
Abstract
While organoids hold immense promise as in vitro three-dimensional (3D) models, their translational utility is fundamentally constrained by passive diffusion limits (>800 μm), which inevitably trigger necrotic core formation and stochastic structural heterogeneity. This review elucidates how bioprinting shatters these physical bottlenecks by executing a paradigm shift toward spatiotemporal determinism. We systematically decode the mechanobiological evolution of bioinks—charting the transition from exogenous static matrices, which now function as temporal controllers via tunable stress relaxation to direct YAP/TAZ mechanotransduction, to the emerging paradigm of “engineerable living bioinks” driven by endogenous, cadherin-mediated fluid-to-solid jamming transitions. Furthermore, we critically evaluate frontier spatial strategies, highlighting how sacrificial networks and deterministic multi-material assembly establish active convective infrastructures and precise biophysical boundary conditions. By enforcing this rigorous baseline, these technologies definitively rectify pharmacokinetic/pharmacodynamic (PK/PD) distortions—eradicating false-positive noise in high-throughput screening and bridging the post-implantation mass transport vacuum—elevating organoids from stochastic clusters to highly predictive pathophysiological macro-models. Ultimately, we posit that transitioning from isolated morphological fabrication to resolving the inherent systemic metabolic paradoxes of multi-lineage integration is the absolute prerequisite for clinical translation.

Keywords
1. Introduction
Since the seminal 2008 demonstration by Yoshiki Sasai’s team that stem cells can spontaneously self-organize into complex neural structures in vitro, organoids have emerged as a cornerstone of biomedical research.1,2 From an engineering perspective, however, this biological “black-box” of random self-assembly is governed entirely by stochasticity, yielding fundamentally uncontrollable core physical parameters. For the ultimate translational mandate of organoids in high-throughput drug screening (HTS), this intrinsic stochasticity constitutes a formidable biophysical barrier. The growth of conventional avascular organoids is rigorously constrained by passive mass transport: computational and empirical models quantitatively confirm that exceeding a physical diffusion limit of approximately 800 μm inevitably triggers O2 starvation-induced central necrosis.3,4 Crucially, this passive diffusion-limited growth does not merely compromise cell viability—it generates severe artificial hypoxic gradients that aberrantly upregulate hypoxia-inducible factors (e.g., HIF-1α), driving artifactual chemoresistance and flooding screening readouts with false-positive noise. 5 This profound metabolic and geometric heterogeneity systematically erodes the standardized, high-signal-to-noise outputs essential for clinical translation. 6
Despite this promise, the translational utility of organoids is currently hindered by critical limitations driven by uncontrollable microenvironmental heterogeneities. These constraints include: (i) poor batch-to-batch reproducibility that triggers phenotypic drift and impedes standardization; (ii) limited nutrient and oxygen diffusion (mass transport limits), which critically distorts pharmacokinetic (PK) drug penetration profiles 7 ; and (iii) the lack of functional vascularization and complex microenvironmental components (e.g., immune cells, neurons), which compromises the fidelity of disease modeling alongside the profound absence of spatiotemporally tunable matrix biomechanics and dynamic mechanical feedback (e.g., fluid shear stress, matrix stress relaxation).8–10
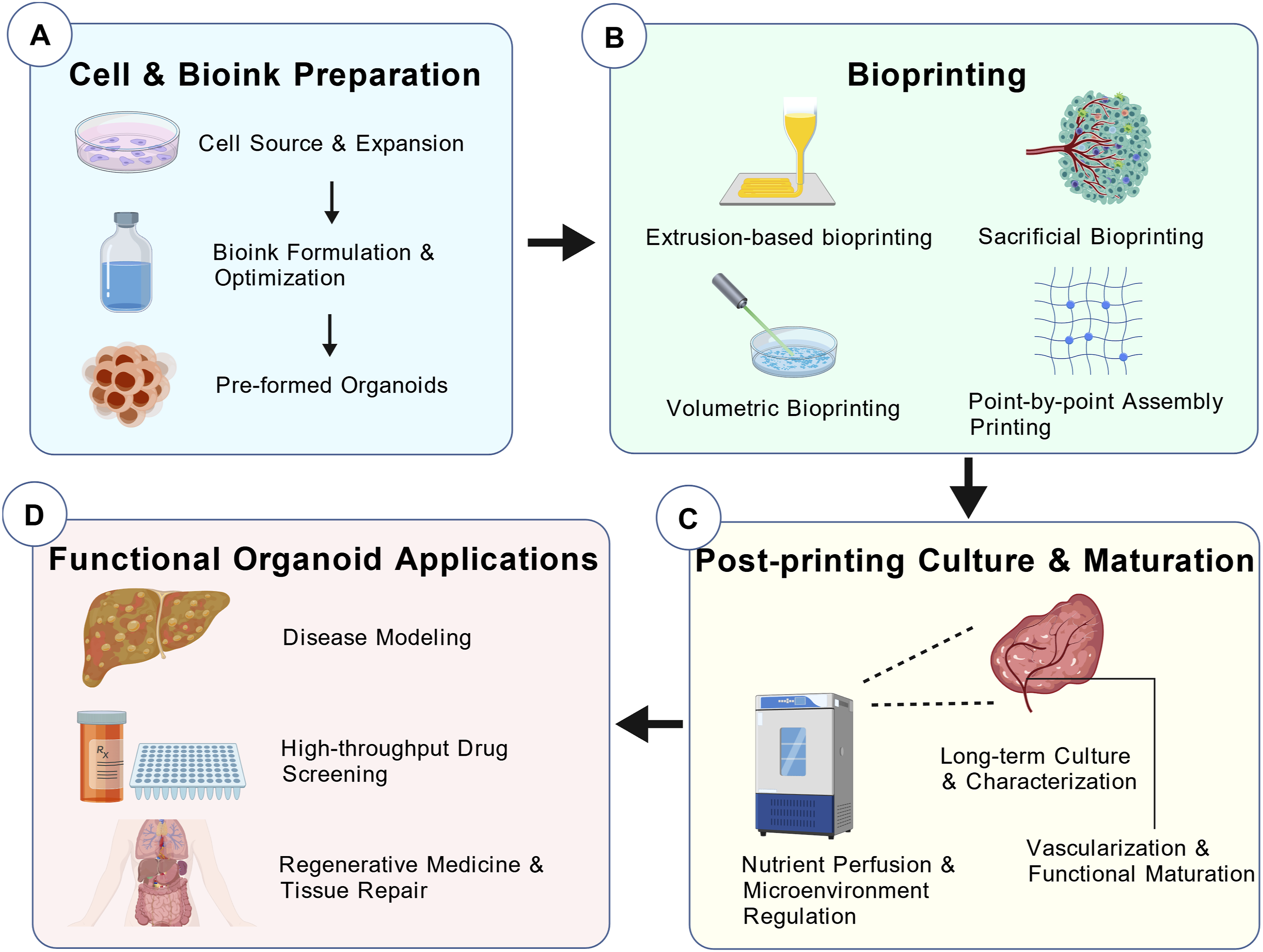
To address these bottlenecks, bioprinting has emerged as a pivotal strategy for engineering “next-generation organoids.” Leveraging high programmability and structural control, bioprinting enables the fabrication of systems with enhanced complexity and functionality (Figure 1 for the bioprinting workflow).11–13 Moving far beyond mere morphological biomimicry, bioprinting functions as an active biophysical intervener—executing spatiotemporally deterministic reconstructions of both the mass transport infrastructure and the biomechanical niche. This review synthesizes innovative advancements in bioprinted organoids over the past five years. Specifically, Section 2 scrutinizes how bioink crosslinking chemistry serves as a “temporal controller,” dictating matrix stress relaxation and mechanotransduction to guide cell fate; Section 3 evaluates how frontier printing modalities shatter spatial mass transport limits through the programmable integration of perfusable vascular architectures; and Section 4 demonstrates how achieving this spatiotemporal determinism definitively rectifies PK/PD distortions, translating into highly predictive precision disease models. Ultimately, we elucidate how these technologies elevate organoids from stochastically self-assembled cellular aggregates to deterministic, highly predictive in vitro digital twins, bridging the systemic gap toward robust clinical application. Spatiotemporally deterministic workflow of bioprinting next-generation organoids (a): Temporal mechanocontrol via bioink preparation; (b): Spatial mass transport infrastructure via bioprinting strategies; (c): Microenvironmental stabilization via post-printing maturation; (d): High-fidelity clinical translation as predictive pathophysiological macro-models)
14
.
2. Bioink innovation: Biochemical and biomechanical controllers in the temporal dimension
Overview of bioink types, core properties and applications.
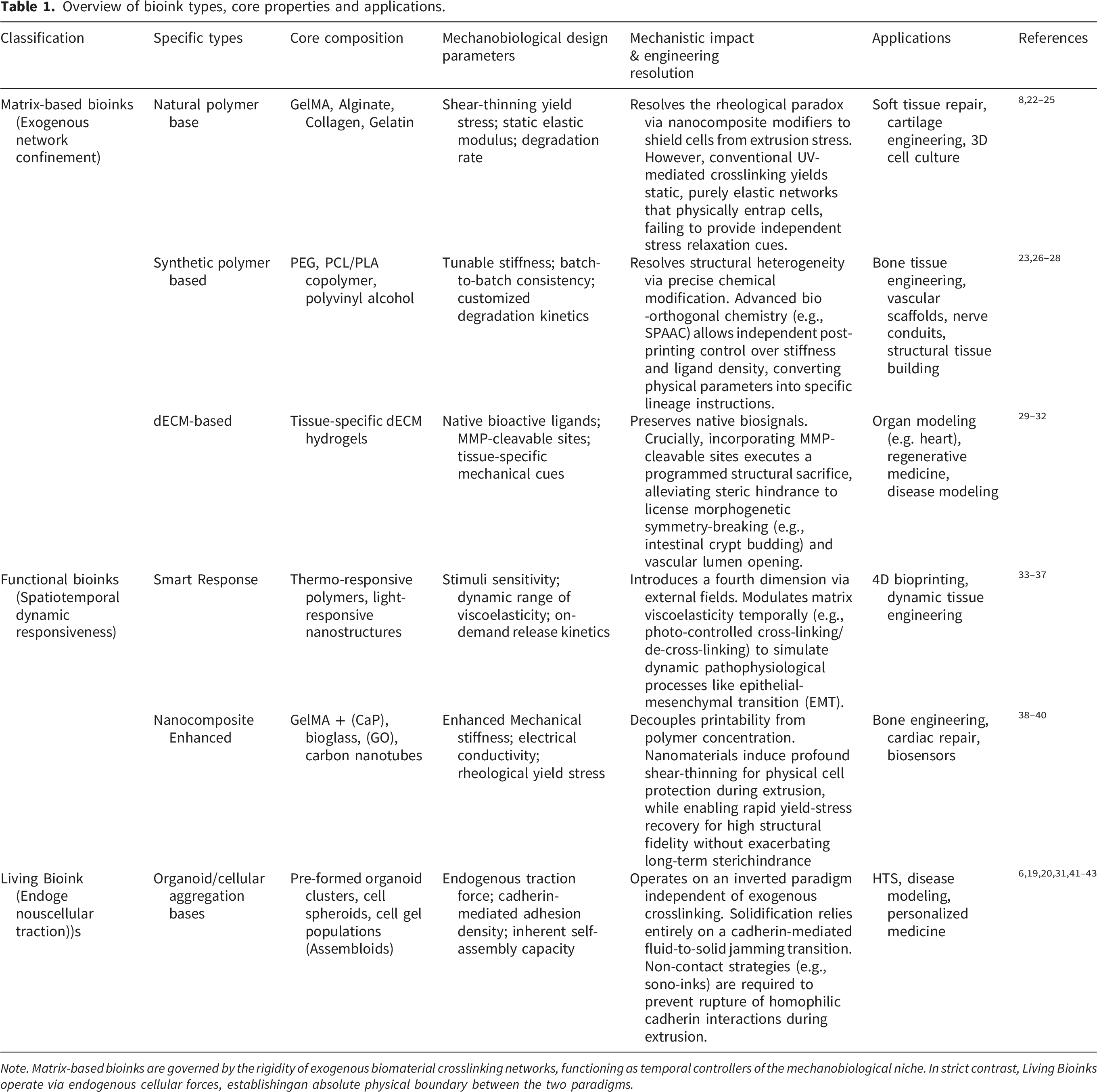
Note. Matrix-based bioinks are governed by the rigidity of exogenous biomaterial crosslinking networks, functioning as temporal controllers of the mechanobiological niche. In strict contrast, Living Bioinks operate via endogenous cellular forces, establishingan absolute physical boundary between the two paradigms.
2.1. Advances in traditional bioink strategies
Conventional bioinks — serve as physical scaffolds replete with bioactive signals. 15 Recent research has shifted from basic material blending to the development of multi-functional, intelligent, and programmable systems that more accurately simulate the dynamic in vivo microenvironment. From an engineering perspective, formulating traditional matrix bioinks necessitates a rigorous rheological compromise. The polymer network must possess specific shear-thinning and yield-stress properties to enable precise spatial extrusion; yet, increasing polymer concentration to enhance structural fidelity invariably elevates nozzle shear stress, inflicting severe mechanically induced cellular damage during printing. To resolve this conflict, recent efforts have focused on modulating polymer molecular weight or incorporating rheological modifiers such as nanoclays.22,23,38 These nanocomposite strategies fundamentally decouple printability from polymer concentration: they induce profound shear-thinning behavior that physically shields cells from extrusion stress, while enabling rapid yield-stress recovery post-printing to guarantee high structural fidelity.8,24 Nevertheless, this approach introduces a significant rheological–biological trade-off—the accumulation of non-degradable inorganic modifiers can inadvertently exacerbate long-term steric hindrance, potentially disrupting endogenous extracellular matrix (ECM) deposition and interfering with organoid expansion.
While rheological properties govern the feasibility of initial spatial deposition, the subsequent crosslinking chemistry rigidly dictates the long-term temporal evolution of the mechanobiological niche. Specifically, the cellular responsiveness of these bioinks is determined by how their crosslinking mechanisms define the matrix’s initial stiffness versus its stress relaxation profiles (a conceptual flowchart delineating the systemic information flow from macro-scale printing parameters to micro-scale mechanotransduction is provided in Figure 2). Crucially, conventional UV-mediated free-radical polymerization (e.g., highly substituted GelMA) predominantly yields static, purely elastic networks that physically entrap encapsulated cells. In contrast, the integration of dynamic covalent bonding—exemplified by reversible imine bonds and host–guest interactions—endows the network with tunable viscoelasticity.33,34,44 These distinct biophysical signals are rapidly transduced via integrin-mediated focal adhesions, triggering cytoskeletal rearrangement of the actomyosin network. This mechanotransduction cascade subsequently activates mechanosensitive nuclear transcription factors, most notably YAP/TAZ, which spatiotemporally direct stem cell lineage specification. Recent paradigm-shifting models necessitate the explicit decoupling of these two physical variables, quantitatively demonstrating that rapid stress relaxation—driven by dynamic bond exchange and wholly independent of absolute matrix stiffness—acts as the primary biophysical license for robust YAP/TAZ nuclear translocation and cellular proliferation.8,45 Systemic mechanobiological information flow from macro-scale bioprinting parameters to micro-scale cell fate. (a) Macro-Scale Inputs: Bioink chemistry fundamentally dictates initial physical boundaries, contrasting static covalent networks (gray) with dynamic, reversibly crosslinked networks (blue). (b) Meso-Scale Niche: These crosslinking chemistries deterministically dictate matrix stress relaxation profiles (representative curves modeled after Chaudhuri et al., 2020
8
). Dynamic networks enable fast stress relaxation, acting as a crucial physical license. (c) Micro-Scale Transduction: Fast stress relaxation permits the physical clustering of integrins and subsequent formation of focal adhesions (FA). This mechanical tension is transduced via the actomyosin cytoskeleton, effectively driving the nuclear translocation of mechanosensitive YAP/TAZ transcription factors to activate target genes and dictate morphogenetic cell fates.
Beyond mechanotransduction via network elasticity, bioink design must be spatiotemporally matched to the intrinsic degradation kinetics of developing organoids. Furthermore, dECM-based bioinks retain native ECM architecture and bioactive ligands, providing a highly biomimetic microenvironment. Frontier synthetic strategies focus on precision functionalization through specific ligand presentation (e.g., RGD cell-adhesion peptides) and the incorporation of matrix metalloproteinase (MMP)-cleavable sites. This biochemical programmability is absolutely imperative: a successful bioink must not function as a permanent physical prison. Instead, it must execute a programmed structural sacrifice, permitting organoids to enzymatically degrade, remodel, and ultimately replace the exogenous scaffold with their own secreted endogenous matrix. Quantitative in vitro models have unequivocally established this causal relationship, demonstrating that precisely tuned MMP-mediated degradability constitutes an absolute physical prerequisite for alleviating steric hindrance, thereby licensing morphogenetic symmetry-breaking events—such as intestinal crypt budding—and enabling the collective cellular invasion required for the physical opening of perfusable vascular lumens.29,30 Collectively, the field is advancing toward bioink formulations that are increasingly programmable, responsive, and physiologically relevant.10,35
2.2. Organoids: As engineerable living bioinks
Conceptualizing organoids—whether as single-cell suspensions, pre-formed spheroids, or macro-scale fused “assembloids”—as engineerable “living bioinks” represents a novel paradigm in bioprinting. 20 To eliminate conceptual ambiguity, it is imperative to enforce the absolute physical boundary established in Section 2.1: whereas traditional strategies rely on exogenous polymers for structural confinement, living bioinks operate on an inverted paradigm of cells engulfing materials. This approach leverages the engineerable attributes of living components—specifically their rheological, biochemical, and self-organizing properties—to drive an active liquid-to-solid phase transition, paving the way for complex tissue fabrication.12,31,46 Unlike chemical crosslinking, this transition depends entirely on robust cadherin-mediated cell–cell adhesions and endogenous ECM secretion; any trace exogenous biomaterials present serve strictly as transient sacrificial binders or yield-stress support baths. In this paradigm, high-density cellular constructs are extruded into a viscoplastic sacrificial matrix, after which a cadherin-mediated fluid-to-solid jamming transition rapidly dictates physical solidification—requiring no permanent exogenous networks whatsoever.21,41
When deploying organoids as bioinks, the synergy between physicochemical properties and biological viability is paramount. 20 Managing the severe fluidic–biological trade-off during extrusion becomes critical. Physically, densely packed cellular aggregates lack the stress-shielding effects of a robust exogenous matrix, rendering them exquisitely sensitive to flow-induced trauma. The high wall shear stresses generated during extrusion do not merely reduce bulk viability—they critically threaten to rupture the homophilic cadherin interactions (e.g., E/N-cadherin) essential for the subsequent phase transition. Preserving this inherent self-assembling capacity therefore demands that bioinks must exhibit profound shear-thinning behavior and appropriate yield stress to ensure extrudability at high cell densities while mitigating mechanical damage.21,42 Recent studies integrating bioprinting with interferometry have optimized the rheology of organoid-laden slurries, achieving high-fidelity deposition while maintaining >90% cell viability. 47 Biologically, stressors such as shear force, phototoxicity, and thermal fluctuations can compromise organoid function. To circumvent these flow-induced mechanical bottlenecks altogether, novel strategies such as viscoelastic “sono-inks” have emerged. By utilizing ultrasound for deep-tissue penetration and assembly, these systems avoid high shear forces and phototoxicity, thereby preserving intact intercellular communication networks required for immediate solidification and organoid functionality. 43
Furthermore, the temporal dimension of living bioinks culminates in spontaneous, biologically driven post-printing architectural evolution—a process fundamentally distinct from the synthetic 4D hydrogel remodeling discussed previously.48,49 Here, the fourth dimension is governed intrinsically by biological morphogenesis rather than externally imposed material stimuli. When constructing complex, multi-regional assembloids—cortico-striatal organoids being a canonical example—ultimate functional integration relies entirely on endogenous mechanobiology.36,50 This integration proceeds via a dual morphogenetic mechanism: localized morphogen gradients at the printed tissue boundary actively break spatial symmetry, 51 while subsequent cadherin-mediated collective cellular migration physically zips the discrete macro-interfaces together.52–54 This autonomous interfacial fusion successfully establishes mature systemic connectivity without disrupting region-specific identities, highlighting the unparalleled capacity of living bioinks to recapitulate dynamic physiological processes such as self-organized tissue polarity and multi-organ crosstalk.26,46,55
3. Bioprinting strategies: Driving structural and functional breakthroughs in organoids
If bioinks function as the temporal controllers governing mechanotransduction (Section 2), bioprinting strategies constitute the indispensable spatial infrastructure for mass transport. Over the past five years, bioprinting has advanced substantially, overcoming the profound physical bottlenecks inherent to conventional organoids by executing a fundamental fluidic shift: from passive, diffusion-limited nutrient gradients to active, convective mass transport systems. This deterministic spatial patterning not only shatters the viability threshold historically imposed by avascular cores, but also precisely re-establishes the macro- and micro-scale heterogeneity essential for physiological fidelity.
13
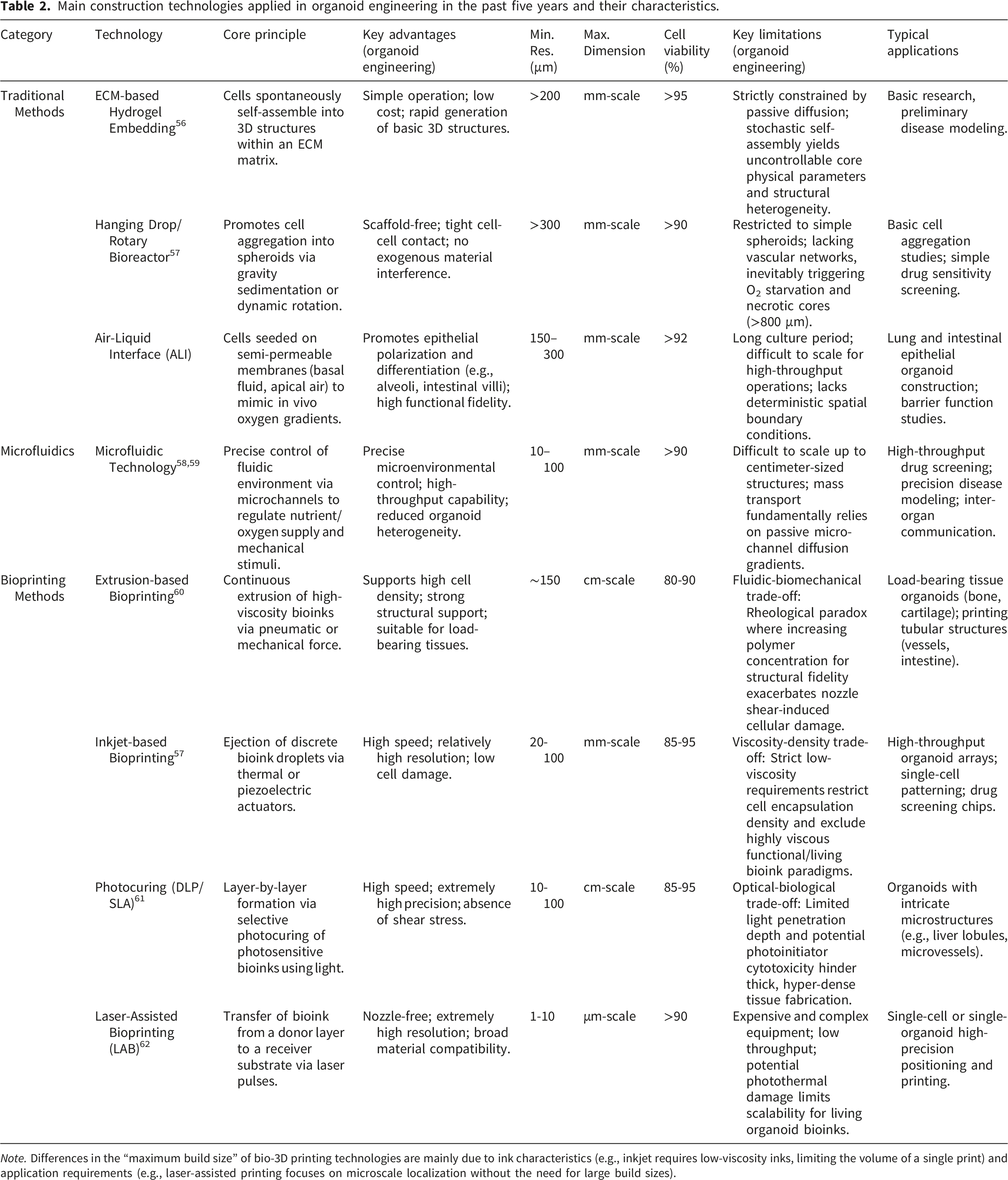
This section highlights the most transformative frontier technologies (Figure 3), while conventional methods are detailed in Table 2. Crucially, moving decisively beyond mere advocacy of technological novelty, we introduce a rigorous evaluation of the fundamental engineering trade-offs inherent to each strategy. Every leap in fabrication speed or spatial resolution inevitably exacts a physiological or biomechanical toll; dissecting these compromises constitutes the absolute prerequisite for evaluating their true translational potential. Frontier spatial bioprinting strategies shattering mass transport limits (a): Active convective infrastructure via sacrificial bioprinting; (b): Deterministic macro-spatial communication via bioprinted organ-on-a-chip; (c): Precise biophysical boundary conditions via multi-material bioprinting. Main construction technologies applied in organoid engineering in the past five years and their characteristics. Note. Differences in the “maximum build size” of bio-3D printing technologies are mainly due to ink characteristics (e.g., inkjet requires low-viscosity inks, limiting the volume of a single print) and application requirements (e.g., laser-assisted printing focuses on microscale localization without the need for large build sizes).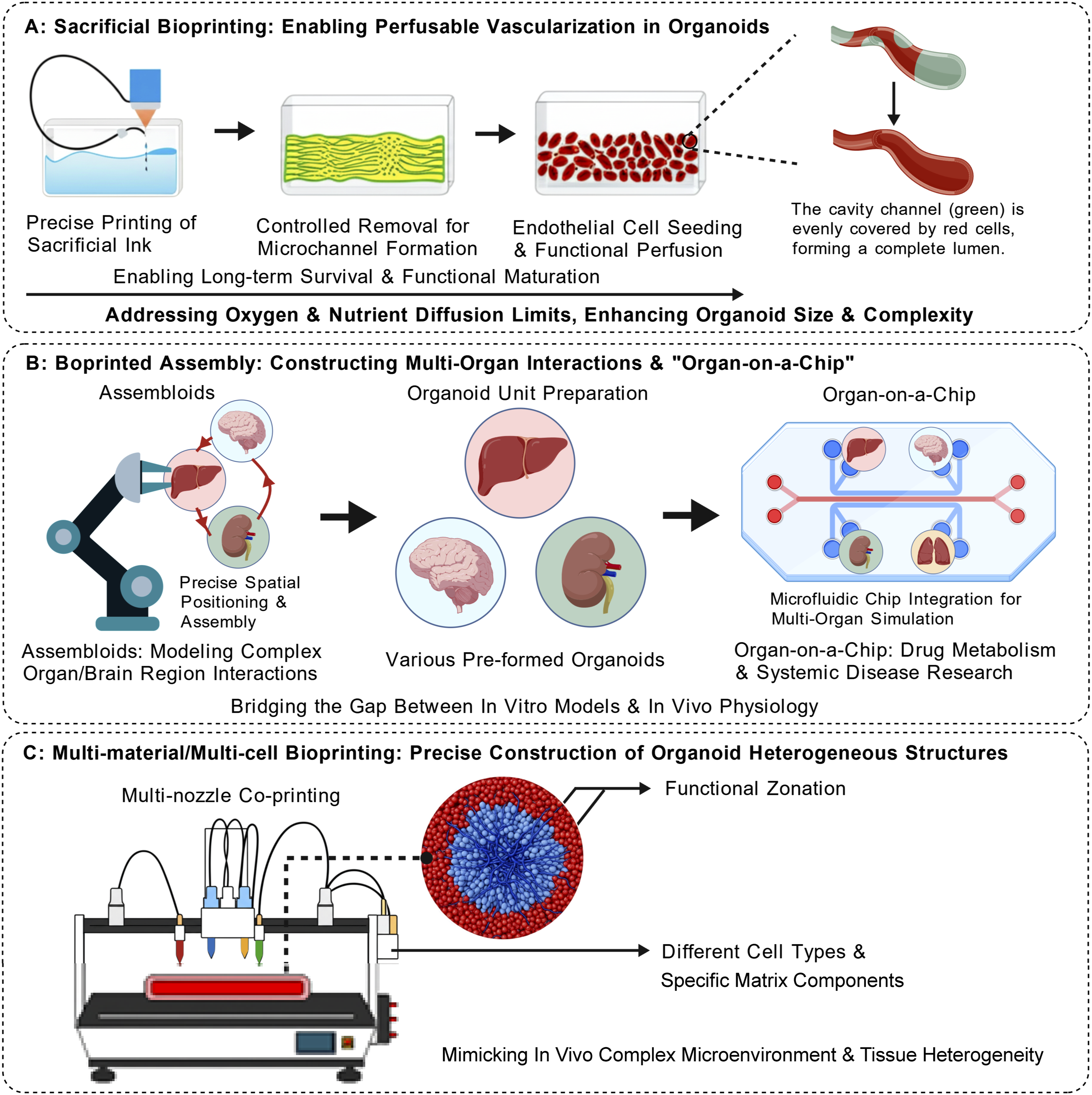
3.1. Volumetric Bioprinting for precise construction of macroscopic complex structures
Volumetric Bioprinting (VBP) represents a paradigm shift in biofabrication. Unlike conventional layer-by-layer additive manufacturing, VBP utilizes tomographic light projections or multi-beam interference to simultaneously fabricate complete, centimeter-scale 3D architectures within seconds.20,63,64 The defining mechanobiological advantage of VBP is its absolute zero-shear environment.
Mechanistically, this eliminates the nozzle-induced mechanical trauma that fundamentally plagues extrusion-based methods, positioning VBP as the ultimate fabrication vessel for the highly fragile, densely packed living bioinks discussed in Section 2.2—meticulously preserving their critical cell–cell communication networks. 64 For instance, Bernal et al. leveraged VBP to construct centimeter-scale bioreactors containing functional liver organoids in under 20 seconds. These constructs exhibited enhanced metabolic functions, including significant ammonia detoxification, a urea synthesis rate of 0.4 μg day-1 mm-3, and a twofold increase in CYP450 activity compared to static 2D controls.63,65 Despite these metabolic triumphs, deploying VBP for organoid engineering necessitates navigating a severe optical–biological trade-off. While VBP excels in stress-free fabrication, its underlying optical physics renders it profoundly susceptible to light scattering when handling the physiologically relevant, hyper-dense organoid bioinks essential for clinical translation. This Mie scattering—driven by the severe refractive index mismatch between lipid-rich cell membranes and aqueous bioinks—exponentially attenuates photon penetration, inevitably producing fatal curing failures at the core of thick tissues. Quantitative analyses confirm that this physical phenomenon critically disrupts the precise tomographic light dose at concentrations exceeding 106–107 cells mL-1. Recent engineering interventions have demonstrated, however, that incorporating precise optical tuning agents to actively match these refractive indices effectively neutralizes the scattering effect, unlocking high-fidelity volumetric fabrication even at hyper-dense concentrations exceeding 108 cells mL-1.65,66 To fully transition VBP from a structural novelty to a routine clinical tool, future research must rigorously focus on optimizing deep-tissue optical penetration through advanced materials engineering and multi-physics field coupling.
3.2. Sacrificial bioprinting and the construction of perfusable vascular networks
The profound absence of functional vascular networks represents a critical translational barrier to organoid scalability and long-term viability — one that does not merely limit viability, but fundamentally distorts pharmacokinetic (PK) penetration profiles, generating artificial diffusion gradients and necrotic cores that produce massive false-positive noise in drug screening. Sacrificial bioprinting decisively addresses this by pre-patterning vascular channels within the host matrix using fugitive inks (e.g., gelatin, Pluronic F127) that are subsequently removed.13,67 This paradigm fundamentally shifts mass transport from limited passive diffusion to active convection, ensuring immediate and uniform solute delivery across centimeter-scale constructs. Notable implementations include the liquid metal-based “ESCAPE” method by Sundaram et al., which exploits the low melting point of gallium (29.8°C) for non-destructive channel formation. 68 Similarly, the “PRINCESS” strategy—combining femtosecond laser holographic processing with DNA-based biolubricants—enables the printing of biomimetic capillary scaffolds (6–9 μm) and overcomes limits in directly printing living cells within 70 μm endothelialized microvessels.69,70 A recent breakthrough involves temporally controlled vascularization, where programmable crosslinking regulates the dissolution rate of sacrificial inks. By dynamically optimizing oxygen gradients in cardiac organoids, this approach yielded a 200% increase in contractility and an 80% reduction in hypoxic regions compared to non-vascularized controls.21,71
Nevertheless, translating these structural networks into truly functional endothelium exposes a severe fluidic–biological trade-off. Extracting conventional high-yield-stress fugitive inks requires aggressive mechanical flushing; this process generates extreme wall shear stresses that frequently rupture the fragile, newly formed tight junctions of the endothelial lining, critically compromising microvascular barrier integrity. To circumvent this flow-induced trauma entirely, frontier biofabrication strategies now employ programmable delayed-dissolution bioinks. By precisely tuning the chemical crosslinking network, these sacrificial templates are engineered to undergo endogenous self-degradation over predefined time windows, completely eliminating the need for destructive physical flushing.18,71,72 Ultimately, this synergistic integration of programmed material degradation and active convection restores physiological mass transport and PK fidelity, serving as the indispensable bridge for advancing bioprinted organ organoids toward clinical translation. 7
3.3. Point-by-point bioprinting: Reconstructing macro-spatial communication
To simulate the intricate systemic interactions between distinct organs or regional brain domains—tumor–immune or cortico-striatal crosstalk being canonical examples—establishing deterministic spatial boundary conditions is an absolute prerequisite. “Assembloid” models have been developed to simulate the intricate interactions between distinct organs or brain regions. Point-by-point bioprinting techniques—such as Aspiration-Assisted Bioprinting (AAB) and Spatial Patterning of Organoids Transfer (SPOT)—offer unprecedented precision in this domain. These methods enable the placement of individual, pre-formed organoid spheroids at predetermined coordinates with exceptional spatial precision. For instance, SPOT technology facilitates non-contact manipulation via magnetic nanoparticle surface modification and programmable magnetic fields, achieving a positioning accuracy of ±18.7 μm with >95% cell viability. 53 Similarly, 3D bio-dot printing utilizing in situ induced microwell spheroid formation (positioning error ≤5 μm) has significantly profoundly amplified directional paracrine communication efficiency. 73 Additionally, a scaffold-free “pure cell” assembly technique has further demonstrated the capacity to construct hyper-dense neural organoids (>108 cells/cm3), where endogenous ECM formation accelerated neuronal migration by 80%.21,74,75
These technologies provide high-precision platforms for investigating heterogeneous tissue interactions, evidenced by their success in quantifying invasive variances in glioma patients (up to 2.8-fold difference) and mimicking spinal cord circuit functions (120% increase in calcium signaling frequency). 53 Translating this spatial precision into scalable functionality, however, introduces a severe spatiotemporal and biomechanical trade-off. While yielding extreme spatial fidelity, the piece-by-piece manipulation inherently bottlenecks manufacturing throughput. More critically, the ultimate functional integration of these discrete modules relies entirely on spontaneous fusion across the printed interfaces—a process that remains fundamentally unpredictable. This reliance creates a profound biomechanical paradox: locking spheroids into macroscopic spatial coordinates requires high-yield-stress bioinks, yet this static mechanical confinement imposes severe microscale steric hindrance that physically arrests the filopodial extension and homophilic cadherin engagement requisite for interfacial tissue fusion.20,74 To decouple this spatial-fusion paradox, frontier biofabrication strategies now leverage parallelized, physics-driven platforms—such as geometrical confinement within micro-molds and field-driven acoustic radiation forces. These systems actively apply mechanical vectors to enforce standardized, high-throughput organoid coalescence, safely overcoming the thermodynamic energy barriers to spontaneous tissue fusion without inducing shear trauma or sacrificing macro-spatial precision.6,10,76
3.4. Multi-material/multi-cellular bioprinting: Simulating complex microenvironments and multi-tissue interactions
Multi-material/multi-cellular bioprinting leverages multi-nozzle or microfluidic switching systems to precisely co-deposit diverse bioinks and cell types in a single run. By establishing highly specific biophysical boundary conditions, this modality faithfully replicates micro-anatomical and mechanobiological heterogeneity—including the programming of precise physical gradients such as the metabolic zonation of hepatic lobules or the defined necrotic-invasive margins of solid tumors. 13 Utilizing core-shell microfluidic printing with coaxial nozzles, Formica et al. synchronously extruded core and shell bioinks to engineer kidney organoids containing a tri-layered structure (glomerulus-proximal tubule-collecting duct) within 14 days thereby establishing critical epithelial polarization. 77 In a hybrid approach, Kiefer et al. integrated a multi-photon lithography module (200 nm resolution) within a multi-material extrusion framework, establishing a technical foundation for constructing capillary-level networks and authentic basement membrane topologies. 78 Furthermore, the Zhejiang University team developed a multi-material projection printing system capable of the simultaneous, high-precision deposition of six distinct materials, significantly advancing biofabrication toward precise biomimicry. 79 Despite these architectural triumphs, co-printing diverse, heterogeneous lineages—tumor cells, stromal fibroblasts, and immune effectors—directly exposes a critical, often-overlooked biological–metabolic paradox. The fundamental absence of a universal culture medium capable of simultaneously satisfying the distinct metabolic demands and optimal differentiation niches of all co-cultured lineages frequently induces severe phenotypic drift, immune exhaustion, or competitive overgrowth, exemplified by the competitive spatial exclusion driven by highly proliferative fibroblasts. To definitively reconcile this inherent metabolic conflict, frontier biofabrication strategies now exploit structural microfluidic compartmentalization. By engineering selectively permeable endothelial or hydrogel barriers, this approach successfully decouples metabolic supply from signaling crosstalk: each distinct tissue niche can be rigorously maintained in its own optimized medium to preserve phenotypic stability, while still permitting the critical trans-barrier diffusion of paracrine morphogens and metabolites necessary for systemic integration. 7
4. Frontier applications of bioprinted organoids: Clinical translation driven by spatiotemporal reconstruction
Biophysical and Pharmacological Fidelity: Conventional Self-Assembled Organoids vs. Bioprinted Organoids.
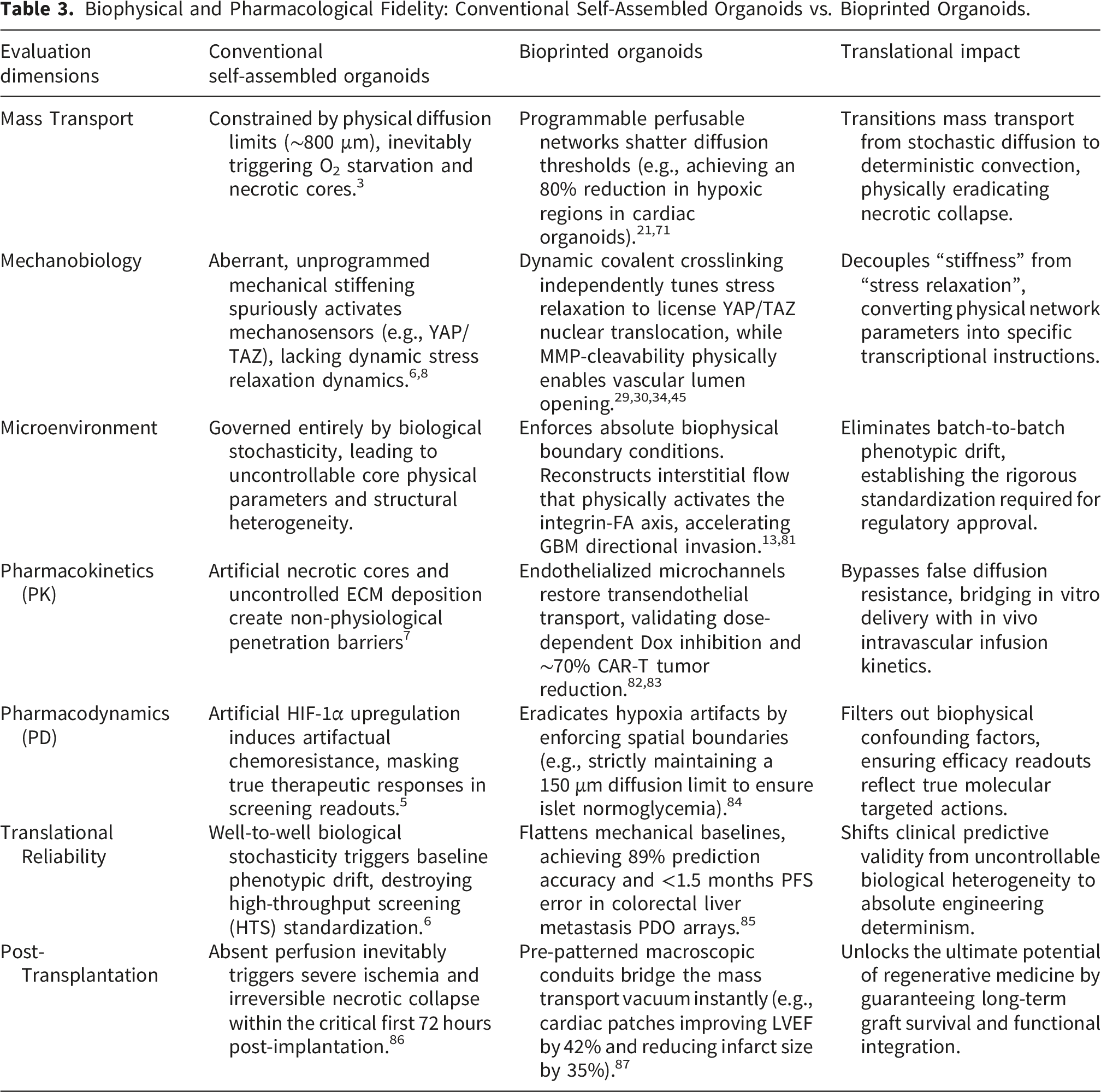
4.1. Precision disease modeling: From morphological mimicry to physical and biomechanical replication
In disease modeling, bioprinted organoids are transitioning from static morphological mimics to dynamic systems capable of recapitulating the core mechanisms of pathogenesis. The fundamental failure of conventional avascular tumor organoids originates in their strict reliance on passive diffusion. This physical limitation inevitably generates artificial necrotic cores and aberrant hypoxia-inducible factor 1-alpha (HIF-1α) upregulation, locking the in vitro model into a state of artifactual chemoresistance. A 2022 study utilizing a 3D bioprinted vascularized breast cancer model demonstrated that enforcing active convection through pre-patterned vascular networks definitively rectifies this pharmacokinetic (PK) distortion: doxorubicin perfusion inhibited tumor growth in a dose-dependent manner (significant volume reduction after 72h at 10/100 μM). Furthermore, perfusion with anti-HER2 CAR-T cells yielded a ∼70% reduction in tumor volume. Because this active mass transport restores physiological immune trafficking and definitively eliminates the biophysical noise of false hypoxia, the resulting chemo-sensitivity and pharmacodynamic (PD) profiles possess a level of clinical predictive validity that static models fundamentally cannot attain.82,83 Beyond restoring mass transport, bioprinting decisively recreates the dynamic biomechanical niche. For neurological disorders, researchers are leveraging microfluidic integration to mimic the cerebrospinal fluid (CSF) circulatory microenvironment. Crucially, these models confirmed that elevated interstitial flow (IF) imposes precise fluid shear stress that physically activates mechanotransduction pathways at the cell–matrix interface—notably, the integrin–focal adhesion axis. This continuous mechanical stimulation drastically accelerates the migration and directional motility of glioblastoma cells, and matrix invasion; the strictly biophysical induction of this malignant invasive phenotype constitutes a spatially driven, dynamic process to which conventional static Petri dish cultures remain entirely blind. 81 Despite these advances, challenges remain: a critical engineering and systemic bottleneck persists. Current bioprinted tumor models predominantly feature static, pre-patterned endothelial channels. While sufficient for initial PK delivery, these rigid geometries fundamentally fail to capture the dynamic, tumor-driven angiogenesis and complex transendothelial immune cell migration required for evaluating next-generation immunotherapies and anti-angiogenic agents.29,88
4.2. High-throughput drug screening: Eliminating the noise of phenotypic drift
The inherent automation and standardization of bioprinting address critical bottlenecks in traditional organoid culture, such as prolonged timelines and batch heterogeneity. Mechanobiologically, conventional manual dispensing is an inherently stochastic process that generates profound well-to-well variations in matrix stiffness and initial cell density. These unprogrammed physical fluctuations arbitrarily activate cellular mechanosensors such as YAP/TAZ, artificially inflating or suppressing baseline survival pathways before therapeutics are even introduced. This physical inconsistency inevitably triggers severe phenotypic drift, flooding screening readouts with massive false-positive noise. 6 By functioning as a systemic biophysical equalizer—flattening the biomechanical baseline across hundreds of microwells through uniform bioink crosslinking (temporal control) and rigorously consistent volumetric deposition (spatial control) — this accelerates high-throughput drug screening (HTS) and personalized medicine.47,89
By constructing patient-derived organoid (PDO) chips on a massive scale, rapid multi-parametric drug evaluation becomes feasible. For instance, a PDO model of colorectal cancer liver metastasis developed by the Peking Union Medical College Hospital team achieved an 89% prediction accuracy (20/23 cases) for chemosensitivity. Notably, the prediction error for progression-free survival (PFS) in oxaliplatin-resistant patients was <1.5 months, offering a vital tool for precise clinical regimen formulation. 85 Crucially, this remarkable accuracy is not merely an intrinsic biological attribute of the PDOs, but a direct consequence of the engineered bioprinting infrastructure. By guaranteeing that chemotherapeutics such as oxaliplatin encounter absolutely identical physical penetration gradients and matrix barriers in every test well, the system extracts pharmacodynamic (PD) data with an exceptionally high signal-to-noise ratio.
Nevertheless, while bioprinting substantially enhances structural standardization, a critical physical bottleneck persists in the readout phase: the dense, crosslinked 3D matrices that confer mechanical fidelity simultaneously impose severe optical scattering and reagent-diffusion barriers. This fundamental optical–mass transport conflict profoundly limits real-time, non-destructive viability assays and continuous metabolic monitoring across high-throughput 3D arrays—an engineering hurdle that must be overcome to fully realize automated HTS.90–92
4.3. Regenerative medicine: Bridging the vascularization valley of death post-transplantation
In regenerative medicine, the ultimate goal is to engineer transplantable tissues that achieve immediate and sustained functional integration at the host–graft interface. Conventional centimeter-scale organoids, however, inevitably encounter a fatal vascularization valley of death: within the critical first 72 hours post-implantation, the inability of host vasculature to rapidly infiltrate the massive cellular bulk leads inexorably to severe ischemia and irreversible necrotic collapse at the core. Bioprinting decisively breaks this impasse by pre-patterning macroscopic perfusion networks that enable immediate convective flow upon surgical anastomosis. 86 Chen et al., for instance, utilized coaxial 3D bioprinting to fabricate cardiac patches containing microvascular networks (50–100 μm diameter). Mechanistically, these engineered conduits function as active physical channels for immediate hemodynamic perfusion, completely bridging the post-implantation mass transport vacuum. 93 These grafts achieved >90% anastomosis with host vessels upon transplantation into a rat myocardial infarction model. Functionally, the patches improved left ventricular ejection fraction (LVEF) by 42% and reduced infarct size by 35% compared to controls, validating the successful functional integration of the vascularized grafts. 87
Beyond macroscopic vessels, bioprinting exerts absolute physical control over microscale diffusion limits. Additionally, the FluidForm Bio team employed Freeform Reversible Embedding of Suspended Hydrogels (FRESH) bioprinting to differentiate gene-corrected iPSCs into islet organoids. Critically, their deterministic spatial patterning guarantees that every individual β-cell resides strictly within the absolute thermodynamic diffusion–reaction limit of 150 μm. By structurally immunizing cells against hypoxic stress, this precise biophysical boundary condition directly translates into long-term functional survival. Implantation into type 1 diabetic mice restored glucose homeostasis (fasting blood glucose <100 mg/dL) with sustained insulin secretion for over 12 weeks, highlighting a novel avenue for cell replacement therapy.84,94
Despite these mass transport victories, severe biomechanical and systemic engineering bottlenecks persist. Current bioprinted hydrogels frequently suffer from profound mechanical impedance mismatch with host tissues, leading to shear-induced graft delamination when implanted onto dynamic, mechanically active organs such as the beating heart. Furthermore, while the field has heavily prioritized arteriovenous perfusion, the complete absence of engineered lymphatic drainage networks in these constructs inevitably triggers interstitial fluid accumulation and graft edema—representing the next critical biophysical frontier in whole-organ biofabrication.95,96
5. Challenges and future perspectives: Towards “living engineered organs”
While bioprinted organoids have achieved substantial milestones, scaling these discrete structural triumphs into robust clinical predictors exposes a critical systemic vulnerability. Realizing the ultimate goal of “living engineered organs” necessitates overcoming multifaceted systemic challenges. 20 The ultimate translational bottleneck is no longer a localized physical limitation, but rather the emergence of coupling disasters in multi-dimensional biological systems. Moving from localized structural fabrication toward systemic clinical translation, the forced spatial integration of conflicting metabolic requirements, asynchronous cellular maturation timelines, and disparate mechanobiological needs—most notably, the highly divergent matrix stiffness and stress-relaxation preferences across co-cultured lineages—inevitably generates compounding biophysical and biological noise.8 This systemic desynchronization critically dismantles the very pharmacokinetic/pharmacodynamic (PK/PD) fidelity that these engineered architectures were designed to establish. Consequently, the future of the field depends not on isolated morphological or technological novelties, but on fundamentally resolving these multi-dimensional systemic paradoxes to achieve genuine physiological integration. 97
5.1. Key challenges: From technical bottlenecks to systemic complexities
Current impediments have evolved from isolated technical bottlenecks into interconnected, systemic complexities. Moving beyond the conventional discourse on structural integration, we must confront the two most consequential blind spots that currently generate massive artifactual data in biofabrication. The first is the metabolic phenotypic drift paradox inherent to multi-cellular co-printing. While advanced bioprinting can physically assemble diverse, heterogeneous lineages—tumor cells, immune components, and stromal fibroblasts—within a single spatial construct, 79 this structural achievement masks a fundamental biological conflict. The field currently lacks a universal culture medium capable of simultaneously satisfying the distinct metabolic pathways and differentiation niches of all co-cultured cells. Consequently, prolonged co-culture inevitably drives severe phenotypic distortion, manifesting as rapid immune cell exhaustion or competitive spatial overgrowth by fibroblasts. 84 Crucially, this metabolic incompatibility does not operate in isolation; it triggers a catastrophic mechanobiological cascade. Overgrowing fibroblasts actively contract and remodel the printed extracellular matrix, imposing aberrant, unprogrammed mechanical stiffening that spuriously upregulates mechanosensors such as YAP/TAZ in neighboring cells. This coupled metabolic–mechanical drift currently represents the single greatest source of false-positive data in advanced disease modeling, severely eroding the clinical predictive validity of multi-tissue constructs. Compounding this challenge, disparate cell lineages exhibit asynchronous maturation timelines, generating desynchronized physiological rhythms that further destabilize systemic functionality.
Realizing these complexities imposes stringent demands on the development of novel bioinks, which must combine superior biocompatibility, tunable mechanical properties, and programmable biological instruction capabilities.79,98 However, this exposes the second critical blind spot residing in the profound data silos surrounding bioink rheology and mechanobiology. The rapid evolution of bioprinting has produced an immense volume of process parameters; yet, owing to highly divergent testing standards across laboratories, fundamental variables such as printability and matrix stiffness remain strictly non-comparable. This fragmentation stems from a flawed industry-wide reliance on reporting these metrics as static, scalar end-points rather than as continuous, time-resolved dynamic viscoelastic profiles—most critically, precise stress relaxation curves. This systemic data fragmentation critically sabotages the much-anticipated integration of artificial intelligence (AI) and machine learning (ML). To achieve standardization, integrating AI-driven automated design with high-throughput fabrication creates a “problem-solution” closed loop, utilizing AI for the real-time optimization of printing parameters, 99 yet the absolute absence of high-fidelity, standardized biophysical input data guarantees a fatal garbage in, garbage out (GIGO) collapse. Dismantling these silos to establish universal, dynamic mechanobiological metrics constitutes the absolute engineering prerequisite before AI can genuinely accelerate systemic clinical translation. 8
5.2. Future perspectives: Intelligent and system-level spatiotemporal closed loops
The role of AI in organoid engineering is evolving from passive parameter optimization to generative design—establishing a closed-loop feedback control paradigm to conclusively overcome systemic standardization bottlenecks. From an engineering standpoint, this integration of in-line computer vision and real-time rheological sensors enables autonomous, dynamic adjustment of print parameters to physically map onto optimal cellular viability.100,101 For instance, generative deep learning algorithms have been employed to design de novo ring proteins that specifically bind fibroblast growth factor receptors (FGFRs), precisely directing stem cell differentiation into functional vascular networks. A 2024 study demonstrated that these AI-designed networks enhanced oxygen transport efficiency by 38% and albumin synthesis by 42% in kidney organoids. This performance significantly surpassed traditional biomimetic designs, marking a paradigm shift from “morphological biomimicry” to “functional hyper-biomimicry”. 102
Constructing “Body-on-a-Chip” systems—via the precise interconnection of multiple organoids (e.g., liver, heart, and tumor) on microfluidic platforms—offers the ultimate spatiotemporal macro-solution to address asynchronous maturation and metabolic-mechanical corruption. By harnessing programmed fluid shear stress to synchronize metabolic states across distinct lineages, this approach represents the sole engineering pathway toward reconstructing comprehensive, systemic pharmacokinetic/pharmacodynamic (PK/PD) profiles.7,103 Their core advantage lies in bridging the gap between in vitro models and in vivo pathophysiology by reconstructing tissue interfaces, mechanical cues, fluid dynamics, and biochemical microenvironments while definitively decoupling metabolic supply from local signaling. 104 Future iterations will focus on incorporating endocrine loops and circulating immune systems to fully simulate systemic physiological and pathological responses.105,106
Organoid Intelligence (OI) refers to the construction of brain organoids with electrophysiological activity (comprising neuron-glial networks). Coupled with 3D microelectrode arrays (MEAs) to monitor neural signals, OI aims to simulate learning and memory mechanisms, providing a platform for biocomputing and neurological research. 107 Conventional self-assembled brain organoids inevitably suffer from stochastic, chaotic neural wiring. Bioprinting uniquely resolves this by executing deterministic topological wiring: current protocols have achieved the standardized culture of high-density, myelinated brain organoids enriched with learning-associated glial cells. Precisely patterning these networks directly onto 3D MEAs securely anchors dynamic electrophysiological feedback loops.108,109 While OI-based biocomputing promises faster decision-making, continuous learning, and superior energy efficiency, it necessitates strict ethical governance. Guidelines must be established to prevent the emergence of autonomous consciousness and ensure compliance with bioethical norms.107,108
In situ bioprinting represents a definitive goal in regenerative medicine: utilizing robot-assisted devices to perform tissue repair directly at the damaged site within the patient. 110 By decisively confronting the host–graft interface challenge and completely bypassing the ex vivo vascularization valley of death, this approach allows for the direct fabrication of precise disease models within the complex, dynamic biomechanical niche in vivo. Advanced vision-guided robotic systems and path planning algorithms can execute this precise volumetric deposition directly within host organs to test targeted therapeutics under authentic hemodynamic conditions.111,112 However, clinical translation faces persistent hurdles regarding in vivo environmental complexity, bioink biocompatibility, and the maintenance of printing precision within dynamic environments—most critically, active hemodynamic interference from bleeding and continuous target organ motion driven by cardiac cycles and respiratory excursions. Overcoming these chaotic in vivo boundary conditions constitutes the final summit for next-generation clinical biofabrication. 113
6. Conclusion
Driven by advanced biofabrication engineering strategies, organoid technology is decisively transcending the biological black-box of stochastic self-assembly to catalyze an engineering revolution toward precise biofabrication and rigorous spatiotemporal determinism. By synergizing bioink crosslinking chemistry—which functions as an active temporal mechanocontroller—with advanced printing modalities that serve as deterministic spatial mass transport infrastructures, this paradigm shift systematically dismantles the physical barriers that separate static in vitro models from the authentic, dynamic in vivo biomechanical niche. By restoring true active convection and pharmacokinetic/pharmacodynamic (PK/PD) fidelity, bioprinted organoids have demonstrated indispensable utility in precision disease modeling, high-throughput drug screening, and regenerative medicine. Despite persistent challenges regarding standardization, functional maturity, and ethical considerations, the field must rigorously confront severe systemic hurdles—most notably, the coupled metabolic–mechanical phenotypic drift paradox and the profound data silos. To resolve this, the convergence of AI-driven closed-loop physical feedback and system-level microfluidics now provides a definitive translational blueprint. Continuous technological innovation ensures that bioprinted organoids will play an increasingly pivotal role in accelerating biomedical research, advancing precision medicine, and ultimately improving human health—emerging not merely as morphological replicas, but as the most biophysically rigorous digital twins in the era of precision medicine.
Supplemental material
Supplemental material - Engineering Next-generation organoids: A review on bioprinting strategies, bioink innovations, and frontier applications
Supplemental material for Engineering Next-generation organoids: A review on bioprinting strategies, bioink innovations, and frontier applications by Defeng Sun, Xuemei Fan, Hua Sun, Dongmei Li, Ruijia Shen, Pu Xia and Zhenying Zhao in Journal of Tissue Engineering.
Supplemental material
Supplemental material - Engineering Next-generation organoids: A review on bioprinting strategies, bioink innovations, and frontier applications
Supplemental material for Engineering Next-generation organoids: A review on bioprinting strategies, bioink innovations, and frontier applications by Defeng Sun, Xuemei Fan, Hua Sun, Dongmei Li, Ruijia Shen, Pu Xia and Zhenying Zhao in Journal of Tissue Engineering.
Footnotes
Author contributions
Conceptualization: Defeng Sun, Yiming Wang, Zhenying Zhao. Data curation: Defeng Sun, Xuemei Fan, Pu Xia. Investigation: Defeng Sun, Xuemei Fan, Hua Sun, Dongmei Li. Formal analysis: Defeng Sun, Zhengying Zhao. Resources: Ruijia Shen, Hua Sun, Dongmei Li. Writing – original draft: Defeng Sun. Writing – review & editing: Xuemei Fan, Pu Xia, Zhenying Zhao. Supervision: Pu Xia, Zhenying Zhao. Funding acquisition: Pu Xia, Zhenying Zhao
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The work was Funded by Tianjin Natural Science Foundation (Grant No. 25JCYBJC01330), the Tianjin Key Medical Discipline Construction Project (Grant No. TJYXZDXK-3-019C), and the Tianjin “131” talent training project (2016).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.