
Abstract
Background
Chronic rhinosinusitis without nasal polyps (CRSsNP) represents a phenotype of CRS, whose immunological mechanisms are still unclear. So far there are neither suitable biomarkers to determine the course of the disease nor an individual therapy.
Objective
The purpose of this study was to characterize the CRSsNP endotype by identifying and validating non-invasive proteomic biomarkers.
Methods
A highly-multiplexed proteomic array consisting of antibodies against 2000 proteins was used to identify proteins that are differentially expressed in the nasal mucus of the CRSsNP and control groups (n = 7 per group). The proteins identified to be most differentially expressed were validated in matched nasal mucus samples using western blots and enzyme-linked immunosorbent assay (ELISA). Validation was also done in a second cohort using western blots (CRSsNP n = 25, control n = 23) and ELISA (n = 30 per group). Additionally, immunohistochemistry in CRSsNP and control tissue samples was performed to characterize the selected proteins further.
Results
Out of the 2000 proteins examined, 7 from the most differentially expressed proteins were chosen to be validated. The validation results showed that 4 proteins were significantly upregulated in CRSsNP mucus, including macrophage inflammatory protein-1beta (MIP-1β), resistin, high mobility group box 1 (HMGB1), and forkhead box protein 3 (FOXP3). Cartilage acidic protein 1 (CRTAC1) was not significantly upregulated. Two proteins were significantly downregulated including scavenger receptor class F member 2 (SCARF2) and P-selectin. All proteins selected are mainly associated with inflammation, cell proliferation/differentiation, apoptosis and cell–cell or cell–matrix interaction.
Conclusion
Proteomic analysis of CRSsNP and control mucus has confirmed known and revealed novel disease-associated proteins that could potentially serve as a new biosignature for CRSsNP. Analysis of the associated pathways will specify endotypes of CRSsNP and will lead to an improved understanding of the pathophysiology of CRSsNP. Furthermore, our data contribute to the development of a reproducible, non-invasive, and quantitative “liquid biopsy” for rhinosinusitis.
Keywords
Introduction
Chronic rhinosinusitis without nasal polyps (CRSsNP) as a subtype of CRS represents a highly complex condition which is mainly characterized by chronic inflammation of the nasal mucosa and fibrotic nasal tissue, while nasal polyps and edema are absent. 1 CRS has been shown to have a significant impact on quality of life, greater in some respects than other chronic diseases such as angina or chronic obstructive pulmonary disease (COPD). 2 Despite the high personal and socioeconomical burden, the etiopathology of the disease is still unknown. CRSsNP and chronic rhinosinusitis with polyps (CRSwNP) are different subtypes of CRS with different endotypes and phenotypes. CRSwNP is associated with a type 2 inflammation, whereas CRSsNP is generally associated with a type 1 or type 3 inflammation. Thus, these subtypes show different pathophysiological features.3–5 Although there are various hypotheses on the pathophysiology of CRSsNP, a generally accepted approach to explain the mechanisms behind the disease remains elusive.1,6 Therefore, endotype classification based on thorough investigation of the underlying pathophysiology is an important task. In complex diseases with mixed pathophysiological mechanisms, a phenotype-driven treatment may not always be sufficient to obtain optimal control. About 25% of CRSsNP patients do not improve with current treatment options, and to date an effective long-term treatment that prevents the recurrence of the disease has not yet been identified. 7 Consequently, the development of optimized diagnostic criteria that can provide relevant prognostic information and improved treatment algorithms will be necessary. Although there are several studies that have demonstrated an array of altered protein expression levels in the mucosa of CRSsNP patients relative to control mucosa, mostly invasive surgical tissue sampling has been required. Moreover, these previous studies have often been restricted by the limited range of proteins tested.4,8–14
Conversely, large-scale analysis of the CRSsNP proteome allows previously unknown disease factors to be revealed, more insight into the pathophysiology to be gained, and novel potential therapeutic targets to be identified. The use of nasal mucus provides an easy, non-invasive, and painless way of biosignature sampling. Additionally, this “liquid biopsy” may be prospectively and repetitively sampled and may be used for therapeutic monitoring.
Therefore, the objective of this study was (1) to identify proteins that are differentially expressed between CRSsNP and controls in nasal mucus using a multiplexed proteomic approach, and (2) to validate these proteins as potential novel non-invasive biomarkers.
Materials and Methods
Patients
This study was approved by the institutional review board of the University of Erlangen-Nürnberg (No: 4_20; No: 269_17) and all patients provided written informed consent.
All samples were taken from patients who had not been exposed to antibiotics or any topical/systemic steroids for at least 4 weeks. A 4-week washout period is frequently used as a standard deprivation time and has also been used in previous studies of our group. 15
Inclusion criteria included patients diagnosed with CRSsNP by the International Consensus Statement on Allergy and Rhinology: Rhinosinusitis (ICAR:RS) 6 criteria and healthy patients (ie, controls). Some of these patients underwent surgery for either CRSsNP (functional endoscopic sinus surgery, FESS, septoplasty and turbinate reduction if needed) or blockage of the nose (controls: septorhinoplasty and/or turbinate reduction). Exclusion criteria included ciliary dysfunction, autoimmune disease, cystic fibrosis, immunodeficiency, fungal sinusitis, malignancies, gastroesophageal reflux disease, chronic rheumatic disease, or any other disease requiring long-term corticosteroid therapy. Patients who had previous sinus surgery were also excluded.
Mucus Collection Technique
Mucus samples were taken from all patients before antibiotic or steroid administration by applying a compressed polyvinyl alcohol sponge (Medtronic, Minneapolis, MN) to the nasal cavity adjacent to the middle turbinate for 5 min, taking care not to abrade the mucosa or contaminate the sponge with blood. For those patients undergoing surgery, tissue was also collected from the mucosa of the middle meatus using Blakesley forceps.
Mucus Extraction
To obtain mucus samples, liquid was extracted from the sponges by adding 400 µL phosphate-buffered saline (pH = 8) with Complete™ protease inhibitor cocktail (Roche Diagnostics GmbH, Germany). After 5 min the sponges were centrifuged at 16 000g for 5 min at 4 °C. Afterwards this procedure was repeated once. Total protein was quantified using a Micro bicinchoninic acid (BCA) Protein Assay Kit (Thermo Fisher Scientific, Bonn, Germany). All samples were aliquoted and stored at −80 °C. This technique was established by our group before. 15
Proteomic Array From Nasal Mucus
Proteomic analysis of 14 mucus samples (n = 7 per group) was performed using the RayBio® L-Series Human Antibody Array 2000 Glass Slide Kit (#AAH-BLG-2000-8, Raybiotech, Peachtree Corners, GA, USA), following the recommended protocol of the manufacturer. For the incubation step, samples were run using 170 µg of total protein each. Extraction of the data by fluorescence was conducted by Raybiotech. Median normalization of the data was performed using the analysis tool provided by Raybiotech. Following this, the values of all proteins that were already known to serve as potential reference genes and against which antibodies were present on the array were examined using the geNorm application. Out of the proteins that showed the most constant expression 5 proteins including glyceraldehyde 3-phosphate dehydrogenase (GAPDH), hypoxanthin-guanin phosphoribosyltransferase (HPRT), phosphoglycerate kinase 1 (PGK1), vascular endothelial growth factor B (VEGF-B), and transaldolase 1 (TALDO1) were chosen for additional normalization of all values. Background signals were then lowered by subtraction of the negative control per subarray from all the other values of the same subarray. The fold change (FC) of the means was obtained by dividing the mean of the CRSsNP samples by the mean of the controls. The FC of the median values was obtained by dividing the median of the CRSsNP samples by the median of the controls.
Validation of the Mucus Proteome by Western Blot
For the validation of resistin, HMGB1, FOXP3, CRTAC1, SCARF2, and P-selectin western blots were performed using the same independent group of CRSsNP and control (n = 7 per group) (cohort A) mucus samples that were used for the array. Additional western blots were performed using a second validation cohort of CRSsNP (n = 18), and control (n = 16) (from cohort B) mucus samples. Due to the lack of evaluable signals from the MIP-1β antibodies, there is no western blot for this protein. Twenty micrograms (20 μg) of mucus protein was used for each lane. After denaturation at 95 °C with denaturing buffer for 5 min, samples were applied on 8% to 15% SDS-PAGE and transferred to nitrocellulose membranes (ROTI®NC; Carl Roth, Karlsruhe, Germany). For resistin, samples were applied without denaturation by heat and by using loading buffer without mercaptoethanol. Results were normalized to GAPDH, because this was the reference protein that showed one of the most regular expressions out of all reference proteins on the antibody array examined by the geNorm application. Values exceeding the mean value plus 3 times the standard deviation were identified as outliers and excluded. The primary detection antibodies listed in Table 1 were incubated overnight, respectively, followed by the secondary antibodies, peroxidase-labeled anti-mouse/rabbit immunoglobulin G (IgG) antibody (Thermo Fisher Scientific, Bonn, Germany). The blots were incubated with either SuperSignal West Dura Extended Duration Substrate or SuperSignal West Femto Maximum Sensitivity Substrate (Thermo Fisher Scientific, Bonn, Germany) and the signals imaged using ChemStudio PLUS (Analytik Jena, Jena, Germany). Quantification of band intensity was performed using VisionWorks version 8.2 (Analytik Jena, Jena, Germany).
Primary Antibodies That Were Used for Western Blots and IHC.
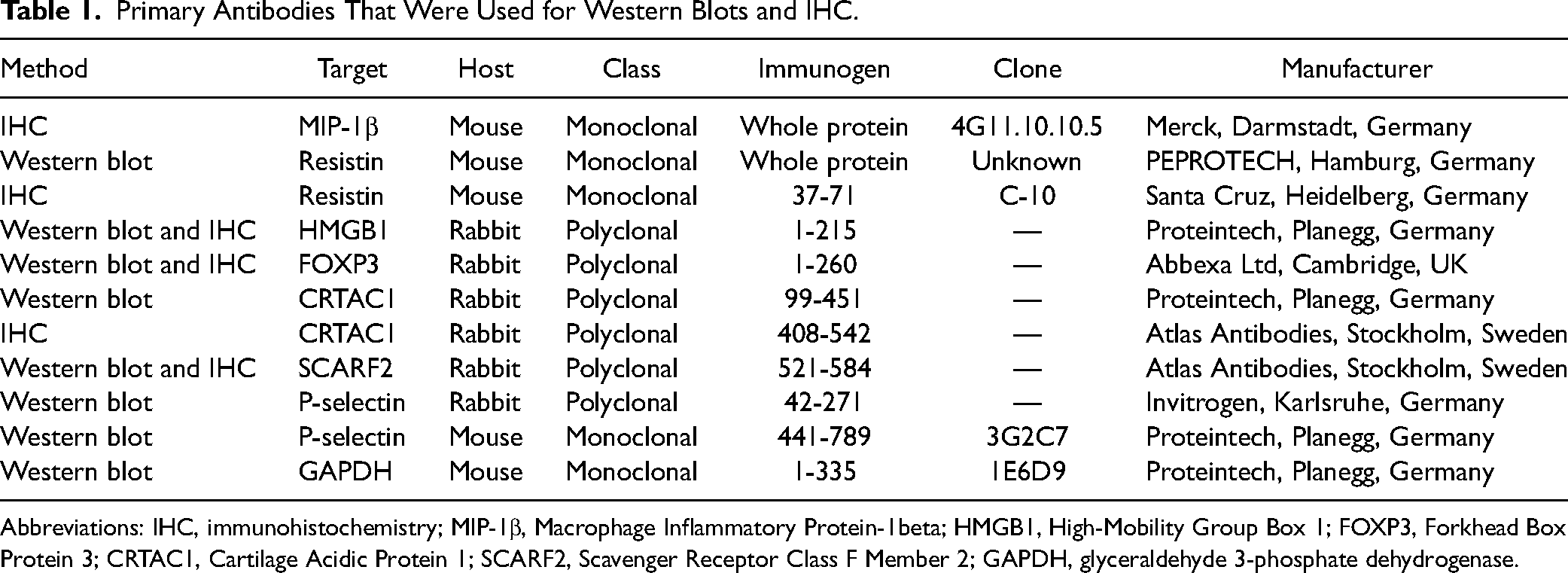
Abbreviations: IHC, immunohistochemistry; MIP-1β, Macrophage Inflammatory Protein-1beta; HMGB1, High-Mobility Group Box 1; FOXP3, Forkhead Box Protein 3; CRTAC1, Cartilage Acidic Protein 1; SCARF2, Scavenger Receptor Class F Member 2; GAPDH, glyceraldehyde 3-phosphate dehydrogenase.
Validation of the Selected Mucus Proteomic Results by Enzyme-Linked Immunosorbent Assay
Enzyme-linked immunosorbent assay (ELISA) tests were conducted for MIP-1β and P-selectin. For each ELISA nasal mucus samples of CRSsNP and control (n = 30 per group) (from cohort A and cohort B) were used. After thawing the aliquots MIP-1β and P-selectin were quantified in the nasal mucus using ELISAs that were performed according to the manufacturer’s protocols. For MIP-1β the nasal mucus was diluted 1:5 with Assay Dilution Buffer (included in the kit), and the MIP-1β Human ProQuantum Immunoassay Kit No. A355975 (Thermo Fisher Scientific, Germany) was used. For P-selectin, the nasal mucus was diluted 1:2 with Assay Diluent (included in the kit), and the RayBio Human P-selectin IQELISA No. IQH-PSELECTIN (Raybiotech, Peachtree Corners, GA, USA) was used. Standard curves were generated and results were calculated by normalizing the values of the total protein concentration of the mucus samples (BCA assay, Thermo Fisher Scientific, Bonn, Germany). Values below the detection limit were excluded. Values exceeding the mean value plus 3 times the standard deviation were identified as outliers and excluded.
Tissue Analysis by Immunohistochemistry
Immunohistochemistry was performed to localize the proteins of interest in the surgically collected tissue. Immunohistochemistry (IHC) on paraffin-embedded sections was carried out with ZytoChem-Plus AP Polymer-Kit (Zytomed Systems GmbH, Berlin, Germany). To make the epitopes available for antibody binding, the sections underwent deparaffinization and heat-mediated antigen retrieval using either phosphate citrate buffer or Tris-EDTA buffer at 95 °C for 20 min. Additional reduction in background staining was achieved by covering the sections with BLOXALL™ Endogenous Peroxidase and Alkaline Phosphatase Blocking Solution (Vector Laboratories, Inc.) for 10 min before the protein block with blocking solution (included in the kit). The antibodies listed in Table 1 were incubated over night at 4 °C. A nonspecific antibody (Cell Signaling Technology, Inc., Danvers, MA) served as negative control. Afterwards, the ZytoChem-Plus AP reagent was applied. Antigens were stained with SIGMAFAST™ Fast Red TR/Naphthol AS-MX tablets. Counterstaining was performed with Harris’ hematoxylin solution (ORSAtec GmbH, Bobingen, Germany). The sections were covered with Aquatex (Merck, Darmstadt, Germany).
Statistical Analysis
The results of the semiquantitative analysis of western blots and ELISA as well as the results of the demographic data using the Mann–Whitney U-test were calculated using Prism (Graphpad Prism 9.2.0; GraphPad Software, La Jolla, CA). A false discovery rate cutoff of P < .05 was used to detect significant differences between the CRSsNP and control groups.
Results
Demographics
Demographic data is shown in Table 2. There were no significant differences between the CRSsNP and control groups (cohort A: n = 7 per group; cohort B: CRSsNP n = 28, and control n = 39) with respect to gender, race, asthma, AERD, or allergic rhinitis for both cohorts. In cohort B, the count of smokers was significantly lower (P = .0013) and the age was significantly higher (P < .0001) in CRSsNP patients than in control patients. No patient was diagnosed with acute fungal rhinosinusitis, and no tissue isolate revealed tissue allergic mucin.
Demographic data.
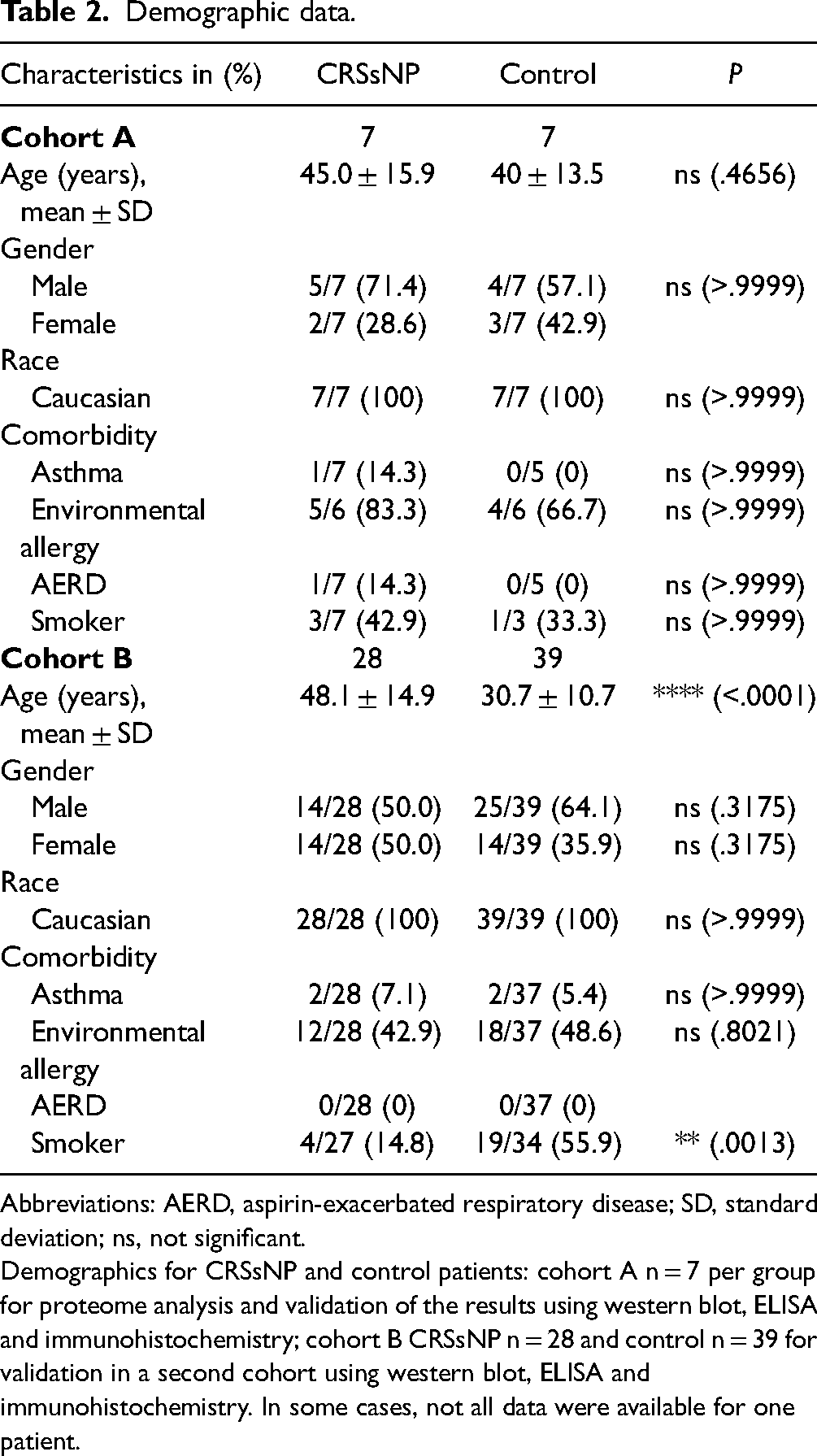
Abbreviations: AERD, aspirin-exacerbated respiratory disease; SD, standard deviation; ns, not significant.
Demographics for CRSsNP and control patients: cohort A n = 7 per group for proteome analysis and validation of the results using western blot, ELISA and immunohistochemistry; cohort B CRSsNP n = 28 and control n = 39 for validation in a second cohort using western blot, ELISA and immunohistochemistry. In some cases, not all data were available for one patient.
Mucus Proteomic Results
Proteomic results of all validated proteins are shown in Table 3 (see Supplemental data 1 for results of all evaluable proteins of proteomic analysis). Out of 2000 proteins examined on the antibody array we chose 7 proteins out of the most differentially expressed proteins for further validation. The decision as to which proteins were worth a more detailed analysis depended on 2 factors: their FC and their previous association with general inflammation in the literature. Four out of these 7 proteins were shown to be overexpressed in CRSsNP mucus compared to the control, including MIP-1β, resistin, HMGB1, and CRTAC1 according to this antibody array analysis. Three out of these 7 proteins were shown to be underexpressed in CRSsNP mucus compared to the control including FOXP3, SCARF2, and P-selectin.
Results of Validated Proteins of Proteomic Array.

Abbreviations: CRSsNP, chronic rhinosinusitis without nasal polyps; MIP-1β, macrophage inflammatory protein-1beta; HMGB1, High-Mobility Group Box 1; CRTAC1, Cartilage Acidic Protein 1; FOXP3, Forkhead Box Protein 3; SCARF2, Scavenger Receptor Class F Member 2; MV, mean value; FC, fold change; SD, standard deviation.
Most of the proteins found to be upregulated in CRSsNP mucus compared to the controls are known for their main functions, that is to contribute to inflammation and cell proliferation/differentiation and apoptosis including MIP-1β, resistin, and HMGB1.16–19 In contrast, SCARF2 was downregulated, although according to current knowledge it is associated with the same functions.20,21 FOXP3, a transcription factor of regulatory T cells (Tregs), 22 was also found to be downregulated.
CRTAC1 is an extracellular matrix (ECM) protein, about which not much is known so far but which is suspected to play a role in cell–cell or cell–matrix interactions. 23 It was shown to be upregulated in CRSsNP mucus according to this initial analysis.
P-selectin is a protein particularly present on platelets and endothelial cells that mediates diapedesis 24 and whose proteomic array result revealed an underexpression in CRSsNP mucus.
Overall, we considered these 7 proteins to be interesting candidates for the following verification of the first results.
Validation of the Mucus Proteomic Results by Western Blots
We performed western blots using matched mucus samples from the proteomic array (cohort A) and an independent set of samples (cohort B). In total, n = 25 CRSsNP and n = 23 control mucus samples were analyzed for HMGB1, FOXP3, CRTAC1, SCARF2, and P-selectin. For resistin n = 25 CRSsNP and n = 26 control mucus samples were analyzed. Interestingly, for HMGB1, several bands at different molecular weights were seen. Directly above the FOXP3 band, the lower part of a non-specific band could be seen. This band was present in all western blots with mucus samples, even after the sole incubation of secondary antibodies without primary antibody. Unfortunately, the location of this band was just above the FOXP3 signal.
Judging by the band pattern, there was hardly any difference in FOXP3 presence, but normalization revealed that, in contrast to the array result, it was overexpressed in CRSsNP mucus. Results showed a significant overexpression of resistin (2 outliers) (FC = 3.03, P = .0062), HMGB1 (2 outliers) (FC = 1.75, P = .0465), and FOXP3 (1 outlier) (FC = 1.42, P = .0388) and a non-significant overexpression of CRTAC1 (1 outlier) (FC = 1.47, P = .1229). Results also showed a significant underexpression of SCARF2 (2 outliers) (FC = 0.46, P = .0002) and P-selectin (2 outliers) (FC = 0.37, P = .0003) in CRSsNP mucus compared to control mucus (Figures 1 and 2) (see Supplemental data 2 for all results including outliers of ELISA and western blot analysis).
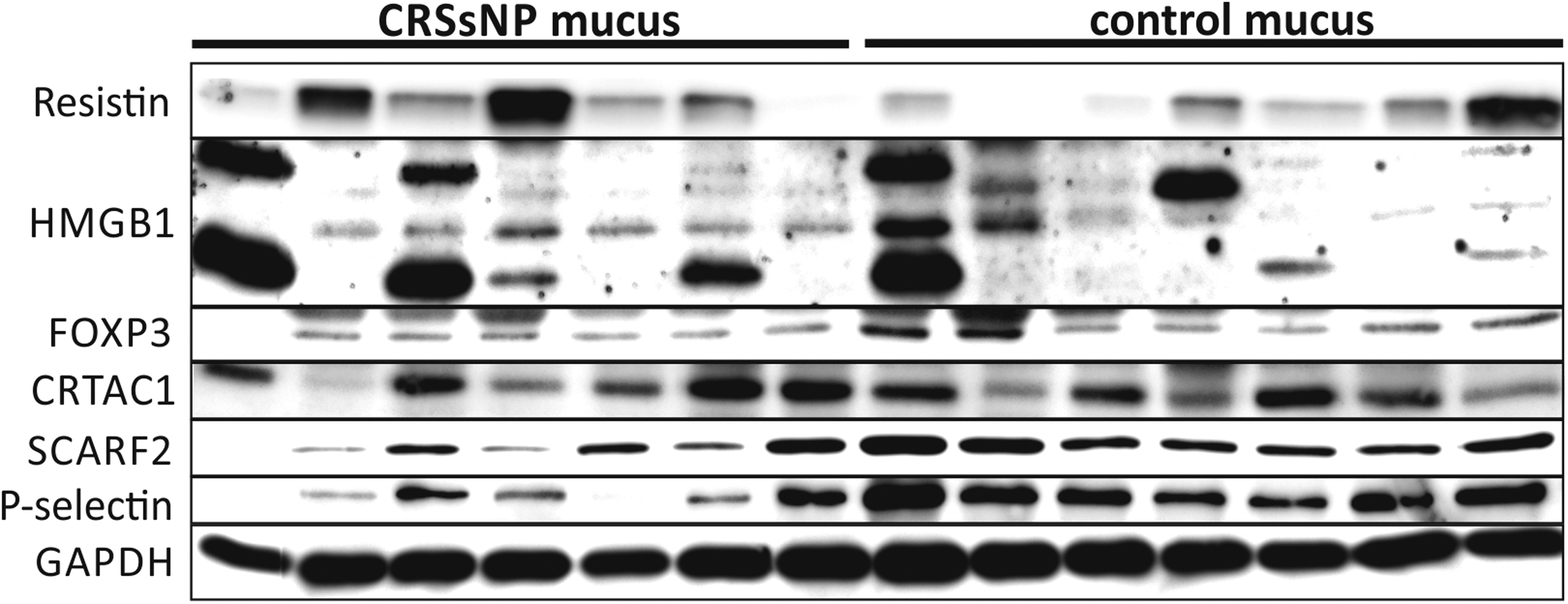
Subset of western blots of CRSsNP and control patients (n = 7 per group) for resistin, HMGB1, FOXP3, CRTAC1, SCARF2, and P-selectin as well as for the reference protein GAPDH in nasal mucus. Since a non-reducing sample buffer was used to detect resistin, the result was normalized to GAPDH signals of the same blot (data not shown).
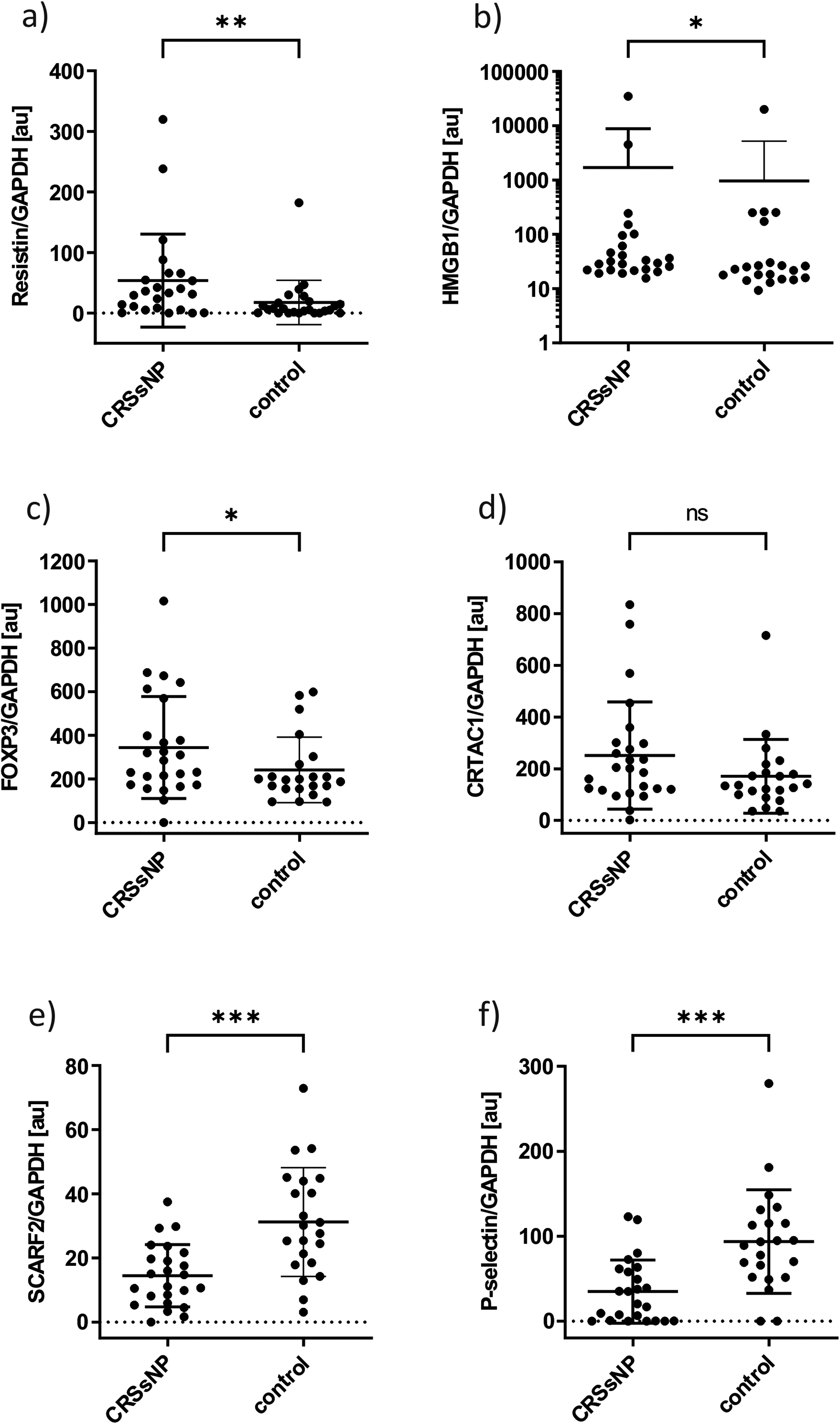
Scatter dot plot for semiquantitative analysis by western blots in nasal mucus for CRSsNP and control patients normalized to GAPDH. Outliers are not included. Due to the high standard deviation, the y-axes are plotted logarithmically for HMGB1. The error bars pointing down are not displayed in this representation. A statistically significant difference between CRSsNP and control patients can be seen for (a) resistin (P < .01), (b) HMGB1 (P < .05), (c) FOXP3 (P < .05), (e) SCARF2 (P < .001), and (f) P-selectin (P < .001). For (d), CRTAC1 no significant difference between CRSsNP and control patients could be found (P = .1229). *<.05, **<.01, ***<.001.
Validation of Mucus Proteomic Results by ELISA
For validation of MIP-1β and additional validation of P-selectin, we performed ELISA from the CRSsNP and control mucus of cohort A and cohort B (n = 30 per group in total).
Results showed a significant overexpression of MIP-1β (1 outlier, 19 values below detection limit) (FC = 3.61, P = .0022) and a significant underexpression of P-selectin (2 outliers) (FC = 0.005, P = .0083) in CRSsNP mucus compared to control mucus (Figure 3) (see Supplemental data 2 for all results including outliers of ELISA and western blot analysis).
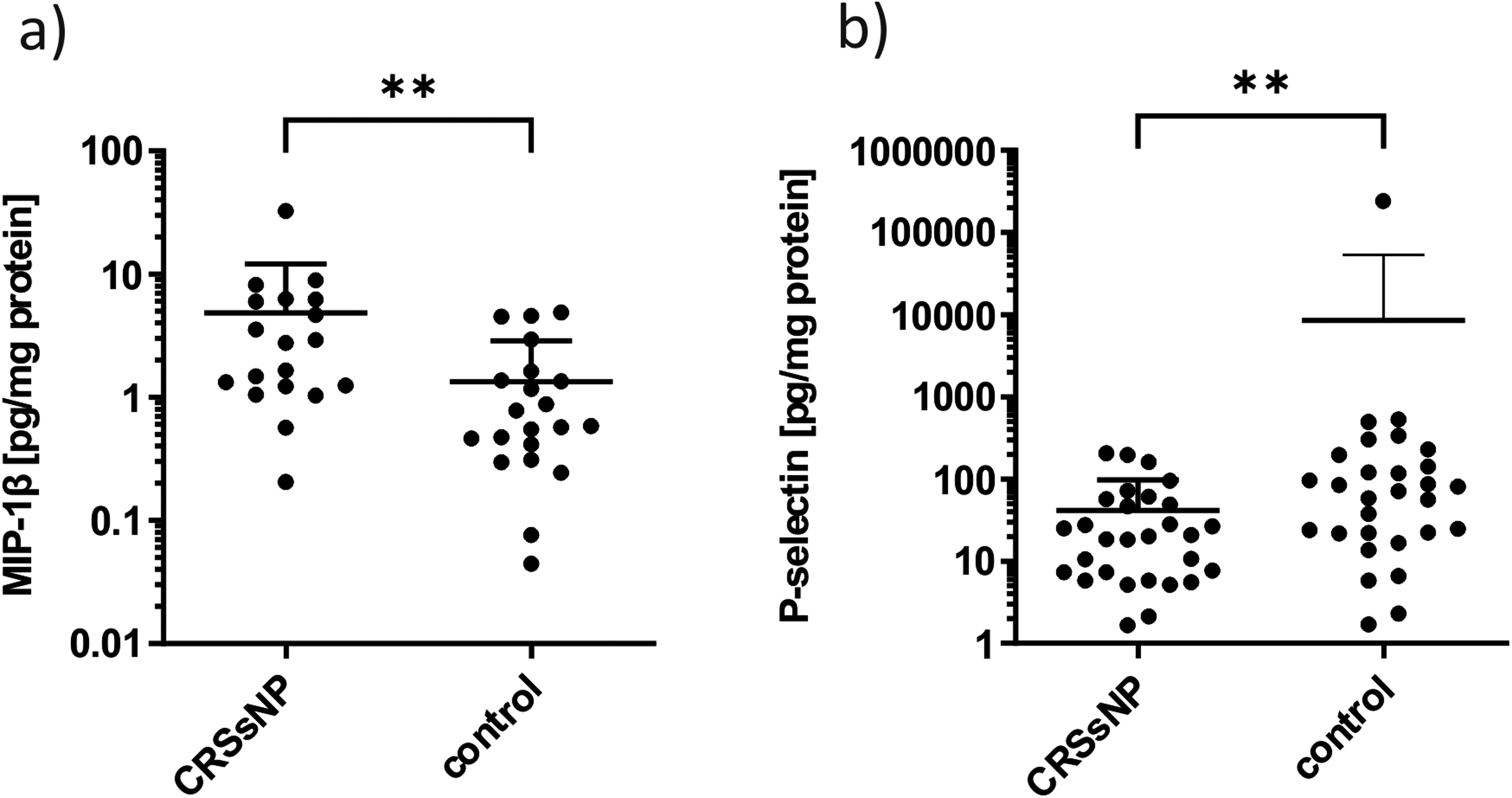
Scatter dot plot for quantitative analysis by ELISA in nasal mucus for CRSsNP and control patients. Outliers are not included. Due to the high standard deviations, the y-axes are plotted logarithmically. The error bars pointing down are not displayed in this representation. A statistically significant difference between CRSsNP and control patients can be seen for (a) MIP-1β (P < .01) and (b) P-selectin (P < .01). **<.01.
Location of the Selected Proteins in the Tissue Using IHC
Immunohistochemical staining was performed for all proteins in CRSsNP ethmoid tissue and control inferior turbinate tissue. For HMGB1 and P-selectin there were only minor visible differences in protein localization and quantitative distribution between CRSsNP and control tissues. MIP-1β was located in the respiratory, ciliated epithelium but also in some immune cells of the stroma, while control tissue revealed a lesser extent of staining. Resistin in CRSsNP tissue was strongly present within the whole epithelial layer, whereas control tissue revealed only a slight staining of the basal epithelial layer and also the glandular epithelium. HMGB1 staining was generally weak, but the protein appeared to be expressed by most cells throughout the whole tissue. FOXP3 was mainly located in the basal and apical epithelial layer. It was also present the glandular epithelium of the control tissue and in immune cells within the stroma of the CRSsNP tissue. CRTAC1 staining revealed some spots within the stroma and the epithelial cells that were differently shaped, whereas the control tissue contained fewer of them, and staining of the basal epithelial layer only in control tissue. The localization of SCARF2 was in the epithelial cells and mainly in the leftover mucus on the cilia, whereas hardly any staining was seen in CRSsNP tissue. P-selectin was expressed by the epithelium, mainly in the apical layer (Figure 4).

Immunohistochemistry of MIP-1β (A,I), resistin (B,J), HMGB1 (C,K), FOXP3 (D,L), CRTAC1 (E,M), SCARF2 (F,N), and P-selectin (G,O) showing differential localizations and staining intensities in CRSsNP and control patients. The NC is displayed in (H) and (P). Antigens were stained with red alkaline phosphatase-substrate, and counterstaining was performed with Harris’ hematoxylin solution.
Discussion
Recent consensus statements on the diagnosis of CRSsNP focus on symptoms, endoscopy, and imaging-based criteria, 6 or on an anatomical and endotype-based (type2, non-type2) approach for primary CRS. 1 In both consensus statements, the underlying pathophysiologic features of CRSsNP are not fully elucidated. 1 Understanding the etiopathology of CRSsNP as well as developing an effective treatment remains challenging, although recent advantages have been made.25–30 Current therapies are based on patient phenotype, and subtypes based on significant underlying immunologic differences are not considered.31,32 Accordingly, these patients’ clinical responses on medical therapy can be highly variable33–35 and relapse surgery is often required.7,36 Hence, there is an urgent need for endotyping in a more precise way, which is especially necessary for the non-type2 group. Simultaneously, a non-invasive combination of biomarkers is necessary to study disease over time and allow more information to be gained about the pathophysiology of CRS subtypes. Therefore, the aim of this study was to identify and validate a promising combination of non-invasive mucus biomarkers to solve this shortcoming.
In order to identify proteins that are differentially expressed between CRSsNP and control mucus we first conducted an antibody array capable of detecting 2000 different proteins at once. After evaluating these results, we picked 7 proteins for further validation by western blot and ELISA in a second step. Proteins that were significantly increased in CRPsNP mucus compared to the controls were MIP-1β, resistin, HMGB1, and FOXP3. The presence of CRTAC1 was also higher in CRSsNP mucus, but the result was not significant. SCARF2 and P-selectin were significantly decreased.
A closer look at the properties of these proteins reveals that their main functions contribute either to inflammation, to angiogenesis, to cell–cell/cell–matrix interactions or to cell proliferation, differentiation and apoptosis in either way.
MIP-1β is a chemokine that participates in host response to invading pathogens. It is produced by a variety of inflammatory cells as well as epithelial and endothelial cells induced by certain stimuli.16,37–39 There are already several other studies that describe an upregulation of MIP-1β in the nasal mucus of chronic sinusitis.11,40
Resistin is a small hormone-like adipokine. Once thought to be the cause of diabetes, it is now linked to many serious medical conditions. Via nuclear factor-κB (NF-kB), phosphatidylinositol 3-kinase (PI3K), and mitogen-activated protein kinases (MAPK) mediated pathways it induces immune cell activity and upregulation of adhesion molecules and growth factors that promote endothelial dysfunction, cell proliferation, and angiogenesis. Induction of oxidative stress in the tissue by resistin leads to a vicious circle that inhibits the process of healing.17,18 In connection with CRSsNP, the development of reactive oxygen species and reactive nitrogen species is often reported, which causes considerable damage to the tissue and contributes to the chronification of the inflammation.1,41 By upregulating mucin genes resistin causes mucus hypersecretion in airway epithelial cells, which is a well-known side effect of various respiratory diseases. 42 Resistin was previously found to be overexpressed in the tissue of non-eosinophilic CRSwNP (NECRSwNP). 43
HMGB1 can be present anywhere inside and outside of all eucaryotic cells dependent on its posttranslational modifications (PTM). Intracellularly bound to chromatin, it participates in the transcription, replication, and repair of DNA. Released either actively by cells or passively by necrosis, it can conduct many different functions by binding to other proteins and different receptors. These receptors work mainly through NF-kB and MAPK pathways which generate pro-inflammatory mediators and enhance proliferation and differentiation of the target cells. 19 By producing defense factors, the airway epithelial cells together with the mucin layer represent the first barrier against invading pathogens.3,8,27,44 However, in case they manage to overcome this barrier, inflammation often leads to necrosis. It is therefore probable that HMGB1 is also increasingly produced by airway epithelium as a first defense factor or released by death cells. Our results are consistent with previous studies that describe HMGB1 as upregulated in the CRS nasal fluid and tissue of patients suffering from different types of CRS.45,46
FOXP3 is a transcription factor that regulates development and function of the Tregs. Presenting a subset of T cells, Tregs are responsible for protection against an excessive immune response. Activity of FOXP3 depends on complex formation with other proteins and numerous PTM. 22 Our array analysis showed a downregulation of FOXP3 in CRSsNP mucus compared to controls, which later turned out to be the other way around by the subsequent validation where more mucus samples were included and outliers were excluded. The fact, that CRSsNP goes along with higher presence of FOXP3 and Treg cells in contrast to CRSwNP, where there is a deficit of Treg cells and FOXP3 in tissue, is interesting and justifies further investigation of T-cell responses in CRS.47,48
SCARF2 is a member of the family of scavenger receptors (SRs) that are membrane-bound multi-domain proteins expressed by a variety of cells. SRs are separated into 8 classes that are distinguished by the expression of characteristic protein domains. 20 In contrast to other scavenger receptors only little is known about SCARF2. The structural determinants of SCARF2 are a long cytosolic tail and several epidermal growth factor (EGF) and EGF-like domains which suggest that it is primarily involved in cell proliferation and differentation.20,21 Generally, SRs are considered to be a subclass of the pattern recognition receptors (PRRs) playing an important role in innate immune response and thus in inflammation. Overall, scavenger activity was originally associated with the ability to bind and to internalize modified lipids and proteins including oxidized low-density lipoprotein. Moreover, it is now known that SRs carry out a wide range of functions and that they are able to recognize diverse ligands including danger-associated molecular patterns (DAMPs) or pathogen-associated molecular patterns (PAMPs). Binding of these ligands while partnering with other receptors including toll like receptors (TLRs) allows them to induce many different signaling cascades depending on the ligand and the cellular context. This adaptive behavior renders SRs extremely versatile. In general, the complex events in which they are involved include phagocytosis, antigen presentation, the clearance of apoptotic cells, the transport of debris and cargo within the cell, lipid transport, and taste sensation. Due to their multiple functions, SRs can also promote the pathogenesis of many neurodegenerative and metobolic diseases and even cancer.20,49 However, there are different reports about whether SCARF2 exerts any scavenger activity 50 or not. 21 Hypothesizing why SCARF2 is downregulated in CRSsNP mucus is difficult. One possible explanation lies in the regulation process of PRRs. In disease state PRRs are often upregulated because of the increased immune response needed. 51 Therefore, in CRS one would actually expect an increased expression of the receptor at the cell membrane. Here it is important to consider alterations in the shedding process of the receptor. Shedding of SRs by exofacial proteases results in the release of soluble products to the circulation, whose functions can differ from those of the precursor protein.20,21 Thus, a possible explanation for SCARF2 underexpression in CRSsNP mucus could be either an unknown function of its soluble ektodomain or an alteration in its shedding process driven by impaired protease activity.
CRTAC1 is an ECM protein mainly synthesized by chondrocytes but also by airway club cells, and alveolar type2 epithelial cells. 52 Its function still remains elusive although the protein's structure and the presence of certain amino-acid sequence patterns such as an EGF-like motif leads to the assumption that it plays a role in cell–cell or cell–matrix interactions.23,53 There is evidence that the spliced variant CRTAC1-A alters genes responsible for cell proliferation, cell migration, and ECM regeneration in human dermal fibroblasts in vitro. In addition, the protein increases the fibroblasts’ viability and energy metabolism and thus contributes to wound healing and tissue homeostasis. 54 Despite a non-significant result, mucus analysis indicated an elevated presence of this protein in CRSsNP mucus compared to control mucus. One reason for the upregulation in nasal mucus could be an increased activity of ECM proteinases that excessively shed ECM proteins. Tissue remodeling along with fibrosis is a concomitant issue in CRSsNP.9,55 In this process components of the ECM are decomposed by shedding to enable cell movement and reconstruction. 56 Recent discoveries about an altered presence of ECM components, tissue remodeling proteins, and adhesion molecules in different forms of CRS underline our result.9,10,57,58
P-selectin is known to be expressed mainly by activated platelets and endothelial cells. It belongs to the family of selectins, a group of adhesion molecules that play an essential role in immunity, inflammatory responses, and tissue healing. The most prominent functions of P-selectin are the stimulation of platelet aggregation and the regulation of the initial interactions between blood vessels, activated platelets, and leucocytes. Binding of P-selectin to its major ligand P-selectin Glycoprotein Ligand 1 (PSGL-1) expressed mainly by leucocytes allows formation of non-covalent interactions. P-selectin permits leucocyte recruitment to the side of inflammation by interaction with immobilized chemokines that convert b2 integrins to high-affinity conformations. As a consequence, rolling velocities of the leucocytes are decreased up to their arrest, followed by their diapedesis. P-selectin is stored inside the cells and translocated to the cell membran after certain stimuli. 24 Although P-selectin reportedly appears to be also expressed by several other cell types, there is only little information about its role in those cells so far.
Since P-selectin is almost exclusively associated with tethering of leukocytes to platelets and vascular endothelial cells, the proteins other functions remain elusive. For instance, P-selectin was shown to be also expressed in pulmonary artery smooth muscle cells (PASMCs) in mice, where it is upregulated by hypoxia in a HIF-1a (hypoxia-induced factor-1a)-dependent manner. Induction of different signaling cascades led to PASMC proliferation and vascular remodeling and thus to pulmonary artery hypertension. 59 Another study noted that P-selectin can be found in human corneal epithelium. Here, it might facilitate the trafficking and accumulation of immune cells on the ocular surface providing local protection from pathogens. 60 To the best of our knowledge, the detection of P-selectin in the nasal epithelium has not been reported previously. Once again, we can only speculate about the reason for the presence of P-selectin in nasal epithelium which apparently releases the protein into the nasal mucus and why it is less present in CRSsNP mucus compared to the control. One possible reason could again be an impaired shedding process of the receptor. Interestingly, after binding to P-selectin whose membrane-bound form is a dimer, leukocytes induce a shedding process by which P-selectin is released into the circulation as a soluble monomer. 61 Since no major differences were seen in the IHC staining between CRSsNP and control tissues, a similar amount of P-selectin production in the epithelium of both tissues may be assumed. However, eventually P-selectin in CRSsNP tissue is bound less by leukocytes and other immune cells, which produces less of the soluble form.
Altogether, our findings of MIP-1β, HMGB1, and FOXP3 upregulation in CRSsNP mucus are consistent with current data in the literature underlining these results. Resistin has been described before as being upregulated in NECRSwNP before. 43 This finding in different subclassifications of CRS is interesting and justifies further analysis. Finally, SCARF2 and P-selectin resemble novel markers for CRSsNP, which therefore also justifies further analysis. We chose this combination of biomarkers in order to characterize the CRSsNP proteome more precisely and try to endotype in more detail. This is important as CRSsNP currently tends to fall into the “non-type2” group rather than being specified with a distinct sub-/endotype. By using a highly multiplexed approach to the CRSsNP proteome, the most promising protein biomarkers could be identified from a large set. To the best of our knowledge, this has been one of the largest multiplexed proteomic analysis of nasal mucus sampled from the middle meatus of CRSsNP and control patients. By validating these results with additional samples, we have laid the foundation stone for further analysis of the biomarkers and CRS pathophysiology.
Despite promising results, there were limitations to our study. First, the analysis of nasal mucus is challenging due to its content of cellular debris and degraded protein fragments, which renders it a highly complex and heterogeneous fluid.62,63 This has probably caused the high standard deviations between the samples and masked additional findings.
Conclusion
A promising combination of known and novel non-invasive mucus protein biomarkers for CRSsNP was identified using the combination of a highly multiplexed proteomic approach, western blots, and ELISAs. All proteins are associated with inflammation, angiogenesis, cell–cell/cell–matrix interactions or cell proliferation, differentiation, and apoptosis. By analyzing these proteins and pathways further, the definition of detailed and precise endotypes of CRSsNP may be possible. Additionally, our data are the basics for the development of a non-invasive “liquid biopsy” for CRSsNP that allows the disease pathophysiology to be studied, the disease state to be discriminated, novel therapeutic targets to be potentially revealed and therapies prospectively monitored.
Supplemental Material
sj-docx-1-ajr-10.1177_19458924221136651 - Supplemental material for Analysis of CRSsNP Proteome Using a Highly Multiplexed Approach in Nasal Mucus
Supplemental material, sj-docx-1-ajr-10.1177_19458924221136651 for Analysis of CRSsNP Proteome Using a Highly Multiplexed Approach in Nasal Mucus by Vanessa-Vivien Pesold, Olaf Wendler, Lisa Morgenthaler, Franziska Gröhn and Sarina K. Mueller in American Journal of Rhinology & Allergy
Supplemental Material
sj-docx-2-ajr-10.1177_19458924221136651 - Supplemental material for Analysis of CRSsNP Proteome Using a Highly Multiplexed Approach in Nasal Mucus
Supplemental material, sj-docx-2-ajr-10.1177_19458924221136651 for Analysis of CRSsNP Proteome Using a Highly Multiplexed Approach in Nasal Mucus by Vanessa-Vivien Pesold, Olaf Wendler, Lisa Morgenthaler, Franziska Gröhn and Sarina K. Mueller in American Journal of Rhinology & Allergy
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Else Kröner-Fresenius-Stiftung.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.