
Abstract
Introduction:
Sperm cryopreservation induces intracellular oxidative stress. Sesamol (SE), as an antioxidant, mitigates cryodamage caused by spermatozoa cryopreservation. This study was designed to evaluate the protective effects of SE-loaded nanostructured lipid carriers (SE-loaded NLCs) on frozen sperm quality.
Methods:
SE-loaded NLCs were synthesized using hot homogenization, followed by characterization for particle size, morphology, and encapsulation efficiency (EE). Normozoospermic semen samples were collected from 20 men and prepared according to the WHO 2021 guidelines. The samples were allocated into experimental groups of fresh, frozen–thawed control, SE-treated, blank NLCs, and SE-loaded NLCs. Following semen cryopreservation and thawing protocols, sperm quality was evaluated regarding motility, vitality, morphology, membrane and acrosome integrity, mitochondrial membrane potential (MMP), and DNA fragmentation using established staining and microscopy techniques. Statistical analysis was also performed at the significance level of 0.05.
Results:
NLCs were successfully prepared with a mean particle size of 154 nm, zeta potential of −17.6 mV, and EE of 72 ± 2.64%. Fluorescence microscopy confirmed the efficient uptake of NLCs by spermatozoa within 60 minutes. The optimal SE dose was determined as 50 µM for cryoprotection. Sperm treated with SE-loaded NLCs showed improvements (p < 0.05) in motility, viability, membrane and acrosome integrity, MMP, and reduced (p < 0.05) DNA fragmentation than other groups.
Conclusion:
Application of SE-loaded NLCs showed accelerated protective effects compared with free SE, highlighting the associated potential in the enhancement of sperm quality during cryopreservation. SE-loaded NLCs effectively enhanced sperm quality indices during cryopreservation. Thus, SE-loaded NLCs represent a promising strategy to mitigate cryoinjury and improve fertility preservation outcomes.
Keywords
Introduction
Sperm cryopreservation is an established technique widely utilized in assisted reproductive technologies to enhance male fertility, particularly in patients undergoing cancer chemotherapy, as well as those affected by urogenital and autoimmune conditions.1,2 During sperm cryopreservation, spermatozoa are subjected to osmotic fluctuations, temperature-induced stress, and predominantly oxidative damage. 3 Oxidative stress is widely recognized as the principal mechanism underlying cryodamage in both reproductive and nonreproductive cells. This phenomenon predominantly manifests during the thawing phase, where the reintroduction of oxygen can lead to a rapid surge in glucose-mediated oxidative metabolism, resulting in markedly elevated production of reactive oxygen species (ROS). 4 The current process is primarily attributable to impaired mitochondrial electron transport chain function, as mitochondria constitute the predominant intracellular source of ROS in human spermatozoa, particularly following cryopreservation.5,6 Thus, one of the key biochemical consequences of the freezing and thawing process is the overproduction of ROS such as O−2, H2O2, and OH, which adversely induce lipid peroxidation of unsaturated fatty acids in the sperm plasma membrane.7,8 Elevated ROS levels and lipid peroxidation in sperm induce motility reduction, DNA fragmentation, alter membrane permeability, and trigger apoptotic signaling pathways, which totally impair the fertilizing capacity of spermatozoa.9–11 Current evidence supports the fact that both enzymatic and nonenzymatic forms of antioxidants enhance sperm quality by ROS neutralization and interrupting peroxidation cascades, thereby mitigating oxidative damage.12,13 According to recent findings, supplementation of sperm cryopreservation media with natural or synthetic antioxidants can mitigate the detrimental effects associated with sperm cryoinjury.14–16 Sesamol (SE), chemically identified as 3,4-methylenedioxyphenol, is a natural phenolic compound isolated from the roasted seeds of Sesamum indicum Linn.17,18 The associated antioxidant activity, anti-inflammatory, and anti-apoptotic properties on the male reproductive system and sperm parameters are substantiated by previous research.19,20 Since SE exhibits limited aqueous solubility (logP = 1.29) and poor bioavailability, incorporation of an appropriate drug delivery system to enhance the associated therapeutic efficacy seems necessary. 21 In recent years, nanotechnology has advanced as a promising approach to address challenges related to drug bioavailability, toxicity, and therapeutic efficacy.22,23 Nanostructured lipid carriers (NLCs), classified as second-generation lipid-based nanocarriers, offer distinct advantages over first-generation solid lipid nanoparticles (SLNs). These benefits include enhanced physicochemical stability, reduced risk of gelation, and increased drug-loading capacity (LC), attributable to the incorporation of a binary mixture of solid and liquid lipids within the NLC matrix.24,25 Previous studies regarding sperm cryopreservation have predominantly relied on the application of free antioxidants with limited stability and bioavailability, while nanocarrier-based strategies remain insufficiently explored. Moreover, SE, a moderately lipophilic antioxidant with well-established properties, has not been evaluated in human sperm cryopreservation. Due to the lipophilic nature of SE, NLCs provide suitable encapsulation of lipophilic compounds, enhanced stability, controlled release, and biocompatibility; thus, these agents appear to be a proper system for SE delivery. 26 Accordingly, the present study aimed to assess the effects of SE-loaded NLCs on human sperm quality parameters following the cryopreservation process.
Materials and Methods
Preparation and characterization of SE-loaded NLCs
The hot homogenization method was used to prepare SE-loaded NLCs. Initially, the lipid phase with various Precirol concentrations (Gattefosse, Saint-Priest Cedex, France) and Miglyol was heated in a warm water bath (70°C). SE (S3003, Sigma-Aldrich) was dissolved in dimethyl sulfoxide 2% (DMSO) and then added to the molten lipid phase. Subsequently, at the same temperature, the aqueous phase containing Poloxamer 407 (Sigma-Aldrich, Germany) was gradually added dropwise to the lipid phase under high-speed homogenization (Heidolph, Germany) at a stirring rate of 22,000 rpm. The temperature of the hot oil–water formulation was subsequently reduced to facilitate the recrystallization of the formed nanoemulsion into NLCs. Particle size, polydispersity index (PDI), and zeta potential of the SE-loaded NLCs were determined using dynamic light scattering (DLS) (Nano ZS, Malvern Instruments, UK). The Z-average value was quantified based on intensity and conveyed particle size. Scanning electron microscopy (SEM) was also used to analyze the morphological properties of the nanoparticles (NPs). 27
Drug encapsulation efficiency and LC assessment
The encapsulation efficiency (EE) of SE was assessed by quantification of the amount of free (unencapsulated) drug using a centrifugal filter tube (Amicon® filter, 100 kDa MWCO, Millipore, UK). In this regard, 1 mL of the formulation was mixed with 1 mL of DMSO (1% vol/vol) to facilitate the dissolution of any unloaded SE, followed by centrifugation (400 g, 8 minutes) to isolate free SE from encapsulated drugs. The measurement of unloaded drugs was then conducted using an ultraviolet-visible spectrophotometer (Ultrascope 2000®, Pharmacia Biotech, UK) at λmax 273.8 nm. 28 The EE was calculated using the following equation: EE (%) = (Ct − Caq)/Ct × 100, where Caq represents the concentration of the unloaded drug in the aqueous phase, and Ct is the total concentration of the drug initially added. Similarly, the LC was determined using the equation: LC (%) = WL/WT × 100, where WL is the weight of the encapsulated SE, and WT is the total weight of lipids used in the NLC formulation.
Releasing pattern and stability assessment
Drug release studies were performed in phosphate-buffered saline (PBS) at 25°C and 4°C by replacement of SE-loaded NLCs into cellulose acetate dialysis bags. At predetermined time intervals, the release medium was analyzed using a UV-Vis spectrophotometer (Ultrascope 2000®, Pharmacia Biotech, UK) at a wavelength of 273.8 nm (λmax). Additionally, the physical stability of the optimized NLC formulation was evaluated over a 2-month storage period at 2°C–6°C by monitoring particle size, PDI, and EE.
Cellular uptake
For evaluating NLCs uptake by sperm cells, fluorescence-labeled NLCs were prepared using rhodamine-B (RH-B). Briefly, RH-B (0.5% w/w regarding the weight of the lipid) was used to prepare fluorescence-labeled NLCs. Then, the dialysis method was used to remove the unloaded RH-B. In the next step, spermatozoa were incubated with fluorescence-labeled NLCs for 30 and 60 minutes. Finally, a fluorescence microscope (Olympus microscope Bh2RFCA, Japan) was used to measure the absorption of NLCs after sperm suspensions were rinsed three times with PBS. 29
Ethical considerations and semen sample collection
The Research Ethics Committee of Tabriz University of Medical Sciences (Tabriz, Iran; IR.TBZMED.REC.1402.243) approved the procedures of the present study. All participants referred to the infertility center at Alzahra Hospital in Tabriz (Tabriz, Iran) were informed regarding the study, and written informed consent was obtained from each individual. Then, semen samples from 20 normozoospermic men (aged 20–45 years) were collected via masturbation following 3–5 days of ejaculatory abstinence. Study exclusion criteria were a history of hormonal treatments, diabetes mellitus, exposure to chemotherapy or radiation therapy, alcohol consumption, varicocele, and smoking. After semen collection, the samples were left to liquefy at 37°C for 30 minutes. All semen samples were analyzed according to the World Health Organization 2021 (WHO 2021) guidelines. 30
Freezing and thawing protocols
Following semen liquefaction, Ham’s F-10 medium containing 5% albumin was added to the samples and followed by centrifugation (500 g, 10 minutes). Then, the supernatant was removed. Next, Ham’s F-10 medium containing 5% albumin was added to the pellet and recentrifuged (500 g, 5 minutes). Finally, the semen samples were equilibrated at room temperature for 10 minutes following the gentle addition of an equal volume of CryoSperm™ medium (Origio, France). Subsequently, the samples were transferred to straws and placed horizontally on liquid nitrogen vapor for 15 minutes prior to plugging into the liquid nitrogen. After 1 week of storage, the frozen straws were thawed in a 37°C water bath.31,32 Subsequently, Ham’s F-10 medium containing 5% albumin was added. The samples were then centrifuged, the supernatant was discarded, and the pellet was resuspended in Ham’s F-10 medium containing 5% albumin.
Experimental design
The present study was divided into two consecutive experiments: first, determination of SE optimal concentration and second, assessment of optimal dose of SE and SE-loaded NLCs on sperm parameters. In the first experiment, seven normozoospermic semen samples were cryopreserved according to the previously described protocol; however, prior to the addition of the cryoprotectant, the spermatozoa were incubated (60 minutes) with varying concentrations of SE (0, 5, 15, 25, 50, and 100 µM) to determine the optimal concentration. Then, sperm motility (total and progressive), vitality, and morphology were measured. In the second experiment, 20 normozoospermic semen samples were each divided into five equal parts and assigned to the following groups: (A) Fresh group (noncryopreserved), (B) Frozen–thawed control group (no treatment), (C) NLC group (treated with unloaded NLC), D) SE group (treated with 50 µM SE as optimal dose), and (E) SE-loaded NLC group (treated with NLCs loaded with 50 µM SE). All treated groups (C, D, and E) were incubated with respective formulations for 60 minutes prior to freezing as described previously.
Assessment of sperm motility
Two expert technicians were hired to evaluate total and progressive sperm motility according to the WHO 2021 guidelines. Briefly, 10 µL of thawed and washed sperm suspension was placed on the slide and covered by a coverslip. Then, 400 sperm were counted using a light microscope (Nikon ECLIPSE E200, Japan) at 400× magnification.
Sperm vitality assessment
Sperm vitality was measured by eosin Y staining. Initially, 10 µL of sperm sample was mixed with 10 µL of eosin solution. Then, a thin smear on a glass slide was prepared immediately. Finally, a coverslip was placed on smear, and at least 200 spermatozoa were examined under a light microscope (Nikon ECLIPSE E200, Japan). In this technique, spermatozoa with noncolor or pale pink head were considered alive, and spermatozoa with a red or dark pink head were considered dead. 33
Sperm morphology assessment
Diff-Quik staining kit (Ideh Varzan Farda, Tehran, Iran) was used to assess sperm morphology according to the manufacturer’s protocols. In this regard, sample smears were mixed with two staining solutions. Morphological criteria were assessed according to WHO 2021 guidelines. In this procedure, at least 200 spermatozoa were examined under a light microscope (Nikon ECLIPSE E200, Japan).
Sperm membrane integrity assessment
Sperm plasma membrane integrity was determined by hypo-osmotic swelling (HOS) test. In this method, 10 µL of each sample was incubated with 100 µL of hypo-osmotic solution at 37°C for 30 minutes. Finally, 10 µL samples were placed on a microscopic slide, and at least 200 spermatozoa were examined under a light microscope (NikonECLIPSE E200, Japan). Spermatozoa with swollen-coiled or curled tails were recorded as intact plasma membranes. 33
Sperm DNA fragmentation detection
The sperm chromatin dispersion (SCD) kit (Halosperm®, Ideh Varzan Farda, Tehran, Iran) was employed for detection of sperm DNA fragmentation (SDF), following the manufacturer’s protocols. Initially, 50 µL of semen sample was combined with melted agarose in a tube, and 30 µL was applied to a microscope slide. Then, the coverslip was removed after 5 minutes of incubation at 4°C. The slide received sequential acid denaturation (7 minutes) and lysis treatment (15 minutes). After washing with distilled water, dehydration was carried out in 70%, 90%, and 100% ethanol for 2 minutes, and then Wright staining was performed. 34 After the sperm staining, bright-field microscopy (Nikon ECLIPSE E200, Japan) was used to examine at least 200 spermatozoa for the status of halos. According to the manufacturer’s instructions, four categories were determined for the status of halos: (A) sperm with a large halo (nonfragmented DNA), (B) sperm with a medium halo (nonfragmented DNA), (C) sperm with a small halo (fragmented DNA), and (D) sperm without a halo (fragmented DNA). The percentage of SDF was calculated by the following formula: SDF (%) = (number of sperm with fragmented DNA/total sperm counted) × 100.
Acrosome integrity assessment
The spermatozoa acrosome status was evaluated using fluorescein isothiocyanate-conjugated Pisum sativum agglutinin staining (FITC-PSA, Sigma-Aldrich, Germany). Initially, sample smears were prepared on slides and dried at room temperature. Then, smears were fixed with methanol and incubated with 50 µL FITC-PSA for 30 minutes in a dark room. Finally, at least 200 spermatozoa were examined under a fluorescence microscope (BX51, Olympus, Japan) at 1000× magnification. Spermatozoa heads with bright green fluorescence were identified as intact acrosome, while spermatozoa heads without or with pale green fluorescence were considered as disrupted acrosome. The percentage of sperm with acrosome integrity was calculated by the following formula: acrosome integrity (%) = (number of sperm with intact acrosome/total sperm counted) × 100. 35
Mitochondrial membrane potential assessment
Sperm mitochondrial membrane potential (MMP) was assessed using JC-1 as a lipophilic cationic dye (T3168, Molecular Probes, Invitrogen Life Sciences, Fullerton, CA, USA). Briefly, sperm samples were incubated with 1 µL of 200 µM JC-1 dye stock solution (diluted in DMSO) in a dark environment at 37°C for 40 minutes. In this technique, at least 200 spermatozoa were examined under a fluorescence microscope (BX51, Olympus, Japan) at 1000× magnification. Spermatozoa with yellow/orange fluorescence in the sperm midpiece were assumed as high mitochondrial membrane activity, and spermatozoa with green fluorescence were considered as low mitochondrial membrane activity. The percentage of sperm MMP was calculated by the following formula: MMP (%) = (number of sperm with high mitochondrial membrane activity/total sperm counted) × 100. 35
Statistical analysis
The analysis of the data was carried out using SPSS software (version 26, Chicago, USA). One-way analysis of variance (ANOVA) was performed, followed by Tukey’s post hoc test to determine significant differences among all evaluated parameters. Data were reported as mean ± standard deviation (SD), and p value <0.05 was considered statistically significant.
Results
Characterization and evaluation of SE-loaded NLCs
SE-loaded NLCs were developed by a modified hot homogenizing method, utilizing various proportions of Precirol, Miglyol, and Poloxamer. The optimal formulation was composed of 60 mg Precirol, 10 mg Miglyol, and 40 mg Poloxamer as the central substance of NPs, stabilizer, and surfactant of NLC, respectively. According to the DLS graph, the average diameter size of NPs was 154.3 ± 28.59 with PDI = 0.25 (Fig. 1A). The average zeta potential value of NPs was −17.6 mV (Fig. 1B). Also, SEM images confirmed the DLS results (Fig. 1C).

Characteristics of SE-loaded NLCs.
Drug EE, LC, releasing pattern, and stability
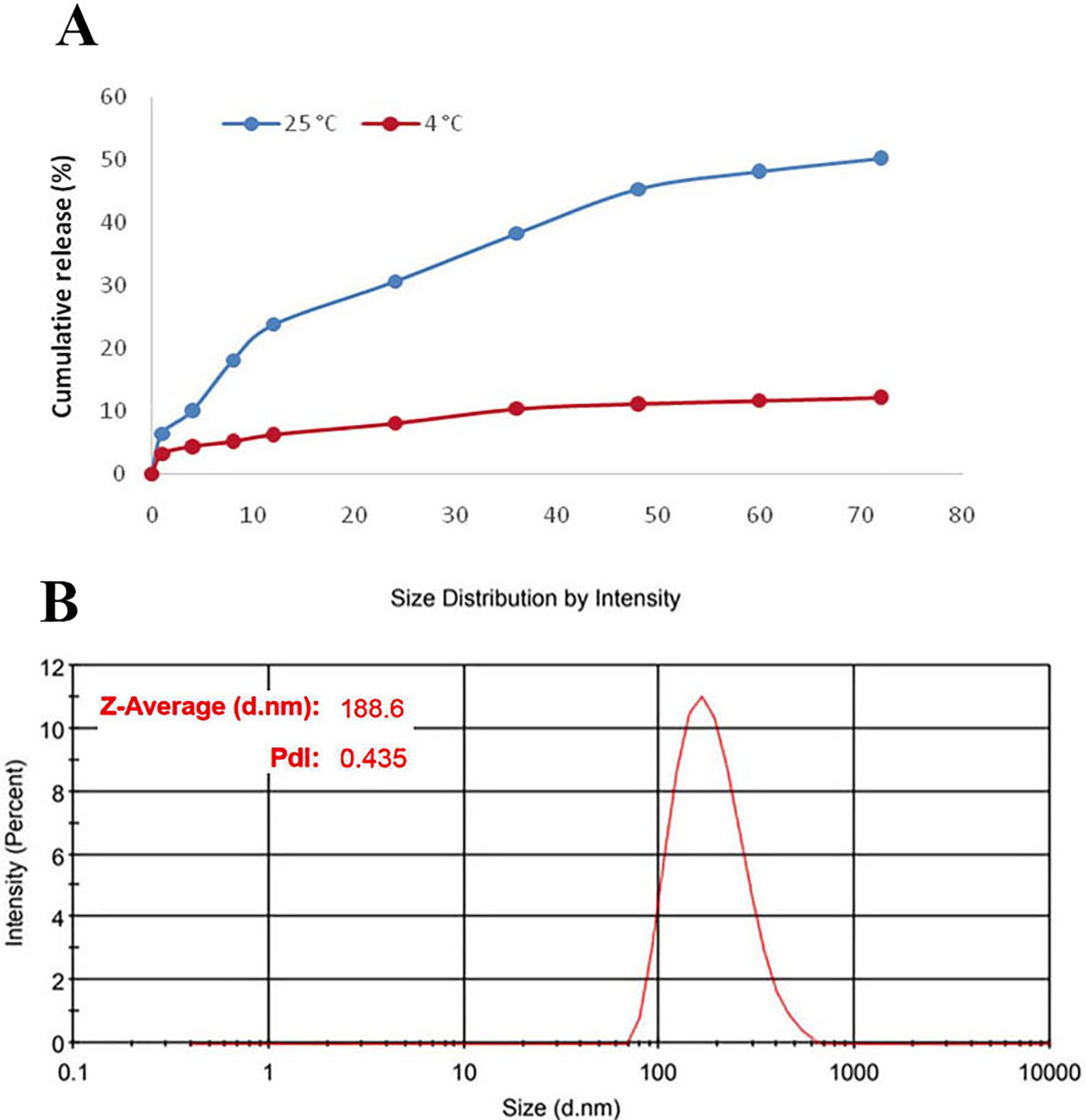
The averages of EE and LC in the optimum formulation were 72 ± 2.64% and 65.45 ± 2.4%, respectively. As shown in Figure 2A, SE released from NLCs at 25°C and 4°C over 72 hours was 50.3% and 12.2%, respectively. After 2 months of storage, the size and PDI of the SE-loaded NLCs were measured as 188.6 nm and 0.435, respectively (Fig. 2B). Also, the stability study illustrated that 12% of SE was released from the NLCs after 2 months of storage at 2°C–6°C. This result supports their potential stability and suitability for biomedical applications.
NPs uptake
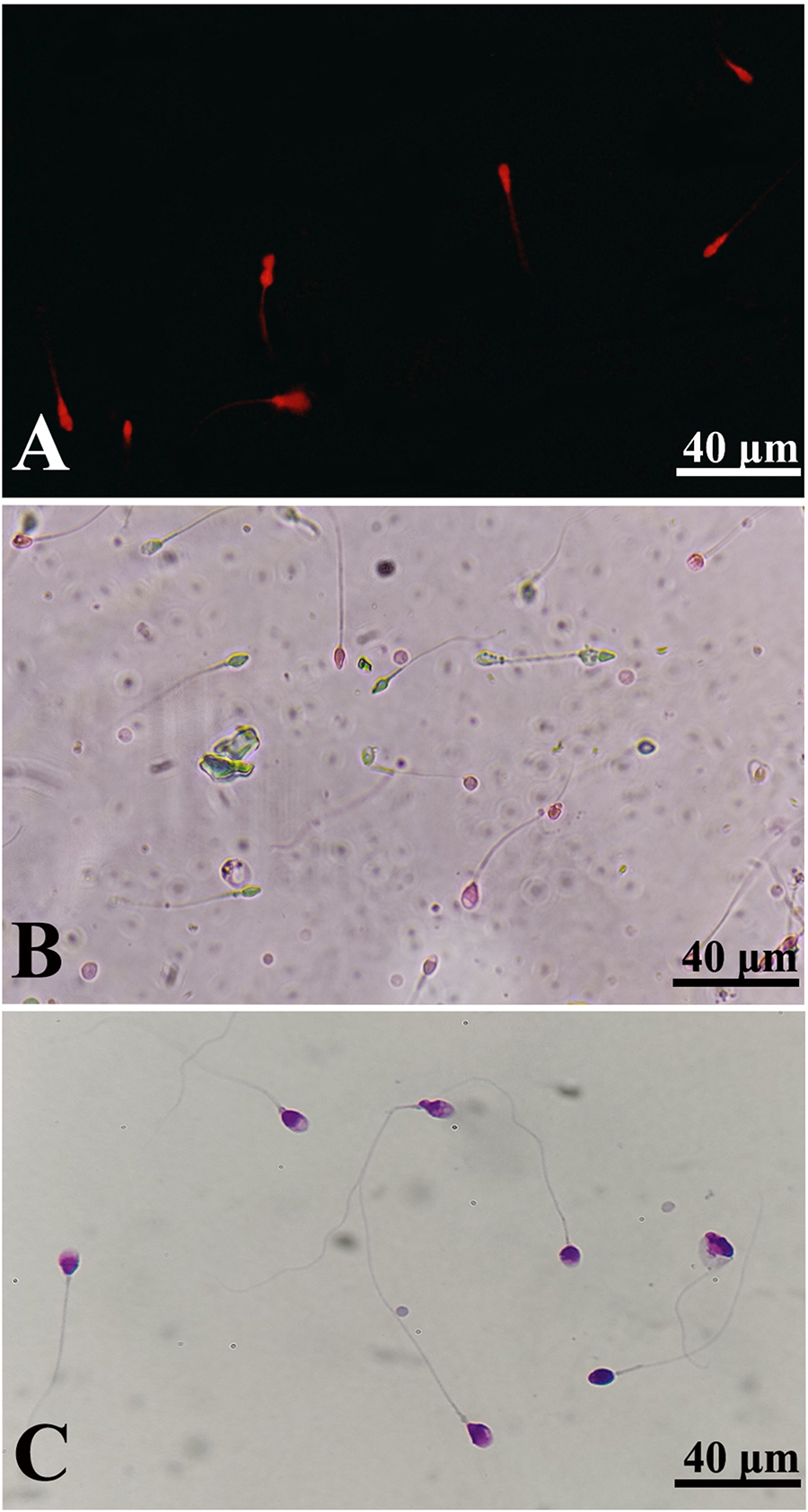
In order to assess the internalization of NLCs into spermatozoa, RH-B-labeled NLCs were incubated with spermatozoa for 30 and 60 minutes. Fluorescence microscopic images showed that approximately 50% of the treated spermatozoa within 30 minutes and more than 95% of the spermatozoa within 60 minutes were positively stained, respectively (Fig. 3A).
Optimal dose of SE for sperm cryopreservation
The results of determining the optimal dose of SE are shown in Table 1. Regarding our findings, among different concentrations of SE on sperm viability, morphology, and motility, SE at a dose of 50 µM was more effective. Therefore, this concentration was selected for further experiments.
Effects of Different Concentrations of Sesamol on the Sperm Parameters

Data are presented as mean ± standard deviation (SD), number of samples (n = 7).
Different letters (a, b) have significant differences (p < 0.05).
One-way analysis of variance (ANOVA) test followed by Tukey’s post hoc test was used to compare the groups.
SE, sesamol.
Sperm analysis
The effects of SE and SE-loaded NLCs administration on sperm total and progressive motility, viability, and morphology parameters are summarized in Table 2. The sperm total and progressive motility and viability were significantly improved in the SE-loaded NLCs group compared with the control, NLCs, and SE groups (p < 0.05) (Fig. 3B). However, no significant improvement was detected in total motility and progressive motility between the SE group and the control and NLCs groups (p > 0.05). According to Diff-Quik staining results, the presence of SE-loaded NLCs or SE in cryomedium had no significant effects on the morphology of spermatozoa in comparison with the control and NLCs groups (p > 0.05) (Fig. 3C).
Comparison of Sperm Parameters Among the Study Groups

Data are presented as mean ± standard deviation (SD), number of samples (n = 20).
Different letters (a, b, c) have significant differences (p < 0.05).
One-way ANOVA test followed by Tukey’s post hoc test was used to compare the groups.
NLCs, nanostructured lipid carriers.
SDF assessment
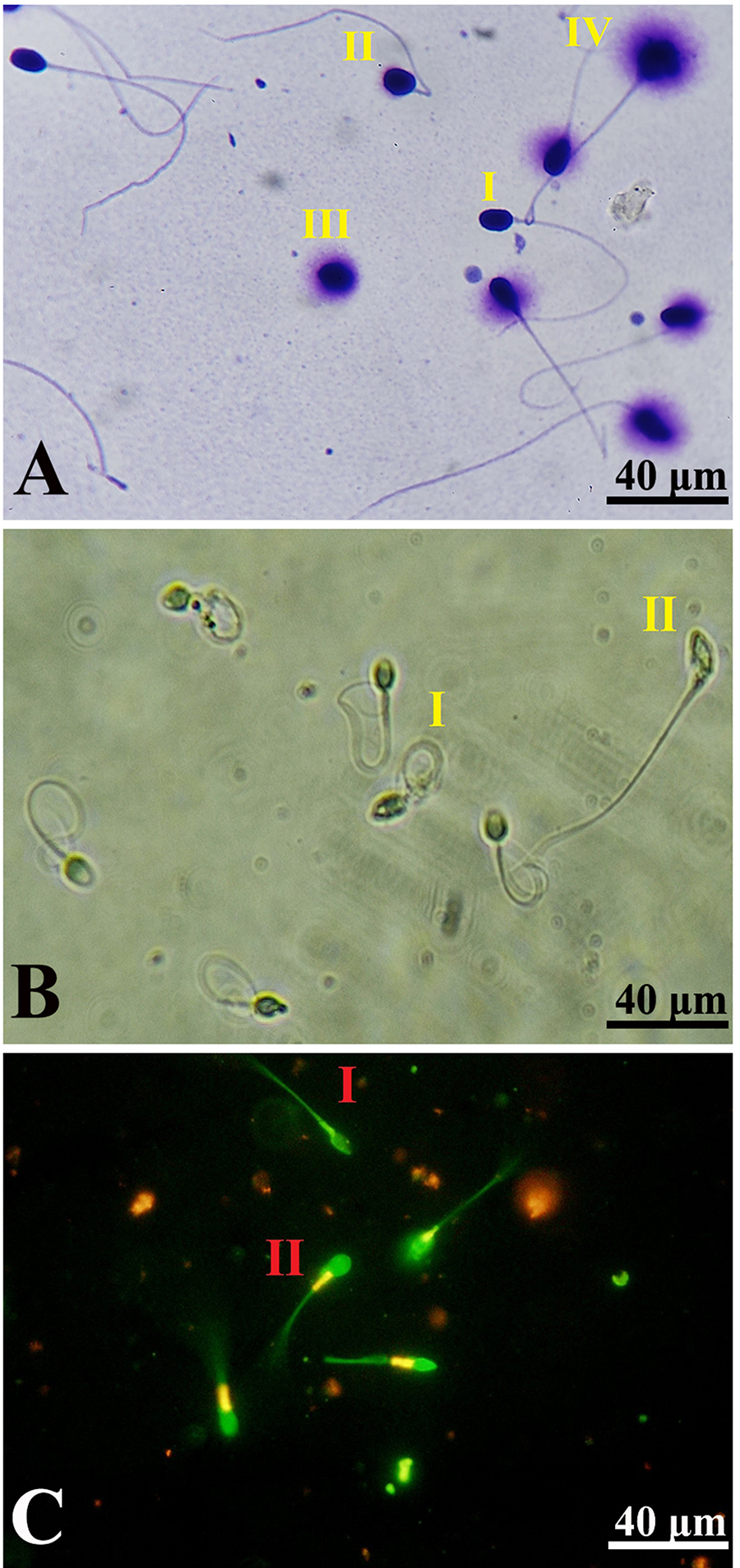
The results of SDF are shown in Figure 4A. Accordingly, cryopreservation induced SDF after thawing in comparison with the fresh group (p < 0.05). No significant difference was detected between the control and NLCs groups (p > 0.05). Also, both SE and SE-loaded NLCs significantly alleviated SDF compared with the control and NLCs groups (p < 0.05). Furthermore, it was found that SE-loaded NLCs led to a significant reduction in SDF compared with the SE group (p < 0.05) (Fig. 5A).

The effect of NLCs, SE, and SE-loaded NLCs on spermatozoa
Sperm membrane integrity
According to Figure 4B, the results of the HOS test showed cryopreservation significantly reduced the spermatozoa membrane integrity compared with the fresh group (p < 0.05). Also, the results revealed membrane integrity increasing in the SE-loaded NLCs group in comparison with the control, SE, and NLCs groups (p < 0.05). Moreover, cryomedium composed with SE had a significant effect on membrane integrity compared with the control and NLCs groups (p < 0.05) (Fig. 5B).
Sperm MMP
Figure 4C shows the outcome of the spermatozoa MMP assessment. The results of JC-1 staining showed that MMP in the control and NLCs groups was significantly decreased compared with the fresh group (p < 0.05). SE-loaded NLCs significantly improved MMP in comparison with control, SE, and NLCs groups (p < 0.05). However, no significant difference was detected between the SE, control, and NLCs groups (p > 0.05) (Fig. 5C).
Sperm acrosome integrity
The results of acrosome integrity are shown in Figure 4D. The results showed that acrosomal integrity was significantly reduced in both the control and NLCs groups compared with the fresh group (p < 0.05). No statistically significant difference was detected between the control and NLCs groups (p > 0.05). Also, supplementation with either SE or SE-loaded NLCs significantly increased acrosomal integrity compared with both the control and NLCs groups (p < 0.05). Furthermore, we found that SE-loaded NLCs led to a significant improvement in acrosome integrity compared with the SE group (p < 0.05).
Discussion
One of the major problems in the sperm freezing procedure is the generation of ROS-induced oxidative stress. 36 ROS can damage the sperm mitochondrial membrane, leading to a decrease in intracellular ATP levels and the release of apoptosis-inducing factors. 13 Additionally, ROS attacks unsaturated fatty acids in the sperm membrane, altering related integrity and permeability. They can also damage DNA, leading to structural disruptions and potentially causing single- or double-strand breaks. 37 Also, it appears that the antioxidant system present in sperm lacks sufficient capacity to neutralize the effects of free radicals during the freezing process. Thus, the application of exogenous antioxidant agents to the sperm or incubating the sperm with these agents prior to freezing seems to be an effective approach for mitigating the adverse effects of ROS. 38 According to several studies, SE possesses strong free radical-scavenging activity.39,40 Despite these capabilities, SE has poor water solubility, which limits the associated applications in biological systems. 41 Thus, the present study aimed to evaluate the effects of SE-loaded NLCs, as a drug delivery system, on the performance of SE during sperm cryopreservation.
In the present study, the physicochemical characterization of the prepared NLCs, including particle size, zeta potential, and high EE, confirms the suitability of the preparation method. The produced NPs were smaller than 200 nm, enabling efficient cellular uptake and avoiding clearance by the reticuloendothelial system.42,43 Additionally, the slightly negative zeta potential enhances nanoparticle stability and prevents detachment from the sperm membrane surface, thereby supporting effective nanoparticle internalization, as validated by uptake experiments. 44 Generally, EE values exceeding 60% are regarded as high and in the optimal range for efficient drug delivery, as they maximize the drug quantity delivered to the target site. 45 Hence, the EE of 72% in this study was considered satisfactory.
Loading SE into nanocarriers such as NLC allows for a more complete manifestation of the associated antioxidant and anti-inflammatory effects by modification of pharmacokinetic and pharmacodynamic properties, increasing stability, 46 facilitating cellular accessibility, 47 and controlled release, 48 thereby enhancing the protection of sperm and testicular tissue. Free SE is unable to maintain therapeutically effective concentrations in target tissues, such as the testis, owing to its short half-life, rapid systemic clearance, and susceptibility to autoxidation; in contrast, NLCs, composed of a combination of solid and liquid lipids, enable high drug-loading efficiency (>90% entrapment) and provide a sustained release profile. 46 In ischemic stroke models, 47 the SE-loaded NLCs represent greater penetration of biological barriers, higher tissue accumulation, and more sustained protection (reduced Malondialdehyde (MDA), increased Glutathione (GSH)/Superoxide Dismutase (SOD)/Catalase (CAT) than the free form. This rationale may be extended to the testis, where oxidative stress and inflammation adversely affect sperm DNA and mitochondrial function. According to the nanometer size and lipophilic surface, NLC enhances the passage of biological barriers, facilitates cellular entry via endocytosis, and provides a controlled release by creating a lipid reservoir that prolongs the duration of active Nrf2, HO-1/NQO1, and antioxidant enzymes. 48 Passive targeting in inflamed testicular tissues and even active targeting (such as mannose-NLC) enables selective accumulation and limits inflammation by more potent inhibition of NF-κB (reduction of TNF-α/IL-1β/COX-2) and activation of PI3K/Akt (increase of Bcl-2, decrease of Bax/caspase-3). 46 At the molecular level, SE activates Nrf2 and inhibits NF-κB, 48 but the nanocarrier with a stable intracellular concentration potentiates these effects and regulates the oxidant-antioxidant balance in favor of ROS reduction, mitochondrial protection, and improved sperm motility. 49 Sesamin (similar to SE) data suggest that reducing oxidative stress normalizes the RNF8-ubH2A/ubH2B pathway for histone–protamine exchange and reduces sperm DNA damage, while PI3K/Akt enhances germ and Sertoli cell survival and improves tubular epithelial thickness. 49
Our results demonstrated that SE significantly improves several critical sperm quality parameters following cryopreservation, including viability, sperm membrane integrity, SDF, MMP, and acrosome integrity. However, SE treatment provided no significant improvements in total motility, progressive motility, or sperm morphology. Notably, when SE was delivered via NLCs, there was a significant improvement across all assessed sperm parameters except morphology, indicating that encapsulation within NLCs enhances the bioavailability and protective effects of SE during the cryopreservation process. These findings align with the study by Ali et al., 38 who reported that canthaxanthin, a carotenoid with low water solubility, similarly improved sperm viability and reduced DNA fragmentation with no significant effect on total or progressive motility or morphology during sperm cryopreservation. This parallel suggests that the limited solubility of antioxidant compounds like SE and canthaxanthin may restrict the efficacy in certain sperm parameters unless delivered through advanced nanocarrier systems, which improve cellular uptake and stability. The application of nanocarriers represents a promising strategy to overcome solubility barriers, enhancing the antioxidant’s protective capacity against cryoinjury by preserving mitochondrial function, membrane integrity, and DNA stability, which are essential for maintaining sperm fertilizing potential.50,51 Notably, our study revealed that the group receiving SE-loaded NLCs demonstrated substantial improvements in sperm vitality, MMP, acrosome integrity, and SDF when compared with those only treated with SE. This pronounced improvement highlights the superior efficiency of NLCs as a drug delivery system, providing controlled and sustained release, facilitating enhanced uptake, and protecting SE from degradation during the cryopreservation process. Mitochondrial integrity and intact MMP are prerequisites for sperm motility. 52 By mitigating oxidative damage, SE preserves mitochondrial integrity, which is critical because intact MMP is essential for ATP production and, consequently, sperm motility and overall functionality. Moreover, the enhanced SE delivery via NLCs probably improves bioavailability and cellular internalization, thereby amplifying the protective effects on mitochondrial function and DNA integrity. Maintaining mitochondrial health through preserved MMP supports energy metabolism and regulation of apoptotic pathways, which collectively contribute to improved sperm vitality and fertilization potential. 52 These findings underscore the importance of advanced nanocarrier systems in optimizing antioxidant therapies aimed at reducing cryoinjury and enhancing fertility preservation outcomes. 52
Recently, studies showed that some antioxidants in different doses had beneficial effects on sperm parameters. Mohammadi-Bardbori et al. 31 demonstrated that incubation of normozoospermic sperm samples with 10 µM astaxanthin, a lipid-soluble carotenoid, resulted in a significant increase in total and progressive motility, viability, and DNA integrity compared with the control group. On the other hand, Ghantabpour et al. 53 and Dede et al. 54 reported that astaxanthin at concentrations of 1 µM and 100 µM, respectively, improved post-cryopreservation sperm quality through effective attenuation of oxidative stress. These differences in effective concentrations may be attributed to the requirement for sufficient antioxidant availability in the sperm microenvironment to facilitate membrane penetration; however, excessive concentrations may adversely affect the cryopreservation process. However, the composition of the freezing medium, including potential synergistic interactions, should not be overlooked. Variations in commercial or laboratory-prepared cryopreservation media across different studies may result in differing constituent profiles, which can influence the efficacy of antioxidant supplementation. Consequently, in formulations exhibiting synergistic effects, lower concentrations of an antioxidant may yield optimal outcomes, whereas in the absence of such interactions, substantially higher doses of the same antioxidant may be required to achieve comparable results. Accordingly, to address this limitation, the present study employed a drug delivery system to facilitate and regulate the intracellular delivery of SE into spermatozoa, thereby achieving enhanced and effective penetration.
High doses of SE (100 µM), despite its antioxidant effects at low concentrations, can paradoxically induce pro-oxidant effects, leading to oxidative imbalance, cytotoxicity, and disruption of sperm membrane integrity, a phenomenon based on the dual behavior of polyphenols and the high sensitivity of sperm to oxidative stress. 55 Polyphenols such as SE, at low concentrations, act as ROS scavengers and neutralize hydroxyl, peroxyl, and superoxide radicals, but at high doses, they themselves become sources of ROS; the main mechanism is the autoxidation of phenols, which generates stable phenoxy radicals, which then react with O2 to release H2O2 and superoxide (O−2), which initiates a chain of lipid peroxidation. 56 Pulse radiolysis studies have shown that SE is primarily antioxidant at concentrations of 5–1000 nmol, but at higher levels, pro-oxidant activity predominates, inducing DNA cleavage and lipid peroxidation. In a study on human platelets, 57 SE ≥0.25 mM induced intracellular ROS production (especially H2O2), reduced thiols, impaired mitochondrial potential, caspase activation, cytochrome c translocation, and phosphatidylserine exposure, and induced mitochondrial-mediated apoptosis, which directly accounts for cytotoxicity. These effects were absent at therapeutic concentrations (≤100 µM), and SE even inhibited platelet aggregation, highlighting its narrow therapeutic window. Sperm is highly susceptible to lipid peroxidation due to its membrane rich in polyunsaturated fatty acids (PUFAs); excess ROS detach hydrogen from methylene PUFAs, generating lipid radicals, then lipid peroxyl radicals, and hydroperoxides (such as MDA and 4-HNE) that disrupt membrane fluidity, integrity, and motility. At high doses of SE, pro-oxidant activity increases MDA (a marker of peroxidation) and induces oxidative imbalance, leading to DNA fragmentation, protein oxidation, and germ cell apoptosis. 10 High doses of SE depolarize ΔΨm, release cytochrome c, and activate caspases, which in sperm destroy motility and viability; increased [Ca2+]i of ER/SR activates calpain and phospholipase A2, further stimulating peroxidation; saturation of GSH peroxidase and SOD accumulates H2O2 and facilitates the Fenton reaction (Fe2+ + H2O2 → OH−), which exacerbates cytotoxicity; disruption of protamine-histone exchange, defective chromatin condensation, and DNA integrity, which causes malformation and infertility. 58 Thus, the deleterious effects of 100 µM SE on sperm parameters are due to a shift to pro-oxidant mode, ROS/H2O2 production, PUFA-rich membrane lipid peroxidation, mitochondrial apoptosis, and saturation of cellular defenses, which are exacerbated in sensitive testicular tissue (with low oxygen tension and high PUFA); dose–response studies (5–1000 nmol) show a safe window of ≤50–100 µM, and higher doses are toxic.
Maintaining the membrane integrity of the sperm is important for cell survival. 59 The findings of the present study indicate that the incorporation of SE into the cryopreservation medium significantly enhances antioxidant capacity and contributes to the protection of the sperm membrane from damage. However, to prevent disruption of the freezing environment while ensuring optimal SE delivery, SE-loaded NLCs were employed to facilitate targeted and controlled administration of the compound. This strategy preserved the optimal equilibrium of the cryopreservation medium while ensuring that SE was delivered to spermatozoa at an effective concentration. By enhancing intracellular antioxidant capacity, the defense against ROS was markedly strengthened, as evidenced by the favorable effects observed with SE-loaded NLCs. This targeted delivery also contributed to the preservation of acrosomal structure. SE-loaded NLC-treated groups exhibited superior acrosome integrity compared with other groups, suggesting that the mechanisms safeguarding the sperm membrane similarly protect acrosomal integrity.
Conclusion
This study demonstrated that SE-loaded NLCs significantly improve multiple parameters of sperm quality following cryopreservation, including motility, viability, membrane and acrosome integrity, MMP, and reduction of DNA fragmentation. The successful synthesis of SE-loaded NLCs with optimal particle size, stability, and high EE facilitated efficient uptake by spermatozoa, enhancing the antioxidant delivery compared with free SE. These findings indicate that SE-loaded NLCs provide a superior protective effect against oxidative stress-induced cryoinjury, thereby preserving sperm functional and structural integrity more effectively during the freeze-thaw process. Consequently, SE-loaded NLCs represent a promising and innovative antioxidant strategy to mitigate cryodamage and improve outcomes in fertility preservation protocols. Further research and clinical validation may establish their potential application in assisted reproductive technologies.
Limitations and Future Directions
This study was subject to several limitations. The relatively small sample size restricts the generalizability of the findings; consequently, future research should involve larger cohorts to validate and extend the observed effects of SE-loaded NLCs during sperm cryopreservation and thawing. Furthermore, the present study exclusively examined semen samples from normozoospermic individuals; inclusion of abnormal samples in subsequent research would provide a more comprehensive assessment of the SE-loaded NLCs’ protective efficacy across diverse semen quality profiles. Another important limitation is that the present study focused exclusively on post-thaw sperm quality parameters (motility, viability, morphology, etc.), while the impact on functional fertility outcomes, particularly fertilization and embryo development rates in IVF/ICSI procedures, was not assessed. Given that the ultimate objective of such interventions is to enhance clinical fertility and achieve successful pregnancies, subsequent studies should prioritize evaluating the effects of SE-loaded NLCs on reproductive success in assisted reproductive technology settings.
Authors’ Contributions
V.S.-A.: Writing—review and editing, writing—original draft, methodology, investigation, software, formal analysis, project administration, and data curation. H.H.: Validation, software, methodology, formal analysis, and data curation. M.P.: Methodology and validation. H.H.: methodology, validation. R.A.: Methodology, investigation. N.N.: Validation. A.F.: Writing—review and editing, writing—original draft, visualization, validation, supervision, project administration, methodology, funding acquisition, formal analysis, and conceptualization. All authors confirm that their research is supported by an institution that is primarily involved in education and/or research.
Footnotes
Data Availability
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
This study has been extracted from the thesis of Vahid Shokri-Asl, registered at the Department of Reproductive Biology, Faculty of Advanced Medical Sciences, Tabriz University of Medical Sciences (Thesis No. 71244, IR.TBZMED.REC.1402.243).
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This study was supported by the