
Abstract
Objective
Exosomes derived from bone marrow mesenchymal stem cells (BMSC-Exos) may modulate the M1/M2 polarization of macrophages during osteoarthritis (OA). However, the underlying mechanisms of BMSC-Exos in this process still need to be elucidated. In this study, we explored the role of BMSC-Exos in the polarization of macrophages in vitro and the OA rats in vivo.
Methods
The effects of BMSC-Exos on RAW264.7 cells were determined, including the production of reactive oxygen species (ROS) and the protein expression of Akt, PINK1, and Parkin. We prepared an OA model by resecting the anterior cruciate ligament and medial meniscus of Sprague–Dawley (SD) rats. Hematoxylin-eosin (H&E) and safranin O-fast green staining, immunohistochemistry and immunofluorescence analyses, and the examination of interleukin 6 (IL-6), interleukin 1β (IL-1β), tumor necrosis factor alpha (TNF-α), and interleukin 10 (IL-10) were performed to assess changes in cartilage and synovium.
Results
BMSC-Exos inhibited mitochondrial membrane damage, ROS production, and the protein expression of PINK1 and Parkin. Akt phosphorylation was downregulated under lipopolysaccharide (LPS) induction but significantly recovered after treatment with BMSC-Exos. BMSC-Exos alleviated cartilage damage, inhibited M1 polarization, and promoted M2 polarization in the synovium in OA rats. The expression of PINK1 and Parkin in the synovium and the levels of IL-6, IL-1β, and TNF-α in the serum decreased, but the level of IL-10 increased when BMSC-Exos were used in OA rats.
Conclusion
BMSC-Exos ameliorate OA development by regulating synovial macrophage polarization, and one of the underlying mechanisms may be through inhibiting PINK1/Parkin signaling.
Introduction
Osteoarthritis (OA) is the most common chronic degenerative joint disease worldwide, characterized by joint pain, swelling, and limited movement, sometimes accompanied by joint stiffness. 1 The pathological changes associated with OA are primarily the degeneration of articular cartilage, changes in the subchondral bone, and macrophage activation. 2 Previously, OA was believed to be a non-inflammatory joint disease. However, increasing evidence has demonstrated the importance of inflammation in OA pathogenesis. 3 The main treatment methods for OA are drug therapy and surgical operation. The main purpose of drug therapy is to relieve pain and control inflammation to improve the quality of the patients’ daily life. However, the long-term effect is limited, and patients with advanced OA require joint replacement or prosthetics. 4 The high rate of morbidity and disability imposes a substantial burden on families and society. Therefore, it is imperative to explore new treatment methods for OA.
Bone marrow mesenchymal stem cells (BMSCs) are pluripotent, non-hematopoietic stem cells capable of tissue repair and multidirectional differentiation. 5 Furthermore, BMSCs have anti-inflammatory and immunomodulatory effects. 6 Therefore, stem cell transplantation is a promising treatment strategy, but risk factors, such as immune rejection and tumorigenesis, complicate the clinical application of BMSCs. 7 Recent studies have shown that stem cells can secrete extracellular vesicles (EVs) that repair damaged tissues and promote cell regeneration. 8 Exosomes with a diameter of 40-120 nm are a type of EVs. 9 Immune-tolerant exosomes have similar biological roles as BMSCs. 10 Thus, many researchers have considered exosomes derived from BMSCs (BMSC-Exos) as a substitute for BMSCs in treating several diseases, including OA. We as well as other groups have demonstrated that BMSC-Exos could help restore the intra-articular environment’s stability and regulate inflammation and play a key role in OA. 11 Macrophages exhibit different phenotypes in response to different stimulating factors; M1 and M2 macrophages are the two extreme phenotypes. 12 M1 macrophages (classically activated macrophages) stimulated by lipopolysaccharide (LPS), interferon gamma (IFN-γ), and tumor necrosis factor alpha (TNF-α) can secrete a large number of inflammatory factors and chemokines, including interleukin 1β (IL-1β), interleukin 6 (IL-6), interleukin 23 (IL-23), TNF-α, inducible Nitric Oxide Synthase (iNOS), and reactive oxygen species (ROS), which participate in killing pathogenic microorganisms and can simultaneously cause tissue damage. 13 IL-4, IL-10, and IL-13 activate M2 macrophages (selectively activated macrophages); these cells secrete anti-inflammatory factors, such as IL-10 and TGF-β, and exert anti-inflammatory and immunosuppressive effects. 14 M2 macrophages promote tissue repair and regeneration but may lead to disease remission. Macrophage polarization from M2 to M1 is a pathophysiological process in OA. 11 Emerging evidence suggests that promoting M2 polarization or inhibiting M1 polarization may be a new therapeutic approach for OA. 15 Studies have shown that exosomes may promote M2 polarization, but the exact mechanisms are unclear.
Elevated ROS production causes chondrocyte damage and apoptosis and is strongly associated with OA. 16 In addition to this, ROS can interact with macrophages, promoting the progression of the inflammatory response in OA. 17 Studies have shown that activating Akt changes the level of ROS, which may be involved in the subsequent PINK1/Parkin signaling pathway. 18 The PINK1/Parkin signaling pathway plays an important role in mitochondrial damage and deterioration of the oxidative state. 19 However, there are few reports of the association between the PINK1/Parkin signaling pathway and BMSC-Exos in macrophage polarization in OA.
In this study, we prepared BMSC-Exos to treat an OA rat model induced by destabilization of the anterior cruciate ligament and medial meniscus. More synovial macrophages transformed from the M1 polarization phenotype to the M2 polarization phenotype after treatment with BMSC-Exos, and cartilage damage was relieved. We also examined the effects of BMSC-Exos in vitro and demonstrated that BMSC-Exos promoted macrophage polarization from M1 to M2 and that PINK1/Parkin signaling might be involved. Collectively, our results support the use of BMSC-Exos as a complementary treatment for OA due to their ability to regulate macrophage polarization.
Materials and Methods
Extraction and Identification of BMSCs
The femurs and tibias of 2-week-old Sprague–Dawley (SD) rats were removed as previously described. 20 BMSCs were flushed from the femurs and tibias with alpha minimal essential medium (α-MEM) (HyClone, Logan, UT) containing 10% fetal bovine serum (FBS) (Gibco, CA) and 1% penicillin/streptomycin (HyClone, Logan, UT). After density gradient centrifugation, the cells were incubated at 37 °C with 5% CO2. The medium was changed every 2 days. When the adherent cells reached 80%-90% confluence, they were digested with 0.25% trypsin (Gibco, CA, USA) and subcultured at a 1:2 ratio.
The morphology of the BMSCs was observed with an inverted microscope (Leica, Wetzlar, Germany), and the expression of surface markers CD29, CD34, CD45, and CD90 (BD Biosciences, San Jose, CA) was analyzed by flow cytometry. After induction with osteogenesis, lipid formation, or cartilage induction medium (Cyagen Biosciences Inc, Santa Clara, CA), the cells were stained with Alizarin red (ARS), oil red O, or Alcian blue, respectively, to evaluate the osteogenic, adipogenic, and chondrogenic potential of BMSCs under an inverted microscope.
Preparation and Identification of BMSC-Exos
BMSC-Exos were extracted by ultracentrifugation. BMSCs were cultured in FBS without exosomes for 48 hours. The culture medium was centrifuged at 10,000 g for 30 minutes to remove dead cells, cell debris, and other pellet materials, and the supernatant was collected. Then, ultracentrifugation was performed at 100,000 g for 120 minutes (HITACHI Limited, Tokyo, Japan), and the pellet was collected. Before the second ultracentrifugation at 100,000 g for 120 minutes, a 0.22-μm sterile filter (Millipore, Boston, MA) was used to remove larger EVs. Finally, BMSC-Exos were carefully re-suspended in phosphate-buffered saline (PBS; Solarbio, Beijing, China) and stored at −80 °C for later use. The concentration of BMSC-Exos was determined by the bicinchoninic acid assay (BCA) (Beyotime, Shanghai, China) method. The morphology of the exosomes was assessed via transmission electron microscopy (TEM; HITACHI Limited, Tokyo, Japan), and the exosome size distribution was analyzed by a nanoparticle sizer (Malvern, Malvern, UK). Western blotting was used to verify the expression levels of surface marker proteins CD81 (Santa, USA), CD63 (Santa, Cruz, CA, USA), and TSG101 (Abcam, Cambridge, MA).
Exosome Uptake
Exosomes were labeled with the red fluorescent dye Dil (Sigma–Aldrich, St. Louis, MO) according to the manufacturer’s instructions. Unbound Dil dye was removed by ultracentrifugation for 60 minutes. RAW264.7 cells were kindly provided by Prof. Liu, Shandong First Medical University. RAW264.7 cells were incubated with Dil-labeled exosomes at 37 °C for 2 hours. After washing with PBS, 4% paraformaldehyde (PFA) was used for fixation at room temperature for 15 minutes. Then, phalloidin (Invitrogen, Waltham, MA) was used to stain the cytoskeleton, and Hoechst 33342 (Beyotime, Shanghai, China) was used to stain the nuclei. Exosome uptake was observed via confocal fluorescence microscopy (Olympus, Tokyo, Japan).
Cell Proliferation Test
RAW264.7 cells were seeded into 96-well plates at a density of 5,000 cells/well. Then, the cells were treated with exosomes at 0, 20, 40, 60, 80, and 100 μg/mL. After culturing for 24 hours, 10 μL of Cell Counting Kit-8 (CCK-8; APExBIO, Houston, TX) solution was added to each well, and the absorbance values were detected at a wavelength of 450 nm (OD450) using a microplate reader (BioTek, Winooski, VT).
Detection of ROS
RAW264.7 cells were seeded into 6-well plates at a density of 2 × 105 cells/well and stimulated with 10 μg/mL LPS (Solarbio, Beijing, China) for 48 hours. Then, the cells were treated with 100 μg/mL exosomes for 24 hours. After digestion and centrifugation, 2′,7′-Dichlorodihydrofluorescein diacetate (DCFH-DA) was added, and the samples were incubated at 37 °C for 20 minutes. The fluorescence intensity was measured by flow cytometry.
Mitochondrial Membrane Potential Assay
Mitochondrial membrane potential was determined with a mitochondrial membrane potential detection kit (JC-1; Beyotime, Shanghai, China). RAW264.7 cells were plated in confocal dishes, stimulated with 10 μg/mL LPS for 48 hours, and then 100 μg/mL exosomes were added. After fixation with 4% PFA, the cells were incubated with the working solution for 20 minutes, and the results were observed and photographed using a confocal fluorescence microscope (Olympus, Tokyo, Japan). Green fluorescence and red fluorescence represented JC-1-monomers and JC-1-aggregates, respectively.
Western Blot Analysis
The procedures for Western blot were described in our previous reports. 20 Cells or exosomes were lysed in RIPA buffer (Beyotime, Shanghai, China) containing protease inhibitors. The protein concentration was quantified using a BCA protein assay kit (Beyotime, Shanghai, China). The protein (30 μg) was subjected to SDS-PAGE and transferred onto a polyvinylidene fluoride (PVDF) membrane (Whatman, Maidstone, Kent, UK). The following primary antibodies (CST, Danvers, MA) were applied: Arg-1 (1:1000), CD206 (1:1000), CD86 (1:1000), AKT (1:1000), P-AKT (1:1000), iNOS (1:1000), PINK1 (1:1000), and Parkin (1:1000). Images were captured using a chemiluminescent imaging system (GE, Boston, USA).
Cell Fluorescence Analysis
RAW264.7 cells were plated into a confocal dish and stimulated with 10 μg/mL LPS for 48 hours. Then, 100 μg/mL exosomes were added. After fixation with 4% PFA, 0.1% TritonX-100 was added (Solarbio, Beijing, China). Cells were incubated overnight with iNOS (1:500) or CD206 (1:50) primary antibodies at 4 °C, and the nuclei were stained with DAPI (Invitrogen, Waltham, MA). The fluorescence-labeled secondary antibodies (Invitrogen, Waltham, MA) were added, and immunoreactivity was recorded via confocal fluorescence microscopy (Olympus, Tokyo, Japan).
Animal Model
The animal experiments were implemented with written confirmation authorized by the Animal Research Ethics Committee of Jinzhou Medical University (2022025). Twenty-four male SD rats (10 weeks old, weighing approximately 220 g) were housed at a 12/12-hour light/dark cycle, with a controlled temperature (23 °C-25 °C) and steady humidity (40%-60%) in the animal center of Jinzhou Medical University (Jinzhou, Liaoning, China). Random number tables were used to allocate 4 groups, with 6 rats in each group. All the rats were anesthetized with isoflurane inhalation through a facemask and randomly divided into 4 groups (6 rats per group), including the Sham group, OA group, BMSCs group, and BMSC-Exos group. The articular cavity of the knee joint was cut open, but the anterior cruciate ligament and medial meniscus were left intact in the Sham group. The anterior cruciate ligament and medial meniscus were excised to establish an animal model of OA 21 in other groups. Four weeks after surgery, the rats in the Sham group received no treatment; those in the OA group, BMSCs group, and BMSC-Exos group received 50 μL of PBS, BMSCs (2 × 106/50 μL), and BMSC-Exos (100 μg/50 μL) by intra-articular injection, respectively. When weight loss of more than 20%, not eating for more than 2 days, or other obvious sufferings were found, the rats would be excluded from the study, and euthanasia (three times the amount of anesthetic) would be implemented. No obvious sufferings happened during our experiment, so after treatment for 6 weeks, all rats were sacrificed for the following examination.
Histological Analysis
Six weeks after the treatment, the rats in each group were killed, and their knee joints and synovium were collected and fixed in 4% PFA overnight. The knee joint samples were decalcified, paraffin-embedded, dehydrated, and longitudinally sectioned at 5 μm. The decalcification procedure was omitted for the synovium samples. Sections were stained with hematoxylin-eosin (H&E) or safranin O-fast green. The results of safranin O-fast green staining were evaluated according to the International Association for the Study of Osteoarthritis (OARSI) scoring system. 22
Immunohistochemistry and Immunofluorescence Analysis
After dewaxing and antigen repair, the sections of the synovium were blocked with 3% bovine serum albumin (BSA) and incubated overnight with primary antibodies (Arg-1, PINK1, and Parkin; CST, Danvers, MA) at 4 °C. Then, the sections were incubated with horseradish peroxidase-labeled secondary antibodies (Abcam, Cambridge, MA) at room temperature. Frozen sections were incubated with a primary antibody against iNOS (Abcam, Cambridge, MA) overnight at 4 °C; the sections were then washed and incubated with a fluorescence-labeled secondary antibody. After staining nuclei with DAPI, the sections were visualized by fluorescence microscopy (Olympus, Tokyo, Japan).
Cytokine Determination
Blood was collected from the heart of the rats at the end of the experiment. The serum was separated by centrifugation. RAW264.7 cell culture supernatant was also collected and centrifuged. The levels of IL-6, IL-1β, TNF-α, and IL-10 in the serum and supernatant were examined by enzyme-linked immunosorbent assay (ELISA) as per the manufacturer’s instructions (Beyotime, Shanghai, China). The serum or supernatant was added into the well at 100 μL/well, and the reaction plate was sealed with a sealing membrane and incubated at room temperature for 120 minutes. Then, biotinylated antibody, horseradish peroxidase-labeled streptavidin, and 3, 3′,5,5′-tetramethylbenzidine (TMB) solution were added, and the plate was incubated at room temperature in the dark. Finally, 50 μL/well of termination liquid was added, and the OD450 was determined using a microplate reader (BioTek, Winooski, VT).
Statistical Analysis
All the experiments were repeated at least 3 times. The data are expressed as the means ± standard derivation and were analyzed with GraphPad Prism 8.0 (GraphPad Software, La Jolla, CA). Student’s t-test was used to compare the differences between the two groups. One-way analysis of variance (ANOVA) was used to evaluate differences among 3 or more groups, and Tukey’s test was performed for the subsequent between-group differences. In the current study, statistical significance was set at P < 0.05.
Results
Identification of BMSCs and BMSC-Exos
Most BMSCs showed a spindle-like shape under an inverted phase contrast microscope (
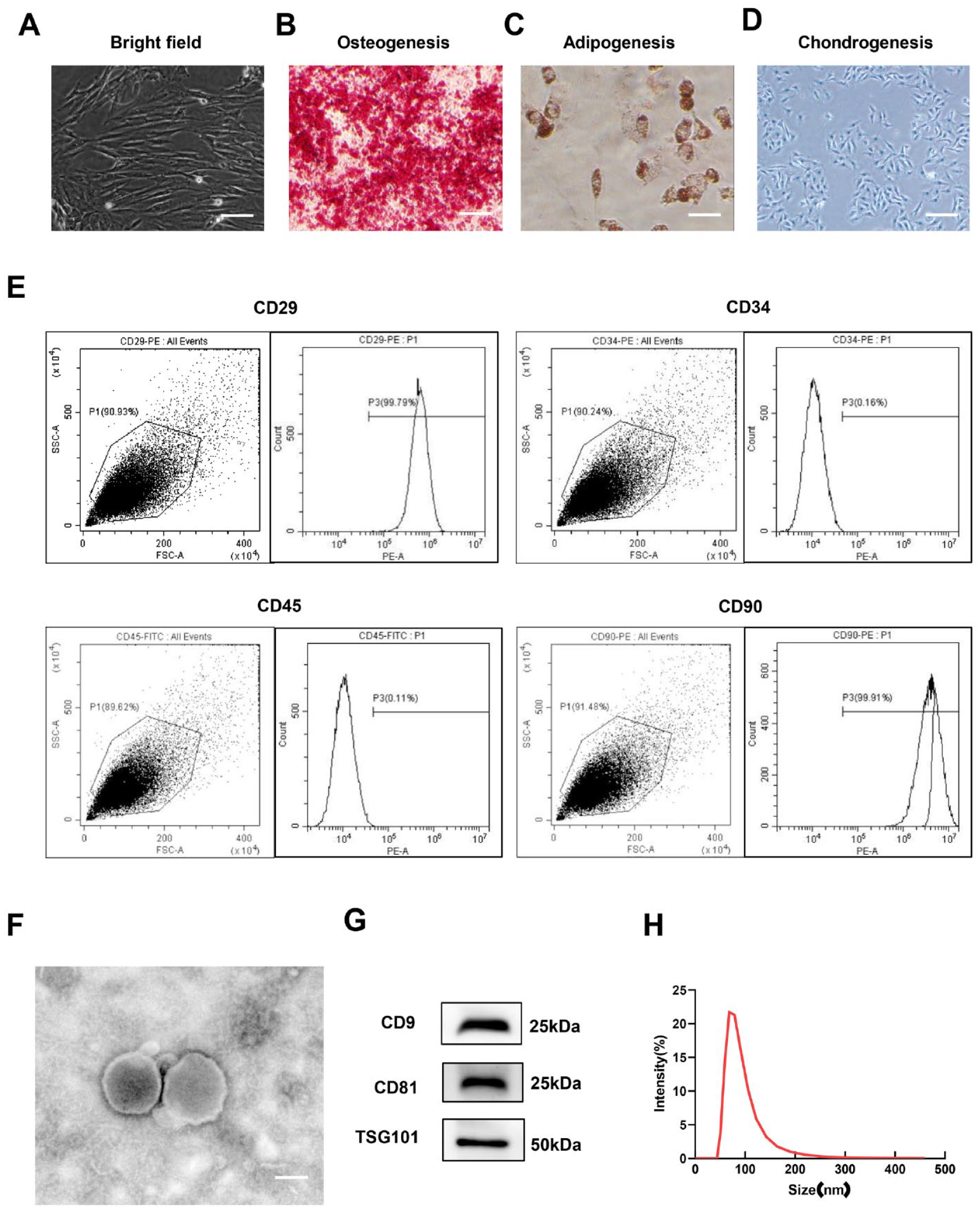
Characterization of BMSCs and BMSC-Exos. (
The hollow spherical microvesicles were observed via TEM (
BMSC-Exos Promoted the M2 Polarization of Macrophages and Reduced Inflammation In Vitro
Exosome uptake is the first step required for their participation in any physiological process.
5
After incubating RAW264.7 cells with Dil-labeled BMSC-Exos (Dil-Exos) for 24 hours, Dil-Exos were observed in the cytosol via confocal microscopy (
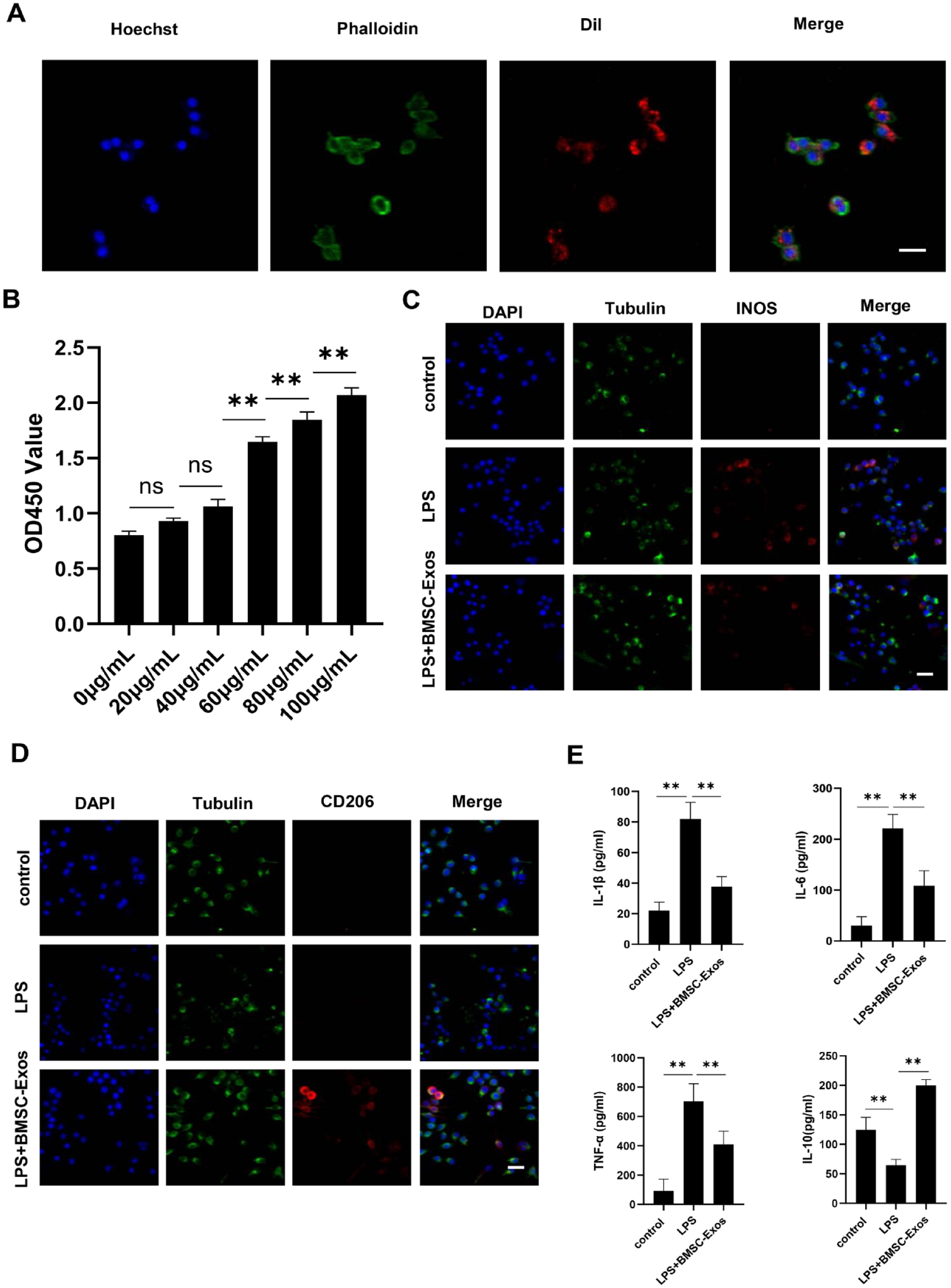
BMSC-Exos promoted M2 polarization of macrophages and reduced inflammation in vitro. (
Macrophages exhibit two phenotypes, M1 and M2. M1 macrophages are pro-inflammatory, while M2 macrophages are anti-inflammatory. After co-culturing with BMSC-Exos, the immunofluorescence expression level of iNOS, the marker of M1 macrophages, significantly decreased (
Involvement of PINK1/Parkin Signaling in the Effect of BMSC-Exos on Macrophage Polarization
Consistent with other reports,
23
the elevated production of ROS by LPS was reduced to a lower level by BMSC-Exos (P < 0.01,
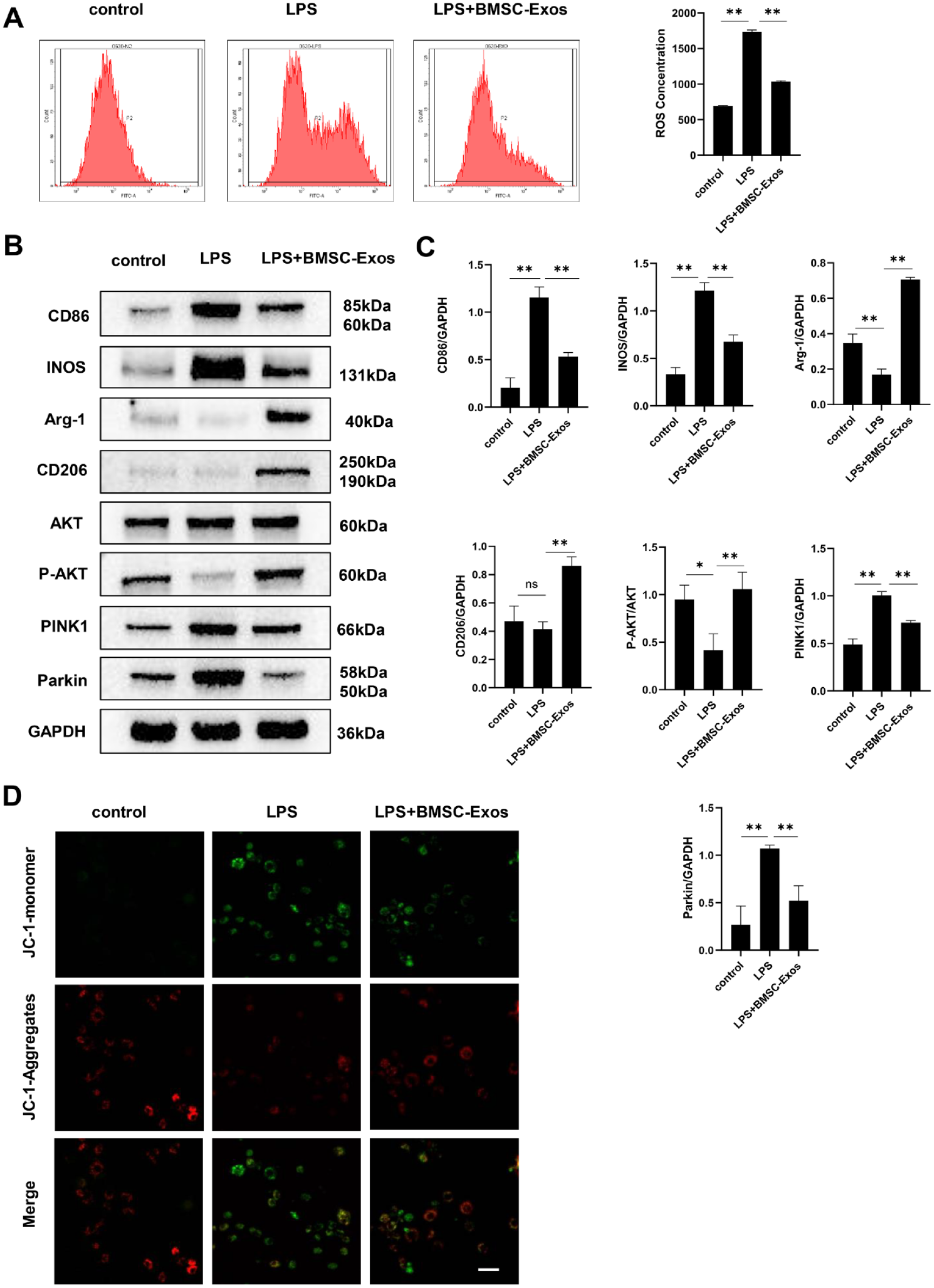
Involvement of PINK1/Parkin signaling in the effect of BMSC-Exos on macrophages. (
The Western blot results showed that after stimulation with LPS for 24 hours, the protein expression levels of CD86 and iNOS (M1 markers) significantly increased, while the protein expression levels of Arg-1 and CD206 (M2 markers) were significantly decreased (P < 0.01,
BMSC-Exos Relieved OA in Rats
A rat model induced by anterior cruciate ligament and medial meniscus was used in our present study to assess the effects of BMSC-Exos in vivo. The cartilage injury was evaluated by H&E and safranine O-fast green staining 10 weeks after the surgery. In the OA group, the cartilage surface was rougher than that in the Sham group, and the integrity of the tide line was destroyed (
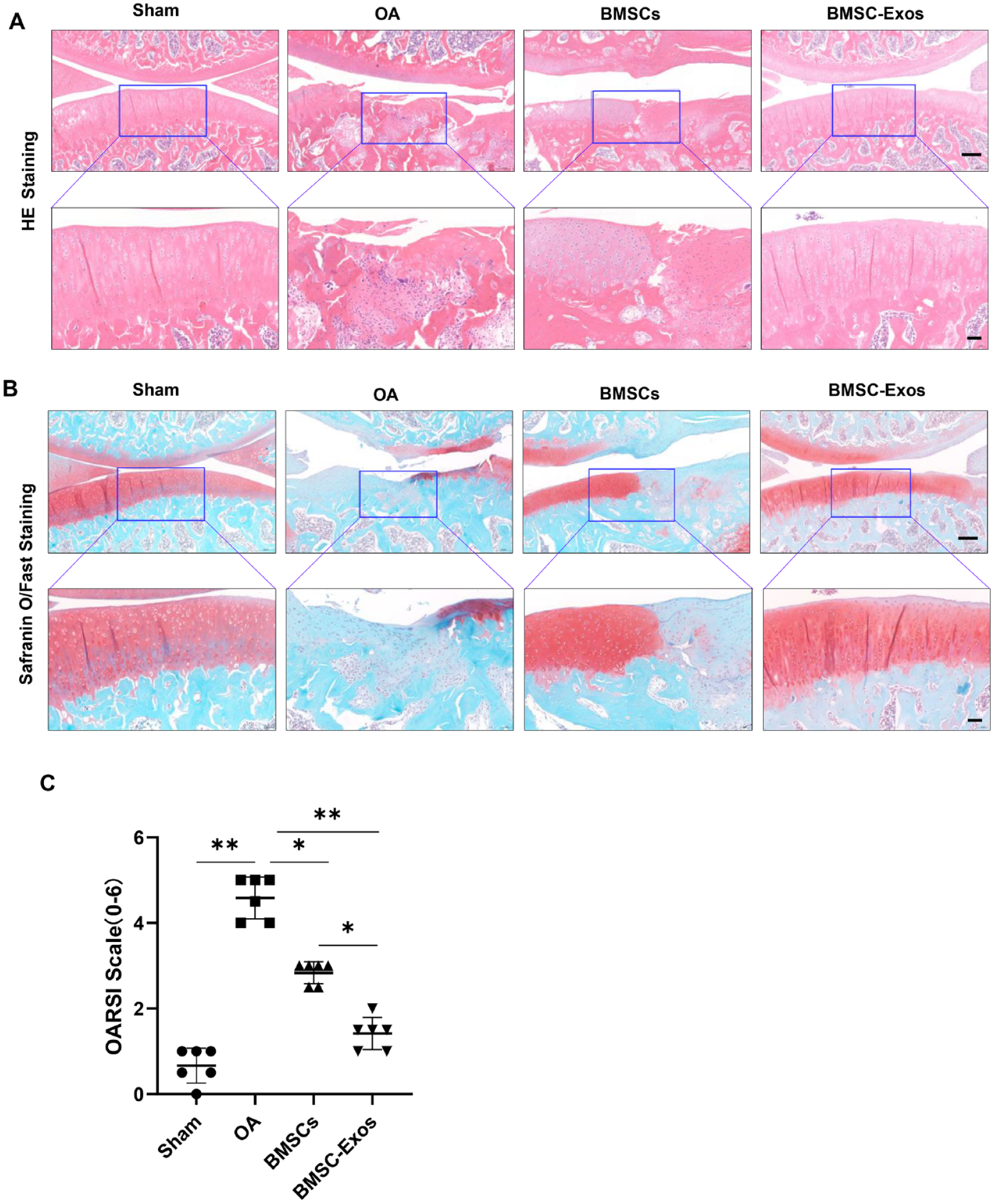
BMSC-Exos alleviated osteoarthritis in a rat model. PBS, BMSCs, and BMSC-Exos were injected intra-articularly into the joint cavity of the OA group, BMSCs group, and BMSC-Exos group, respectively, after 4 weeks of surgery and once a week for 6 consecutive weeks. (
BMSC-Exos Changed the Polarization of Synovial Macrophages
Immunohistochemistry and immunofluorescence analyses were applied to examine the effects of BMSC-Exos on the polarization of synovial macrophages in vivo. According to the results of HE staining, obvious synovial hyperplasia and inflammatory cell infiltration were found in the OA group (
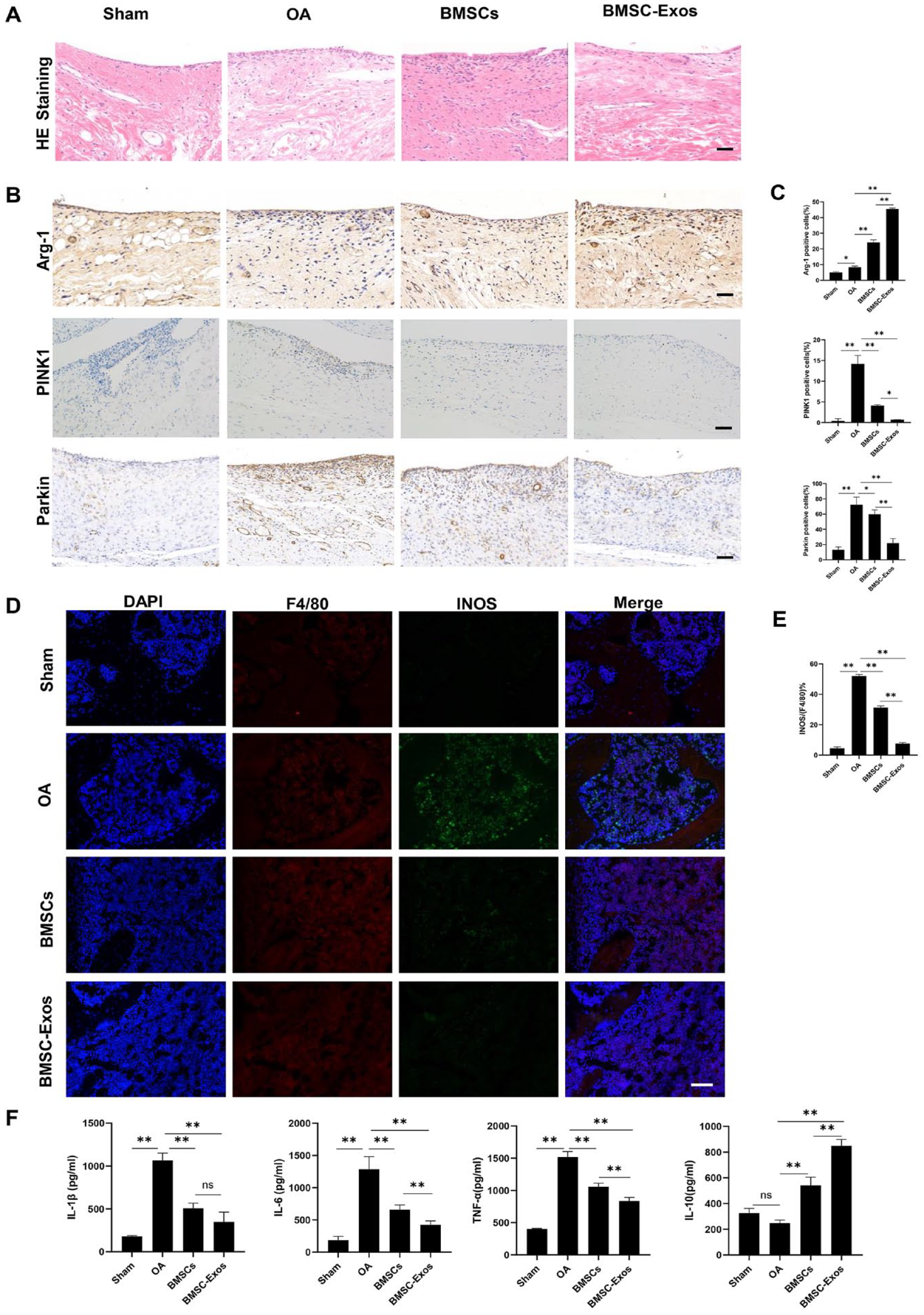
BMSC-Exos induced the transformation of M1 macrophages into M2 macrophages in the synovial tissue and reduced inflammation in the rat OA model. Treatment in each group was the same as in Figure 4. (
The ELISA results showed that the serum levels of pro-inflammatory cytokines IL-1β, IL-6, and TNF-α were the highest in the OA group, while the level of anti-inflammatory cytokine IL-10 was the lowest (
Discussion
BMSC-Exos have provided new hope for OA treatment since they have similar effects as BMSCs on cartilage regeneration but fewer risks. 25 The macrophage phenotype shift may be involved in this process, but the mechanisms remain unclear. In this study, we found that BMSC-Exos have anti-inflammatory functions both in vitro and in OA rats, in which M2 polarization is promoted and PINK1/Parkin signaling pathway plays a key role.
As components of the innate immune system, the primary role of macrophages is to protect the host from infection. 26 Macrophages show different polarization phenotypes, M1 and M2, in response to stimuli in their microenvironment. 27 M1 macrophages are also referred to as classically activated macrophages and exert pro-inflammatory effects, while M2 macrophages are referred to as alternatively activated macrophages and usually exhibit anti-inflammatory effects. 28 OA is an immune disorder, and increasing evidence has suggested that macrophages accumulate and become polarized in the synovium and articular cavity during OA development. 29
BMSC-Exos-based treatment has been widely studied regarding anti-inflammation and immune regulation. 5 For example, small EVs (including exosomes) derived from human umbilical cord MSCs can regulate macrophage polarization and promote the proliferation of chondrocytes, thereby delaying OA progression. 30 Another study demonstrated that BMSC-Exos can protect the brain in patients with traumatic brain injury by promoting the transformation of microglia into M2 macrophages, reducing neuroinflammation. 31 In our study, we prepared OA rat models and found that the polarization of M1 synovial macrophages was inhibited by treatment with either BMSCs or BMSC-Exos, relieving cartilage damage. Notably, the levels of pro-inflammatory factors in the serum, such as IL-1β, IL-6, and TNF-α, were reduced, while that of the anti-inflammatory factor IL-10 was increased after treatment with BMSCs or BMSC-Exos, indicating that the systemic inflammatory state was also altered. Other studies have demonstrated that a higher ratio of M1/M2 macrophages in the synovial fluid and peripheral blood is found in OA patients than in healthy controls, 32 indicating that changes in the local and systemic inflammatory states contribute to OA. 33
Studies have revealed that macrophage polarization is regulated in many physiological and pathological states 2 ; for example, human umbilical cord mesenchymal stem cell (hucMSC)-derived exosomes promote the healing of spinal cord injuries by reducing inflammation at the injury site and inducing the conversion of macrophages from the M1 phenotype to the M2 phenotype. 34 However, the mechanisms by which exosomes modulate macrophage polarization remain unclear. S1P/SK1/S1PR1 signaling might be involved in M2 macrophage polarization induced by adipose-derived mesenchymal stem cell (ADSC)-derived exosomes to improve cardiac damage, 35 and pknox1 might participate in M2 macrophage polarization by BMSC-Exos and enhance cutaneous wound healing. 36 PINK1/Parkin signaling is important for macrophage polarization. 37 The activation/phosphorylation of Akt plays a negative role in the PINK1/Parkin signal pathway. 38 The PINK1/Parkin signaling pathway mainly mediates the autophagy of cells and can modulate mitochondrial damage. 39 In our study, the decrease in Akt phosphorylation and the increase in PINK1 and Parkin expression induced by LPS were partly reversed by BMSC-Exos. The markers of M1 macrophages, CD86 and iNOS, were downregulated by BMSC-Exos; however, the markers of M2 macrophages, Arg-1 and CD206, were upregulated. Thus, PINK1/Parkin signaling may be involved in macrophage polarization. The reduction of ROS and the recovery of the mitochondrial membrane potential induced by BMSC-Exos further confirm their beneficial role.
However, further studies are required to confirm the molecular mechanisms of BMSC-Exos in macrophage polarization. Many biological molecules, including proteins, lipids, and nucleic acids, are encapsulated in BMSC-Exos, among which many miRNAs regulate multiple signal transduction pathways. miR-140-5p can enhance the proliferation and migration of chondrocytes, and miR-140-5p might promote the anabolism of the extracellular matrix. 40 More studies should be conducted on these biological molecules and their related mechanisms to further explore macrophage polarization by BMSC-Exos in OA.
Conclusion
In summary, BMSC-Exos can reduce articular cartilage damage and synovium inflammation by regulating the polarization of macrophages to M2, a process in which the PINK1/Parkin signal pathway might play an important role. Although other underlying mechanisms of BMSC-Exos in macrophage polarization still need to be explored, our results confirm that a therapeutic strategy based on BMSC-Exos is superior to that based on BMSCs in OA.
Footnotes
Author Contributions
Danping Liu and Xu Jiang were responsible for the conception, design, and interpretation of data. Beibei Li, Enpu Shen, and Zhiwen Wu performed the experiments. Cheng’ai Wu and Hui Qi analyzed data. Beibei Li wrote the paper.
Acknowledgments and Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the National Natural Science Foundation of China (grant number: 81572140), Beijing Municipal Health Commission (grant numbers: BMC2019-9, BMC2021-6), Beijing Natural Science Foundation—Haidian Original Innovation Joint Fund (L222089), and Beijing Research Institute of Traumatology and Orthopedics Foundation (QN-2023-08).
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethics Approval
The animal experiments were approved by the local ethics committee: Animal Research Ethics Committee of Jinzhou Medical University (2022025). All methods were carried out and reported in accordance with ARRIVE guidelines.
Availability of Data and Materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.