
Abstract
Background
Chronic rhinosinusitis (CRS) comprises heterogeneous eosinophilic (type-2) and neutrophilic (type-1/3) endotypes, yet existing murine models rarely distinguish them or compare their therapeutic responsiveness.
Methods
Seven-week-old female C57BL/6N mice received intranasal instillations three times weekly for 4 or 12 weeks with either (1) a clinically relevant airborne allergen cocktail (house dust mite [HDM], Aspergillus fumigatus, Alternaria alternata, and Staphylococcus aureus protease) to induce eosinophilic CRS (ECRS) or (2) an innate stimulus mixture (lipopolysaccharide, β-1,3-glucan, and S. aureus protease) to induce neutrophilic CRS (NCRS). A parallel cohort received weekly dexamethasone administration (2 mg/kg). Endpoints included flow cytometry, enzyme-linked immunosorbent assays, quantitative PCR, histology, and immunofluorescence assays.
Results
Four-week allergen exposure produced a pure ECRS phenotype characterized by robust tissue eosinophilia, type-2 cytokines, and marked responsiveness to dexamethasone. Extending allergen delivery to 12 weeks generated mixed CRS with superimposed neutrophilia, upregulation of Il1b/Tgfb1, mucus hypersecretion, and attenuated steroid efficacy. Innate stimulus treatment yielded a steroid-resistant NCRS phenotype dominated by neutrophils, elevated Ifng/Il17a expression, and minimal type-2 biomarkers. Systemic eotaxin and HDM-specific immunoglobulin E mirrored those found in local eosinophilia, whereas circulating granulocytes remained unchanged across groups.
Conclusions
By adjusting the stimulus type and duration, we established tunable murine models that recapitulate pure ECRS, mixed CRS, and steroid-resistant NCRS within a single genetic background. These paired models provide a versatile platform for dissecting endotype-specific mechanisms and evaluating tailored interventions, highlighting the potential importance of early, phenotype-directed CRS therapy.
Introduction
Chronic rhinosinusitis (CRS) is a prevalent and heterogeneous inflammatory condition affecting the sinonasal mucosa, typically categorized into eosinophilic (type 2) and neutrophilic (type 1 or type 3) endotypes. 1 Eosinophilic CRS (ECRS) is distinguished by significant Th2 cytokine production (interleukin [IL]-4, IL-5, and IL-13), elevated immunoglobulin E (IgE) levels, tissue eosinophilia, and frequently coincides with the formation of nasal polyps.1,2 Conversely, neutrophilic CRS (NCRS) is characterized by predominant Th1/Th17 inflammation, marked by the production of interferon-gamma (IFN-γ), IL-17, and IL-1β with pronounced neutrophil infiltration. 3 Many patients with CRS exhibit overlapping inflammatory profiles, highlighting the necessity for a comprehensive understanding of how mixed or fluctuating immune responses contribute to disease severity and recurrence. 4 Therefore, robust animal models are indispensable for deciphering CRS pathophysiology and pre-clinical testing of candidate therapies.
The preponderance of existing murine CRS models is predicated by ovalbumin (OVA) in conjunction with adjuvant protocols originating from asthma research.1,5–8 Nevertheless, OVA is not an aeroallergen in humans and fails to elicit clinically relevant rhinosinusitis. Additionally, numerous OVA models require intraperitoneal sensitization to mitigate tolerance, a route of exposure that lacks a parallel in human disease, where allergen priming transpires across the airway epithelium.9–11 The attributes of these models impose limitations on physiological relevance and may obscure the translatability of immunological insights or therapeutic efficacy. Although a variety of CRS mouse models have been described, few studies have clearly delineated the resulting endotype, and none have generated and compared eosinophilic and neutrophilic phenotypes side-by-side within a single experimental framework.
In response to these constraints, our research group previously devised a physiologically relevant ECRS model that replaces OVA with a consortium of airborne allergens commonly associated with human disease—namely, house dust mite (HDM), Aspergillus fumigatus, Alternaria alternata, and Staphylococcus aureus (S. aureus) protease—administered exclusively via the intranasal route. 12 This methodology emulates natural exposure to aeroallergens and engenders substantial eosinophilic infiltration and epithelial remodeling in the absence of systemic priming, thereby offering a more accurate representation of human type-2 CRS.
Nonetheless, significant deficiencies persist. A complementary neutrophil-dominant CRS model developed under similar physiological conditions has yet to be established. Furthermore, the lack of a direct comparative analysis between eosinophilic and neutrophilic paradigms precludes the evaluation of endotype-specific therapeutics, such as corticosteroids. We therefore hypothesized that intranasal delivery of clinically relevant aeroallergens or innate danger signals would generate discrete eosinophilic and NCRS models amenable to direct comparison of immunopathology and steroid responsiveness. Here, we combined our allergen-based protocol with a lipopolysaccharide (LPS)/β-1,3-glucan/S. aureus protease regimen to establish parallel endotypes, enabling side-by-side assessment of stimulus-dependent inflammation and glucocorticoid sensitivity.
Materials and Methods
Animals and Ethical Approval
The use of animals in this study was approved by the Ethics Committee and Institutional Animal Care and Use Committee (protocol #2024-0059), and the study was conducted according to international guidelines (ARRIVE) on animal experiments. Seven-week-old female C57BL/6N mice (Orient Bio, Seongnam, Korea) were housed under specific-pathogen-free conditions (22 ± 2 °C, 50 ± 10% humidity, 12-hour light/dark cycle) with free access to food and water.
Establishment of Eosinophilic and Neutrophilic Chronic Rhinosinusitis Models
To generate ECRS, mice received intranasal instillations (30 µL per dose) of a clinically relevant airborne-allergen mixture—HDM (XPB82D3A25, Greer Laboratories Inc., 20 µg), A. fumigatus (XPM3D3A25, Greer Laboratories Inc., 20 µg), and A. alternata (XPM1D3A25, Greer Laboratories Inc., 20 µg), and a serine protease derived from S. aureus (P5254, Abnova, 1 µg)—three times weekly for either 4 or 12 weeks. Doses were selected based on our previous allergen model. 12 NCRS was induced with an innate stimulus cocktail consisting of LPS (L2630-10MG, Sigma, 10 µg), β-1,3-glucan from Euglena gracilis (89862-1G-F, Sigma, 10 µg), and protease from S. aureus (P5254, Abnova, 1 µg) on an identical schedule. Phosphate-buffered saline (PBS) served as a vehicle control. A parallel cohort received weekly intraperitoneal dexamethasone (D2915, Sigma, 2 mg/kg) beginning in week 3 (short-term protocol) or week 8 (long-term protocol) to examine corticosteroid responsiveness. Mice were euthanized 24 hours after the final intranasal challenge (Figure 1).
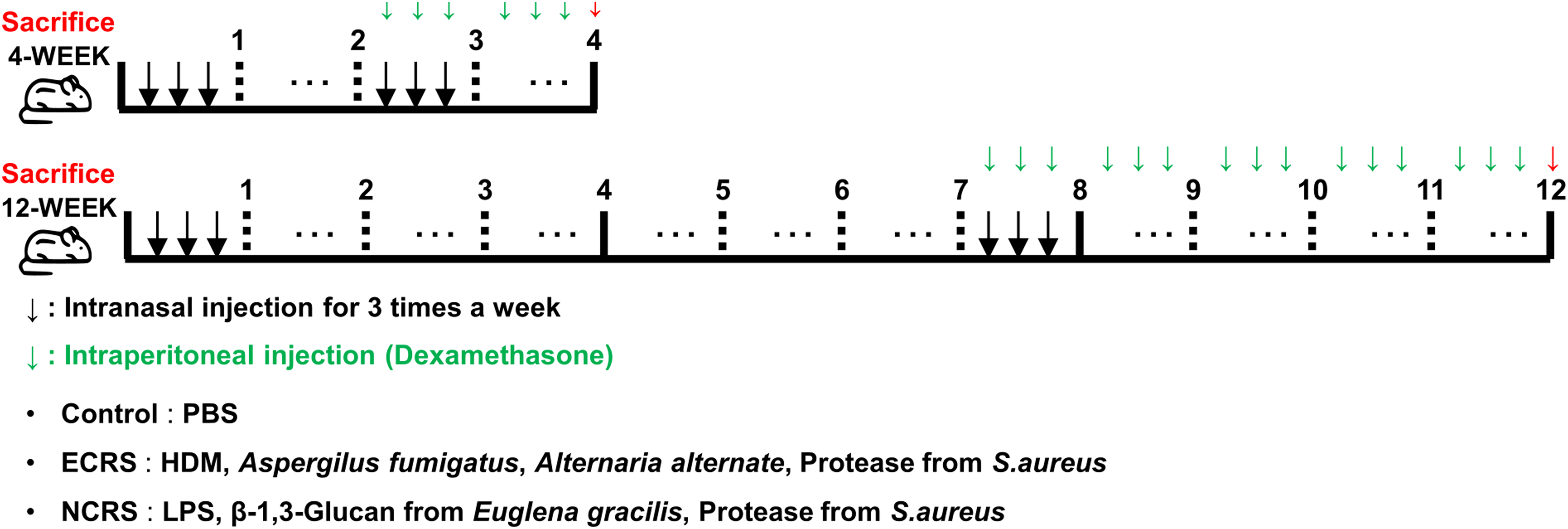
Experimental design for CRS induction and dexamethasone treatment. Schematic timeline for the 4-week (upper) and 12-week (lower) protocols in C57BL/6N mice. Downward black arrows denote intranasal instillations three times per week: control (PBS); ECRS (HDM 20 µg, Aspergillus fumigatus 20 µg, Alternaria alternata 20 µg, S. aureus protease 1 µg); NCRS (LPS 10 µg, β-1,3-glucan 10 µg, S. aureus protease 1 µg). Downward green arrows indicate weekly intraperitoneal dexamethasone (2 mg/kg) starting at week 3 (4-week cohort) or week 8 (12-week cohort). Red arrows mark sacrifice 24 hours after the final intranasal challenge. Specific sample sizes for each endpoint are provided in Figures 2-6. CRS, chronic rhinosinusitis; ECRS, eosinophilic CRS; NCRS, neutrophilic CRS; HDM, house dust mite; PBS, phosphate-buffered saline.
Stimulus Preparation and Standardization
To reflect the mixed environmental exposures in ECRS, we used an intranasal cocktail of HDM, fungal antigens (A. fumigatus, A. alternata), and a S. aureus-derived protease. HDM serves as a principal perennial indoor aeroallergen facilitating type-2 inflammation,13,14 and A. fumigatus exposure induces ECRS, effectively replicating sinonasal eosinophilia and tissue remodeling.8,15,16 Additionally, A. alternata quickly triggers epithelial IL-33 and type-2 responses, 17 while fungal proteases further amplify type-2 immunity via barrier disruption. 18 Bacterial proteases synergize with fungal allergens to promote barrier dysfunction,19,20 with final dosages established based on prior research. 12
To stimulate NCRS, we used pathogen-associated molecular patterns by intranasal delivery of LPS, β−1,3-glucan, and S. aureus protease. The repeated administration of LPS at doses of 5-10 µg in mice consistently triggers neutrophilic rhinosinusitis along with potential polyp formation; thus, we opted for 10 µg per dose as a reliable and well-tolerated amount.21,22 β-1,3-glucan serves as a Dectin-1 ligand, enhancing Th1/Th17-skewed airway responses and modulating HDM-driven inflammation; intranasal co-exposure studies indicate that β-glucan elevates airway inflammatory markers, and we opted for a 10 µg dosage to ensure efficacy while mitigating morbidity and regulating mucosal immunity.23–25 Lastly, S. aureus protease (1 µg) was incorporated to further emphasize epithelial barrier impairment and innate cytokine release without promoting type-2 bias, consistent with the known barrier-disrupting properties of protease allergens.19,20
Flow Cytometric Analysis
Whole blood collected into EDTA tubes was centrifuged to harvest plasma (stored at −70 °C), and the buffy coat underwent red blood cell lysis. Nasal mucosa was minced and digested in RPMI 1640 containing 2% fetal bovine serum and collagenase II (10 mg/mL, LS004176, Worthington), filtered through a 70-µm mesh, and resuspended. The resulting cell suspensions were then incubated with anti-CD16/32 (553141, BD Biosciences) to block Fc receptors and stained with Live/Dead™ Fixable Near-IR viability dye (L34976, Invitrogen) and fluorochrome-conjugated antibodies to CD45 (564279, BD Biosciences), Ly6G (75915, BD Biosciences), and Siglec-F (155508, BioLegend). Cells designated for intracellular analysis were subsequently fixed and permeabilized with the FOXP3 Fixation/Permeabilization Set (00-5523-00, Invitrogen). Samples were acquired on a BD Symphony A5SE flow cytometer. CD45+Ly6G−Siglec-F+ cells were defined as eosinophils, and CD45+Ly6G+Siglec-F− cells were defined as neutrophils.
Enzyme-Linked Immunosorbent Assay
On the day after completing the 4-week or 12-week modeling, blood plasma was collected from mice and analyzed for Eotaxin, total IgE, and HDM-specific IgE levels using enzyme-linked immunosorbent assay (ELISA) kits. Eotaxin levels were measured using the Mouse CCL11/Eotaxin DuoSet ELISA (DY420, R&D Systems, Bio-Techne) according to the manufacturer's protocol. Total IgE levels were quantified using the mouse IgE Uncoated ELISA Kit (88-50460-88, Invitrogen) following the product's instructions. HDM-specific IgE levels were determined using the Mouse Serum Anti-HDM IgE Antibody Assay Kit (3037, Chondrex) according to the manufacturer's protocol.
Gene Expression Analysis
Total RNA (TRIzol 15596026) was extracted, precipitated with isopropanol, and washed in 70% ethanol. cDNA was synthesized from 1 µg RNA (M-MLV, Invitrogen 28025-013) and amplified using KAPA SYBR® FAST (#KK4605) on a QuantStudio 3 system. The primer sequences are listed in Supplementary Table 1. Relative expression was calculated using the ΔΔCt method.
Histology and Immunofluorescence
Following PBS perfusion, mouse heads and lungs were fixed in 4% paraformaldehyde (4 days) and decalcified in 10% EDTA (20 days). Paraffin blocks were sectioned at 4 µm. Slides underwent hematoxylin and eosin (H&E), periodic acid-Schiff (PAS), and Sirius-red staining. For immunofluorescence, antigen retrieval (IHC-TeK IW-1100-1L) was followed by blocking in 5% bovine serum albumin and overnight incubation at 4 °C with anti-PRG2 (PA5-30130, 1:500; Thermo Fisher Scientific), anti-MPO (AF3667, 1:400; R&D Systems), or anti-MUC5AC (45M1, 1:500; Invitrogen). Alexa-568 secondary antibody (1:500) and Fluoromount™ + DAPI were applied. Imaging was performed on a Zeiss LSM 780 confocal microscope.
Statistical Analysis
All experiments were performed using 3-16 mice per group (n = 3-16), based on the specific assay, and the exact sample sizes are indicated in the respective figure legends. For histological assessments, although n = 3 mice per group may appear modest, statistical reliability was ensured by analyzing four non-overlapping fields per mouse and averaging these values as one biological replicate. This approach (3 mice × 4 fields = 12 measurements per group) provides adequate statistical power while accounting for intra-animal spatial heterogeneity and inter-animal biological variation.
Results
Eosinophilic Chronic Rhinosinusitis Model Exhibits Robust Eosinophilic Infiltration, Whereas Neutrophilic Chronic Rhinosinusitis Model Remains Neutrophil-Dominant
Repeated intranasal instillation in the ECRS model generated a pronounced eosinophilic infiltrate in the nasal mucosa as early as week 4, which intensified by week 12 (Figure 2A). At this later time point, a parallel increase in neutrophils became evident, indicating that chronic exposure drives a transition toward a mixed eosinophil-neutrophil inflammatory pattern. In contrast, the NCRS model remained consistently neutrophil-dominant with minimal eosinophil infiltration at both time intervals (Figure 2B).
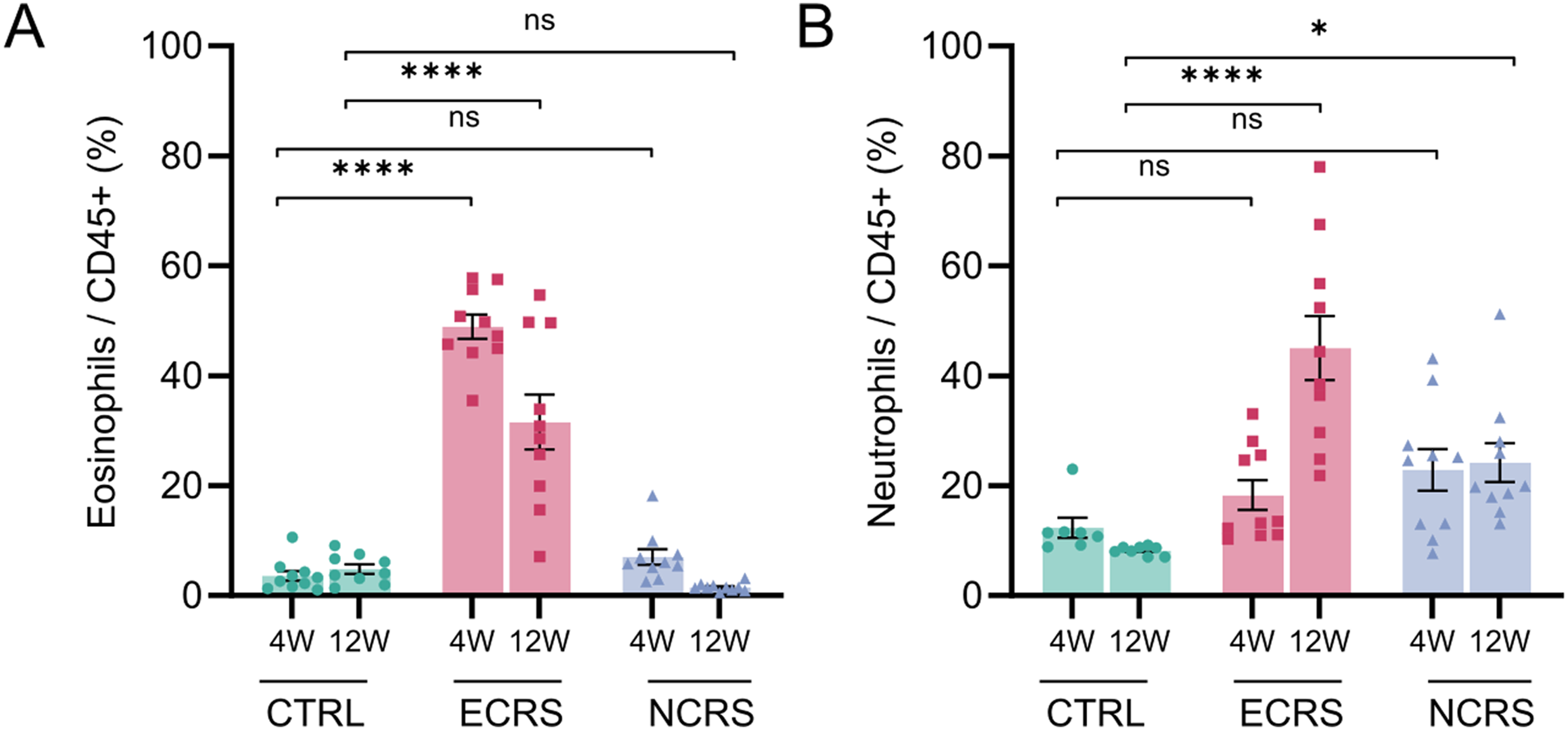
Eosinophil- and neutrophil-dominant inflammation in the nasal mucosa. (A) Percentages of CD45+Ly6G−SiglecF+ eosinophils and (B) CD45+Ly6G+SiglecF− neutrophils among CD45+ cells determined by flow cytometry at 4 weeks (4 W) and 12 weeks (12 W). Bars represent the mean ± SEM (n = 9-10 mice per group); P < .05, P < .01, **P < .0001; ns, not significant (two-way ANOVA with Tukey's post-hoc test).
Systemic Type-2 Biomarkers Reflect Local Eosinophilia, Although Circulating Granulocytes Remain Unchanged
Plasma analysis revealed significant elevations of eotaxin, total IgE, and HDM-specific IgE in ECRS, mirroring the local Th2 microenvironment (Figure 3A-C). NCRS values were comparable to those of controls, indicating limited systemic type-2 spillover. Despite marked mucosal infiltration, the proportions of circulating eosinophils and neutrophils in peripheral blood did not differ significantly across groups, underscoring the compartmentalized nature of CRS inflammation (Figure 3D, E).
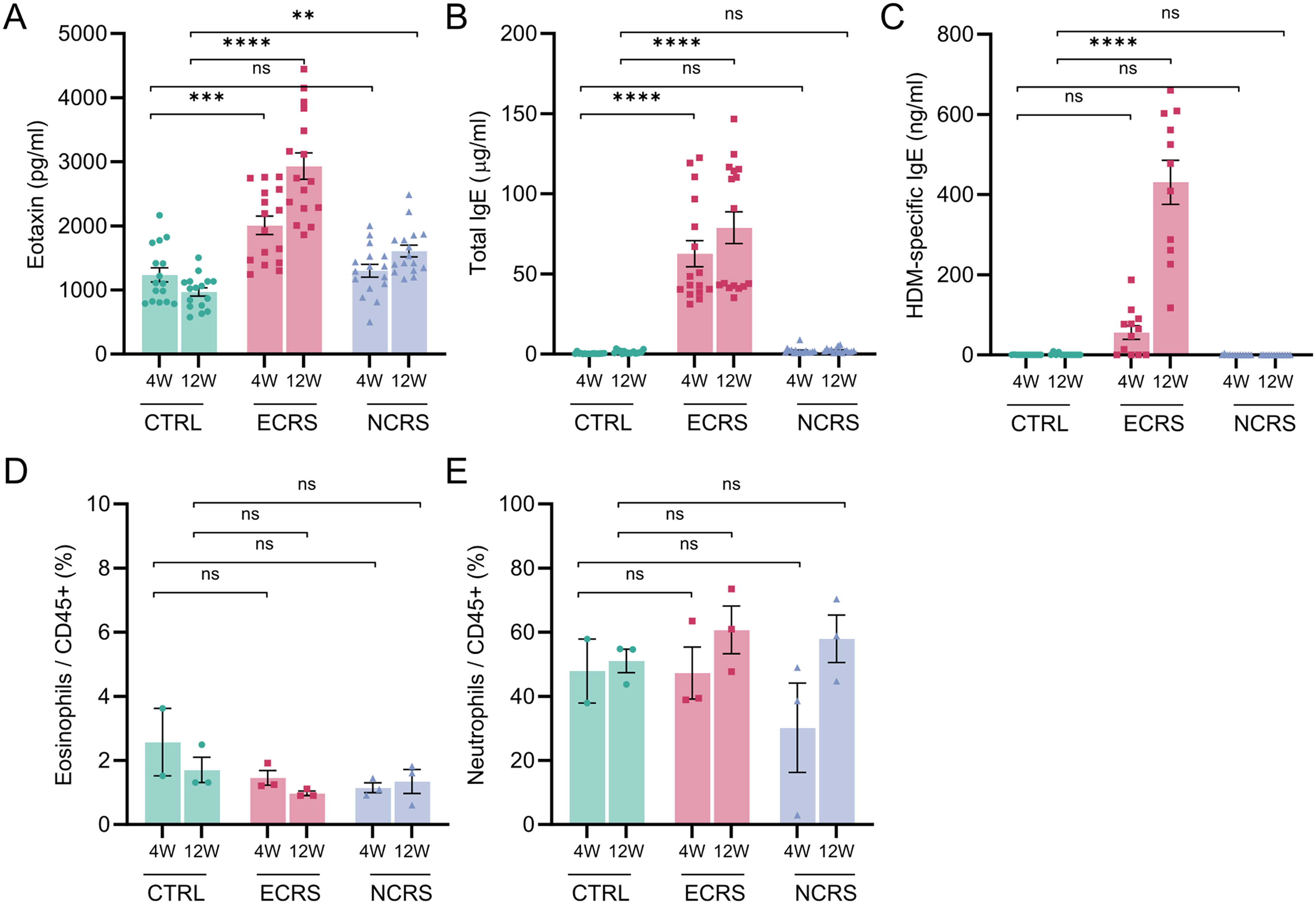
Systemic biomarkers and circulating granulocytes. (A) Plasma eotaxin (CCL11), (B) total IgE, and (C) HDM-specific IgE concentrations at 4 W and 12 W measured by ELISAs. (D) Percentages of circulating eosinophils (CD45+Ly6G−SiglecF+) and (E) circulating neutrophils (CD45+Ly6G+SiglecF−) among CD45+ blood leukocytes. Bars show the mean ± SEM (n = 16 mice per group for A-B, n = 12 for C, and n = 3 for D-E); P < .05, *P < .01, **P < .001, ***P < .0001; ns, not significant (two-way ANOVA with Tukey's post-hoc test). HDM, house dust mite; ELISA, enzyme-linked immunosorbent assay.
Distinct Cytokine Signatures Confirm Type-2 Versus Type-1/Type-17 Polarization
Quantitative PCR analysis of nasal tissue demonstrated significant upregulation of Ccl11, Il4, Il5, Il13, and Tslp in ECRS, which aligns with the established paradigm of type-2 signaling (Figure 4). Interestingly, transcripts typically associated with Th1/Th17 or tissue repair mechanisms—Il1b, Il6, Tnf, and Tgfb1—exhibited notable increases in the 12-week ECRS mucosa, in contrast to their persistently low levels in NCRS. This selective induction observed in late-phase ECRS coincided with the onset of neutrophil infiltration, implying that chronic eosinophilic inflammation transitions toward a heterogeneous eosinophil-neutrophil environment, accompanied by signals pertinent to wound healing. Conversely, the NCRS model was characterized by upregulation of Ifng and Il17a at both assessed time points, whereas Th2-associated genes remained at relatively low levels (Figure 4). These findings corroborate a type-2 bias in early ECRS that evolves over time, alongside a sustained Th1/Th17 profile in NCRS.
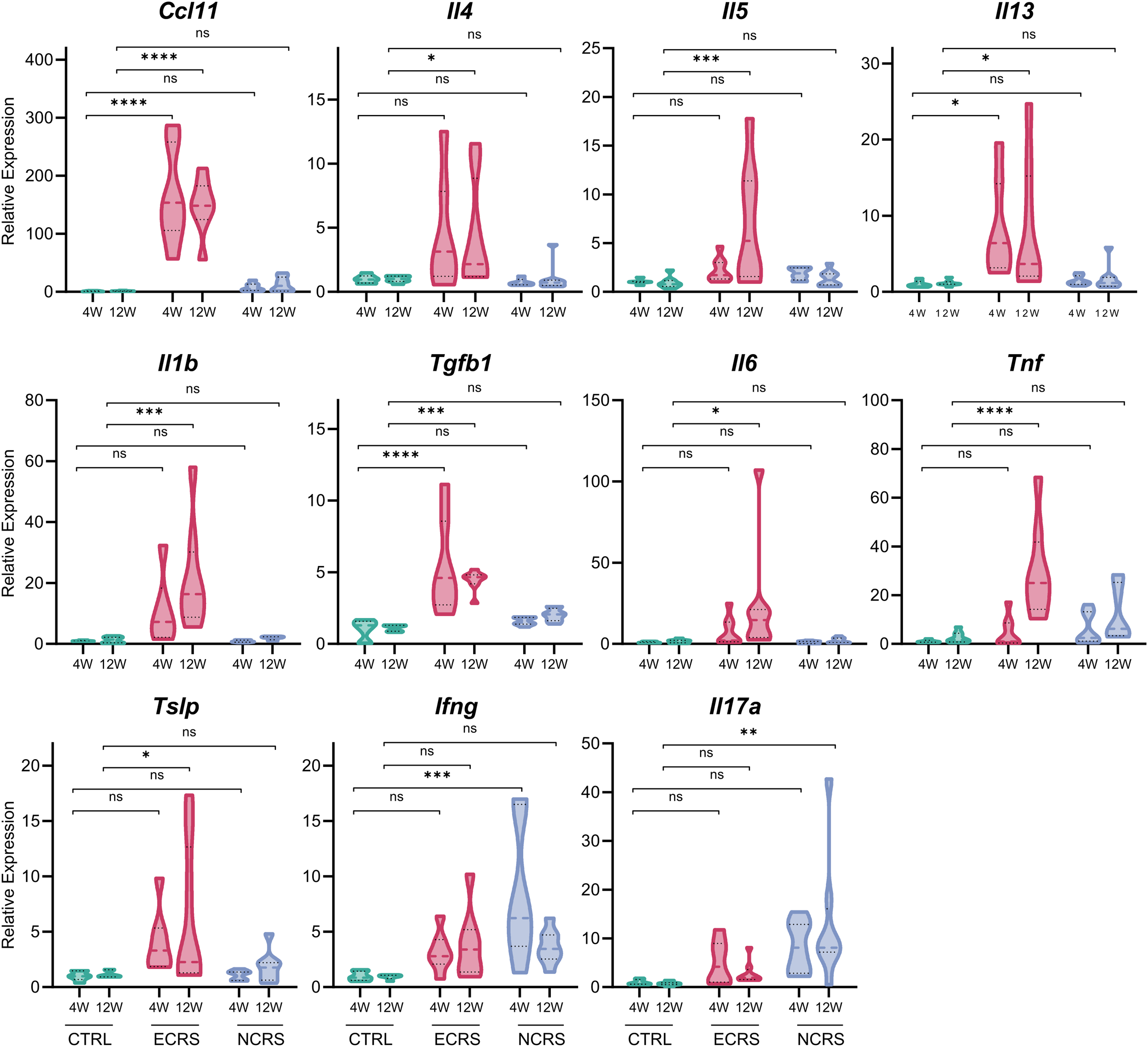
Nasal cytokine gene expression distinguishes Th2- and Th1/Th17-skewed endotypes. Violin plots display the relative mRNA abundance of Ccl11, Il4, Il5, Il13, Il1b, Tgfb1, Il6, Tnf, Tslp, Ifng, and Il17a in the nasal mucosa at 4 W and 12 W, normalized to B2 m and expressed as fold-change versus control (n =7-8 mice per group). Horizontal bars denote the median ± interquartile range. P < .05, *P < .01, **P < .001, ***P < .0001; ns, not significant (two-way ANOVA with Tukey's post-hoc test).
Histopathology Reveals Differential Tissue Remodeling and Mucus Hypersecretion
H&E staining highlighted epithelial thickening and dense inflammatory infiltrates in both models, with greater severity in ECRS (Figure 5A). PAS staining and MUC5AC immunofluorescence revealed extensive goblet cell hyperplasia and mucus accumulation in NCRS at both 4 and 12 weeks, a pattern consistent with reports that neutrophil-derived elastase and IL-17A can up-regulate MUC5AC in NCRS,26–28 whereas ECRS showed modest mucus changes at week 4 that became pronounced by week 12 (Figure 5B, F). Sirius-red staining identified abundant eosinophils in ECRS but sparse deposition in NCRS (Figure 5C). Immunofluorescence supported these findings; Figure 5D shows MBP-positive eosinophils restricted to ECRS, while Figure 5E reveals strong MPO-positive neutrophil signals in both models at both 4 and 12 weeks, with signal intensity further increased at the 12-week time point for each model. Collectively, these data indicate that NCRS drives early and persistent mucus hypersecretion with neutrophil predominance, whereas ECRS evolves from an eosinophil-dominated state to a mixed granulocyte profile accompanied by late-onset goblet cell expansion.

Histological and immunofluorescent assessment of the nasal mucosa. (A) H&E-stained sections illustrating the epithelial architecture at 4 W and 12 W with corresponding epithelial thickness quantification. (B) PAS staining and goblet cell counts. (C) Sirius-red staining and eosinophil enumeration. For all quantitative histological analyses, four non-overlapping mucosal regions were counted per mouse and the average was taken as one biological replicate. Data represent the mean ± SEM (n = 3 mice per group); P < .05, *P < .01, **P < .001, ***P < .0001; ns, not significant. (D-F) Immunofluorescence for MBP, MPO, and MUC5AC with DAPI-stained nuclei. Scale bars = 20 µm (A-F). H&E, hematoxylin and eosin; PAS, periodic acid-Schiff.
Dexamethasone Selectively Attenuates Eosinophilic, But Not Neutrophilic, Inflammation
Flow cytometry analysis elucidated that weekly administration of dexamethasone significantly diminished eosinophil populations within the nasal tissue (Figure 6A) and simultaneously reduced plasma levels of eotaxin and total IgE (Figure 6C, D). Conversely, the neutrophil quantities exhibited no significant alterations across either experimental model (Figure 6B). Histological evaluation corroborated that the epithelial layer thickness was reduced following dexamethasone treatment in both models; however, goblet cell hyperplasia—assessed through PAS staining—remained evident, particularly in NCRS (Figure 6E, F). Sirius-red staining indicated a pronounced decrease in eosinophilic aggregates in ECRS, whereas MPO immunofluorescence demonstrated that neutrophil accumulation remained largely unchanged in both models (Figure 6G, H). Collectively, these results suggest that glucocorticoids effectively attenuate eosinophilic responses while exerting negligible effects on neutrophilic or mucus-secreting components associated with CRS, a response pattern that aligns with established clinical observations.
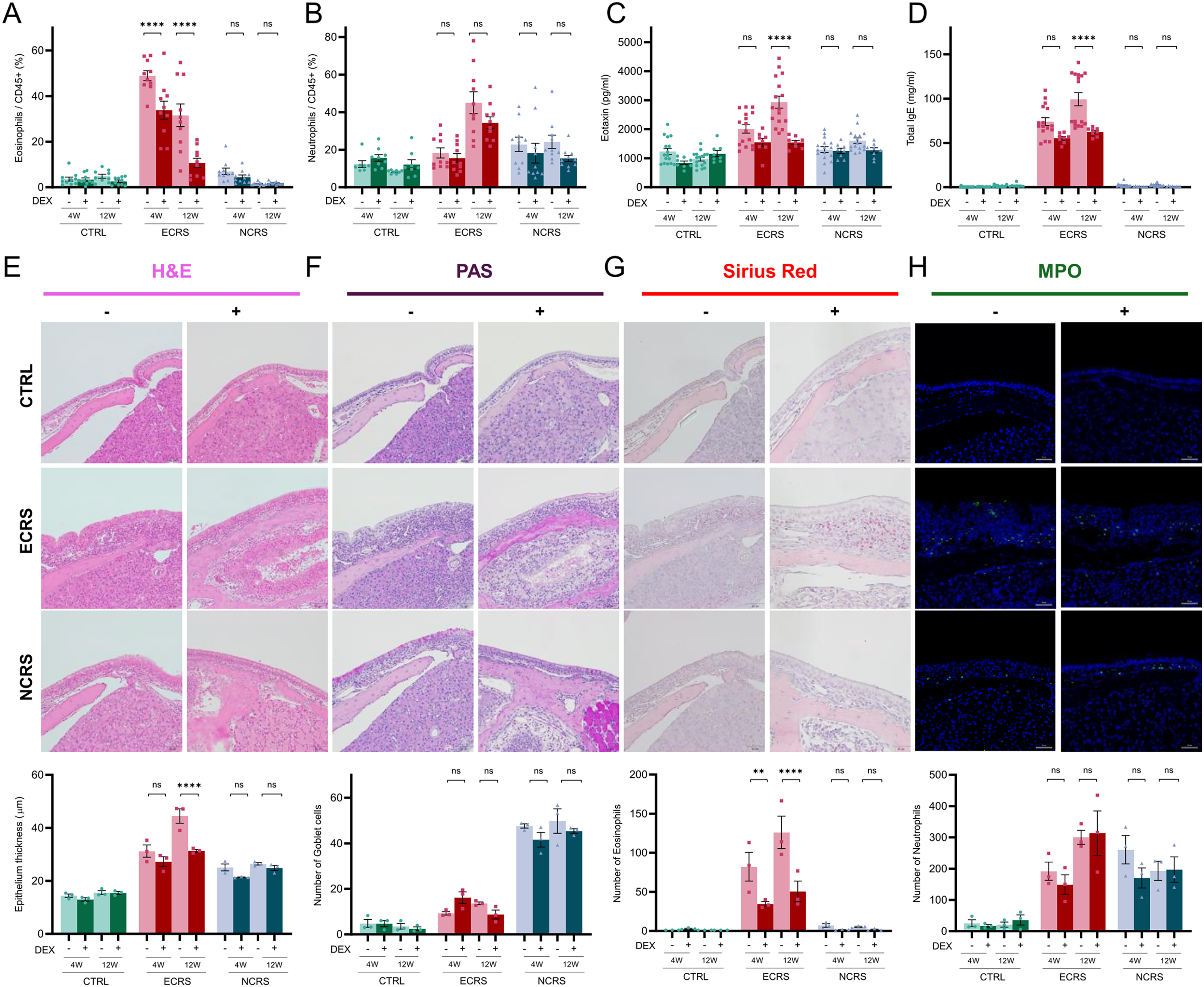
Dexamethasone selectively suppresses eosinophil-predominant inflammation. (A, B) Nasal CD45+Ly6G−SiglecF+ eosinophils and CD45+Ly6G+SiglecF− neutrophils with or without dexamethasone at 4 W and 12 W (n = 8-10 mice per group). (C, D) Plasma eotaxin and total IgE responses (n = 16 for Dexamethasone (-) groups and n = 8 for Dexamethasone (+) groups). (E-H) Histological and MPO immunofluorescent evaluation of the effects of dexamethasone on epithelial thickness, goblet cell hyperplasia, tissue eosinophils, and MPO+ neutrophils. The upper photomicrographs depict representative 12-week samples, and the accompanying bar graphs present pooled quantification from both the 4-week and 12-week cohorts. Each data point in panels (E-H) represents the mean of four non-overlapping fields per mouse (n = 3 mice per group). Scale bars = 20 µm (E-H). P < .05, *P < .01, **P < .001, ***P < .0001; ns, not significant.
Discussion
In this study, we demonstrated side-by-side generation and characterization of eosinophilic, neutrophilic, and mixed-type CRS endotypes using exclusively intranasal, physiologically relevant stimuli. Our objective was to establish physiologically relevant murine models that accurately reflect the two predominant inflammatory endotypes of CRS. By delivering clinically encountered airborne allergens or microbial danger signals exclusively through the intranasal route, we avoided the artificial intraperitoneal priming that compromises traditional OVA systems and more accurately mimicked the physiological process of natural airway sensitization. The resulting eosinophilic and neutrophilic models therefore provide a unified platform for investigating endotype-specific mechanisms and therapeutic approaches.
A comparative analysis of existing murine CRS models reveals that traditional OVA models, which typically involve intraperitoneal sensitization with alum followed by intranasal or aerosol exposure, consistently demonstrate significant Th2 polarization. However, OVA is not a major human aeroallergen, and systemic priming does not accurately reflect physiological conditions, while the variety of endotypes is limited.29–31 Bacterial infection models, such as intranasal administration of S. aureus or Pseudomonas aeruginosa, are essential for investigating infection-related diseases, but they usually lead to acute inflammation rather than chronic, potentially failing to reflect the chronic inflammatory condition seen in human CRS.32,33 Fungal models, like exposure to Aspergillus conidia or proteases, simulate allergic fungal diseases but mainly lead to eosinophilic/type-2 responses with potential batch variability.29,34 In contrast, our intranasal ECRS/NCRS system, using clinically relevant allergens/PAMPs, creates unique endotypes within the same genetic context and allows for precise timing (4 or 12 weeks) without the inconsistencies of live cultures, thereby improving current methods.
The two paradigms exhibited markedly different immunopathological profiles. ECRS displayed canonical type-2 features—eosinophil predominance, Th2 cytokine expression, and elevated IgE—but, by week 12, also showed up-regulation of Il1b and Tgfb1 and a secondary neutrophil influx, yielding a mixed granulocytic milieu. This evolution parallels clinical observations that indicate severe or recalcitrant ECRS can harbor combined eosinophil-neutrophil signatures.1,35 NCRS, in contrast, remained stably neutrophil-dominant with persistent Ifng and Il17a expression, offering a robust model of type-1/3 CRS. Our findings indicate that a primarily type-2 inflammatory response may evolve to exhibit neutrophilic characteristics, transitioning into a mixed CRS phenotype, thus underscoring the potential significance of early medical or surgical intervention for long-term disease management and outcomes. The initiating stimulus shapes the early endotype, with innate danger signals leading to a neutrophil-dominant phenotype; however, additional allergenic cues or prolonged exposure may attract eosinophils, resulting in a mixed profile.
Several hypotheses elucidate the shift from a Th2-dominant phenotype to a mixed phenotype. Chronic allergen and protease exposure compromises the epithelial barrier, triggering damage-associated molecular patterns that boost type-2 immune responses and promote microbial translocation, activating TLR and Dectin-1 pathways, which results in increased IL-1β and IL-6 levels.36,37 Subsequently, increased IL-1β plays a pivotal role in driving epithelial-mesenchymal transition and fibroblast activation, which are critical processes in chronic inflammatory conditions.38,39 IL-1β not only enhances the secretion of neutrophil-recruiting chemokines but also, in conjunction with IL-6 and TGF-β, promotes Th17 differentiation, creating a feedback loop that exacerbates inflammation.40,41 Future studies using loss-of-function approaches, pharmacologic blockade, and multi-timepoint single-cell RNA-seq will test these mechanisms and define causal nodes.
Corticosteroid testing highlighted these differences. Dexamethasone effectively eliminated eosinophilia, reduced epithelial hypertrophy, and decreased IgE levels in ECRS, yet failed to diminish the neutrophil burden or goblet cell hyperplasia in either model. This finding mirrors clinical experience, where steroids effectively control type-2 disease but demonstrate limited efficacy in neutrophil-predominant CRS.42–45 Importantly, our data indicate that the 4-week protocol generates a near-pure eosinophilic phenotype suitable for evaluating anti-type-2 interventions, whereas the 12-week protocol models late-stage mixed inflammation and steroid resistance. By modulating the type of intranasal stimulus and the duration of exposure, our platform can be tuned to model the full clinical spectrum: a 4-week allergen regimen yields a pure eosinophilic phenotype; extending allergen delivery to 12 weeks produces mixed ECRS with combined eosinophilic and neutrophilic features; and substituting allergens with LPS/β-glucan generates a neutrophilic profile that is inherently corticosteroid-resistant. This flexibility enables direct comparison of endotypes within a single genetic background and facilitates pre-clinical evaluation of phenotype-specific therapies.
Given the distinct physiological and immune kinetics across species, the definition of CRS in humans as symptoms persisting for ≥12 weeks is not directly translatable from murine models. 1 Instead, we prioritize histopathological chronicity, marked by persistent inflammatory infiltrates and tissue remodeling observable within a 4-week timeframe. Prior murine work similarly shows that 3-6 weeks of repeated allergen or PAMP exposure yields stable histologic/molecular hallmarks of chronic sinonasal inflammation. For example, a model utilizing OVA along with fungal protease over 7 weeks demonstrates advanced eosinophilic remodeling 6 ; a 3-week model employing intranasal LPS reveals mucosal thickening, neutrophilic infiltration, and olfactory dysfunction that stabilizes by the third week 46 ; and protocols involving Aspergillus protease at 6 and 12 weeks yield reproducible ECRS, with more pronounced remodeling noted at the 12-week mark. 16 Collectively, these data support 4 weeks as a reasonable murine threshold for chronic sinonasal inflammation and 12 weeks as a progression window capturing endotypic drift—emergence of mixed Th2 with superimposed Th1/Th17 and neutrophilia—and attenuated steroid responsiveness, consistent with long-standing, refractory CRS in patients.
Should this trajectory be confirmed clinically, early phenotype-guided intervention—pharmacological or surgical—may improve the long-term prognosis. Notably, contemporary cohort studies report substantial fractions of CRS patients with concurrent Th2 and Th17/neutrophil signatures, particularly in Asian populations, and these mixed phenotypes exhibit higher recurrence and reduced steroid responsiveness—paralleling our 12-week ECRS findings.47–50
Several limitations warrant consideration. Murine sinonasal anatomy restricts polyp size, high-dose stimuli may exaggerate pathological changes, and our models omit microbial community dynamics that influence human CRS. Furthermore, although the present investigation offers an extensive immunopathological and histological analysis, it predominantly focuses on these evaluations rather than on functional outcomes. We concur that integrating functional assessments, such as behavioral investigations to assess olfactory deficits or advanced imaging modalities like micro-CT for quantifying sinus opacification, would yield a more holistic appraisal of the model's functional implications. Moreover, despite our transcript-level profiling, we did not quantify these cytokines at the protein level in this dataset. Future prospective cohorts will incorporate nasal lavage and tissue homogenate assays such as ELISA or multiplex bead-based platforms to confirm protein expression over time. Although the current work does not delineate every molecular pathway driving each endotype, the establishment of a flexible in vivo platform that captures key features of diverse patient phenotypes constitutes a significant advance for subsequent mechanistic and therapeutic studies.
In conclusion, our physiological ECRS and NCRS models effectively capture the spectrum, evolution, and steroid responsiveness of human CRS endotypes. They provide a versatile platform to elucidate pathogenic mechanisms, identify biomarkers, and evaluate endotype-tailored interventions that may ultimately improve clinical outcomes across the CRS spectrum.
Supplemental Material
sj-docx-1-ajr-10.1177_19458924261456360 - Supplemental material for Murine Eosinophilic and Neutrophilic Chronic Rhinosinusitis Models Reveal Phenotype-Specific Steroid Responses
Supplemental material, sj-docx-1-ajr-10.1177_19458924261456360 for Murine Eosinophilic and Neutrophilic Chronic Rhinosinusitis Models Reveal Phenotype-Specific Steroid Responses by Miran Kang, Dachan Kim, Juhee Seo, Yosep Kim, Subin Kim, Min-Seok Rha, Chang-Hoon Kim and Hyung-Ju Cho in American Journal of Rhinology & Allergy
Footnotes
Acknowledgements
This work was supported by National Research Foundation of Korea (NRF) grants funded by the Korean government (MSIT) (No. RS-2023-00220853 and NRF-2021R1A2C2010811) to H.J. Cho. The authors thank BioScience Writers, LLC, for technical editing support.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Research Foundation of Korea (NRF-2021R1A2C2010811) to HJC and also supported by a grant of the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (grant number: RS-2025-25459824).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.