
Abstract
Self-enhancement, the motive to view oneself in positive light, and its manifestations have received wide attention in behavioral sciences. The self-enhancement manifestations vary on a continuum from a subjective level (agentic narcissism, communal narcissism, narcissistic grandiosity) through an intermediate level (better-than-average judgments) to an objective level (overclaiming one’s knowledge). Prior research has established the heritability of self-enhancement manifestations at the subjective and intermediate levels. The present twin study demonstrated that (1) the objective level of self-enhancement manifestation is also heritable; (2) a common core, which is moderately heritable, underlies the three levels of self-enhancement manifestations; (3) the relation between self-enhancement (manifested at all three levels) and psychological well-being is partly heritable; and (4) environmental influences, either shared by or unique to family members, are evident through (1), (2), and (3). The findings deepen understanding of the etiology of individual differences in self-enhancement and their links to psychological well-being.
Keywords
People are motivated, albeit to a varying degree, to view themselves in a positive light. This motive is known as self-enhancement and works to nourish one’s self-regard (Sedikides & Gregg, 2008). One of several self-evaluation motives (Sedikides & Strube, 1997), self-enhancement has received wide attention in the social/personality, clinical, developmental, and neuroscience literature (Alicke & Sedikides, 2011), as well as in the education (Liem & McInerney, 2018), sports science (Matosic et al., 2017), and organizational behavior (Ferris, Johnson, & Sedikides, 2018) literature. A key reason for this attention is the psychosocial benefits to which the motive conduces such as well-being, goal pursuit, leadership selection, and sexual selection (Sedikides, 2018). In an effort to uncover etiological similarity (or dissimilarity) underlying various self-enhancing manifestations and their connections with psychosocial benefits, we engaged in a behavioral genetic analysis of individual differences in self-enhancement and the covariation between self-enhancement and well-being.
Self-Enhancement: Manifestations and Individual Differences
Manifestations of self-enhancement vary on a subjective-to-objective continuum. At the subjective level, the motive is manifested in terms of endorsement of attributes or statements that flatter the self. These could reflect agentic narcissism (believing that one is an authoritative, entitled, and special figure, while taking pride in one’s vanity and exhibitionism), communal narcissism (believing that one is a saintly figure, likely to be super helpful or caring, and the beacon of world justice and peace), or narcissistic grandiosity (believing that one is endowed with a set of glowing traits such as extraordinary, brilliant, honorable, and acclaimed). Note that, when individuals endorse these attributes or statements, they do not compare the self directly with a referent, although a referent is implied (Biernat, 2005). At the intermediate level, the self-enhancement motive is manifested in terms of favorable comparisons of the self to a referent. In the case of the better-than-average task (BTAT; Alicke, 1985), for example, participants compare themselves to their typical or average peer on a variety of traits. Finally, at the objective level, the presence of an impartial standard is required. This standard is often met by the so-called criterion-discrepancy measures such as the overclaiming task (Paulhus, Harms, Bruce, & Lysy, 2003). Here, participants manifest self-enhancement when they claim knowledge about bogus test items. Yet, regardless of the level at which it is manifested (subjective, intermediate, or objective), self-enhancement is associated with, or confers, psychological well-being (Dufner, Gebauer, Sedikides, & Denissen, 2019).
Although the self-enhancement motive and its manifestations are generally potent (Alicke, Zell, & Guenther, 2013) and also prevalent cross-culturally (Sedikides, Gaertner, & Cai, 2015), they nevertheless vary among individuals (Foster, Campbell, & Twenge, 2003; Hepper, Gramzow, & Sedikides, 2010; Hepper, Sedikides, & Cai, 2013). Such individual differences are partly due to parental practices (Brummelman et al., 2015; Thomaes, Brummelman, & Sedikides, 2017) and are embedded within hormone systems (Cheng, Tracy, & Miller, 2013; Reinhard, Konrath, Lopez, & Cameron, 2012) and neural networks (Cai, Wu, Shi, Gu, & Sedikides, 2016; Yang et al., 2018a, 2018b) in adults.
Recently, research has started to focus on the etiological origins of individual differences in self-enhancement. Twin studies on agentic narcissism (Luo, Cai, Sedikides, & Song, 2014), communal narcissism (Luo, Cai, Sedikides, et al., 2014), narcissistic grandiosity (Luo, Cai, & Song, 2014), and the better-than-average effect (BTAE—considering oneself superior to peers; Luo, Liu, Cai, Wildschut, & Sedikides, 2016) have revealed that individual differences in self-enhancement manifestations have genetic bases both at the subjective and intermediate levels. Environmental influences also contribute to the variance in self-enhancement manifestations at both levels. However, no twin study has examined the extent to which self-enhancement manifestations at the objective level are heritable. Likewise, no study has examined whether individual differences in self-enhancement manifestations at different levels are underpinned by the same or different genetic sources. The same goes for studies on environmental influences.
Overview
We aimed to uncover the genetic and environmental bases underlying individual differences in objective-level self-enhancement by using the overclaiming task. We had two other key objectives. For the first time, we incorporated simultaneously manifestations of self-enhancement at all three levels (subjective, intermediate, and objective) in an attempt to probe their etiologic commonality. Is there a core construct underlying the three levels, and, if so, to what extent is it heritable or environmentally shaped? Also for the first time, we examined whether the (cross-cultural; Dufner et al., 2019) relation between self-enhancement and psychological well-being—at the subjective, intermediate, and objective levels—was genetically, as well as, environmentally, influenced.
Agentic and communal narcissism, narcissistic grandiosity, the BTAE, and overclaiming are moderately interrelated on phenotypic levels (Crowe, Carter, Campbell, & Miller, 2016; Gebauer, Sedikides, Verplanken, & Maio, 2012; Luo, Cai, Sedikides, et al., 2014; Paulhus et al., 2003). As phenotypic covariance usually indicates genetic and/or environmental similarity (Knopik, Neiderhiser, DeFries, & Plomin, 2017), we expected that common genetic and environmental factors would contribute to variations in all these self-enhancement manifestations besides genetic and environmental influences unique to each manifestation. We tested two competing hypothetical models, the independent pathway model and the common pathway model. Both models acknowledge common and unique genetic as well as environmental effects on each manifestation. However, the independent pathway model assumes that some common genetic and environmental factors influence each manifestation through pathways that are independent from those influencing other manifestations, whereas the common pathway model assumes that common genes or environments influence all manifestation via the same pathway, namely, a latent trait. This latent trait stands for the core of manifestations. If all manifestations converge into one core, the common pathway model would outperform the independent pathway model.
To examine potential genetic and environmental bases between self-enhancement manifestations and psychological well-being, we focused on both the cognitive and affective components of well-being (Diener, Emmons, Larsen, & Griffin, 1985; Diener, Suh, Lucas, & Smiths, 1999). Self-enhancement manifestations (in general) and psychological adjustment (including well-being) are positively related (r = .18), as a meta-analysis established (Dufner et al., 2019). As such, we hypothesized that the genetic effects on self-enhancement and those on well-being would be positively associated. Stated otherwise, self-enhancement and well-being will share similar genetic bases to some degree. We formulated similar hypothesis for environmental effects.
Method
Participants
We recruited twin pairs from the Beijing Twin Study (BeTwiSt). Twins in the BeTwiSt are sociodemographically representative of Beijing adolescents (Chen et al., 2013). To ensure the measures were accessible to all participants, we only invited twins who had finished middle school by the time of testing. Eventually, 304 twin pairs agreed to participate (55.9% female; age: range = 15–27 years, M = 18.29, SD = 1.96). Of them, 152 pairs were monozygotic (MZ), and 152 pairs were dizygotic (DZ; 94 same sex, 58 opposite sex). We used DNA testing, with classification accuracy approaching 100%, to determine zygosity for 95% of the twin pairs. For the remaining 5% of twin pairs, we established zygosity with a combination of parent reports and children self-reports of co-twin physical similarity and frequency of confusion—tests that feature predictive accuracy of 90.6% (Chen et al., 2010). We excluded a DZ twin pair due to missing data. We obtained ethical approval from the ethics committee of Institute of Psychology, Chinese Academy of Sciences. We collected written consent from the twins and their parents before we began testing.
Measures
Participants completed measures of the abovementioned self-enhancement manifestations and psychological well-being components (along with measures irrelevant to the purposes of this research) on a computer in a private room. We used translation and back-translation procedures to ensure language equivalence. We provide Cronbach’s αs for all measures in Table 1.
Reliabilities of, Means (SD) of, and Zero-Order Correlations Among All Measures.
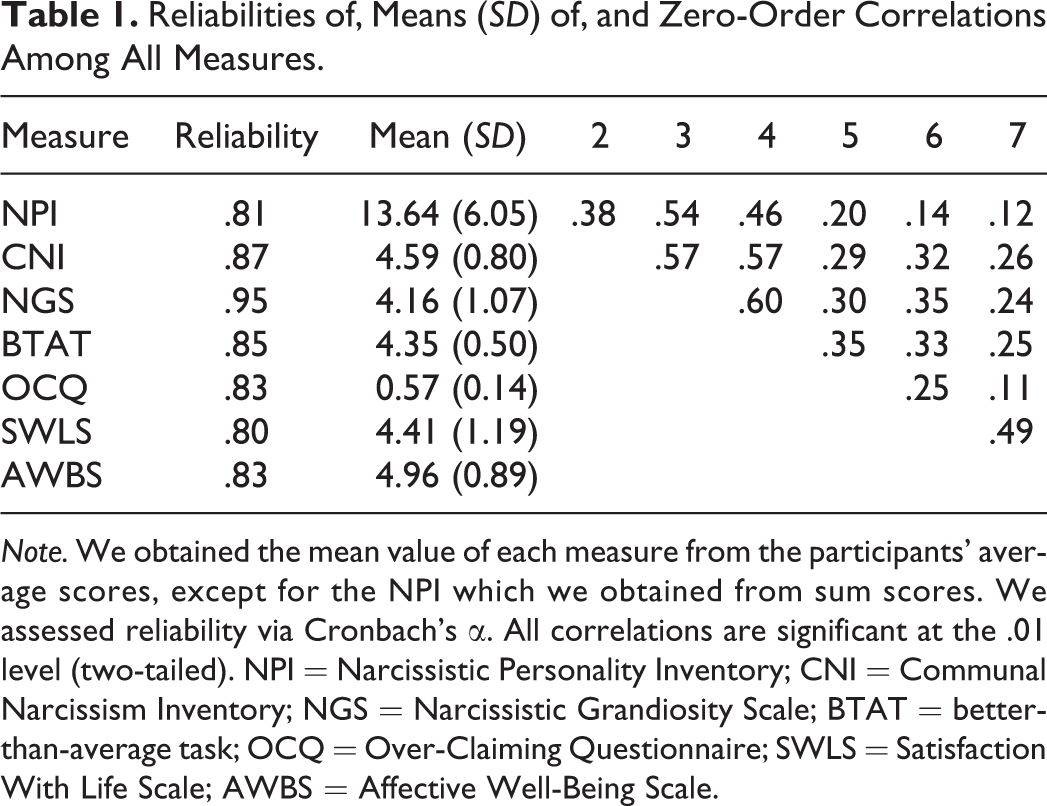
Note. We obtained the mean value of each measure from the participants’ average scores, except for the NPI which we obtained from sum scores. We assessed reliability via Cronbach’s α. All correlations are significant at the .01 level (two-tailed). NPI = Narcissistic Personality Inventory; CNI = Communal Narcissism Inventory; NGS = Narcissistic Grandiosity Scale; BTAT = better-than-average task; OCQ = Over-Claiming Questionnaire; SWLS = Satisfaction With Life Scale; AWBS = Affective Well-Being Scale.
Self-enhancement
We measured agentic narcissism with the 40-item Narcissistic Personality Inventory (NPI; Raskin & Terry, 1988). Each item includes a pair of statements, one narcissistic (e.g., “I have a natural talent for influencing people,” “I like to look at myself in the mirror”) and one nonnarcissistic (e.g., “I am not good at influencing people,” “I am not particularly interested in looking at myself in the mirror”). For each item, participants indicate which statement describes them better. We coded agreements with the narcissistic statements as 1 and with the nonnarcissistic statements as 0, and summed up the scores for each participant.
We measured communal narcissism with the 16-item Communal Narcissism Inventory (CNI; Gebauer et al., 2012). Sample items are “I am the most helpful person I know,” “I am going to bring peace and justice to the world” (1 = disagree strongly, 7 = agree strongly). We calculated the mean score for each participant.
We measured narcissistic grandiosity with the Narcissistic Grandiosity Scale (NGS; Rosenthal, Hooley, & Steshenko, 2007; see also Crowe et al., 2016). The NGS consists of 16 trait adjectives (advanced, admirable, brave, enviable, excellent, extraordinary, honorable, omnipotent, outstanding, perfect, powerful, prestigious, respectable, talented, unique, and vigorous) responded to on a scale from 1 (not at all) to 7 (extremely). Higher ratings indicate better applicable to participants. We averaged scores across the 16 items.
We measured the BTAE effect with a modified version of the BTAT (Alicke, 1985), that is, by asking participants to compare themselves with the average person of the same age and sex on 16 traits: attractive, capable, considerate, cooperative, creative, easygoing, friendly, gifted, independent, kind, loyal, polite, self-reliant, sincere, smart, unique (1 = much less than the average person, 6 = much more than the average person). We averaged ratings into an aggregate score.
Finally, we measured overclaiming with the Over-Claiming Questionnaire (OCQ; Paulhus et al., 2003). Participants rated their familiarity with items belonging to seven categories: art works, celebrities, classical literature, famous buildings, historical events, leading intellectuals, national government (1 = never heard of it, 7 = know it very well). Each category comprised 2 real items and 1 foil. Claiming knowledge for foils reflects self-enhancement, also called in this context “response bias.” We calculated a bias index using standard signal detection formulas (Macmillan & Creelman, 1991). We calculated indices at each of the six cutoffs on the 1–7 rating scale (i.e., 1, 2, 3, 4, 5, 6). The hit rate was the proportion of the 14 real items on which a participant gave a rating above the cutoff. Similarly, the false-alarm rate was the corresponding proportion of the seven foils on which the participant gave a rating above the cutoff. We first set the cutoff as ratings above 1, yielding an overall hit rate and a false-alarm rate. The bias was the mean of the hit rate and the false-alarm rate. We repeated these calculations for the other five cutoffs and averaged the six values of bias to form the final value of OCQ bias for each participant.
Psychological well-being
We assessed the cognitive component of well-being with the 5-item Satisfaction With Life Scale (SWLS; Diener et al., 1985). A sample item is “In most ways my life is close to my ideal” (1 = strongly disagree, 7 = strongly agree). We used mean scores as the index of life satisfaction. We assessed the affective component of well-being with the 8-item Affective Well-Being Scale (AWBS; Diener et al., 2010). Participants rated the frequency with which they experienced negative (unpleasant, sad, angry, afraid) and positive (happy, pleasant, joyful, contented) affect (1 = never, 7 = always). We reversed the negative affect scores and obtained a mean score across the 8 items as the index of affective well-being.
Genetic Analyses
In principle, by comparing the resemblance of MZ and DZ twin pairs on observed trait(s), we can estimate additive genetic (A), shared environmental (C), and nonshared environmental (E) contributions to (1) variance within a trait and (2) covariance between traits (Knopik et al., 2017). MZ twins are 100% genetically identical, whereas DZ twins are on average 50% identical for additive genetic effects. In the usual case where twins are reared together, greater resemblance between MZ twins than between DZ twins indicates that the trait is heritable. Heritability denotes the proportion of the variance of a trait, or the covariance between traits, explained by additive genetic effects. A shared environment contributes to the similarity of twins growing up in the same family. A nonshared environment is unique to each individual (this component also includes measurement error).
More sophisticated analyses are possible through structural equation modeling (SEM). In order to examine how genes and environments contributed to all five self-enhancement manifestations in simultaneity, we tested two models: an independent pathway model and a common pathway model (Young, Stallings, Corley, Krauter, & Hewitt, 2000). The independent pathway model (Figure 1) partitions the variances and covariances of the five self-enhancement measures into two components: (1) common variance (Ac, Cc, Ec), which represents the variance that is shared by all measures and (2) unique variance (A1-5, C1-5, E1-5), which represents the variance that is not shared between the measures. This model posits that the variation common to the five self-enhancement measures is influenced by a set of common genetic and environmental factors, whereas the variation unique to each measure is influenced by specific genetic and environmental factors.
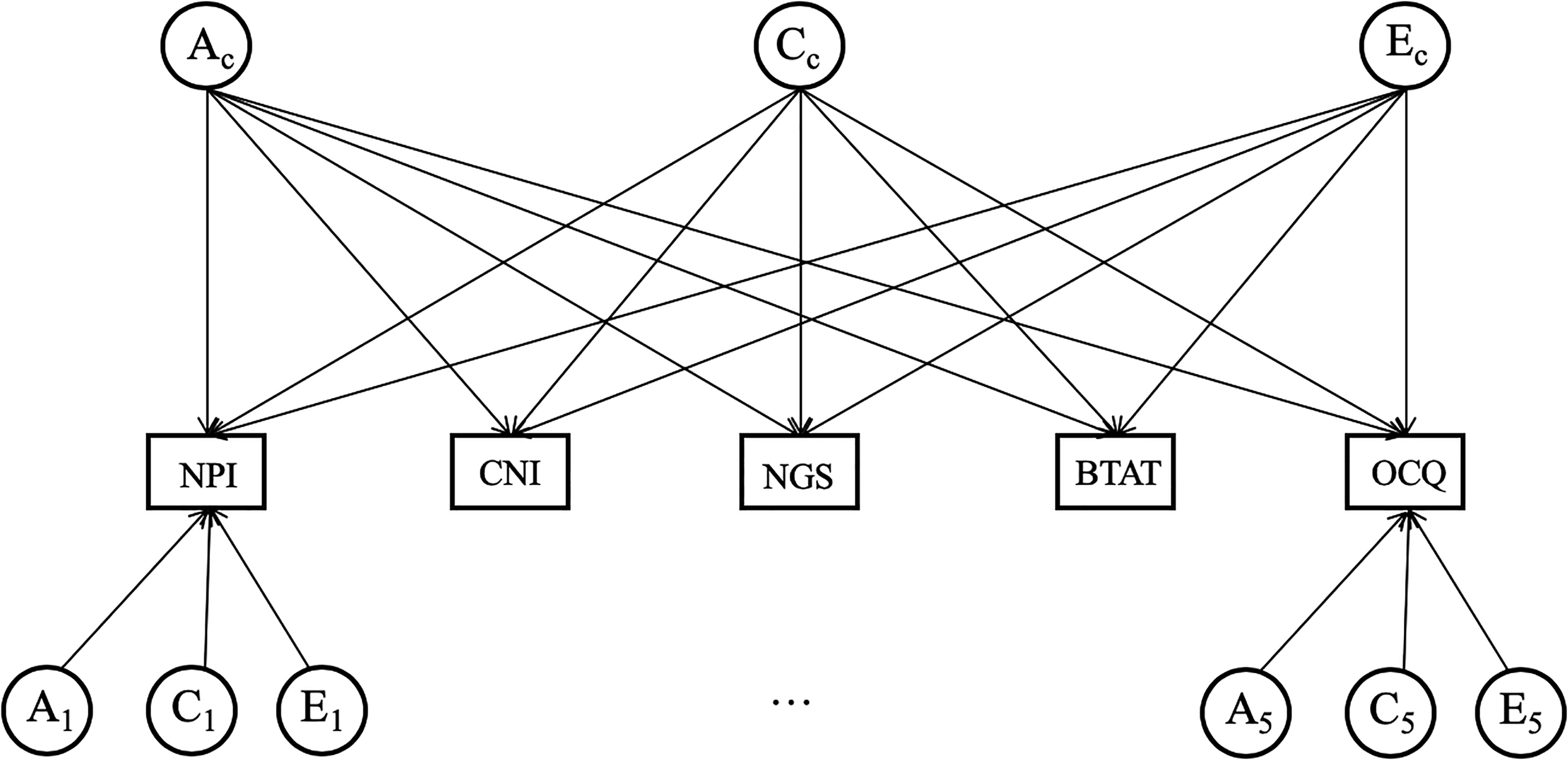
An illustration of the independent pathway model. We depict measured variables in rectangles. NPI = Narcissistic Personality Inventory; CNI = Communal Narcissism Inventory; NGS = Narcissistic Grandiosity Scale; BTAT = better-than-average task; OCQ = Over-Claiming Questionnaire. We present latent factors A (additive genetic factor), C (shared environmental factor), and E (nonshared environmental factor) in circles. Ac, Cc, and Ec are factors influencing all measures. A1-5, C1-5, and E1-5 are factors unique to each measure.
In contrast, the common pathway model (Figure 2) posits that genetic and environmental factors influencing variation common to the five measures do not affect the measures directly; rather, they impact on each measure via a latent factor. This factor represents the underlying common variation across measures of self-enhancement. Stated differently, this model extracts the overlap among all measures, which represents the commonality in self-enhancement reflected by different measures. This model is nested in independent model and more parsimonious because it estimates fewer parameters (Briley & Tucker-Drob, 2012).
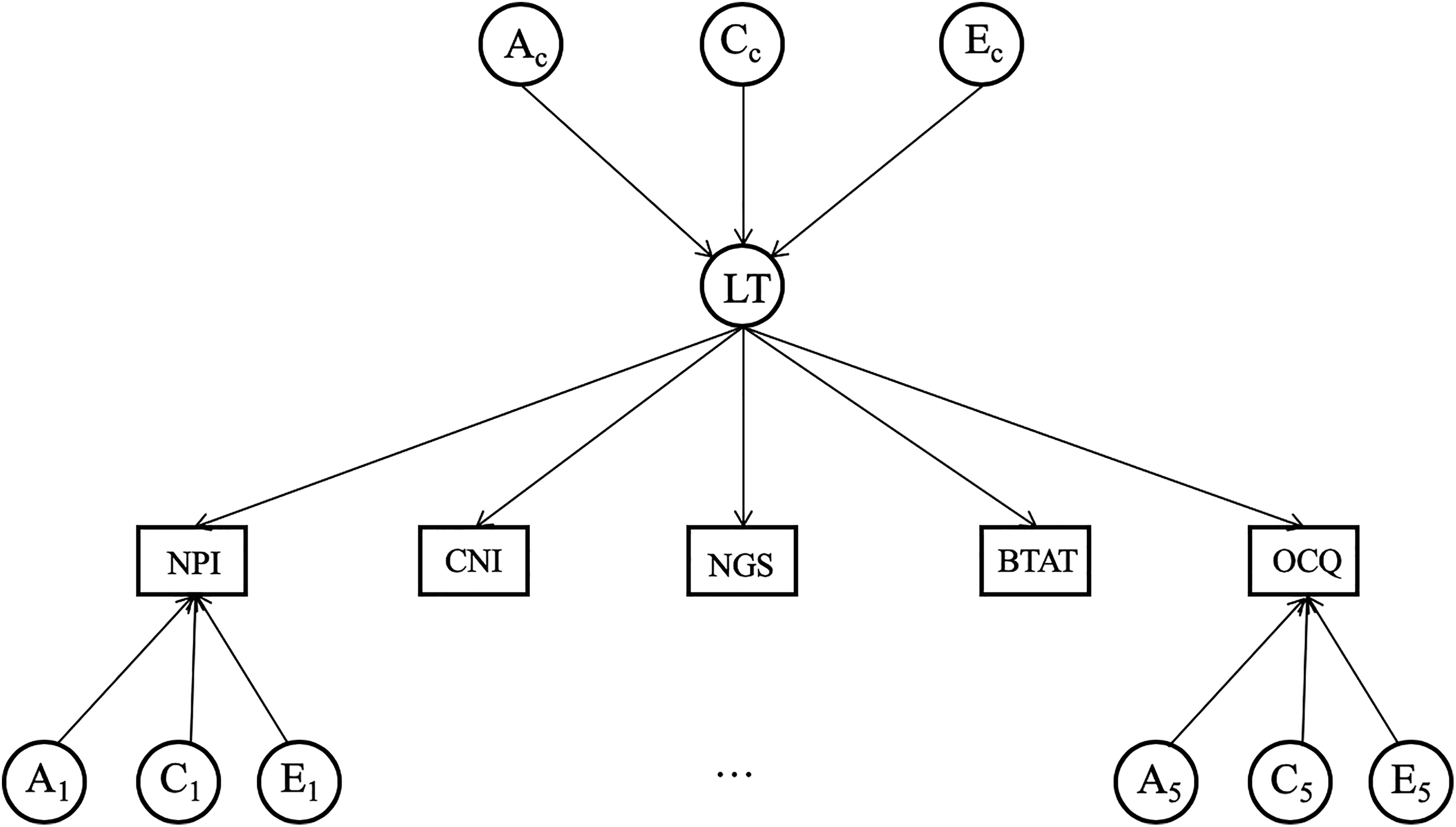
An illustration of the common pathway model. We depict measured variables in rectangles. NPI = Narcissistic Personality Inventory; CNI = Communal Narcissism Inventory; NGS = Narcissistic Grandiosity Scale; BTAT = better-than-average task; OCQ = Over-Claiming Questionnaire. We present latent trait (LT), A (additive genetic factor), C (shared environmental factor), and E (nonshared environmental factor) in circles. Ac, Cc, and Ec are factors influencing the latent trait for self-enhancement. A1-5, C1-5, and E1-5 are factors unique to each measure.
We used three statistics to guide model selection. The first one was the change in χ2. When two models are nested, the difference in model fit (i.e., twice the negative log-likelihood, −2LL) between them can be evaluated via the χ2 difference, with the degree of freedom (df) difference of the two models as the df. A nonsignificant χ2 difference suggests that the more parsimonious model should be chosen because including additional constraints or parameters does not improve the model fit (Bollen, 1989). The second and third statistics were the Akaike’s (1987) information criterion (AIC) and the Bayesian information criterion (BIC; Raftery, 1995). For both indices, a lower value indicates better fit. In particular, differences of BIC value between 6 and 10 provide strong evidence in favor of the model with the smaller value (Raftery, 1995). We carried out genetic analyses with Open Mx package (Version 2.6.9; Boker et al., 2012) for R (Version 3.2.5).
Statistical Power for Genetic Analyses
In order to gauge whether we had sufficient power to detect genetic or environmental effects, we estimated the minimum effect size that could be reliably observed within the current sample. We implemented the power analysis via the package “pwr” (Version 1.2.0) in R (Champely, 2018), which functions along the lines of Cohen (1988). Given a sample size of N = 303 (the total number of twin pairs; Table 2), level of significance at .10, 1 dfs at 1 (estimating one effect each time), and power at the conventional level of .80, we could detect effects whose size (w) was no less than .14. 2 Put otherwise, if the standardized estimate of a path parameter in the independent pathway model or common pathway model (e.g., Ac to NPI in Figure 1 or Ac to LT in Figure 2) were no less than .14, we would have sufficient power to examine the corresponding genetic or environmental effects.
Twin Intraclass Correlations (ICC) for All Measures.
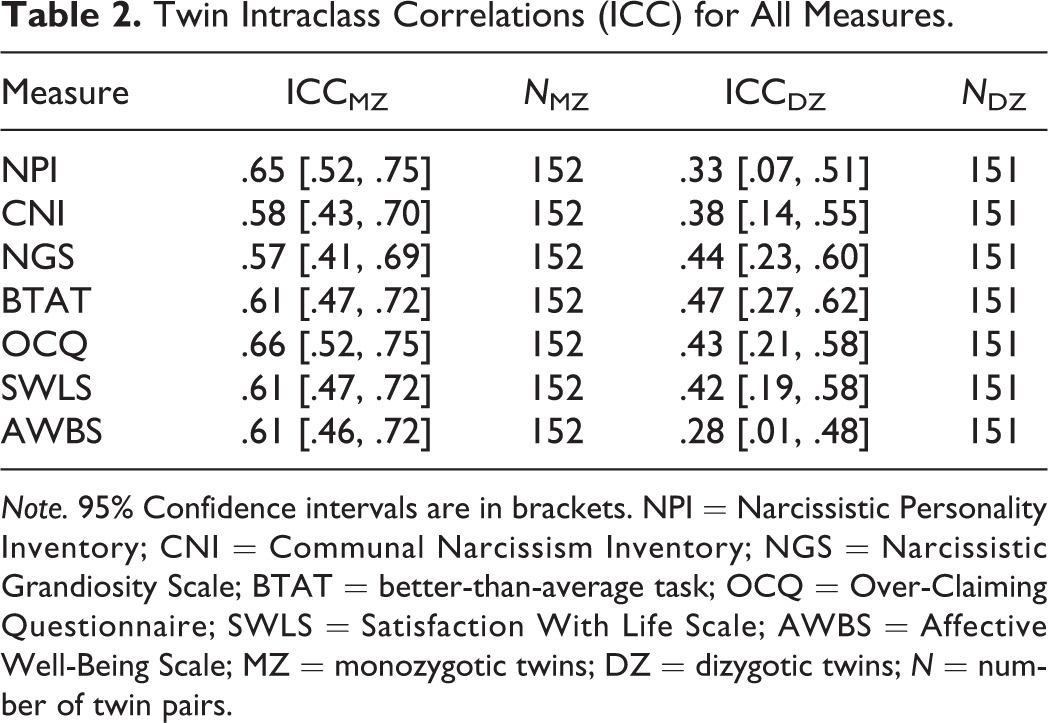
Note. 95% Confidence intervals are in brackets. NPI = Narcissistic Personality Inventory; CNI = Communal Narcissism Inventory; NGS = Narcissistic Grandiosity Scale; BTAT = better-than-average task; OCQ = Over-Claiming Questionnaire; SWLS = Satisfaction With Life Scale; AWBS = Affective Well-Being Scale; MZ = monozygotic twins; DZ = dizygotic twins; N = number of twin pairs.
Beyond testing genetic and environmental effects, we engaged in a comparison between the independent and common pathway models. We wondered whether we had adequate power for that comparison. This analysis is different from the former one, with the exception of one step (the first step estimating the minimal effect size w). First, we estimated the smallest effect size that could be detected within the current sample (N = 303). Given a level of significance at .10, df = 8 (the difference in df between the two models; Table 3), and power at .80, the minimal effect that we could assess with confidence was w = .20. Then, based on that value, we estimated the threshold of test statistic—that is, χ2—according to the formula χ2 = w2 × N. We obtained a χ2 of 12.12. Thus, if the difference in model fit (i.e., −2LL) between the independent and common pathway models were no less than 12.12, we would have adequate power for model comparison.
Multivariate Genetic Model-Fitting for Self-Enhancement Manifestations: Model Fit Statistics.
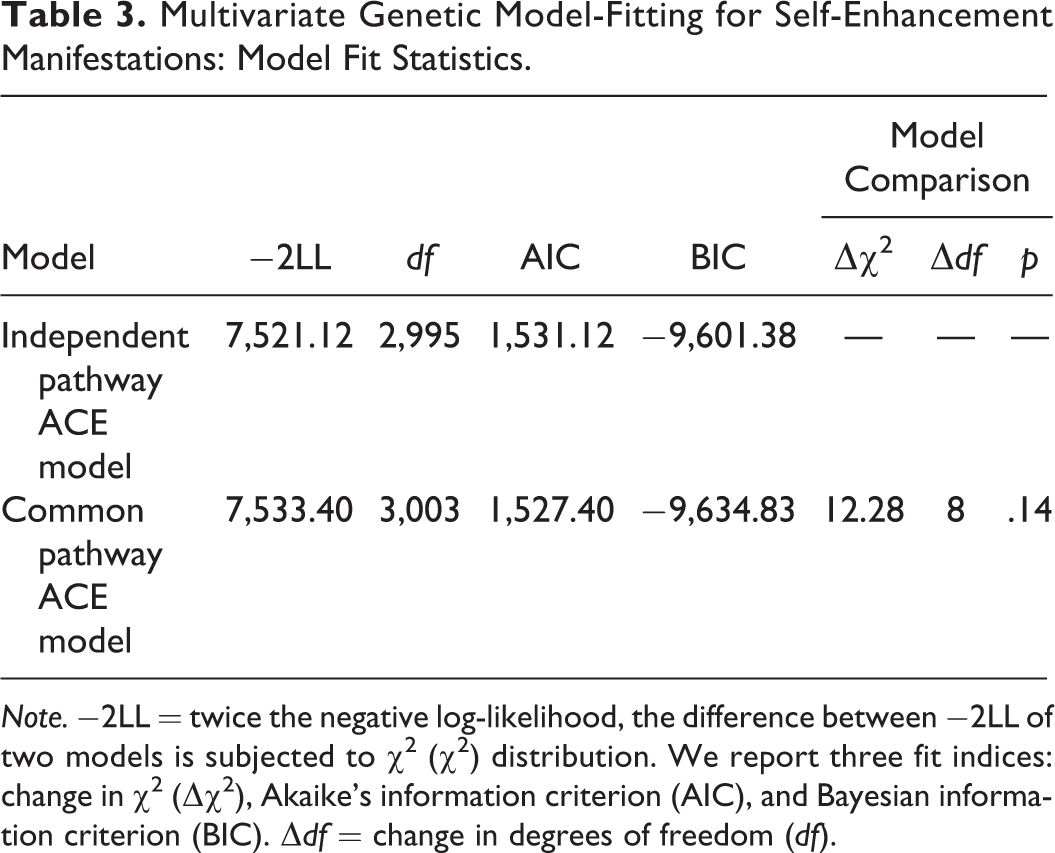
Note. −2LL = twice the negative log-likelihood, the difference between −2LL of two models is subjected to χ2 (χ2) distribution. We report three fit indices: change in χ2 (Δχ2), Akaike’s information criterion (AIC), and Bayesian information criterion (BIC). Δdf = change in degrees of freedom (df).
Results
Descriptive Statistics
In Table 1, we present correlations, means, and standard deviations for all measures. All correlations, no matter within or across domains (i.e., self-enhancement manifestations or well-being), were significant, ranging from moderate to high (rs: .11∼.60).
Given that twins are perfectly correlated for age, same-sex twins are perfectly correlated for sex, and variations associated with age and/or sex would inflate the correlation between twins, we (1) regressed each measure onto age, sex, age 2 , and age × sex, respectively, and (2) saved standardized residuals for genetic analyses (McGue & Bouchard, 1984). In Table 2, we present the intraclass correlations for all seven measures. MZ twins resembled each other more than DZ twins did on every self-enhancement or well-being measure, suggesting that these indicators were all heritable.
Multivariate Genetic Model-Fitting for Self-Enhancement Manifestations
We examined genetic and environmental influences on individual differences in self-enhancement manifestations by testing two hypothetical genetic models, the independent pathway model and the common pathway model (Figures 1 and 2). 3 Compared to the common pathway model, the independent pathway provides a less constrained account of the patterns of global and specific genetic and environmental effects on self-enhancement manifestations (Table 3). Based on parameter estimates from the independent pathway ACE model (Table 4), we noticed the following about the genetic and environmental structure of the five self-enhancement indicators. First, they shared some genetic influence, but to different extent (.22∼.63). Second, some of them (NPI, BTAT, OCQ) were also influenced by genetic effects unique to each. Third, although certain nonshared environments influenced all self-enhancement manifestations (.16∼.61), the nonshared environmental influence on these manifestations was substantially due to specific factors (.52∼.70). Finally, the shared environmental influence on all self-enhancement manifestations was largely in common (.11∼.45), except that NGS was also affected by a distinct shared environment.
Standardized Estimates of Model Parameters From the Independent Pathway Model of Self-Enhancement Manifestations.
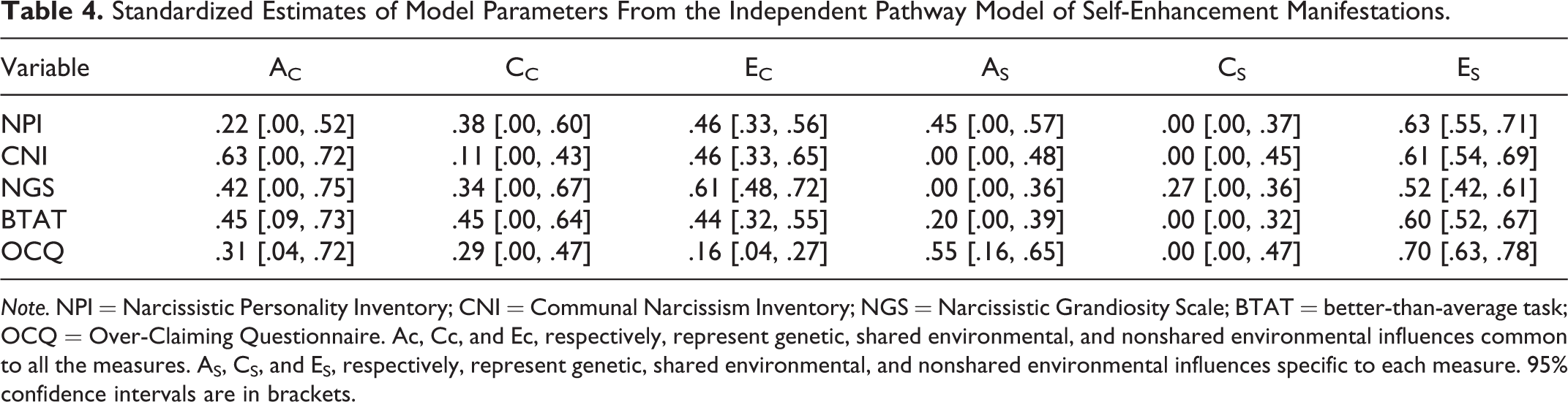
Note. NPI = Narcissistic Personality Inventory; CNI = Communal Narcissism Inventory; NGS = Narcissistic Grandiosity Scale; BTAT = better-than-average task; OCQ = Over-Claiming Questionnaire. Ac, Cc, and Ec, respectively, represent genetic, shared environmental, and nonshared environmental influences common to all the measures. AS, CS, and ES, respectively, represent genetic, shared environmental, and nonshared environmental influences specific to each measure. 95% confidence intervals are in brackets.
As we proposed earlier, the five self-enhancement manifestations might converge onto one latent trait whose genetic and environmental bases would further contribute to each manifestation. To examine this possibility, we designed a common pathway model and tested whether it accounted for the data better than the independent pathway model. As common pathway models are nested in independent pathway models (Briley & Tucker-Drob, 2012), we conducted a χ2 difference test and found that the more parsimonious common pathway model fitted the data equally well with the independent pathway model did (p > .05; Table 3). Further, the χ2 difference in model fit between the two models (12.28) was just above the threshold (12.12) set in the power analysis. Therefore, we had enough power in comparing the common pathway model against the independent pathway model. In addition, the common pathway model had lower AIC and BIC values (Table 3). Of note, the difference in BIC between the two models was greater than 10. Hence, we opted for the common pathway model.
According to parameter estimates of this model (Table 5), all five self-enhancement manifestations reflected some part of a latent trait as indicated by moderate to large factor loadings on the measures (.40∼.81). Moreover, the analysis revealed that the variation in the latent trait was mainly accounted for by genetic (42%) and nonshared environmental (42%) factors, with the remaining part (16%) accounted for by shared environments. Further, we decomposed the individual differences in every self-enhancement measure into common and unique effects from genes, shared environments, and nonshared environments (Table 5). The following results emerged. First, common genetic effects (via the latent trait) accounted for part of the heritability of all measures, whereas specific genetic effects mattered for only three of these measures (NPI, BTAT, OCQ). Second, both global and specific nonshared environmental effects contributed to variation in each self-enhancement manifestation, though the specific ones exerted more influence. Third, the moderate shared environmental effects on the five manifestations were largely in common, albeit specific shared environmental effects still gave rise to some variance in two measures (CNI, NGS). Notably, these results were comparable to those from the independent pathway model. In addition, our finding that variations in self-enhancement latent trait and manifestations were mainly due to genes and nonshared environments, and secondarily due to shared environments, was consistent with previous findings about self-enhancement manifestations on subjective and intermediate levels (Luo, Cai, Sedikides, et al., 2014; Luo, Cai, & Song, 2014; Luo et al., 2016).
Standardized Estimates of Model Parameters, as well as Genetic and Environmental Effects, From the Common Pathway Model of Self-Enhancement Manifestations.
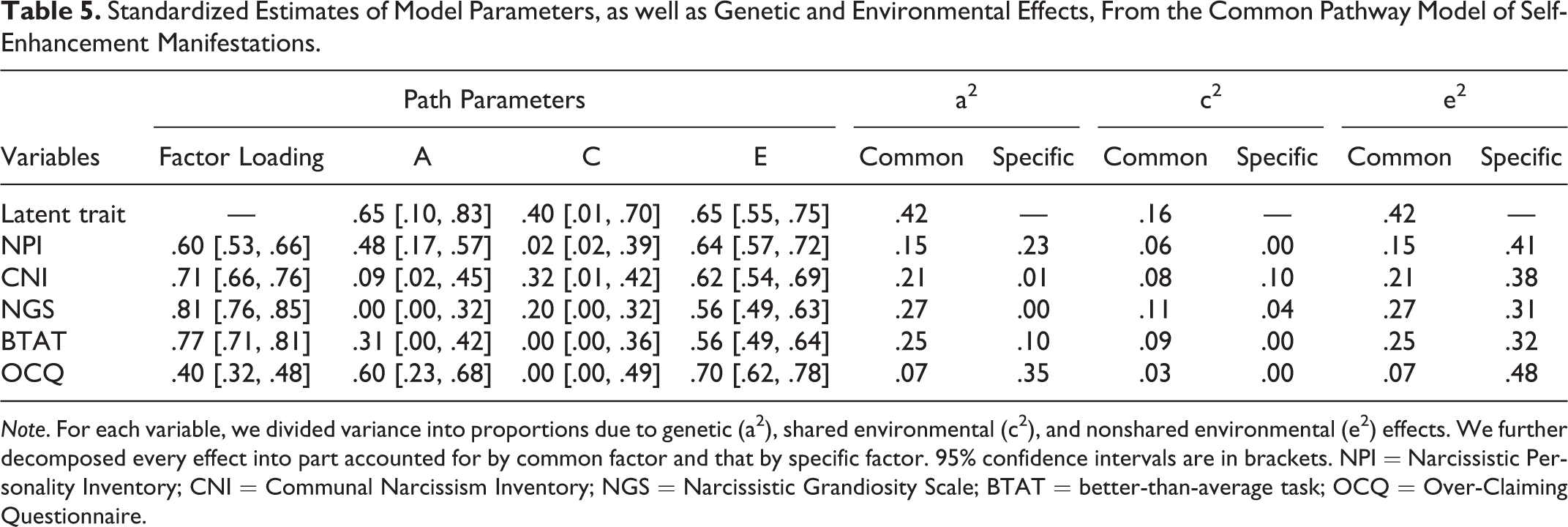
Note. For each variable, we divided variance into proportions due to genetic (a2), shared environmental (c2), and nonshared environmental (e2) effects. We further decomposed every effect into part accounted for by common factor and that by specific factor. 95% confidence intervals are in brackets. NPI = Narcissistic Personality Inventory; CNI = Communal Narcissism Inventory; NGS = Narcissistic Grandiosity Scale; BTAT = better-than-average task; OCQ = Over-Claiming Questionnaire.
Multivariate Genetic Model-Fitting for Self-Enhancement Manifestations and Psychological Well-Being
To examine the genetic and environmental contributions to the relation between self-enhancement and well-being, we extended the previous common pathway model by incorporating another group of parameters for well-being as we did for self-enhancement manifestations (Figure 3). We arrived at this practice based on previous findings of common genetic and environmental influences on cognitive and affective well-being, especially given that the common influences are attributed to latent factors underlying the two well-being indicators (Franz et al., 2012; Haworth, Carter, Eley, & Plomin, 2017). Furthermore, we tested whether the genetic influences common to the self-enhancement manifestations and those common to the well-being indicators were interconnected (i.e., a genetic correlation between the two common genetic factors, rA). Similarly, we examined shared and nonshared environmental correlations (rC, rE) between corresponding common factors.
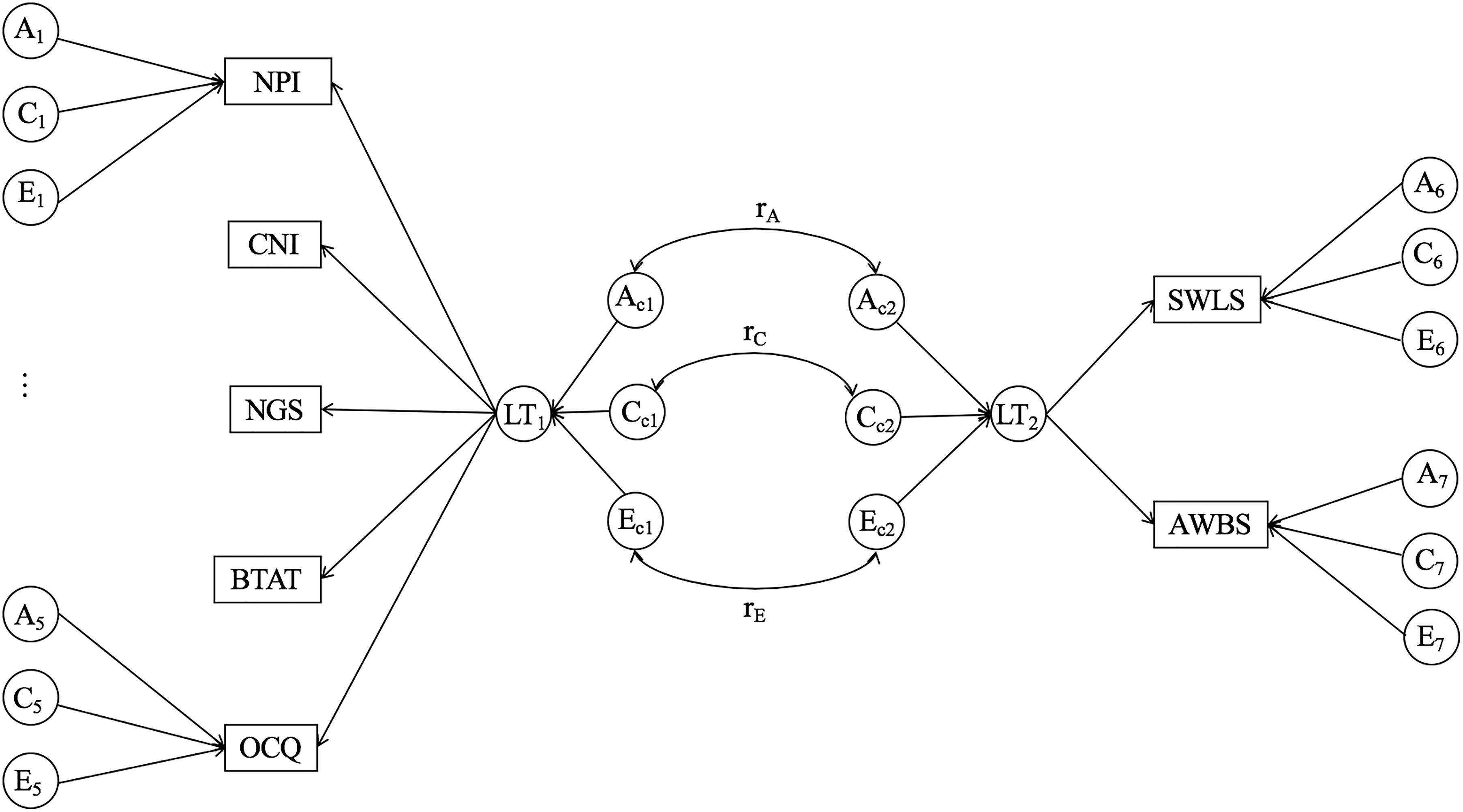
An illustration of the common-pathway model incorporating both self-enhancement and psychological well-being. We depict measured variables in rectangles. NPI = Narcissistic Personality Inventory; CNI = Communal Narcissism Inventory; NGS = Narcissistic Grandiosity Scale; BTAT = better-than-average task; OCQ = Over-Claiming Questionnaire; SWLS = Satisfaction With Life Scale; AWBS = Affective Well-Being Scale. We present latent traits (LT1, LT2), A (additive genetic factor), C (shared environmental factor), and E (nonshared environmental factor) in circles. Ac1-2, Cc1-2, and Ec1-2 are factors influencing latent variables. A1-7 and E1-7 are factors specific to each measure.
According to the full ACE model (−2LL = 10,617.91, df = 4,200, AIC = 2,217.91, BIC = −13,393.61), the results concerning self-enhancement manifestations per se replicated the findings from the common pathway model targeting self-enhancement manifestations (Table 6). For well-being, we found moderate genetic (.39) and nonshared environmental (.49) effects as well as small shared environmental effects (.12) on the latent trait for well-being. Such effects influenced both well-being measures (Table 6). Crucially, we found that the global genetic influence underlying self-enhancement manifestations was positively related to that underlying two well-being components (rA = 0.32, 95% confidence intervals (CI) = [0.02, 0.84]). The shared and nonshared environmental influences on self-enhancement and well-being were also related (rC = 1.00 [1.00, 1.00]; rE = .58 [0.38, 0.75]).
Standardized Estimates of Model Parameters, as well as Genetic and Environmental Effects From the Common Pathway Models, Incorporating Self-Enhancement Manifestations and Psychological Well-Being.
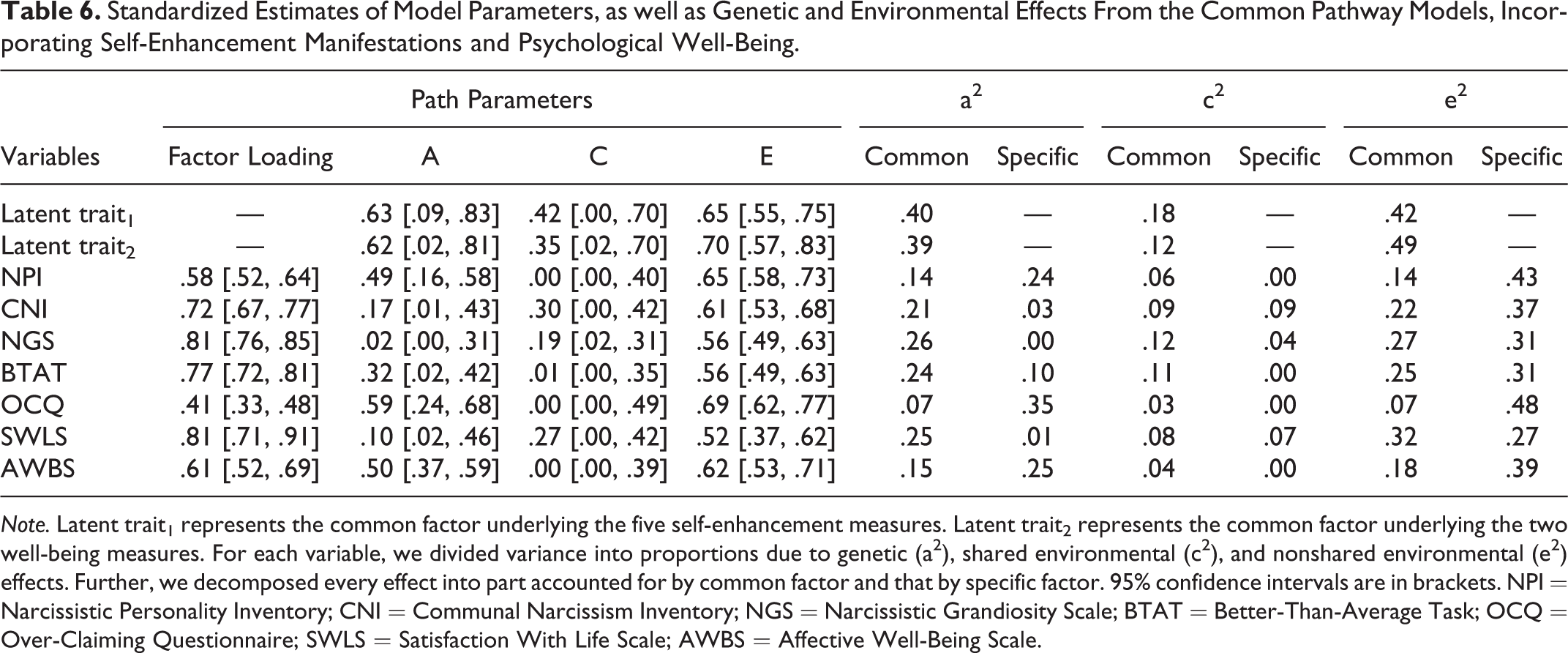
Note. Latent trait1 represents the common factor underlying the five self-enhancement measures. Latent trait2 represents the common factor underlying the two well-being measures. For each variable, we divided variance into proportions due to genetic (a2), shared environmental (c2), and nonshared environmental (e2) effects. Further, we decomposed every effect into part accounted for by common factor and that by specific factor. 95% confidence intervals are in brackets. NPI = Narcissistic Personality Inventory; CNI = Communal Narcissism Inventory; NGS = Narcissistic Grandiosity Scale; BTAT = Better-Than-Average Task; OCQ = Over-Claiming Questionnaire; SWLS = Satisfaction With Life Scale; AWBS = Affective Well-Being Scale.
Discussion
We focused in this research on the etiological bases of self-enhancement and its association with psychological well-being. Using a genetically informative design with a sample of Chinese twins, diverse measures of self-enhancement manifestations and well-being, and sophisticated genetic modeling, we identified a latent core underlying diverse manifestations of self-enhancement, from subjective to objective levels (i.e., agentic narcissism, communal narcissism, narcissistic grandiosity, BTAE, overclaiming). We further showed that this core was heritable, with some shared and nonshared environmental influences. Finally, we demonstrated that genetic, shared environmental, and nonshared environmental influences on self-enhancement manifestations overlapped with those on well-being to some extent.
Implications
Our findings have several implications. To begin, the findings improve understanding of the genetic and environmental sources of variation in self-enhancement. For the first time, we identified a latent core for self-enhancement manifestations and established its heritability. Although individuals may overestimate themselves in diverse ways, these ways are more or less sustained by the same core motive to view oneself in a positive light. Moreover, the genetic and environmental factors underlying the core also account for variance in self-enhancement manifestations, at least partly explaining why they are interconnected despite marked differences in operationalizations. To concretize these implications, some genes and environments increase hunger for self-positivity, and this motivation contributes to narcissism, boasting in social comparisons, and overclaiming one’s knowledge.
Nevertheless, we should view this implication with caution in light of alternative explanations. The latent core for self-enhancement had only a moderate influence on the objective-level manifestation (the OCQ), though its influences on the subjective- and intermediate-level manifestations were substantial. This pattern becomes more evident when we compare the global genetic or environmental influence across the three levels. The relatively low convergence on the objective level may be due to the overclaiming task operating differently than other measures (i.e., signal detection vs. self-report). Alternatively, the low convergence may be due to self-enhancing manifestations at the objective level being concurrently subjected to influences beyond the latent core identified in this study. Future research could test these explanations by implementing a more comprehensive assessment of self-enhancement along the objective–subjective continuum (e.g., selective self-memory, favorable interpretation of ambiguous feedback, self-serving bias; Hepper et al., 2010; Sedikides & Alicke, 2012, 2019) in a genetically sensitive design.
Besides self-enhancement per se, our findings inform the relation between self-enhancement manifestations and psychological well-being. Although a subset of the literature has reported a negative link between self-enhancement manifestations and well-being (e.g., Colvin & Griffo, 2008), the bulk of the literature (Alicke & Sedikides, 2009; Taylor & Brown, 1988), including a recent meta-analysis (Dufner et al., 2019), has established a positive link. Our research not only replicated the positive association between self-enhancement manifestations and well-being but also illustrated a positive genetic correlation between the latent traits corresponding to self-enhancement and well-being. In addition, our research demonstrated that the shared and nonshared environments shaping self-enhancement manifestations are positively related to those shaping well-being. To concretize these findings, genes and environments that contribute to self-positivity could also contribute to feeling satisfied with life and happy. Taken together, our findings bolster the positive relation between self-enhancement and well-being and further reveal a possible etiological mechanism for this relation.
Nonetheless, we cannot rule out the possibility of other influences at work. Besides common genes and environments, other factors may increase the resemblance between self-enhancement and well-being on phenotypic, genetic, or environmental level. For example, as six of our seven measures were self-report, it is possible that individuals’ response styles (e.g., acquiescence or extremity), which could implicate all the self-report measures, inflated the phenotypic correlation between self-enhancement and well-being. Because response styles are partly heritable (Melchers et al., 2018), the genetic factors underlying response styles may concurrently account for a portion of the individual differences in both self-enhancement and well-being measures, which could then increase the genetic similarity between self-enhancement and well-being. By the same logic, response styles, which are also influenced by nonshared environments (Melchers et al., 2018), may amplify the nonshared environmental correlation between self-enhancement and well-being. Future investigations would do well to control for response styles.
Limitations
We acknowledge limitations of our research and point to empirical directions. First, we sampled in a single culture. Follow-up studies would need to expand the scope and generalizability of the current findings by sampling in a variety of cultures, a very challenging goal to implement in genetics research due to forbidding costs. Second, we used the classic ACE models, which treated genetic and environmental effects as independent. This overlooks the possibility that genetic factors may correlate or interact with environmental factors in complex ways (Purcell, 2002). Such gene–environment correlations or interactions are critical for gaining nuanced knowledge about the origins of self-enhancement. Future investigations might include measures of environments (e.g., economic conditions; Bianchi, 2014) to test for potential interactions or correlations with genes. Third, our sample was moderate in size. This entailed drawbacks including wide CIs for model parameters and, more important, limited statistical power to detect small genetic or environmental effects, especially shared environmental effects (Verhulst, 2017). Indeed, the magnitude of several genetic and shared environmental effects, especially those originated from factors specific to a particular self-enhancement manifestation (Tables 4 and 5), was below the minimal effect size (.14) that we can detect with sufficient power. Hence, the estimates of these genetic and shared environmental effects should be treated with caution. Future studies would need to replicate our findings with larger samples.
Conclusion
We identified a core trait underlying various self-enhancement manifestations and demonstrated genetic and environmental composition of this latent core. In addition, we found that self-enhancement is associated with psychological well-being possibly due to similar genetic and environmental bases. The findings advance understanding of the origins and correlates of self-enhancement.
Supplemental Material
Supplemental Material, Heritability_of_Self-enhancement_R2_Supplement_submit - On the Etiology of Self-Enhancement and Its Association With Psychological Well-Being
Supplemental Material, Heritability_of_Self-enhancement_R2_Supplement_submit for On the Etiology of Self-Enhancement and Its Association With Psychological Well-Being by Yu L. L. Luo, Constantine Sedikides and Huajian Cai in Social Psychological and Personality Science
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Chinese national funding of social sciences (17ZDA324), CAS Key Laboratory of Behavioral Science, Institute of Psychology (Y5CX052003), and National Natural Science Foundation of China (31300871 and 31571148).
Supplemental Material
The supplemental material is available in the online version of the article.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.