Finally, this review critically addresses the prevailing challenges in clinical translation, particularly the lack of standardized treatment parameters, and proposes future research directions. This comprehensive overview aims to solidify the scientific foundation of PBM and inspire the design of integrated, precision therapeutic strategies for diabetic wound management.
Impact Statement
This review delineates the critical role of photobiomodulation (PBM) for overcoming diabetic wound healing impairment by restoring mitochondrial homeostasis. It integrates multifaceted evidence on the mechanisms of PBM—from improving cellular energetics and redox balance to modulating inflammation and activating prohealing pathways. Furthermore, it systematically explores the combination of PBM with next-generation therapeutic strategies. These insights not only advance the fundamental understanding of PBM but also directly inform the rational design of novel, effective treatments for diabetic chronic wounds, a major unmet clinical need.
Keywords
Introduction
Diabetes mellitus is a chronic metabolic disorder characterized by persistent hyperglycemia, which leads to progressive damage to blood vessels, nerves, and other organs. 1 In 2022, an estimated 828 million adults (aged 18 years and older) worldwide were living with diabetes, and this number continues to rise, posing a significant global public health challenge. 2 Among the various complications associated with diabetes, diabetic foot ulcers are particularly common. It is estimated that approximately 15–25% of patients with diabetes develop diabetic foot ulcers. Owing to the challenges in treatment, about 85% of these patients eventually require amputation, severely impairing their quality of life. 3
Mitochondria, central to cellular energy and redox homeostasis, are critically involved in impairing diabetic wound healing. Under hyperglycemic conditions, mitochondrial electron transport chain (ETC) dysfunction leads to reduced adenosine triphosphate (ATP) production, excessive reactive oxygen species (ROS) release, and disruption of mitochondrial homeostasis. These alterations trigger chronic inflammation, apoptosis, and vascular injury, ultimately delaying the wound healing process. 4
Confronting this multifaceted challenge, photobiomodulation (PBM) has emerged as a promising noninvasive therapeutic modality. PBM harnesses specific wavelengths of light, primarily in the red and near-infrared spectrum, which are absorbed by mitochondrial chromophores like cytochrome c oxidase (COX). This initial photonic event triggers a cascade of restorative processes, leading to improved ATP synthesis, attenuated oxidative stress, and resolution of inflammation. 5 While the therapeutic potential of PBM is recognized, a comprehensive and systematic synthesis of its mechanisms, particularly through the lens of mitochondrial homeostasis, is urgently needed to bridge the gap between experimental evidence and clinical translation.
Thus, this review is structured to build a comprehensive and mechanistic framework for understanding PBM’s role in diabetic wound healing. The narrative begins by deconstructing the pathogenic interplay between mitochondrial dysfunction and diabetic wound, establishing the foundational rationale for mitochondrial-targeted therapy. It then systematically examines the evidence for PBM’s capacity to recalibrate core aspects of mitochondrial homeostasis—including dynamics, bioenergetics, and quality control—as the central pillar of its therapeutic action. Building upon this mitochondrial premise, the discussion deciphers the ensuing cascade of molecular signaling that translates cellular restoration into tangible tissue repair, encompassing enhanced angiogenesis, proliferation, and inflammation resolution. Furthermore, the review looks ahead to the next frontier in treatment: the strategic integration of PBM with cutting-edge adjuncts such as smart biomaterials, stem cells, and nano-therapeutics, critically appraising the synergistic potential of these combinatorial approaches to overcome the complexity of diabetic wounds. Through this structured exploration, this review seeks to solidify the theoretical foundation for PBM and accelerate the development of effective, personalized treatment paradigms for diabetic wounds.
Mitochondria-Centered Oxidative Stress Delays Wound Healing in Diabetes
Mitochondrial homeostasis is crucial for wound repair, governing energy supply, redox balance, and immune modulation. In diabetes, hyperglycemia disrupts this homeostasis, leading to oxidative stress, dynamic imbalance, and impaired mitophagy. These dysfunctions collectively sustain inflammation, hinder angiogenesis, and promote apoptosis, ultimately delaying wound healing. The following sections dissect these interconnected mechanisms (Fig. 1).
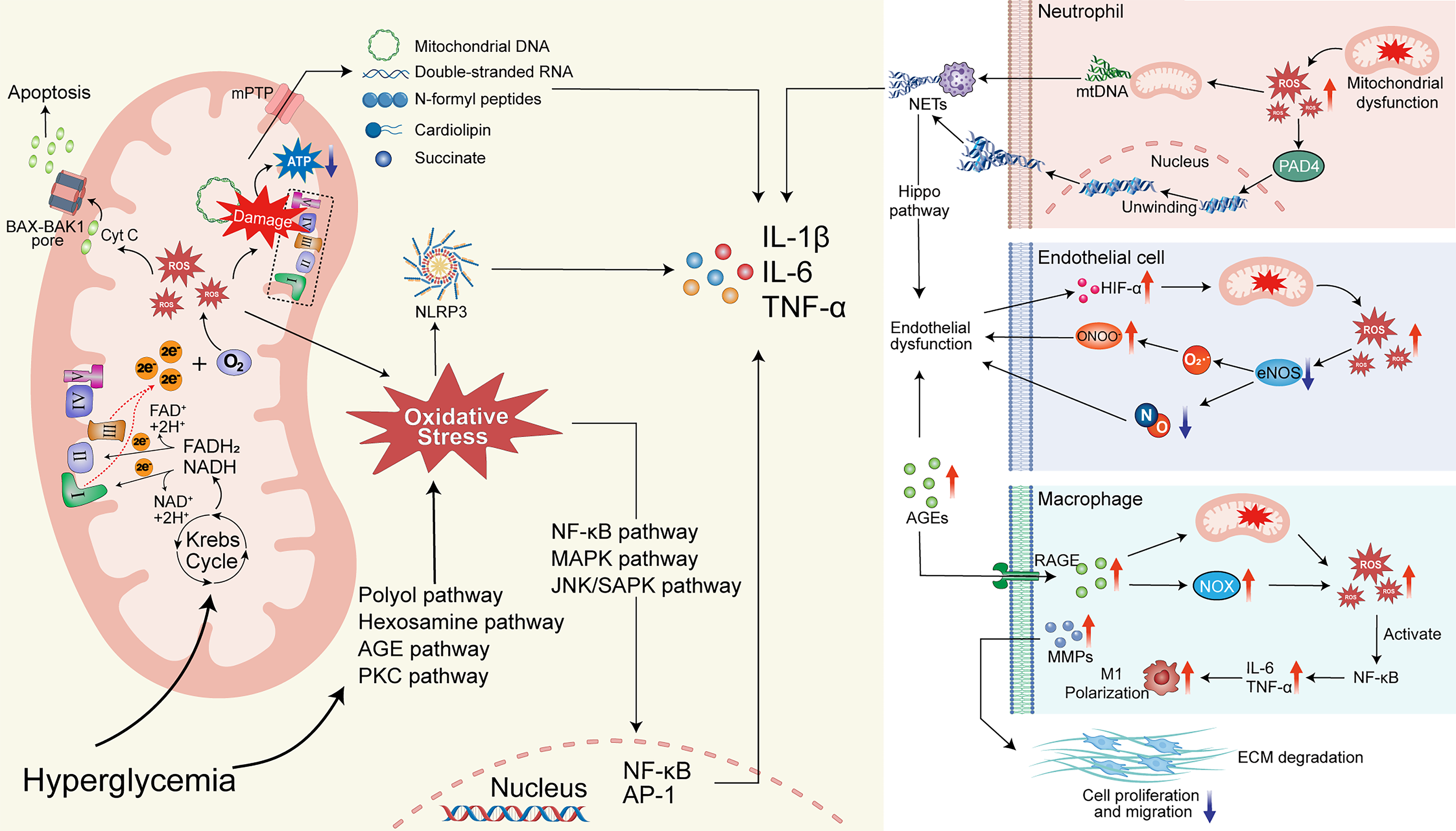
Mechanisms of mitochondrial dysfunction in diabetic cells and resulting changes in neutrophils, endothelial cells, and macrophages. The left panel depicts the mechanism by which high glucose levels in diabetic cells cause mitochondrial dysfunction, leading to increased oxidative stress, leakage of mitochondrial contents, elevated proinflammatory factor secretion, and cell apoptosis. The right panel illustrates the mechanism whereby increased oxidative stress from mitochondrial dysfunction in diabetic wounds promotes neutrophil-derived NETs release, disrupts endothelial cell function, and drives macrophage M1 polarization, resulting in heightened wound inflammation, endothelial dysfunction, and delayed wound healing.
Role of mitochondria in wound healing
Cellular energy metabolism relies on glycolysis in the cytoplasm and oxidative phosphorylation in mitochondria. 6 Under normal conditions, oxidative phosphorylation provides over 90% of cellular ATP. 7 The wound healing process progresses through hemostasis, inflammation, proliferation, and remodeling. Early hypoxia due to vascular injury shifts cellular metabolism toward glycolysis to meet energy demands, 8 with metabolic intermediates supporting cell proliferation and migration. 9 As oxygen supply is restored during proliferation and remodeling, oxidative phosphorylation becomes the primary energy source. 10 This metabolic reprogramming adapts to the changing energy requirements of healing. In addition, mitochondria generate metabolites (e.g., branched-chain amino acids, glutamine) that participate in signaling and biosynthesis, further promoting repair. 11
Mitochondria are a major source of ROS, which play a dual role in healing: moderate ROS act as signaling molecules that facilitate cell proliferation, migration, and antimicrobial defense, whereas excessive ROS cause oxidative damage and impair healing. During the early inflammatory phase, glycolysis and pro-inflammatory factors elevate ROS, promoting M1 macrophage activation for pathogen clearance. Later, enhanced mitochondrial oxidative phosphorylation drives macrophage polarization toward the pro-repair M2 phenotype, resolving inflammation and supporting tissue reconstruction. 10 Thus, mitochondria contribute to wound healing through energy supply, ROS regulation, and immune modulation.
Oxidative stress and mitochondrial homeostasis imbalance in diabetic wounds
Diabetic chronic wounds exhibit persistent oxidative stress driven by multiple pathways. Hyperglycemia activates alternative metabolic routes (polyol, hexosamine, PKC, AGE accumulation), overwhelming antioxidant defenses. 12 Concurrently, excessive glucose flux through the TCA cycle overloads the mitochondrial ETC, leading to electron leakage and superoxide production. 13 The resulting ROS damage ETC components, mitochondrial DNA, and metabolic enzymes, causing mitochondrial dysfunction, 14 which in turn generates more ROS—establishing a vicious cycle. 15
Elevated oxidative stress disrupts mitochondrial dynamics (fusion/fission) and mitophagy. Normally, fusion proteins (Mfn1/2, OPA1) and fission regulators (Drp1, FIS1) maintain mitochondrial morphology,16–18 while mitophagy (mediated by PINK1/Parkin/LC3) clears damaged mitochondria.19,20 Under oxidative stress, OPA1 hydrolysis and Mfn1/2 degradation impair fusion,21,22 while sustained Drp1 activation promotes excessive fission, 23 collectively causing mitochondrial fragmentation. This fragmentation perpetuates ΔΨm loss, ETC uncoupling, and further ROS bursts.24,25 Impaired PINK1/Parkin function also hampers mitophagy, leading to accumulation of dysfunctional mitochondria and exacerbating cellular damage. 17
Multidimensional mechanisms through which mitochondrial homeostasis impairment delays diabetic wound healing
Wound healing is energy-intensive. Mitochondrial dysfunction reduces ATP production, 26 failing to meet the high energy demands of keratinocytes, fibroblasts, and endothelial cells, thereby directly delaying repair. 27
Mitochondrial-derived oxidative stress triggers persistent inflammation via multiple pathways. It activates transcription factors (NF-κB, MAPK, NLRP3 inflammasome) that upregulate pro-inflammatory cytokines and chemokines.28–30 The NLRP3 inflammasome serves as a critical hub linking mitochondrial stress to inflammation during wound healing. Mechanistically, NLRP3 activation is regulated by mitochondria through multiple pathways, including the production of ROS, release of mitochondrial DNA, and modulation of cellular metabolism. 31 Damaged mitochondria also release mitochondrial DAMPs (mtDNA, dsRNA, succinate) via mPTP opening, intensifying innate immune activation.32–34 Oxidative stress furthermore accelerates AGE formation, which through RAGE/NF-κB signaling sustains M1 macrophage polarization and inhibits M2 transition, prolonging inflammation.29,35–38 Excessive inflammation upregulates MMPs and degrades ECM, hindering cell migration.30,39 Neutrophils in the diabetic wound environment become hyperactivated, forming excessive NETs via PAD4.40–42 NETs perpetuate M1 polarization via NLRP3, 43 inhibit angiogenesis via the Hippo pathway, 44 and induce fibroblast ER stress and apoptosis. 45
Increased mtROS in diabetes activates the PKC/NOX pathway while uncoupling eNOS, reducing vasodilatory and pro-angiogenic NO.46,47 NO also reacts with ROS to form peroxynitrite, further uncoupling eNOS and impairing angiogenesis. 48 Together with direct endothelial damage by AGEs, this causes severe vascular dysfunction and hypoxia, which further exacerbates mitochondrial dysfunction via HIF-α – forming a vicious cycle. 49
Mitochondria are central to the intrinsic apoptosis pathway. Oxidative stress-induced ΔΨm loss and outer membrane permeabilization allow cytochrome c release via BAX-BAK1 pores, activating caspases and triggering apoptosis.50–52 This depletes repair cells at the wound site, directly impeding healing.
PBM Promotes Diabetic Wound Healing by Modulating Mitochondrial Function
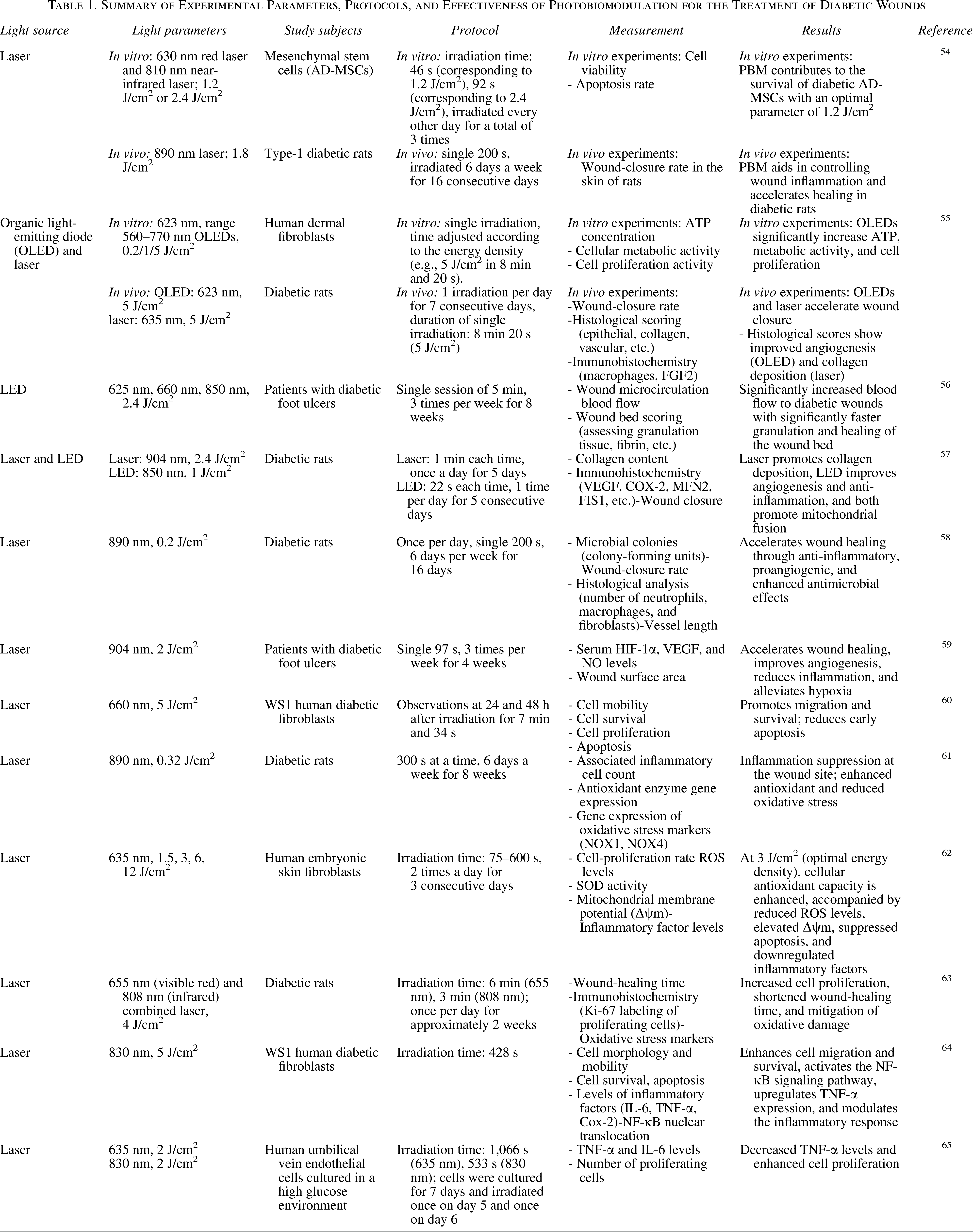
Given that mitochondrial homeostasis imbalance is a core mechanism underlying impaired diabetic wound healing, therapies targeting mitochondria have become a research hotspot. As a noninvasive physical therapy, PBM exhibits a mechanism of action highly aligned with mitochondrial function regulation. By modulating mitochondrial activity, PBM initiates a series of cellular cascades that promote diabetic wound healing. 5 PBM is considered a promising technology for treating diabetic wound healing due to its ability to stimulate tissue regeneration, alleviate pain and inflammation, and protect damaged or weakened tissues. 53 It has been extensively applied in various animal models, in vitro studies, and clinical research (Table 1).
Summary of Experimental Parameters, Protocols, and Effectiveness of Photobiomodulation for the Treatment of Diabetic Wounds
PBM alleviates oxidative stress and inflammation by improving mitochondrial function
In diabetic wounds, PBM effectively mitigates oxidative stress and suppresses excessive inflammatory responses by enhancing mitochondrial efficiency and reducing ROS production. Moheghi et al. demonstrated that in streptozotocin (STZ)-induced diabetic rat models, PBM treatment significantly reduced local inflammatory cell infiltration at the wound site. Along with this reduction, antioxidant enzyme activity increased while oxidative stress marker expression decreased, indicating that PBM regulates inflammatory responses and accelerates healing by improving mitochondrial redox status. 61
PBM restores fibroblast function and promotes extracellular matrix synthesis
Fibroblasts are responsible for synthesizing extracellular matrix components such as collagen during wound healing, 66 but their function is often suppressed in diabetic environments. In a single-cell transcriptomic study of diabetic foot ulcers, a unique subset of fibroblasts was found to be overexpressed in diabetic wounds: healing-enriched fibroblasts, which overexpress MMP1, MMP3, MMP11, and HIF1A, are associated with ECM remodeling and immune-/inflammation-related gene expression, 67 indicating their crucial role in wound healing. As opposed to that in normal wounds, healing is delayed in diabetic wounds due to a hyperglycemic environment, which results in impaired phenotypic switching of fibroblasts, imbalance in ECM synthesis and degradation, impeded angiogenesis, and decreased cell proliferation and migration. 68 PBM directly enhances fibroblast proliferation and metabolic activity by boosting mitochondrial energy metabolism. Studies demonstrate that PBM-treated human dermal fibroblasts cultured in high-glucose conditions exhibit increased ATP production and enhanced proliferation capacity. In diabetic animal models, PBM also promotes collagen deposition, granulation tissue formation, and angiogenesis at wound sites, thereby accelerating the repair process. 55
PBM enhances angiogenesis and improves local wound microenvironment
Impaired angiogenesis is a key factor in diabetic wound healing failure. PBM promotes endothelial cell function and neovascularization by upregulating nitric oxide levels and modulating pathways such as vascular endothelial growth factor. 59 Both clinical and animal studies confirm that PBM treatment improves local wound blood flow, alleviates hypoxia, and normalizes the expression of angiogenesis-related factors. This indicates that PBM can create a more favorable microenvironment for healing by improving mitochondrial-mediated vascular regulatory mechanisms. 69
Molecular Mechanisms and Signaling Pathways of PBM in Regulating Mitochondrial Homeostasis
The core mechanism of PBM involves specific targeting of endogenous chromophores within mitochondria. COX, a terminal enzyme of the mitochondrial ETC, is a primary photoreceptor for PBM, containing light-absorbing heme and copper centers.5,70,71 Under diabetic conditions, COX dysfunction impairs electron transport, increasing ROS and reducing ATP. 72 PBM promotes NO dissociation from COX, restoring its enzymatic activity.5,73 Specifically, PBM delivers photons in the red (600–700 nm) and near-infrared (700–1,100 nm) range that are absorbed by the heme α3–CuB binuclear center of COX. 74 This absorption primarily targets the CuB+-NO+ complex, which exhibits a broad absorption band spanning 600–900 nm arising from d–d transitions of CuB+ under different redox and ligand environments.74,75 Photon absorption promotes an electron in the d orbital of CuB+ to an excited state, weakening the Cu+-NO+ coordination bond and leading to NO dissociation from the active site.75,76 Concomitantly, CuB2+ reestablishes its coupling with heme α3 (Fe³+), allowing oxygen to bind to heme α3. This sequence restores COX activity, reestablishes the proton gradient, and reduces electron leakage and oxidative stress.74–77 This enhances mitochondrial membrane potential (ΔΨm), improves oxidative phosphorylation efficiency, increases ATP and cAMP production, and reduces oxidative stress. 78 This initial mitochondrial response triggers downstream signaling to restore homeostasis (Fig. 2).
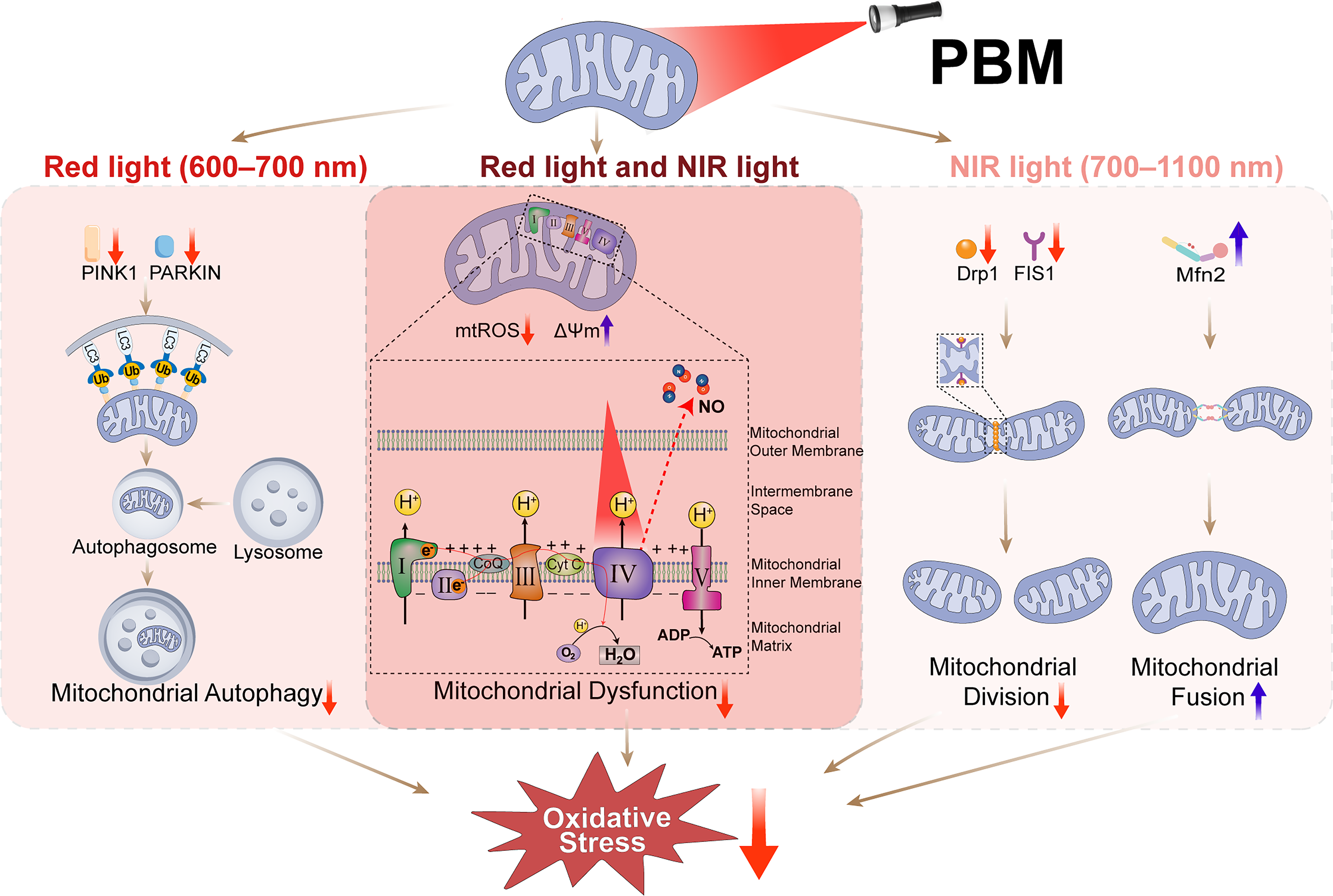
Distinct effects of red and near-infrared (NIR) PBM on mitochondrial homeostasis in diabetic cells. PBM employs red (600–700 nm) and NIR (700–1,100 nm) wavelengths, which are absorbed by distinct chromophores within COX. This absorption promotes NO dissociation, enhances electron transfer efficiency, reduces ROS accumulation, and restores mitochondrial ΔΨm. Red light predominantly suppresses excessive mitophagy by downregulating PINK1 and PARKIN expression. NIR light primarily restores mitochondrial dynamics by downregulating fission proteins (Drp1, FIS1) and upregulating the fusion protein Mfn2, shifting the balance from pathological fragmentation toward protective fusion. COX, cytochrome c oxidase; PBM, photobiomodulation; ROS, reactive oxygen species.
PBM improves mitochondrial dynamics imbalance
In diabetic conditions, excessive mitochondrial fragmentation is primarily driven by upregulation of the fission regulator Drp1 and its adaptor FIS1, alongside downregulation of the fusion mediators Mfn1, Mfn2, and OPA1. 79 Studies have demonstrated that PBM can reverse this pathological imbalance. In diabetic wound tissues, PBM (850 nm LED) significantly increased Mfn2 expression and decreased FIS1 expression. 57 Similarly, in diabetic peripheral neuropathy, PBM (904 nm laser) reduced Drp1 and increased Mfn2 in dorsal root ganglia and sciatic nerve. 80 In a rat model of Alzheimer’s disease, long-term PBM treatment decreased Drp1, FIS1, and MFF while increasing Mfn1 and OPA1. 81 Collectively, these findings indicate that PBM promotes the formation of elongated, interconnected mitochondrial networks by coordinately modulating fission and fusion protein expression, thereby supporting efficient ATP production and maintaining mitochondrial integrity.57,80–82
PBM improves mitochondrial dysfunction and regulates metabolic reprogramming
Mitochondrial dysfunction in diabetic wounds forces cells into excessive glycolysis. PBM, via mitochondrial activation, corrects this state. It reduces oxidative stress and alleviates mitochondrial dysfunction in diabetic mouse wounds. 69 In diabetic adipose-derived stem cells (ADSCs), PBM reduces mtROS, enhances ΔΨm, and increases ATP synthesis. 83 Importantly, PBM restores physiological metabolic balance by improving ETC efficiency while reducing excessive ROS production—rather than forcibly driving OXPHOS in an uncontrolled manner.83,84 This improvement drives metabolic reprogramming from glycolysis to efficient oxidative phosphorylation, supporting the high energy demands of reparative cells (fibroblasts, endothelial cells) and promoting macrophage polarization from M1 toward the prorepair M2 phenotype. 85 Consequently, proinflammatory factors (IL-6, TNF-α) decrease, anti-inflammatory IL-10 rises, and NLRP3 inflammasome activation is suppressed, optimizing the wound microenvironment. 84
Regulatory role of PBM in mitochondrial autophagy
Mitophagy, crucial for clearing damaged mitochondria, is impaired in diabetes. 86 Interestingly, PBM reduces oxidative stress but does not upregulate key mitophagy proteins like PINK1 and PARKIN; it may even decrease their expression.83,87 This does not represent a conflict but rather reflects a shift in the mode of mitochondrial quality control: by directly enhancing mitochondrial function and reducing organellar damage, PBM lowers the burden of dysfunctional mitochondria, thereby diminishing the compensatory demand for sustained mitophagy flux.
Regulation of key signaling pathways
Building upon the restoration of mitochondrial homeostasis (the primary response), PBM orchestrates a secondary wave of signaling reprogramming that is crucial for diabetic wound repair. By rectifying the core mitochondrial deficits—reducing oxidative stress (ROS), replenishing cellular energy (ATP), and restoring key signaling molecules (NO, Ca2+)—PBM creates a microenvironment conducive to the reactivation of multiple healing-related pathways.88–91
This mitochondrial-mediated signaling shift manifests as the coordinated activation of prorepair and proliferative pathways. Specifically, the VEGF pathway is upregulated via HIF-1α stabilization, rescuing angiogenesis.89,92–94 Key metabolic and survival pathways, including PI3K/AKT/mTOR/GSK3-β and RAS/MAPK, are reactivated, promoting cell proliferation, migration, and inhibiting apoptosis.57,95–106 Concurrently, the AMPK pathway is activated to restore cellular energy homeostasis.90,91,103,107,108 In parallel, PBM stimulates repair-oriented immune signaling, such as the JAK/STAT and TGF-β/Smad pathways, which regulate inflammation and promote macrophage polarization toward the prohealing M2 phenotype.109–117 This is coupled with the suppression of the overarching proinflammatory drive mediated by the NF-κB pathway.62,118–124
These pathways do not operate in isolation but form a synergistic network (Fig. 3). For instance, AMPK activation can cross-talk with PI3K/AKT signaling to further enhance cell proliferation, 103 while the dampening of NF-κB activity is reinforced by the TGF-β/Smad pathway.115,116 Collectively, this PBM-induced signaling network counteracts the diabetic impairment by simultaneously promoting angiogenesis, enhancing the proliferation and migration of fibroblasts and keratinocytes, and resolving chronic inflammation. Thus, the regulation of downstream signaling pathways represents a critical integrative mechanism through which primary mitochondrial improvements translate into accelerated tissue repair.62,89,98,106,120
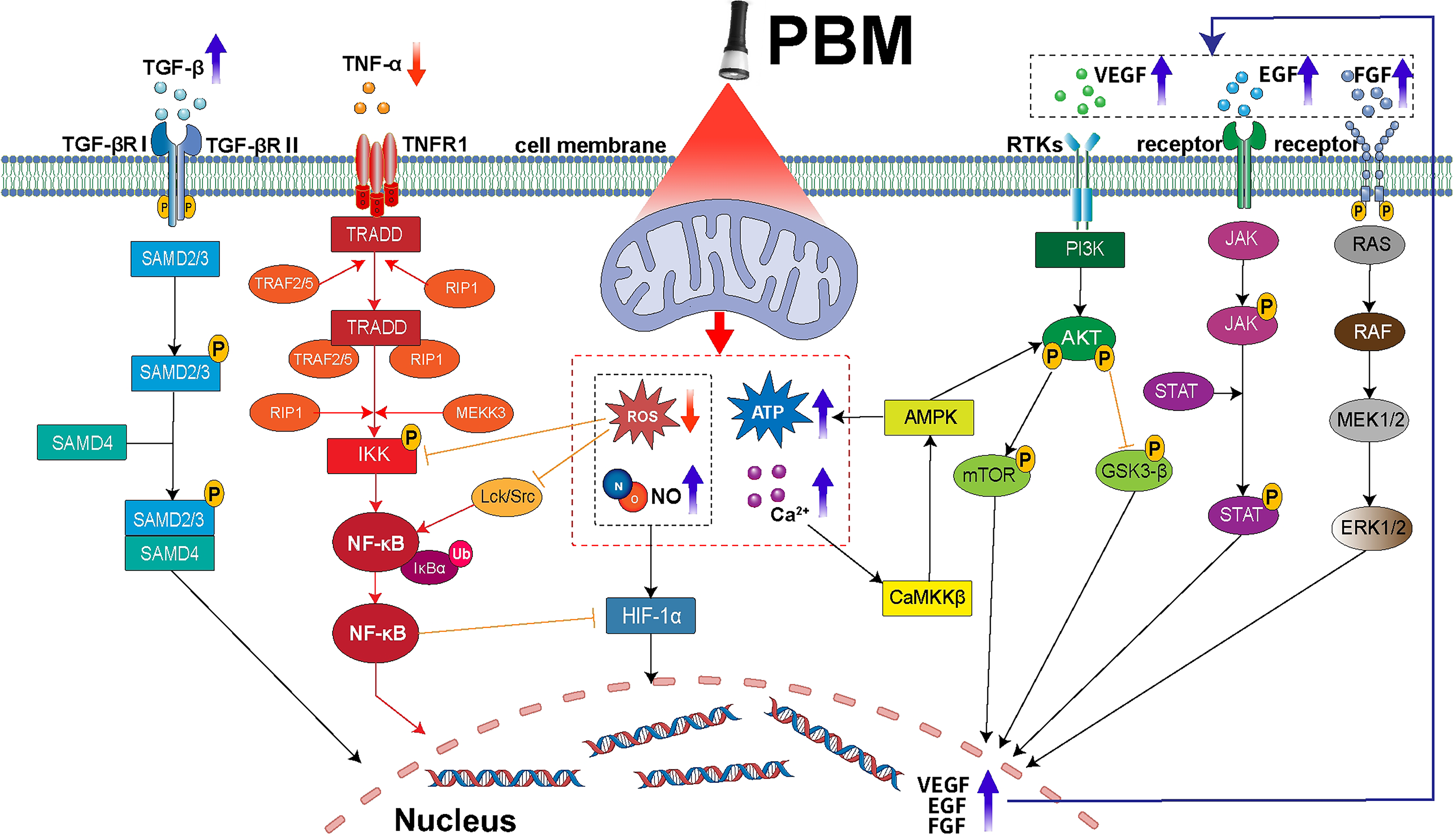
Schematic diagram of intracellular signaling pathways in diabetic cells associated with PBM effects. After PBM treatment on diabetic cells, intracellular ROS levels decrease, while ATP, NO, and Ca2+ concentrations increase. This leads to elevated secretion of cytokines such as VEGF, EGF, FGF, and TGF-β, along with reduced TNF-α levels. Consequently, the VEGF, PI3K/AKT/mTOR/GSK3-β, AMPK, RAS/MAPK, JAK/STAT, and TGF-β/Smad signaling pathways are activated, while the NF-κB pathway is inhibited. Shifts in these signaling pathways promote angiogenesis and cell proliferation, and differentiation in diabetic wounds, thereby controlling inflammation and accelerating wound healing. PBM, photobiomodulation; ROS, reactive oxygen species.
Challenges in Clinical Translation and Prospects of Multimodal PBM Strategies for Diabetic Wounds
Despite demonstrating clear molecular mechanisms and substantial preclinical evidence in regulating mitochondrial homeostasis and promoting wound healing in diabetes, PBM faces a series of practical challenges in clinical translation and widespread application. First, the significant heterogeneity and lack of standardized treatment parameters constitute a core bottleneck limiting the reproducibility of its efficacy. Currently, no optimal parameter set has been established for wound healing effects.30,125 The biphasic dose–response is a hallmark of PBM: low-to-moderate energy densities typically elicit stimulatory effects on cellular metabolism, proliferation, and repair, whereas excessive energy densities may lead to inhibitory or even detrimental outcomes, including increased oxidative stress, apoptosis, or thermal damage (Table 2).117,136 This nonlinear relationship underscores the critical importance of precise dosimetry, as the optimal therapeutic window varies depending on cell type, tissue context, and disease state. Importantly, the biphasic response is not limited to energy density alone; other PBM parameters such as power density and treatment frequency also exhibit dose-dependent effects on cellular behavior. For example, applying the same energy density at different power densities may yield different biological outcomes, highlighting the need for comprehensive parameter optimization rather than focusing solely on total flux.126,127 Furthermore, clinical translation faces multiple challenges: patient variability and complex local wound microenvironments (e.g., hypoxia, infection) significantly influence phototherapy response, yet personalized treatment protocols capable of real-time monitoring and dynamic parameter adjustment remain lacking. 124 Concurrently, high costs and specialized operation requirements for medical devices, coupled with the imprecision of home-use devices and the burden of long-term treatment, all impact treatment accessibility and patient compliance.
Summary of Biphasic Dose Responses in PBM Across Different Cell Types and Experimental Models
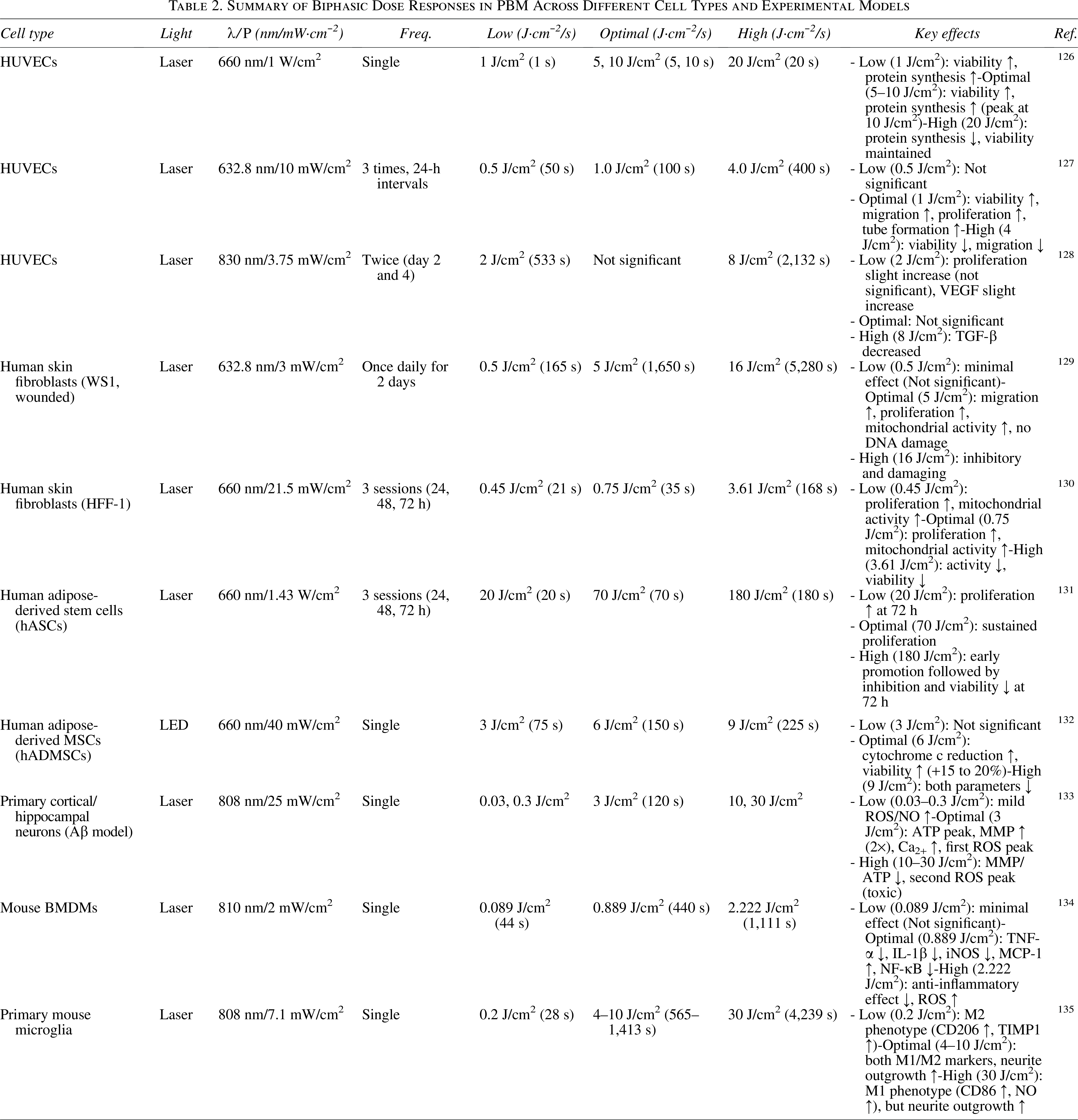
In addition to parameter standardization, clinical translation of PBM for diabetic wound healing faces challenges in light delivery and tissue penetration. Light penetration is wavelength-dependent: longer near-infrared wavelengths achieve deeper penetration (2–15 mm) than red light (1–3 mm). 5 However, diabetic foot ulcers often present with thick eschars and necrotic tissue that act as physical barriers, attenuating photon delivery to the viable wound bed. In addition, skin pigmentation (melanin) acts as a competitive chromophore. Individuals with skin of color exhibit increased photosensitivity, with a 50% lower maximum tolerated dose compared with lighter-skinned individuals, and are at higher risk for adverse effects at higher fluences.5,125 These factors necessitate individualized dosimetry accounting for wound depth, tissue composition, and pigmentation. Future advances may include real-time optical monitoring to ensure adequate tissue penetration and uniform dosing.
Precisely due to these challenges, monotherapy with PBM demonstrates limited efficacy when addressing the highly heterogeneous pathological microenvironments of diabetic wounds. Moreover, the pathogenesis of diabetic wounds involves multiple intertwined pathways such as persistent inflammation, impaired angiogenesis, bacterial biofilm formation, and cellular dysfunction,137,138 against which single PBM therapy shows limited efficacy. 139 Consequently, current research trends are shifting toward synergistically integrating PBM with other therapeutic strategies to develop multitargeted, synergistic comprehensive treatment regimens. This chapter will systematically explore the potential mechanisms and application prospects of combining PBM with various strategies—including hydrogels, nanomaterials, small-molecule drugs, ADSCs, and extracellular vesicles (EVs)—in diabetic wound repair (Fig. 4), aiming to provide theoretical guidance for achieving more optimal healing outcomes.
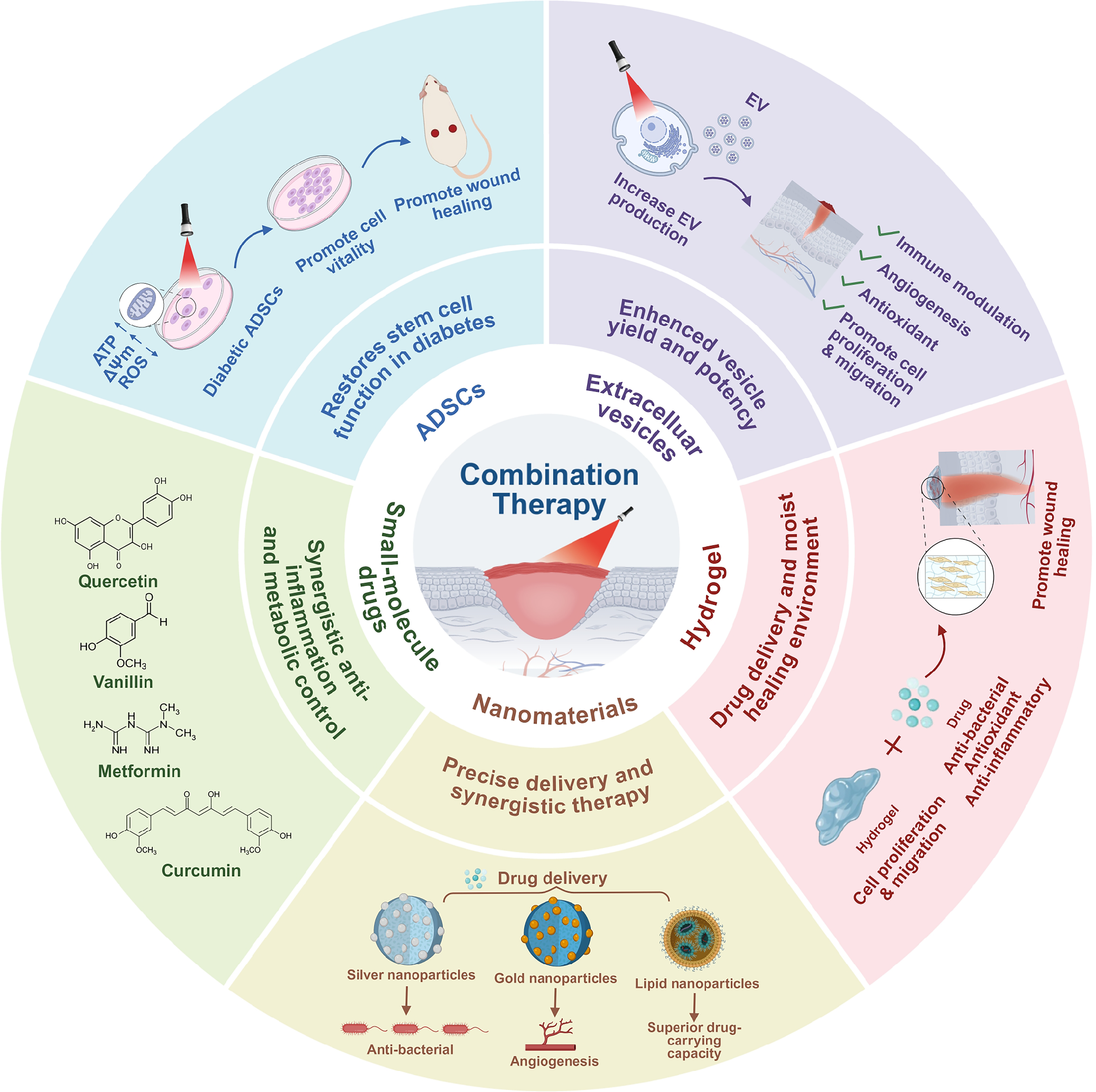
PBM promotes diabetic wound healing when combined with multiple therapeutic strategies. By improving mitochondrial function, reducing oxidative stress and inflammatory responses, and promoting angiogenesis and cell proliferation, PBM synergistically regulates the diabetic wound microenvironment through multiple targets alongside hydrogels, nanomaterials, small-molecule drugs, ADSCs, and EVs. This approach accelerates tissue repair and regeneration. ADSC, adipose-derived stem cell; EV, extracellular vesicle; PBM, photobiomodulation.
Combined application of PBM and hydrogels
Wound dressings are crucial in diabetic wound care. Conventional dressings like gauze provide hemostasis, absorb exudate, and protect wounds but cannot actively regulate the healing microenvironment. 140 In contrast, hydrogels—made from natural or synthetic polymers with a 3D network—offer high biocompatibility, water content, and breathability. They mimic the native ECM, providing a moist environment and structural support for cell migration and tissue regeneration. In addition, hydrogels can locally deliver antimicrobial and anti-inflammatory agents to promote healing.141,142
Combining PBM with hydrogels enables light-responsive synergistic therapy, depending on the hydrogel’s optical properties. For example, Wachał et al. confirmed that the HydroAid hydrogel transmits over 92% light at PBM wavelengths (660 nm and 808 nm), 143 minimizing energy loss and allowing effective light delivery. Building on this, functionalized hydrogels enhance synergy with PBM. Aragão-Neto et al. utilized a policaju-chitosan (POLI-CHI) hydrogel, which maintained a moist, anti-inflammatory environment, making cells more responsive to PBM and improving migration, proliferation, and collagen synthesis. 144 He et al. designed a catechol-modified gelatin methacrylamide (GelMAc) loaded with mesoporous polydopamine nanoparticles (MPDA@Ce6) (Fig. 5a), integrating photodynamic therapy with PBM. High-power 660 nm light triggered ROS for sterilization, while low-power PBM then promoted fibroblast activity and tissue repair after infection control (Fig. 5b and c). 146
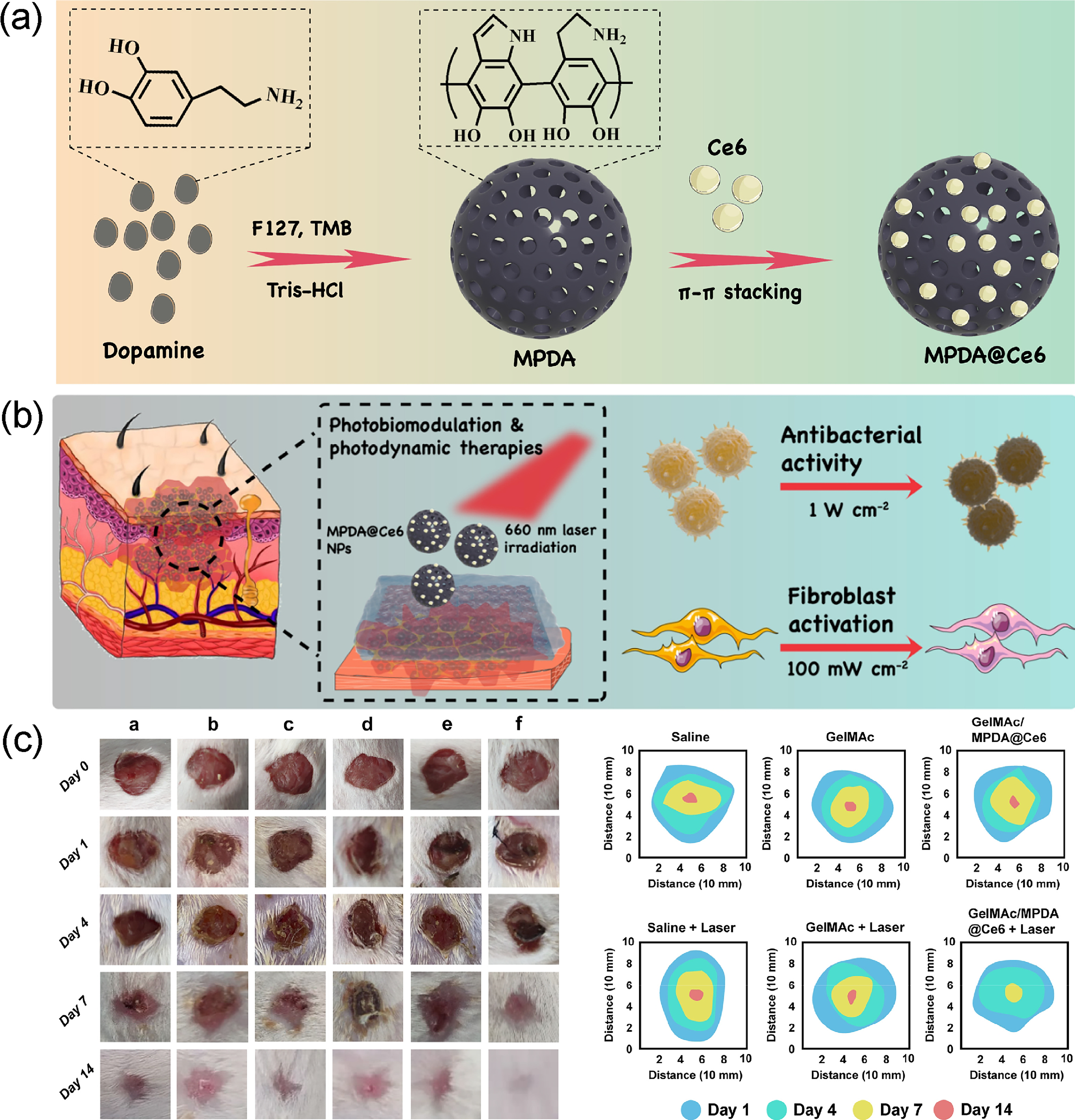
Combined application of PBM and nanomaterials
Nanomaterials, defined as substances with diameters of 1–100 nm, possess unique physicochemical and biological properties that enable multifunctional promotive effects across wound healing stages such as hemostasis, antibacterial action, inflammation modulation, and proliferation (Fig. 6).148–150 The combination of PBM with nanomaterials offers synergistic benefits for diabetic wounds. PBM modulates cellular energy metabolism, reduces inflammation, and enhances the proliferation and migration of fibroblasts and endothelial cells. 151 Meanwhile, nanomaterials—through intrinsic properties or as carriers for drugs and growth factors—enable targeted delivery and controlled release, helping to establish a healing-conducive regulatory network in the diabetic wound microenvironment. 152

Therapeutic strategies of diabetic wounds nanomedicines. Nanomaterials can promote diabetic wound healing by downregulating blood glucose, promoting homeostasis, anti-inflammatory, anti-infection, regulating angiogenesis, et al. Reproduced from ref. 147 with permission from Wiley-VCH GmbH. Copyright (2022).
Studies have confirmed this synergistic effect: silver nanoparticles combined with photobiostimulation enhance antimicrobial activity and promote wound healing 153 ; gold nanoparticles combined with photobiostimulation improve angiogenesis 154 ; and curcumin-loaded iron oxide nanoparticles combined with photobiostimulation overcome curcumin’s poor solubility, significantly enhancing antimicrobial, anti-inflammatory, and healing effects. 155 Functionalized nanocarriers such as solid lipid nanoparticles—loaded with lauric acid and tea tree oil and used with PBM—significantly promote fibroblast migration. 156 Hyaluronic acid-modified gold nanoparticles combined with PBM increased levels of anti-inflammatory cytokines (IL-4, IL-10) and growth factors (TGF-β, FGF), promoted collagen deposition, and facilitated the transition from acute to chronic inflammation in rat models. 157
Despite their efficacy, nanomaterials’ potential toxicity—such as allergic reactions, DNA damage, hemolysis, and organ toxicity—requires careful consideration in clinical applications.158–161 Future research should prioritize biosafety evaluations to enable broader and safer clinical use.
Synergistic therapy of PBM and small-molecule drugs
Small-molecule drugs, whether natural or synthetic, show great promise for treating diabetic wounds due to their established pharmacological profiles and cost-effectiveness. Studies indicate that these drugs synergize effectively with PBM to accelerate diabetic wound healing.162–164 Regarding anti-inflammatory and antioxidant effects, Ahmed et al. found that quercetin combined with PBM reduces proinflammatory cytokines (IL-1β, TNF-α), raises IL-10, and alleviates oxidative stress. 162 Erukainure et al. showed that vanillin with PBM boosts antioxidant enzyme activity, restores glyoxalase 1 function to reduce AGE accumulation, and inhibits 5-lipoxygenase, thus improving the wound’s glyco-oxidative stress microenvironment. 163 In terms of tissue repair, Bagheri et al. reported that metformin plus PBM in type 2 diabetic rats enhanced wound tensile strength, promoted fibroblast proliferation and neovascularization, and accelerated the transition from inflammation to proliferation. 164 In type I diabetic rats, Soleimani et al. demonstrated that curcumin with PBM exerts a strong synergistic antibacterial effect, significantly reducing S. aureus colonies and enhancing the wound’s anti-infective capacity to support regeneration. 165
Combined application of PBM and adipose-derived stem cells
Stem cells are widely studied in tissue engineering and regenerative medicine due to their multipotency and self-renewal capacity. 166 ADSCs are commonly used for skin wound healing because of their differentiation and paracrine functions. 167 However, in diabetic wounds, ADSC efficacy is limited by the hostile microenvironment and intrinsic dysfunction of diabetic-derived ADSCs, including impaired angiogenesis.168–171 Enhancing cellular function is therefore crucial for autologous ADSC therapy in diabetes. PBM reduces mtROS, increases ΔΨm, and boosts ATP synthesis in diabetic ADSCs, improving their viability and function. 83 Both in vivo and in vitro studies show that combining PBM with ADSCs synergistically enhances diabetic wound healing.58,145,147,172 For example, Tadi et al. reported that PBM plus ADSCs produced stronger anti-inflammatory and proliferative effects than either alone in a type 1 diabetic rat model of skin wound healing, 145 and Ebrahimpour-Malekshah et al. corroborated these findings and further indicated that this combinatorial approach possesses antibacterial and proangiogenic properties. 58 In addition, this combination modulates inflammation via IL-1β and microRNA-146a regulation. 147
Combined application of PBM and extracellular vesicles
EVs are cell-secreted phospholipid bilayer-enclosed vesicles carrying functional nucleic acids, proteins, and lipids. They serve as key mediators of intercellular communication, regulating cellular behavior and tissue homeostasis. 173 EVs are classified by size into exosomes, microvesicles, membrane particles, and apoptotic vesicles. 174 In skin regeneration, EV-based cell-free therapies show promise by influencing wound repair through homing/recruitment, immunomodulation, antiapoptosis, anti-inflammation, and angiogenesis (Fig. 7a).175,176 EVs for skin repair are derived from mesenchymal stem cells, plant extracts, engineered cells, and probiotics,177–180 but their clinical use is limited by low yield and unstable components. PBM has emerged as a noninvasive method to enhance EV production and function. Chang et al. found PBM (830 nm, 5 J/cm2) increased EV concentration from ADSCs by 6.25-fold while improving cell viability and migration. 181 Mechanistically, PBM promotes EV secretion by boosting mitochondrial function, metabolism, and membrane transport. Hyun et al. observed that red light (610 nm) irradiation of stem cells produced nanovesicles (R-NVs) that restored aged fibroblast viability to youthful levels, showing strong healing potential (Fig. 7b–e). These R-NVs upregulated stem cell factors (e.g., KLF4, c-Myc) and angiogenesis-related factors (e.g., VEGF, FGF2) and were enriched with miRNAs linked to antiaging and proliferation. 182 Although promising, PBM-EV combined strategies remain early-stage. Future work should clarify how PBM parameters (wavelength, energy density, etc.) affect EV yield, composition, and function from different sources. High-throughput proteomics and transcriptomics could reveal molecular mechanisms, expanding the strategy’s potential in regenerative medicine.

Summary and Future Perspectives
PBM has emerged as a powerful noninvasive therapeutic strategy demonstrating significant potential in restarting the stalled healing process of diabetic wounds. This review elucidates that PBM’s promotion of diabetic wound repair primarily stems from its core capability to regulate mitochondrial homeostasis. PBM restores cellular energy supply by enhancing ATP synthesis, reestablishes redox balance by scavenging excess ROS, and resolves pathological inflammation by promoting macrophage polarization toward the M2 phenotype. These actions, mediated through the activation of multiple signaling pathways, collectively reverse the adverse microenvironment of diabetic wounds, promoting angiogenesis, cell proliferation, and tissue remodeling. Furthermore, current research frontiers have expanded to the combined application of PBM with various advanced strategies, including hydrogels, nanomaterials, small-molecule drugs, ADSCs, and their exosomes. These combination strategies demonstrate significant potential to overcome the complex pathological environment of diabetic wounds through multitarget synergistic effects, representing a crucial shift toward comprehensive, precision-based therapeutic paradigms.
However, translating PBM into a frontline standard therapy still faces major challenges. The most pressing issue is the lack of standardized treatment parameters. Significant variations in wavelength, energy density, power density, and treatment protocols across existing literature hinder interstudy comparability and consistent clinical efficacy replication. Future large-scale, multicenter clinical trials are needed to systematically optimize parameters, aiming to replace one-size-fits-all approaches with personalized protocols dynamically adjusted in spatiotemporal terms according to distinct wound healing phases (e.g., early inflammation and late proliferation). Adaptive dosing strategies can be guided by computational modeling and real-time sensing technologies. Furthermore, key mechanistic questions remain unresolved. Resolving these requires leveraging advanced tools like animal models of mitochondrial autophagy and single-cell RNA sequencing to dissect dynamic, cell-specific responses within the wound microenvironment.
Most of the preclinical studies included in this review utilized STZ-induced type 1 diabetes models, which do not fully replicate the insulin resistance and chronic low-grade inflammation characteristic of type 2 diabetes. Although both conditions share mechanisms involving mitochondrial dysfunction and oxidative stress, future studies will need to employ type 2 diabetes models (such as diet-induced obesity or db/db mice) to validate the translational value of PBM in the context of type 2 diabetes.
In summary, PBM undoubtedly represents a cornerstone in the future arsenal of diabetic wound therapy. However, fully unlocking its potential requires concerted efforts to standardize its application, deepen mechanistic understanding, and creatively integrate it with advanced adjunct therapies. By bridging these gaps, PBM holds promise to evolve from a promising physical therapy modality into a core component of precision medicine for chronic wounds.
Authors’ Contributions
Conceptualization, C.W. and S.X.; methodology, F.L. and Y.M.; investigation, F.L. and Z.W.; writing–original draft preparation, F.L.; writing–review and editing, C.W., Y.M., and S.X.; visualization, Z.W.; supervision, C.W. and S.X.; project administration, C.W.; funding acquisition, C.W. and S.X. All authors have read and agreed to the published version of the article.
Footnotes
Author Disclosure Statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Funding Information
This work was supported by the Stomatological Hospital, School of Stomatology, Southern Medical University, under Grant number KQIIT2021001; and PY2024007.