
Abstract
Objective
Multilineage differentiating stress-enduring (Muse) cells, a pluripotent stem cell subset of mesenchymal stem cells (MSCs), have shown promise for various tissue repairs due to their stress tolerance and multipotent capabilities. We aimed to investigate the differentiation potential in vitro, the dynamics in vivo, and the reparative contribution of Muse cells to osteochondral lesions.
Design
Labeled MSCs were cultured and sorted into Muse and non-Muse (MSCs without Muse cells) groups. These cells were then formed into spheroids, and chondrogenic differentiation was assessed in vitro. Twenty-one immunocompromised mice were used as the in vivo models of osteochondral lesions. Live imaging, macroscopic evaluation, and histological and immunohistochemical analyses were conducted at the 4- and 8-week time points.
Results
Muse cell spheroids were formed, which were larger and stained more intensely with toluidine blue than non-Muse spheroids, indicating better chondrogenic differentiation. Live imaging confirmed luminescence in all 4-week model knees, but only in a few knees at 8 weeks, suggesting cell persistence. Macroscopically and histologically, no significant differences were observed between the Muse and non-Muse groups at 4 and 8 weeks; however, both groups showed better cartilage repair than that of the vehicle group at 8 weeks. No collagen type II generation was observed in the repaired tissues.
Conclusion
The implantation of the spheroids of Muse and non-Muse cells resulted in better healing of osteochondral lesions than that of the controls, and Muse cells had a higher chondrogenic differentiation potential in vitro than non-Muse cells.
Introduction
Articular cartilage lesions lack the ability to regenerate and cannot be repaired by normal articular cartilage, leading to osteoarthritis.1,2 Treatments for articular cartilage lesion have included procedures such as bone marrow (BM) stimulation, 3 mosaicplasty osteochondral bone graft, 4 autologous chondrocyte implantation (ACI), 5 and other methods. However, a treatment that can reproduce the complete vitreous cartilage and successfully repair the cartilage has yet to be established. The repair of articular cartilage is very challenging. Nevertheless, because of the significant benefits in terms of the subsequent condition of the knee joint, many researchers are still developing new therapies from various angles.
Cell-based therapy is one such therapy. Since ACI was first reported by Brittberg et al. 5 in 1994, many researchers have been actively studying and reporting on various cell-based therapies. Chondrocyte-based therapies have been studied in terms of the method of administration, combination with various scaffolds, and transplantation after in vitro culture.6-8 Induced pluripotent stem cell–derived chondral particles have been used in cartilage regeneration. 9 Mesenchymal stem cells (MSCs) have multilineage potential and are one of the most common cell types being investigated for experimental cell-based therapies to treat human diseases. 10 MSCs have been reported as useful in the treatment of various organ damages; however, their effects are mainly indirect such as the paracrine action to protect the damaged area,11,12 MSCs have also been used for osteochondral lesions with mixed results.13-16 Researchers have suggested that the direct reparative effects of injured cartilage by MSCs are minimal, and that their effects are mainly indirect such as their role in paracrine action. 17 However, recent studies have shown that MSCs show high chondrogenic differentiation ability by the administration of aggregate formation and that they are excellent for osteochondral repair.18-22
Multilineage differentiating stress-enduring (Muse) cells, first reported by Kuroda et al., 22 are unique multipotent cells, existing in approximately 1-5% of adult human MSC populations. These cells are pluripotent-like stem cells which can differentiate into endodermal, ectodermal, and mesodermal cells from a single cell and have the abilities of self-renewal and stress tolerance.
Muse cells are collectible as stage-specific embryonic antigen (SSEA)-3–positive cells from MSCs derived from various tissues such as BM, fat, skin, and synovium.23-26 As they normally reside in adult tissues, Muse cells have low telomerase activity and are not tumorigenic.23-25 Muse cells have been studied and their efficacy has been reported in diseases of various organs such as brain, skin, liver, heart, lung, and cornea.27-37 In these studies, Muse cells were found to differentiate directly into damaged cells. MSCs without Muse cells lack this ability. Therefore, superiority of Muse cells in tissue repair has been confirmed. One known report on osteochondral lesions using Muse cells investigated osteochondral repair by the intra-articular administration of Muse cells; however, the study did not include detailed verification of the cell pathology or whether the cells actually contributed to the repair. 38 Whether Muse cells contribute to the direct repair of articular cartilage injuries remains unclear.
The purpose of this study was to examine the healing processes, including in vivo dynamics, contribution to replenishment by differentiation into new bone and cartilage cells, and repair of osteochondral tissue after implantation of human BM Muse cells into osteochondral lesions in an immunosuppressed mouse model, and compare the effects with those after implantation of human BM MSC without Muse cells.
Methods
Labeling of MSCs with Venus-Akaluc (VA)
Human BM MSCs were purchased from Lonza (Basel, Switzerland) and expressed VA 39 after transfection with the pcDNA3 VA-labeled lentivirus (CS2-EF-MCS). 40 Transformation was performed on LentiX-293T using 3 promoters (VA on CS2-EF-MCS, pCMV-VSV-G, and pCAG HIV). The resulting supernatant was used for transfection into MSCs. The pcDNA3 VA and CS2-EF-MCS were provided by RIKEN BRC through the National BioResource Project of MEXT/AMED, Japan. CS-EF-MCS was developed by Hiroyuki Miyoshi.
Preparation of Muse and Non-Muse Cells
MSCs after VA labeling were cultured at 37°C with 5% CO2 in low-glucose Dulbecco’s minimum essential medium (DMEM) supplied with 10% fetal bovine serum (FBS; HyClone, Logan, UT), 1 ng/ml human FGF-2 (Miltenyi Biotec, Bergisch Gladbach, Germany), 0.1 mg/ml kanamycin (Invitrogen, Carlsbad, CA), and 1% GlutaMAX (Life Technologies, Carlsbad, CA). After 90% confluence was reached, the cells were subcultured at a ratio of 1:2 with 0.25% trypsin-EDTA (Life Technologies, Carlsbad, CA). MSCs from the 8th to 10th subcultures were used in this study. Fluorescence-activated cell sorting (Aria III, Becton Dickinson Biosciences, Franklin Lakes, NJ) was used to isolate Muse and non-Muse cells and to select VA-expressing cells. MSCs were first incubated with rat anti-SSEA-3 IgM antibody (clone MC-631; Millipore, Billerica, MA) and further incubated with an Allophycocyanin AffiniPure F(ab’)2 Fragment Goat Anti-Rat IgM (immunoglobulin M) antibody (Jackson ImmunoResearch, West Grove, PA). VA-labeled Muse cells were collected and sorted as Venus-positive/SSEA-3-positive, while VA-labeled non-Muse cells were sorted as Venus-positive/SSEA-3-negative, as described previously, with several modifications. 23
Muse (M)-Cluster Formation in Single-Cell Suspension
Individual Muse cells were dispensed into each well of a 96-well dish coated with poly 2-hydroxyethyl methacrylate (Merck, Darmstadt, Germany) after the dilution was limited. Single cells were cultured in low-glucose DMEM supplemented with 10% FBS, FGF-2, kanamycin, and GlutaMAX, as described above. At 7 to 10 days after starting the culture, the formation of the M-cluster was confirmed when diameters were >25 μm.23,25
Muse and Non-Muse Spheroids
Muse cells and non-Muse cells were cultured in non-adherent U-shaped 96-well plates with 50,000 cells in 200 μl medium per well for 3 days. The presence and size of cell aggregates (spheroids) were determined.
In Vitro Chondrogenic Differentiation
Spheroids of 50,000 cells were transferred to 15-ml polypropylene tubes containing 500 μl of respective specific media using StemPro® Chondrogenesis Differentiation Kit (Life Technologies, Carlsbad, CA) and incubated at 37°C and 5% CO2 with the lids loosened for 21 days. The medium was changed every 3 days.
Toluidine blue staining was performed after 21 days. After chondrogenic differentiation, the spheroids were embedded in optimal cutting temperature compound (Sakura Fine Technical, Tokyo, Japan). The cryoprotected samples were frozen and kept at −80°C until use. The 5-μm sections were prepared with a cryostat, mounted on slides, and stained with 0.05% toluidine blue (Muto pure chemicals, Tokyo, Japan). Images were obtained using an all-in-one microscope, BZ-X810 (Keyence, Osaka, Japan).
Animals
All animal procedures were approved by the ethics committee of our institution, followed the National Institutes of Health guidelines for the care and use of laboratory animals, and complied with the ARRIVE (Animal Research: Reporting of In Vivo Experiments) guidelines (Supplemental material). Twenty-one skeletally mature 6-week-old male C.B-17/Icr-scid/scidJcl mice (CLEA Japan, Fujinomiya, Japan) with a mean weight of 23.0 g were used in this study (
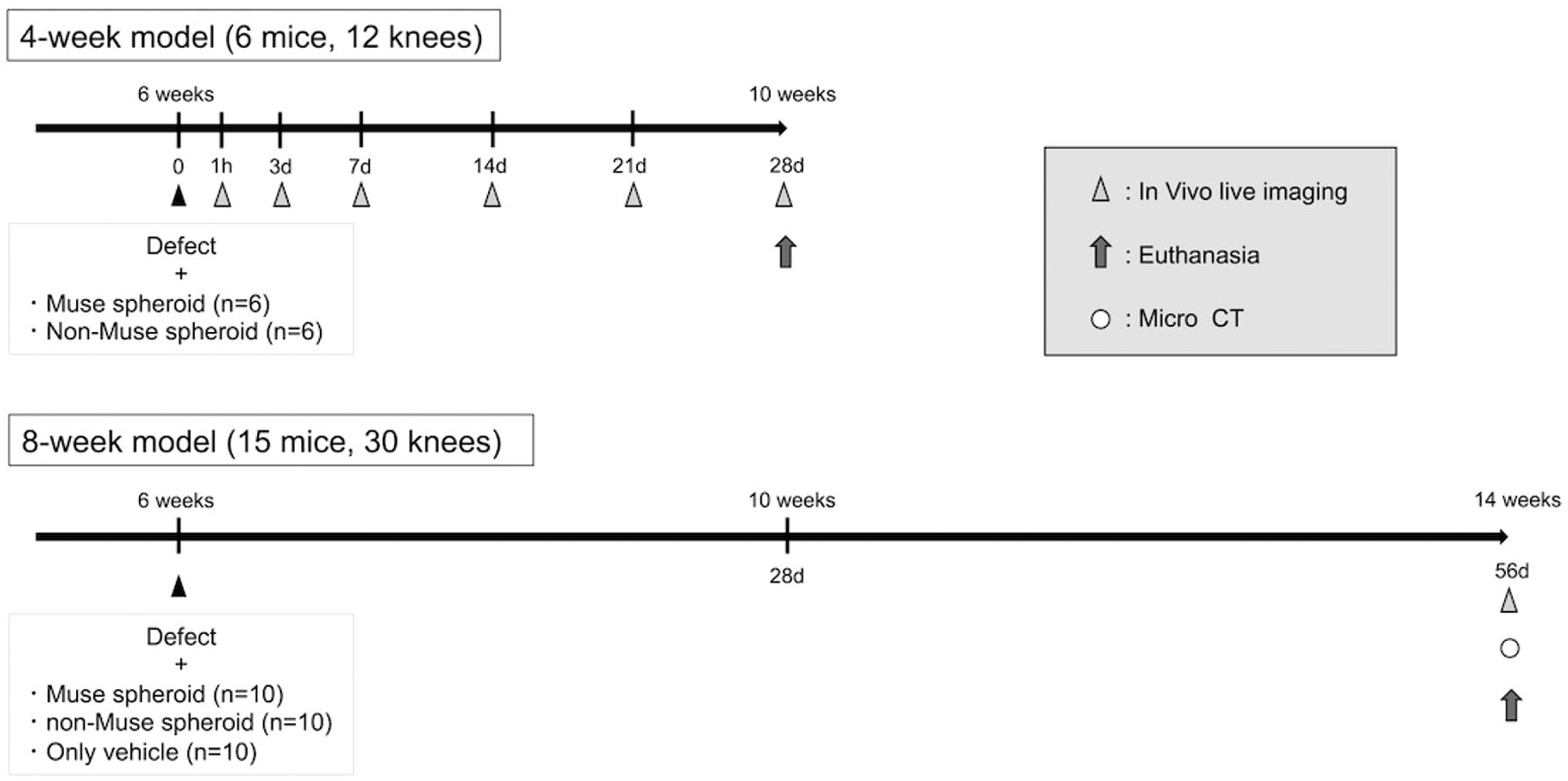
Animal models schedule. Four-week models were divided into 2 groups for cell tracking investigation; the 8-week models were divided into 3 groups. CT = computed tomography.
In Vivo Transplantation of Muse versus Non-Muse Spheroids
All animals were anesthetized by intraperitoneal injection of medetomidine (0.3 mg/kg), midazolam (4 mg/kg), and butorphanol (5 mg/kg) and underwent bilateral knee surgery. A medial parapatellar approach was performed in each knee. The patella was laterally dislocated to allow full exposure of the articular surface, and an electric microdrill (
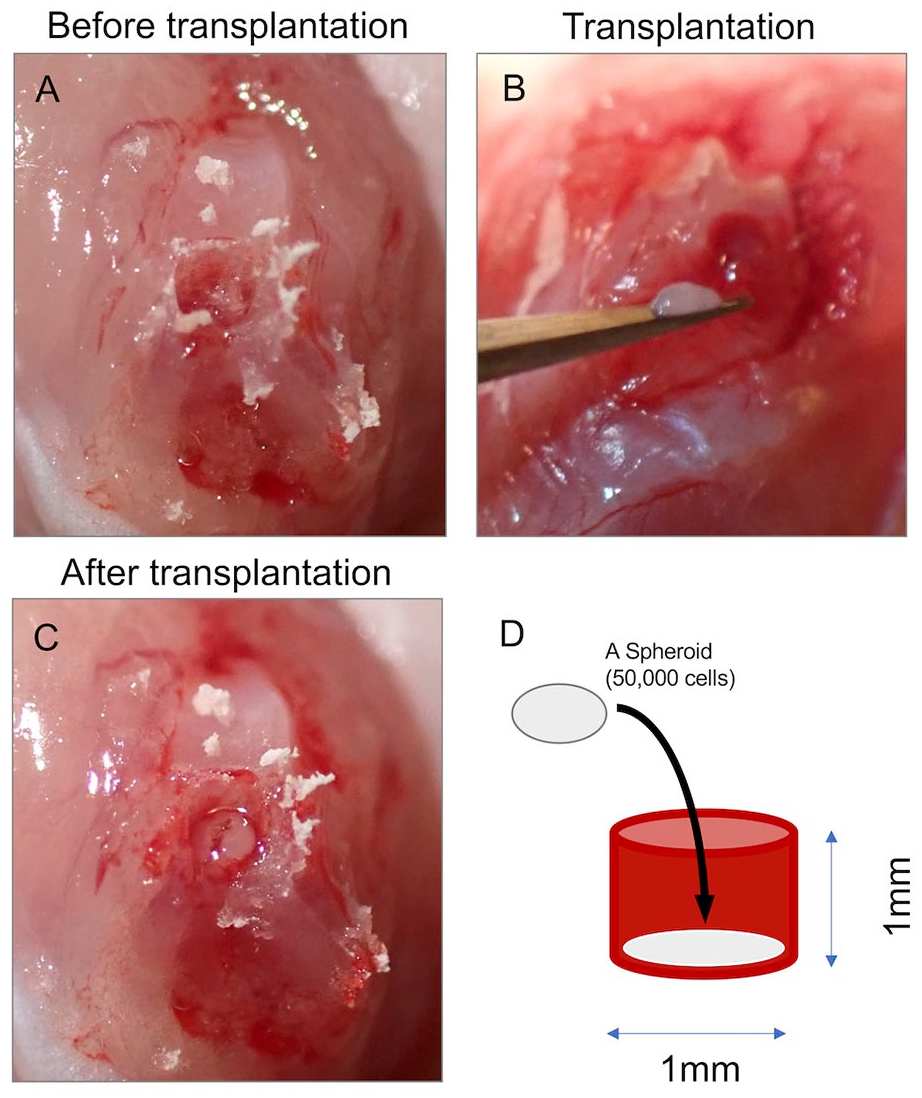
Transplantation of Muse and non-Muse spheroids. (A) An osteochondral defect was created at the femoral groove using an electric micro drill. (B, C) The spheroid was transplanted to the defect. (D) Schematic of the defect shows the diameter of 1 mm and depth of 1 mm.
In Vivo Live Imaging with AKABLI System
Three mice (6 knees) in each group (M and N), as 4-week models, were subjected to live imaging examinations at 1 hour, 1 day, 3 days, 1 week, 2 weeks, 3 weeks, and 4 weeks postoperatively. Five mice (10 knees) from each group, for a total of 30 knees of 15 mice, as 8-week models, were also subjected to live imaging examinations, but only at 8 weeks postoperatively (
IVIS Lumina II (PerkinElmer, Boston, MA) was used with the AKABLI system 39 activated by the reaction between the introduced enzyme, Akaluc, and intraperitoneal administration of the substrate, 100 μl of 30 mM AkaLumine-HCl. Ten minutes after administration, images were acquired under the following conditions: open for total bioluminescence, exposure time = 300 seconds, binning = medium: 4, field of view = 10 or 12.5, and f/stop = 1. Bioluminescent images were analyzed using Living Image 3.2 software (PerkinElmer) developed specifically for the IVIS system.
Macroscopic Evaluation
Macroscopic evaluations were performed by two blinded examiners. The International Cartilage Repair Society (ICRS) macroscopic cartilage evaluation score and grading system (grade 1, normal, 12 points; grade 2, nearly normal, 8-11 points; grade 3, abnormal, 4-7 points; and grade 4, severely abnormal, 0-3 points) were used. 41
Micro–Computed Tomography (CT) Scanning
Micro-CT was performed by wrapping the entire distal femur in parafilm and then scanning at 38-mm resolution, 70 kVP, and 112 mA using a Viva CT-40 (Scanco Medical). For micro-CT analysis, a Gaussian support of 0.8, sigma of 1, and a threshold of 220 for 3-dimensional reconstruction were used to reveal overall chondral defect healing.
Histological Evaluation
The harvested knees were fixed with 4% PFA for 2 days at room temperature, decalcified with 10% ethylenediaminetetraacetic acid for 3 weeks, split in the sagittal direction, processed, and embedded in paraffin blocks, then dehydrated with an ethanol series and cleared with xylene. A 5-μm-thick section was prepared using a microtome and histologically stained with safranin-O and fast green (SOF) for histological evaluations which were blindly rated by two researchers using the O’Dricolll score. 42 A score of 0 was assigned for properties considered indicative of poor quality and 24 was assigned for good-quality cartilage (i.e., as would be observed in normal articular cartilage).
Immunohistochemistry
Rabbit polyclonal antibodies against human type I collagen (anti-COL1; #ab34710, Abcam, Cambridge, UK) and mouse monoclonal antibodies against human type II collagen (anti-COL2; #F-57, Kyowa Pharmaceuticals, Tokyo, Japan) were used. Before staining, the specimens were treated with the following enzymes after deparaffinization. For the anti-COL1 antibody, pretreatment was performed by soaking for 2 minutes in an enzyme solution of Proteinase K (S3004; DAKO, Glostrup, Denmark) at room temperature. For anti-COL2, the pretreatment was performed by soaking for 60 minutes in an enzyme solution of hyaluronidase (H3506; Sigma-Aldrich Corporation, St. Louis, MO) dissolved to 0.05% in 100 mM acetate buffer (pH 5.0) at 37°C. For anti-COL1, anti-rabbit IgG goat polyclonal antibody (HISTOFINE #424144; Nichirei Corporation, Tokyo, Japan) labeled with horseradish peroxidase (HRP) was used as the secondary antibody and allowed to react with the specimens for 30 minutes at room temperature. For anti-COL2, the HISTFINE Mouth Stain Kit (HISTOFINE #414321; Nichirei Corporation, Tokyo, Japan) was used to block nonspecific reactions, and HRP-labeled anti-mouse IgG (immunoglobulin G) goat polyclonal antibody (HISTOFINE #424134; Nichirei Corporation, Tokyo, Japan) was added to the specimens for 30 minutes at room temperature. Positive sites were visualized as dark brown using 3,3′-diaminobenzidine tetrahydrochloride (K3468, DaKO, Glostrup, Denmark) as a chromogenic substrate; then, the nuclei were contrast-stained to a light purple color using hematoxylin.
Immunofluorescence (IF)
The anti-green fluorescent protein (anti-GFP) rabbit polyclonal antibody (#598; Medical-Biological Laboratories, Aichi, Japan) was diluted 1,000x and allowed to react with the specimens for 60 minutes at room temperature. The secondary antibody, Alexa Fluor 488 anti-rabbit IgG(H+L) goat polyclonal antibody (A11034; Invitrogen, Carlsbad, CA), was diluted 500x and allowed to react with the specimens for 60 minutes at room temperature. The contrast for staining, 4′-6-diamidine-2′-phenylindole, dihydrochloride (DAPI: #340-07971; Dojindo Kumamoto, Japan) was diluted 5,000x and allowed to react at room temperature for 10 minutes to stain the nuclei blue. Images of the SOF-stained and enzyme-antibody-immunostained specimens were observed under an optical microscope (BX-43; OLYMPUS Corporation, Tokyo, Japan). The IF images were obtained using a fluorescence microscope (BZ-X810; Keyence Corporation, Osaka, Japan), and merged images of anti-GFP+DAPI were produced.
Statistical Analysis
The mean scores of the ICRS macroscopic cartilage evaluation and O’Dricoll scores of the two investigators were analyzed using the Kruskal-Wallis and Welch’s t test. Statistical significance was set at P < 0.05. All statistical analyses were performed using STATA version 18 (StataCorp, College Station, TX).
Results
Muse and Non-Muse Cells
VA-labeled Muse cells were extracted at less than 5% of Venus-positive cells, respectively (
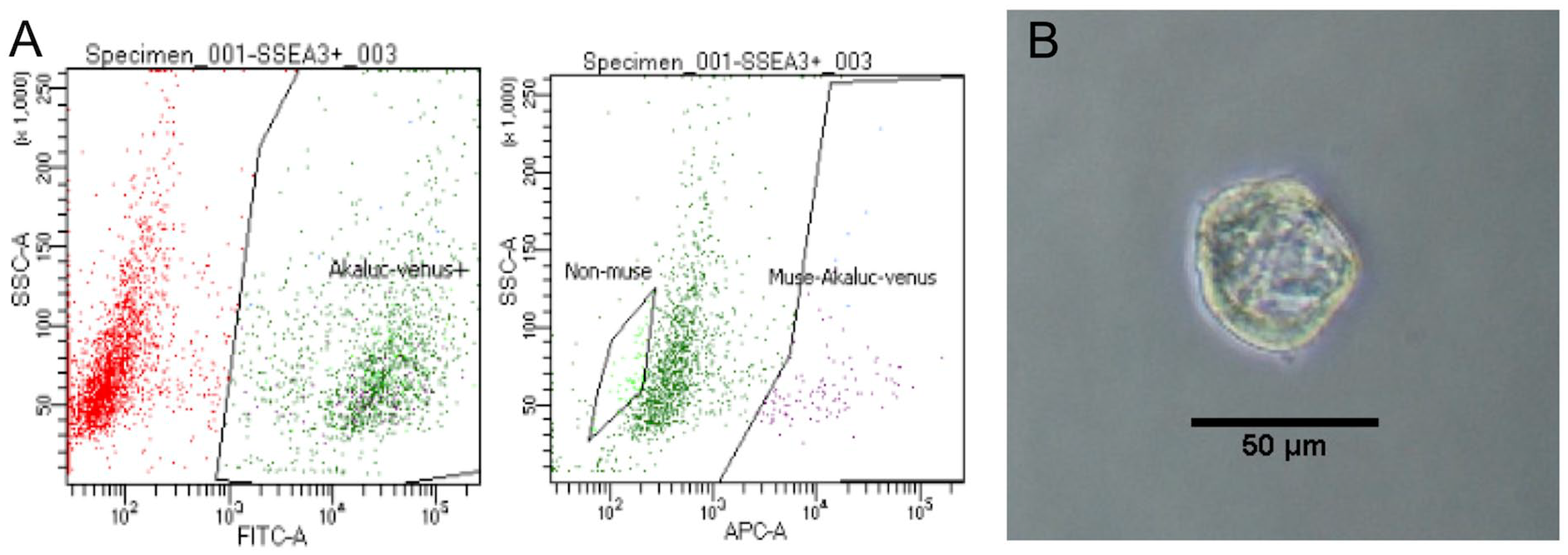
Muse cell sorting and confirmation of the pluripotency. (A) Venus-Akaluc-labeled cells were sorted. SSEA-positive cells were sorted as Muse cells and SSEA-negative cells were sorted as non-Muse cells. (B) M-cluster formation in single-cell suspension. SSEA = stage-specific embryonic antigen; -A = -Area; SSC = side scatter; FITC = fluorescein isothiocyanate; APC = allophycocyanin.
In Vitro Chondrogenic Differentiation
After chondrogenic culture, 3 masses were created in each M or N group; masses were somewhat firm to the touch and resembled external cartilage fragments. The masses of the Muse spheroids were larger than those of the non-Muse spheroids (

Size of cartilage masses comparing Muse and non-Muse cells. After 3 weeks of chondrogenic culture, every spheroid resembled a small piece of cartilage.
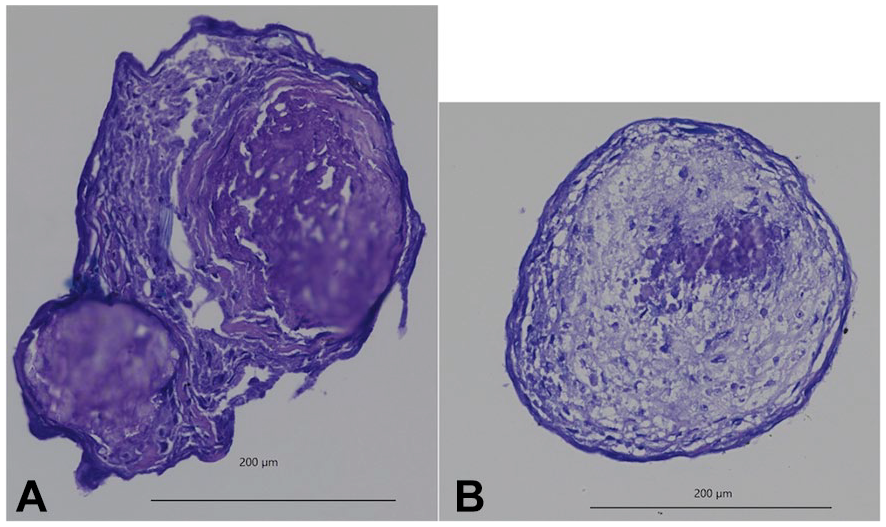
Toluidine blue staining after chondrogenic culture. Cartilage fragments from Muse spheroids (A) stained better with toluidine blue than those from non-Muse spheroids (B).
In Vivo Live Imaging for Cell Tracking with AKABLI
Among the 4-week models, one mouse in the N group died between 3 and 4 weeks, and one mouse in the M group died between 1 and 2 weeks. These deaths were likely from an infection caused by too many injections of anesthesia and substrate. The remaining 4 mice in each group were each subjected to 6 live imaging shots for up to 4 weeks. The luminescence of the administered cells was confirmed in all remaining mice (48 knees in 24 shots;
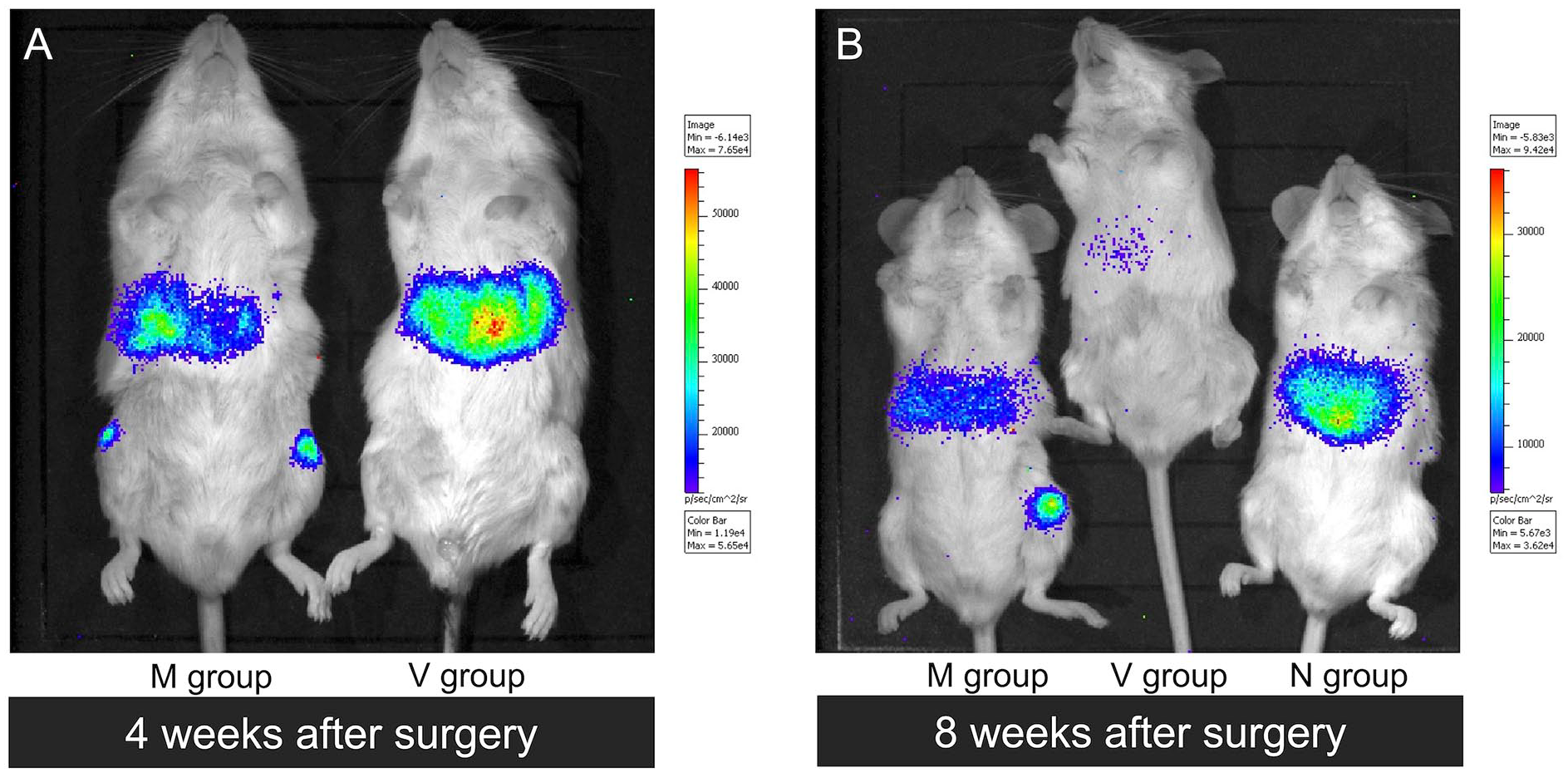
In vivo live imaging for cell tracking. Non-specific luminescence can be observed in the livers. (A) In the 4-week model, luminescence was observed in all knees in the M and N groups. (B) In the 8-week model, luminescence was observed in only 2 of 10 knees in the M group and 3 of 10 knees in the N group.
In the 8-week model, 15 mice were subjected to live imaging shots 8 weeks after the procedure. Luminescence of the administered cells was confirmed in only 5 knees: 2 in the M group and 3 in the N group (
Macroscopic Evaluation of Cartilage Regeneration Using the ICRS Score
At the time of euthanasia, no obvious synovitis was observed macroscopically in any of the 38 knees, except in two mice that died during the experiment. The defect in the femoral groove was covered with cartilage-like tissue on both sides of each knee, although there were various levels of differences.
No significant difference was observed in the ICRS scores between the M and N groups of the 4- or 8-week models. Comparisons of the V and MSC groups and the combined M and N groups showed that the MSC group had better scores in the ICRS overall repair assessment than the V group in the 8-week model (P < 0.05;
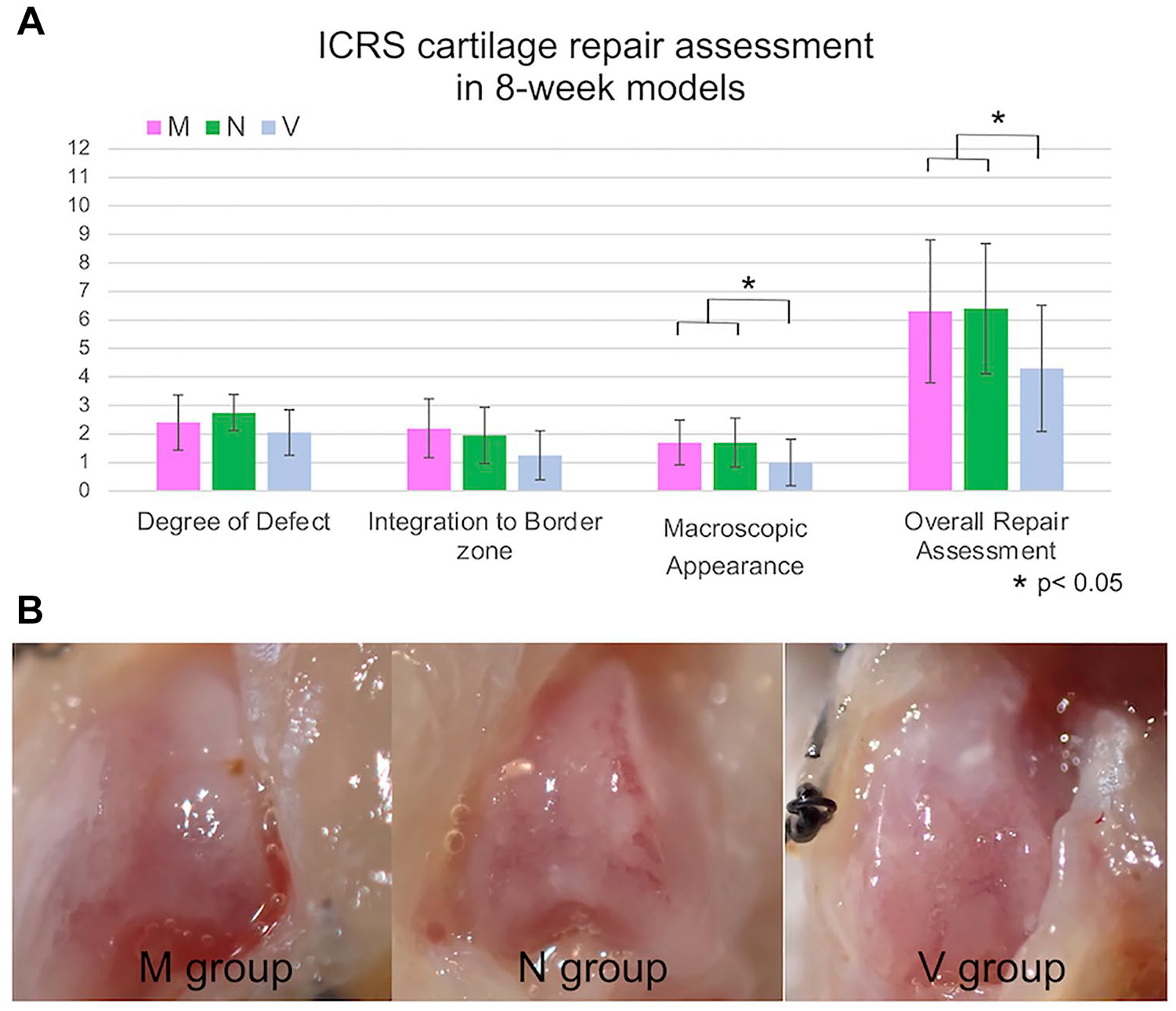
Macroscopic evaluation in 8-week models. (A) The ICRS macroscopic cartilage evaluation score showed that the MSC group with the combined M and N groups had a better score in the ICRS overall repair assessment than the V group in the 8-week models (Welch’s t test). (B) Injured lesions in the 3 groups are shown. ICRS = International Cartilage Repair Society; MSC = mesenchymal stem cell. Statistically significant differences are indicated by *P < 0.05.
Micro-CT Evaluation of Subchondral Bone Repair
Micro-CT was performed on 30 knees of the 8-week models at the time of euthanasia. In all cases, regardless of the group, there was no obvious difference in the regeneration of the subchondral bone, regardless of whether the surface was covered with tissue (
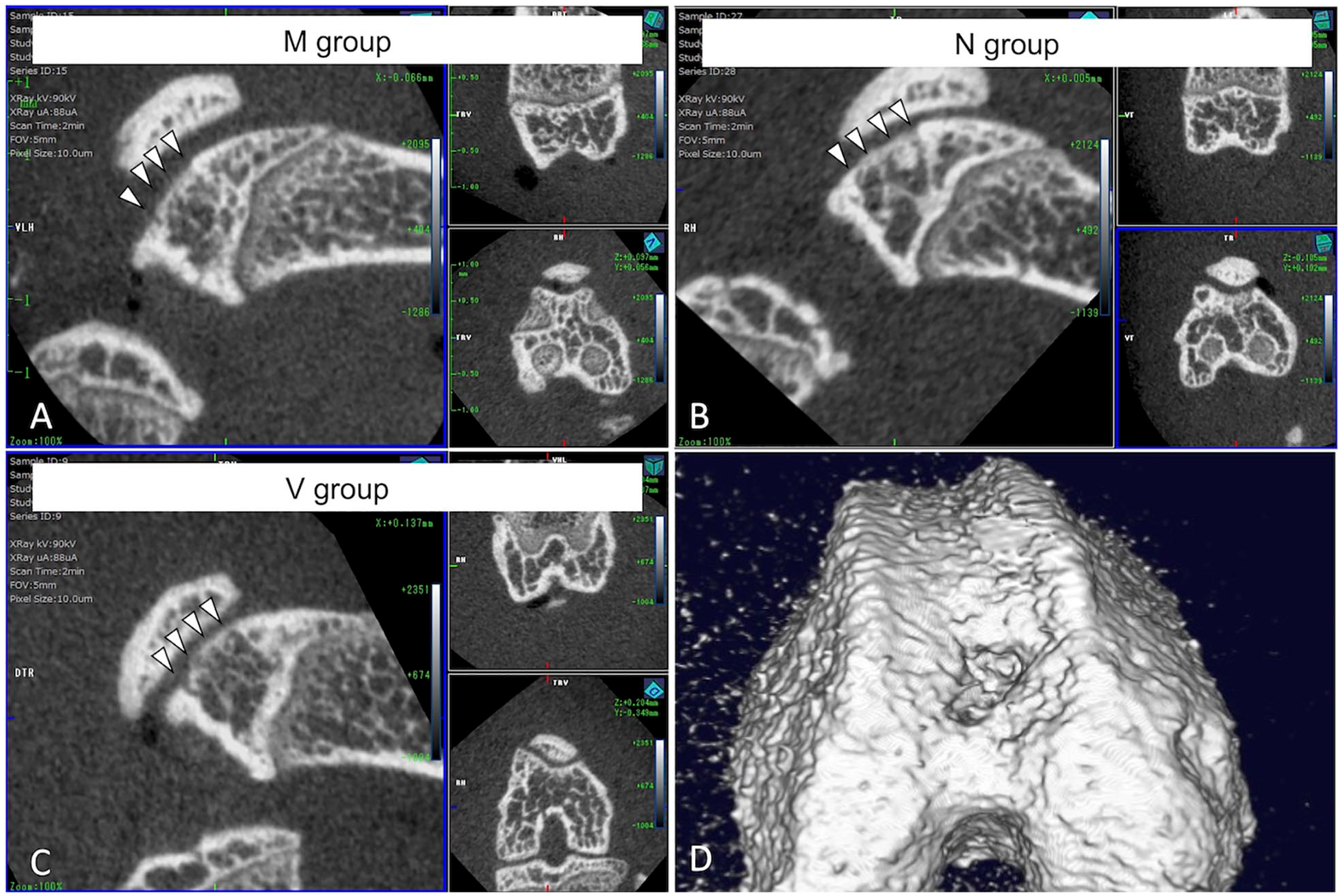
Micro-CT evaluation in 8-week models. The representative images show no obvious differences in subchondral bone regeneration among the 3 groups (A, B, C, and D). CT = computed tomography.
In Vivo Histological Evaluation
Histological evaluations were performed using the O’Dricoll score and SOF staining in 30 knees of the 8-week models. Each individual scoring item and total scores were not significantly different between the groups. However, the MSC group, combining the M and N groups, had better scores in cellular morphology, matrix staining, freedom from degenerative changes in adjacent cartilage, and total O’Dricoll score than the V group in the 8-week models (Welch’s t test, P < 0.05;
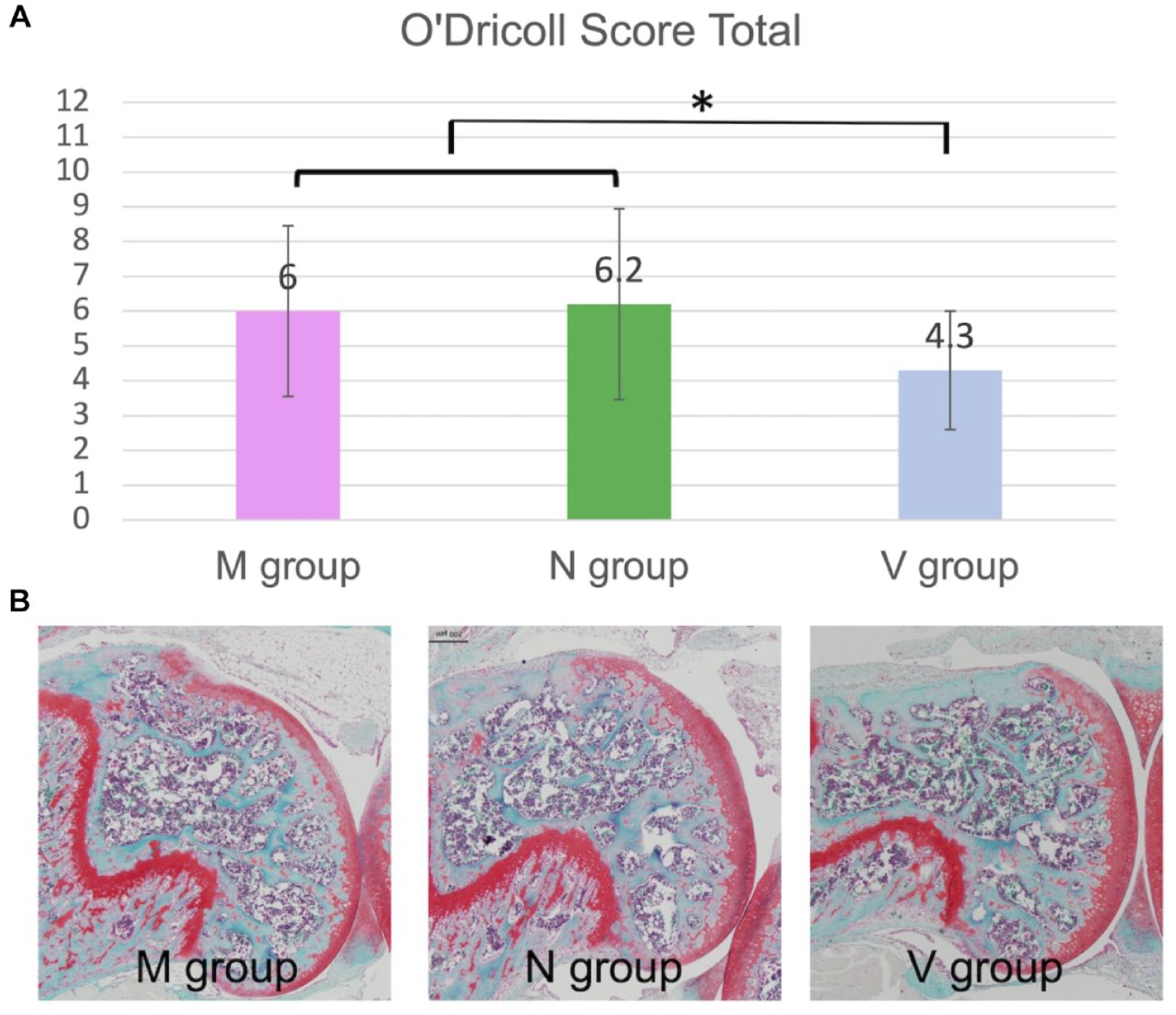
O’Dricoll score in 8-week models. The individual score items and total scores were not significantly different among the 3 groups. The MSC group, combining the M and N groups, had a better O’Dricoll score than the V group in the 8-week model (Welch’s t test). MSC = mesenchymal stem cell. Statistically significant differences are indicated by *P < 0.05.
In Vivo Muse Cell Differentiation on Osteochondral Lesions
Anti-COL1 was positive and anti-COL2 was negative in the repair tissue. Anti-GFP was negative in the osteochondral defect area. Positive cells that were administered as Muse or non-Muse cell spheroids 8 weeks prior were in another area of the knee joint (
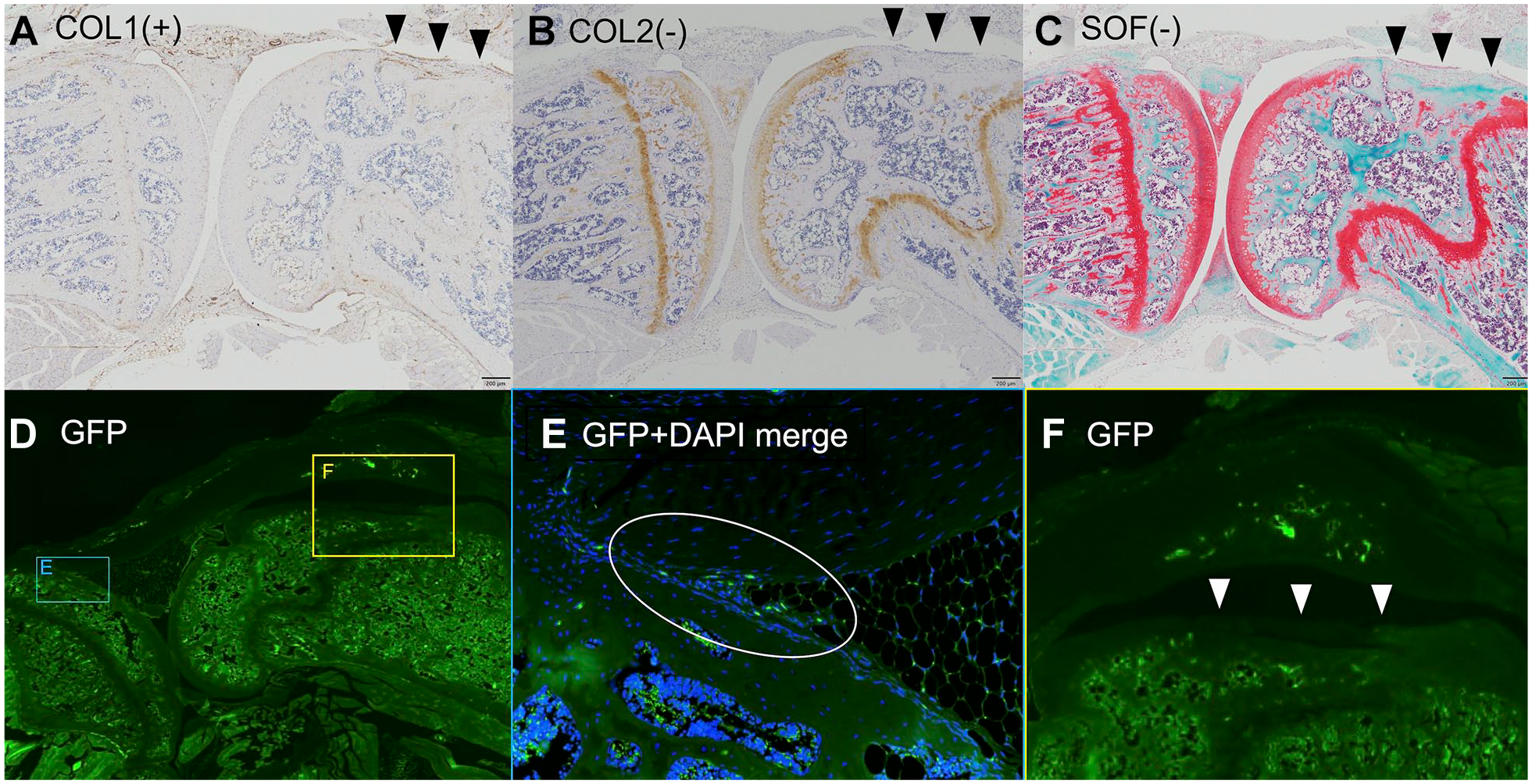
Microscopic evaluations for the osteochondral lesions. (A) Anti-COL1 was positive with the repair tissue cell; (B) anti-COL2 was negative with the repair tissue cell; (C) SOF staining shows no regenerated articular cartilage; (D, E, and F) anti-GFP was negative in the osteochondral defect area (F), but positive in another area of a knee joint (E). COL = collagen; SOF = safranin-O and fast green; GFP = green fluorescent protein; DAPI = 4′-6-diamidine-2′-phenylindole.
Discussion
Our study had valuable findings. MSC spheroids, including both Muse and non-Muse cells, resulted in good repair of osteochondral lesions based on macroscopic evaluation compared with the control group. No significant difference was observed in the repair of osteochondral lesions between the Muse and non-Muse spheroids. Tracking the administered cells using IVIS showed MSCs in the knee joint 4 weeks after administration, and almost all cells disappeared by 8 weeks after administration. Muse cells did not directly repair the injured cartilage and subchondral bone tissue.
The effects of MSCs, such as their paracrine action, are considered mainly indirect. These effects are exerted for 4-8 weeks before they discontinue and serve to help repair the damaged area with type I collagen. These considerations are consistent with previous reports.15-17 The current study is practical in that we used transgenic cells with luminescence and fluorescence labeling instead of painting, and followed changes over time in the same subjects without slaughtering the mice. In general, intra-articular MSC administration in mice knees leads to the distribution of MSCs around the synovial membrane for up to 4-8 weeks. The MSCs exert indirect effects which disappear when they fail to remain at the site of injury. Although we expected that MSCs would remain at the site of injury if they were implanted in the form of spheroids rather than injected with cell suspensions, we found that MSCs did not remain in situ or initiate differentiation.
These results may have been different if the Muse cells had remained in situ as a cell mass. Although non-Muse cells have chondrogenic differentiation potential, Muse cells have higher chondrogenic differentiation potential and visibly better chondrogenesis. The differentiation of Muse cells into cells at the site of injury has been reported in many diseases27-37; however, the mechanism of action has not been fully elucidated. Wakao et al. 42 recently reported that differentiation of Muse cells occurs earlier and in a different way than by the usual stimulation of surrounding factors. Rather, the authors determined that the differentiation and stimulation occurred by phagocytosis of dead cells at the site of injury. Moreover, the Muse cells exhibit field ethics and differentiate into the required cells. Cartilage is a specialized tissue with few chondrocytes, most of which are composed of cartilage matrix. Therefore, the number of dead chondrocytes is expected to be small, which is unfavorable for Muse cell differentiation. In addition, the osteochondral injury created in the current study was affected by hemorrhage, and even a 1-mm depth injury may have been too deep for the mouse knee compared to the thickness of the cartilage.
Other major issues must be resolved when considering using Muse cells for cartilage regeneration. One is the question of whether the cells can go directly to the injured areas of cartilage and bone, as has been reported in other organs, by performing the homing ability. During transvenous administration, there are two possible routes for Muse cells to reach the injured site: through the BM in the vascular system, or through the vascular system to the synovium, where they leave the synovium and reach the injured site. It is unlikely that cells would reach the injury site through the BM without bleeding. Furthermore, it is unlikely that large numbers of Muse cells would reach the injury site through the synovial side as undifferentiated cells. Even if the cells were delivered via the bleeding process on the BM side, they may likely diffuse into the joint space, and subsequent bleeding would promote cartilage fibrosis. In contrast, during intra-articular administration, cells suspended in the joint fluid attach to the damaged area. However, most cells attach to other tissues, including the synovium (our previous unpublished study on intra-articular seeding showed that the cells were incorporated into the synovium and disappeared earlier than did the cells in the present study). Even when the formed spheroids were transplanted, they eventually migrated to the synovium. Second, it is important to consider whether the Muse cells stay in the injured area, even if they ultimately end up there. The cartilage surface faces the joint fluid; therefore, it is difficult for the cartilage to adhere, and an appropriate scaffold is required. Placing cartilage particles that have differentiated to some degree could be considered; however, this would erase the attractive feature of Muse cells, in that they differentiate in accordance with the tissue properties of the location. We believe that we should first consider a form of administration that allows the cells to remain and enables them to combine with the scaffolds. Verifying even homing capability all at once would then be a difficult challenge with cartilage damage. Therefore, we used the direct administration method in the present study.
Our literature search revealed only one study reporting on osteochondral lesions using Muse cells; that study investigated osteochondral repair by intra-articular administration of Muse cells. However, the study did not include detailed verification of cell pathology or whether the cells directly contributed to the repair. 38 To our knowledge, our study is the first study to examine in detail the role of cell tracking after Muse cell administration in cartilage injury, investigate direct healing by Muse cells, and to suggest a higher chondrogenic differentiation potential in Muse cell spheroids than that in non-Muse cell spheroids.
This study had some limitations that must be considered when clinically applying the results. First, the sample size was small, which means that we potentially missed the chance to detect differences between Muse and non-Muse cells. Second, the ratio of the depth of the osteochondral lesions differed from that in clinical situations. Third, we evaluated the histological evaluation and the differentiation on the osteochondral lesions only for the 8-week model. For the 4-week model, we just evaluated the live imaging evaluation to find out whether the implanted cells are alive or not. Considering the results, it was more obvious to check the status of the cells even at 4 weeks. However, the fact remains that the cells were not viable at the end of the study. Fourth, to evaluate the chondrogenic differentiation potential of Muse in vitro, we judged only the size and appearance of the actual chondrogenic changes and the staining of the tissue specimens, but did not perform any quantitative evaluation such as reverse transcription-quantitative polymerase chain reaction. Furthermore, we did not investigate transplantation methods using Muse cells for osteochondral lesions other than transplantation as spheroids.
In conclusion, the implantation of MSCs, including Muse and non-Muse cells, resulted in better healing of osteochondral lesions in the mice models than in the controls, and in vitro, the aggregates of the Muse cells as a spheroid had a higher chondrogenic differentiation potential than that of non-Muse cells. Potentially, one could induce in vivo cartilage regeneration by using Muse cells as somatic in vivo. To investigate effective transplantation methods, further studies are needed to treat a larger animal model of cartilage defects and to include the use of a scaffold or coating method to keep the cells in the injured location.
Supplemental Material
sj-pdf-1-car-10.1177_19476035241262020 – Supplemental material for Investigating the Potential of Multilineage Differentiating Stress-Enduring Cells for Osteochondral Healing
Supplemental material, sj-pdf-1-car-10.1177_19476035241262020 for Investigating the Potential of Multilineage Differentiating Stress-Enduring Cells for Osteochondral Healing by Shotaro Watanabe, Hiroaki Hosokawa, Takuya Sakamoto, Manato Horii, Yoshimasa Ono, Seiji Kimura, Satoshi Yamaguchi, Seiji Ohtori and Takahisa Sasho in CARTILAGE
Footnotes
Authors’ Note
Location: The work reported was done at Chiba University.
Acknowledgment and Funding
The authors would like to thank Masahiro Kiuchi for technical assistance with the experiments. They also thank Editage (![]() ) for English language editing. The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by JSPS KAKENHI (grant no. JP22K16705), JST SPRING, Japan (grant no. JPMJSP2109), Grant of the Japanese Orthopaedic Society of Knee, Arthroscopy and Sports Medicine, 2019, and Grants from Chiba Foundation for Health Promotion Disease Prevention.
) for English language editing. The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by JSPS KAKENHI (grant no. JP22K16705), JST SPRING, Japan (grant no. JPMJSP2109), Grant of the Japanese Orthopaedic Society of Knee, Arthroscopy and Sports Medicine, 2019, and Grants from Chiba Foundation for Health Promotion Disease Prevention.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The research protocol for this study was approved by the Institutional Review Board and complied with the ARRIVE guidelines.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.