
Abstract
Bone healing is a complex and well-organized process, regulated by various factors ranging from growth factors to hormones, cytokines, mechanical stimuli, and aging. Recently, numerous techniques have been devised to efficiently induce the differentiation of human induced pluripotent stem cells (hiPSCs) to osteoblasts. However, enhancing the efficiency of osteoblast differentiation remains a challenge. Thus, we induced the differentiation of hiPSCs to mesodermal cells through Wnt/BMP signaling based on the generation of hiPSCs. After successful generation of hiPSCs, we induced the differentiation of mesodermal cells to osteoblasts. The results revealed that the runt-related transcription factor 2 (RUNX2)-encoding gene was upregulated from the early differentiation stage; hence, the expression of the mature osteoblast marker was higher compared to that observed in other differentiation stages. In addition, the deposition of substrates in mature bones was observed. The results were confirmed via real-time PCR, Alizarin Red Staining, and Von Kossa staining. The coactivation of Wnt and BMP signaling was shown to rapidly and effectively promote the differentiation of osteoblasts. The findings of this study will provide a foundation for future studies on the mechanism of osteoblast development, as well as the biological and pathological investigations of drug screening and bone regeneration tracing.
Introduction
Human-induced pluripotent stem cells (hiPSCs), which are directly generated from somatic cells, represent a pluripotent cell source capable of differentiating into derivatives of all three embryonic germ layers: endoderm, mesoderm, and ectoderm. 1 This high differentiation potential makes them a promising candidate for tissue engineering.2,3 Currently, treatments for large bone defects and diseases often involve autologous or allogeneic bone grafts.4–6 However, these approaches are limited by supply constraints, potential inflammation, infection, immune responses, lack of mechanical stability, and the limited regenerative capacity of native bone tissue.7–10 Stem cell-based strategies are actively being investigated to overcome these challenges.11,12 While mesenchymal stem cells (MSCs) are commonly employed for osteoblast differentiation, their clinical utility is hindered by insufficient cell numbers and prolonged differentiation periods.13–15 In contrast, hiPSCs, including hiPSCs, are highly suitable for bone tissue engineering due to their robust self-renewal capacity and sustained differentiation potential.
Traditional protocols for the osteogenic differentiation of hiPSCs typically involve the direct application of differentiation media—containing components such as fetal bovine serum (FBS), dexamethasone, beta-glycerophosphate, and ascorbic acid (vitamin C)—to the pluripotent cells.16,17 However, to more closely mimic embryonic osteogenesis in vivo, we introduced an intermediate mesodermal induction step, which has been previously shown to enhance differentiation efficiency.12,13 Given the critical role of paraxial mesoderm in embryonic bone formation, 18 this lineage is of particular interest. Specifically, we have provided a more detailed discussion on the synergistic roles of Wnt/β-catenin, Bone Morphogenetic Protein 4 (BMP4), and FGF signaling, which are critical for activation during the primitive streak-like stage. Furthermore, we have elaborated on how these pathways trigger key transcription factors, such as T (Brachyury) and TBX6, to drive paraxial mesoderm commitment. While numerous studies have explored the direct osteogenic differentiation of hiPSCs,19,20 a fully elucidated method for sequential differentiation via the mesodermal lineage remains challenging.
Osteogenesis proceeds either through the direct differentiation of MSCs into osteoblasts or, in the context of hiPSCs, via differentiation into mesodermal cells followed by subsequent osteoblast commitment.21,22 This process is governed by the sequential expression of key genes and proteins involved in bone mineralization, formation, absorption, and degradation. Essential markers expressed during this lineage progression include collagen type I (COL1), alkaline phosphatase (ALP), and non-collagenous bone matrix proteins such as osteocalcin (OCN) and osteonectin (ON). RUNX2 is the master regulator of osteoblast differentiation and osteogenesis, binding to promoter regions (e.g., OCN) to induce expression and regulate early osteoblast proliferation. 23 Crucially, the Wnt and BMP signaling pathways are indispensable upstream activators of RUNX2 expression during early osteoblast commitment.24,25 Therefore, their co-activation during the initial mesoderm differentiation phase is hypothesized to enhance the rate and efficiency of osteoblast generation.
In this study, we aimed to establish an efficient hiPSC osteogenic differentiation strategy by optimizing the mesodermal induction stage prior to terminal osteogenic differentiation. To this end, hiPSCs were subjected to various experimental conditions during a 9-day mesodermal induction process (Figure 1). Gene and protein expression were quantitatively analyzed using qPCR and immunofluorescence staining, respectively, and mineralization and calcium deposition were assessed by Alizarin Red Staining (ARS) and Von Kossa staining. By comparing the expression of bone-specific genes and functional mineralization outcomes, we identified the conditions most conducive to efficient osteogenic differentiation. Our findings demonstrate that mesodermal induction is a critical determinant of hiPSC osteogenesis. In particular, differentiation toward the paraxial mesoderm lineage, facilitated by co-activation of Wnt/BMP signaling, resulted in the most rapid and effective differentiation into osteoblasts. A schematic showing BMP4-mediated osteogenesis maturation through CHIR-induced mesodermal differentiation through Wnt activation and BMP signaling, efficient osteoblast differentiation with hiPSCs is synergistically promoted through Wnt/BMP signaling. hiPSC, Human induced pluripotent stem cells; CHIR, CHIR99021; BMP4, Bone morphogenetic protein 4.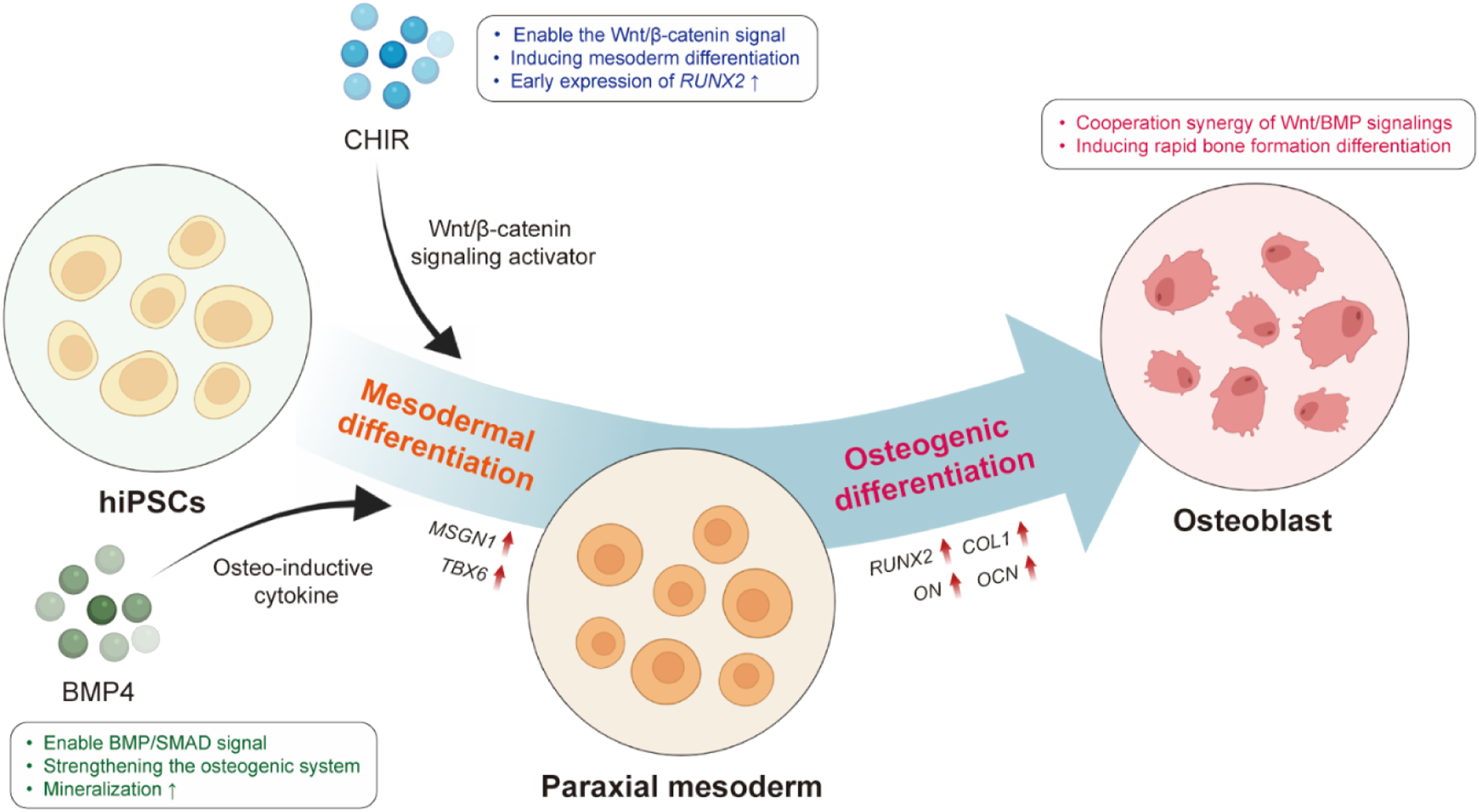
Materials and methods
hiPSCs culture
The hiPSCs used in this study was provided by the National Stem Cell Bank of South Korea, a commercial supplier that received initial ethical approval (Cat. CMC-hiPSC-022). These lines were authenticated by STR analysis and found to be free of mycoplasma, and showed normal nucleoid type (46,XX). The hiPSCs line were maintained in Essential 8 (E8) Basal medium (A15169-01, Gibco, USA) on 6-well plates (Corning, USA) pre-coated with recombinant human vitronectin (rhVTN-n; A14700, Gibco, USA). The hiPSCs were passaged every five days, and the culture medium was replaced daily. For subculturing, hiPSCs were washed once with 1 mL of phosphate buffered saline (PBS) (SH30028.02, HyClone, USA) and subsequently incubated with 500 μL of Accutase (SCR005, Millipore, USA) at 37°C for 3 minutes to facilitate dissociation. Following incubation, 1.5 mL of PBS was added to the culture vessel, and the cell suspension was gently triturated to ensure uniform dissociation before being transferred to a 15 mL conical tube. The suspension was then centrifuged at 850 rpm for 3 minutes. For routine maintenance, the cells were reseeded at a split ratio of 1/40 onto VTN-coated dishes. The remaining cell population was collected by an additional round of centrifugation for cryopreservation. After the supernatant was aspirated, the resulting cell pellet was resuspended in 1 mL of Synth-a-Freeze (A12542-01, Gibco, USA) and transferred into a cryogenic vial (368632, Thermo SCIENTIFIC, USA). The vials were then stored at -80°C. For recovery, cryopreserved vials were rapidly thawed in a 37°C-water bath for 3 minutes. The cell suspension was immediately transferred to a 15 mL tube containing 1 mL of pre-warmed E8 medium and centrifuged at 850 rpm for 3 minutes. The cell pellet was resuspended in fresh growth medium and plated onto VTN-coated dishes for further expansion. hiPSCs between passages 15 and 30 were utilized for differentiation experiments.
Induction of paraxial mesoderm cells from hiPSCs
hiPSCs were differentiated into paraxial mesoderm cells following a published protocol with minor modifications. 18 Briefly, hiPSCs were dissociated using Accutase (SCR005, Millipore, USA) and re-seeded onto Matrigel-coated (354277, Corning, USA) 6-well plates in mesoderm induction medium. The medium consisted of STEMdiff™ APEL™2 Medium (APEL; 05270, STEMCELL, Canada) supplemented with 6 ng/mL StemMACS CHIR99021 (CHIR; 130-103-926, Miltenyi Biotec, Germany) and 50 ng/mL BMP4 (CYT-1093, ProSpec, Korea) for two days. The duration of induction was established based on the temporal expression patterns of T (Brachyury) and TBX6.
To optimize the differentiation efficiency into the paraxial mesoderm lineage, several culture conditions were evaluated using four distinct media formulations. The basal APEL medium was used alone for the MM1 condition, while MM2 was supplemented with CHIR to initiate mesodermal signaling. In MM3, both CHIR and BMP2 (CYT-627, ProSpec, Korea) were added to the APEL medium to assess their synergistic effects, and MM4 involved the combined treatment of CHIR99021 and BMP4 within the same basal medium. These conditions were systematically compared during the early induction phase to identify the most effective signaling combination for robust paraxial mesoderm commitment.
Induction of osteoblasts from paraxial mesoderm
The differentiated paraxial mesoderm was cultured in osteoblast induction medium. This medium comprised Dulbecco’s Modified Eagles Medium (DMEM) high glucose (SH30243.01, HyClone, USA) supplemented with 10% FBS (SH30919.03, HyClone, USA), 1% penicillin/streptomycin (15140-122, Gibco, USA), 0.1 M Dexamethasone (D2915, Sigma-Aldrich, USA), 50 M L-Ascorbic acid (49752, Sigma-Aldrich, USA), and 10 mM beta-glycerophosphate (G9422, Sigma-Aldrich, USA). The medium was replenished every two days.
RNA extraction, cDNA synthesis, and real-time PCR (qPCR) analysis
Total RNA was extracted using the AccuPrep Universal RNA Extraction Kit (K-3140, Bioneer, Korea). Complementary DNA (cDNA) was synthesized from 1 µg of RNA using the MG cDNA Synthesis Kit (MR01611, MGmed, Korea). Quantitative PCR was performed on a QuantStudio 3 system (A28137, Applied Biosystems, USA) using SYBR Green (RR430A, Takara, Japan). Target gene expression levels were normalized to GAPDH and calculated using the ΔΔCt method. Primer sequences are listed in Table S1.
Alizarin Red Staining
Cultured cells were washed twice with PBS and fixed with 4% paraformaldehyde (PFA) (PC2031-100-00, Biosesang, Korea) for 15 min. Fixed cells were stained with 2% ARS solution (0223, ScienceCell, USA) for 30 min at room temperature. After staining, the solution was removed, and the cells were washed five times with distilled water. ARS staining was imaged using a microscope (IX73P2F, Olympus, Japan). Quantitative analysis of mineralization was conducted by eluting the bound stain with a 10% Cetylpyridinium chloride (CPC) solution. The CPC elution protocol was: 10 mL of 10 mM sodium phosphate (BSP-9570, T&I, Korea) mixed with 1 g of CPC (C0732, Sigma-Aldrich, USA). The absorbance of the elevator was measured at 540 nm using an Epoch microplate reader (BioTek, USA).
Von Kossa staining
Cultured cells were washed twice with PBS and fixed with 4% PFA for 30 min at 37°C. Fixed cells were stained using the Von Kossa Stain Kit (KTVKO, MasterTech, USA). After staining, the solution was removed, and the cells were washed three times with distilled water. Von Kossa staining was analyzed using a microscope, and the area of calcium deposition was quantified using ImageJ software (National Institutes of Health, USA).
Immunofluorescence staining
Cells were washed with PBS (SH30028.02, Hyclone, USA) and fixed with 4% PFA at 37°C for 10 min. Permeabilization was performed by treating cells with 0.1% Triton X-100 (1.12298.1001, Merck, USA) at room temperature for 15 min. Non-specific binding was blocked using 2% bovine serum albumin (BSA) (30063-572, Gibco, USA) at room temperature, followed by overnight incubation with primary antibody solution at 4°C. The primary antibodies used were: RUNX2 (D1L7F) Rabbit mAb (1:6400; 12556, Cell Signaling, USA) and COL1A1/Collagen I Recombinant Rabbit Monoclonal Antibody (1:200; Huabio, China). After three washes with PBS, cells were incubated with Alexa Fluor 594 donkey anti-rabbit secondary antibody (1:1000; ET1609-68, Invitrogen, USA) for 1 h at room temperature. Nuclei were counterstained using Antifade Mounting Medium with DAPI (H-2000, Vector Laboratories, USA). Immunofluorescence staining was analyzed and quantified (fluorescently stained areas) using a fluorescence microscope and ImageJ software, respectively.
Chondroitin sulfate cryogel scaffold fabrication
Chondroitin sulfate methacrylate (CSMA) was synthesized by reacting 0.5 g of chondroitin sulfate A sodium salt (GC6763, Glentham, UK) dissolved in PBS with 0.5 mL (74 mM) glycidyl methacrylate (779342, Sigma-Aldrich, Germany) for 10 days. The CSMA solution was purified using a dialysis membrane cassette for 48 h and subsequently dialyzed against deionized water for 24 h. After lyophilization, CSMA powder was stored at -20°C. Chondroitin sulfate (CS) cryogels were prepared by dissolving CSMA in deionized water at 10% w/v at room temperature. Whitlockite was added to the polymer solution at 6% w/v and vortexed to ensure particle dispersion. The mixture was cast into a plastic mold to achieve the desired shape. Cross-linking was initiated using 2.5% v/v thermal initiators (100 mg/mL, APS; 9913, Sigma-Aldrich, USA) and 0.12% v/v accelerators (N, N, N, N-tetramethylethylenediamine, TEMED; T7024, Sigma-Aldrich, USA). The mixture was then lyophilized at -20°C for 20 h to create the cryogel for in vivo experiments.
Cell culture for transplantation
A 4-well plate was covered with cover glass, and both the cover glass and the CSMA cryogel scaffold were coated with Matrigel for 2 h. A total of 1×106 hiPSCs were suspended in Matrigel solution. In parallel, human bone marrow-derived mesenchymal stem cells were prepared via LONZA (PCS-500-012, Switzerland). BMSCs were used in the experiment with cells between passages 4 and 8. They were prepared with BMSCs at the same density as hiPSCs and transplanted under the same in vivo conditions as positive controls. A 100 µL aliquot of the cell-Matrigel mixture was dispensed onto the CSMA cryogel scaffold to facilitate swelling. The scaffold was incubated in a 37°C incubator for 30 min. Subsequently, 1 mL of E8 medium was added, and the culture was stabilized for one day before initiating the differentiation protocol. Upon completion of differentiation, the scaffolds were transplanted subcutaneously into mice and harvested at 1, 4, and 6 weeks.
In vivo osteogenesis differentiation studies
All experimental procedures were carried out in compliance with ethical regulations and were approved by the 4D Biomaterials Animal Care and Use Committee (KNUT IACUC 2024-02). A total of 20 female BALB/c nude mice (CoreTec, Korea) underwent a 1-week acclimatization period in an environmentally controlled facility. Breeding conditions were maintained at a temperature of 23 ± 3°C, a relative humidity of 50 ± 10 %, and a 12-hour light/dark cycle, and feed and water were freely ingested. The experimental groups were divided into: Scaffold only, Bone mesenchymal stem cells (BMSC), CHIR (AC), and CHIR+BMP4 (ACB). Samples were prepared by seeding the cells into the prefabricated CSMA cryogel scaffolds. Before surgery, the animals were treated after obtaining sufficient anesthesia under isoflurane inhalation (657801261, Hana Pharm, Korea) anesthesia, and after the surgery was completed, the incision was treated using a local disinfectant. During the procedure and recovery period, the animal’s condition was closely monitored and managed to meet the institutional welfare standards. At the end of the experiment, euthanasia was performed using the carbon dioxide overabsorption method, and the treatment was performed in a way that gradually increased the carbon dioxide concentration. Scaffolds were taken at 1, 4, and 6, respectively, and fixed with 4% PFA (47608, Sigma-Aldrich, Switzerland).
Micro-computed tomography (Micro-CT) analysis
To evaluate the extent of osteogenic mineralization within the scaffolds, high-resolution micro-computed tomography (micro-CT) imaging was performed using a Quantum GX3 micro-CT imaging system (Revvity, Waltham, MA, USA). Scans were conducted using the standard acquisition mode for 2 minutes at an X-ray tube voltage of 60 kV and a current of 160 µA. An aluminum (Al) filter with a thickness of 0.5 mm was applied. For comprehensive 3D visualization and quantitative analysis, the reconstructed volumetric datasets were imported into Analyze 15.0 software (AnalyzeDirect, Overland Park, KS, USA). The mineralized regions were digitally segmented from the scaffold background by applying a globally uniform radiodensity threshold (Hounsfield Units, HU). Based on the thresholded regions, 3D volumetric models were rendered, and the total volume of the mineralized tissue (mm3) within each scaffold was calculated to quantify the degree of bone-like tissue formation.
Histological staining of CSMA cryogel scaffolds
Scaffolds were processed for tissue staining by embedding in paraffin (CelltisBio, Korea) and sectioning. Paraffin-embedded slides were deparaffinized in xylene (534056, Sigma-Aldrich, USA), gradually rehydrated using a graded series of 70 – 100% Ethanol (NBM203, Duksan, KR), and finally washed with Distilled Water (D.W). For general histological analysis, slides were stained using the Hematoxylin and Eosin (H&E) Stainless Kit (H-3502, Vector Laboratories, USA). Samples were incubated in Hematoxylin solution for 5 min, washed twice with D.W., treated with a Bluing agent solution for 10 – 15 min, washed twice with D.W., briefly exposed to 100% Ethanol, and finally stained with Eosin Y solution for 2 - 3 min. Dehydration was performed by washing three times for 1 – 2 min with 100% Ethanol. To evaluate calcium deposition, fixed samples were incubated with 2% ARS solution (0223, Science Cell™, USA) for 20 min at room temperature, washed three times with D.W. to remove excess dye, and then stored in D.W. to prevent drying. Images were collected using a Zeiss microscope and quantified using ImageJ. For the analysis of calcium salt detection, samples were stained with a Von Kossa staining kit (ab150687, Abcam, UK) according to the manufacturer’s protocol. Image collection and quantification were performed using a Zeiss microscope and ImageJ, respectively.
Immunofluorescence staining of CSMA cryogel scaffolds
Paraffin-removed tissue slides underwent antigen retrieval by incubation with Proteinase K at 37°C for 20 min, followed by cooling at room temperature for 10 min. Permeabilization was achieved by incubating sections with 0.5% Triton-X (T8787, Sigma-Aldrich, USA) solution for 15 min, followed by washing. Blocking was performed using a 1:1 mixture of 0.2% BSA (A7906, Sigma-Aldrich, USA) and 0.1% Triton-X. After washing, primary antibodies were diluted 1:500 in PBS and incubated overnight at 4°C. The primary antibodies used were: COL1A1 (ET1609-68, Huabio), RUNX2 (12556S, Cell Signaling), Osteocalcin (OCN; SC-74495, Santa Cruz), and Osteopontin (OPN; SC-21742, Santa Cruz). Following three washes with 0.1% Triton-X solution, sections were incubated with secondary antibodies (Alexa-Fluor 594, ab150080; Alexa-Fluor 488, ab150113; Abcam, UK) at room temperature for 1 h. After three final washes, the scaffolds were mounted and counterstained with Mounting Medium containing DAPI (AB104139, Abcam, UK) and observed under a Zeiss microscope.
Statistical analysis
Statistical analyses were performed using GraphPad Prism 9. Data are presented as means ± SD. Before statistical tests, the normality of the data distribution was assessed using a diagnostic procedure embedded in the Prism. Sample sizes for each experiment are plotted in the corresponding figure legends. n denotes independent biological repeats, and when technical repeats were included, values were averaged within each biological repeat and then used for statistical analysis. One-way ANOVA was applied to assess differences between groups according to one independent variable was used for comparison between three or more groups, and Tukey’s multiple comparison test was used for postmortem tests. *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001.
Results
Co-activation of Wnt/BMP signaling directs hiPSC differentiation toward paraxial mesoderm
The initial phase of this study aimed to optimize the conditions for the efficient differentiation of hiPSCs into paraxial mesoderm (PM). hiPSCs were cultured on Matrigel-coated dishes using mesoderm differentiation medium (MM) supplemented with varying activators. Traditionally, PM differentiation is carried out over 4 days. However, to account for potential rapid differentiation into off-target lineages (e.g., vascular cells) within 2 days, a comparative time-course analysis was performed at day 2 (D2) and day 4 (D4) to determine the optimal differentiation time point (Figure 2(a)). For paraxial mesenchymal induction, hiPSC was treated in 6 ng/mL CHIR and 50 ng/mL BMP4 for 2 days. Quantitative polymerase chain reaction (qPCR) analysis of the PM-specific marker Mesogenin 1 (MSGN1) revealed an increase in expression at D2 in groups treated with the Wnt activator CHIR (MM2, MM3, and MM4), with average fold changes of 18.35, 21.42, and 20.8, respectively (Figure 2(b)). Similarly, T-box transcription factor 6 (TBX6), another PM marker, showed increased expression at D2 in MM2, MM3, and MM4, with fold changes of approximately 12.4, 8.72, and 14.85, respectively (Figure 2(c)). Notably, the MM4 group, which received simultaneous treatment with CHIR and BMP4, demonstrated the most significant upregulation of both MSGN1 and TBX6. To assess lineage specificity and potential off-target differentiation, we analyzed the expression of lateral mesoderm markers ISL LIM Homeobox 1 (ISL1) and NK2 Homeobox 5 (NKX2.5) (Figures 2(d) and 2(e)). While a general upregulation in both ISL1 and NKX2.5 was observed across the groups, the differences between the MM2, MM3, and MM4 groups at D2 were statistically insignificant. Consequently, the optimal PM differentiation period was established as 2 days. These results strongly suggest that the co-activation of the Wnt signaling pathway (via CHIR) and the BMP signaling pathway (via BMP4) is crucial for robust PM induction. Coactivation of Wnt/BMP signaling specifies the differentiation of hiPSCs to a paraxial mesoderm. (a) A schematic diagram of the protocol of mesoderm according to the differentiation period. (b-e) Graphs of mRNA expression of MSGN1, TBX6, ISL1, and NKX2.5 in mesenchymal-induced cultures (n=3). (f-g) Graphs of MSGN1 and RUNX2 mRNA expression according to the concentrations of BMP2 and BMP4 (n=3 and 5). (h-i) Graphs of TBX6 and RUNX2 mRNA expression for BMP4 (n=5). Groups: PM, Pluripotent stem cell Medium; MM, Mesoderm differentiation Medium; AC, CHIR; ACB, CHIR+BMP4. hiPSC, Human induced pluripotent stem cells; CHIR, CHIR99021; BMP2, Bone morphogenetic protein 2; BMP4, Bone morphogenetic protein 4. All data represent mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001.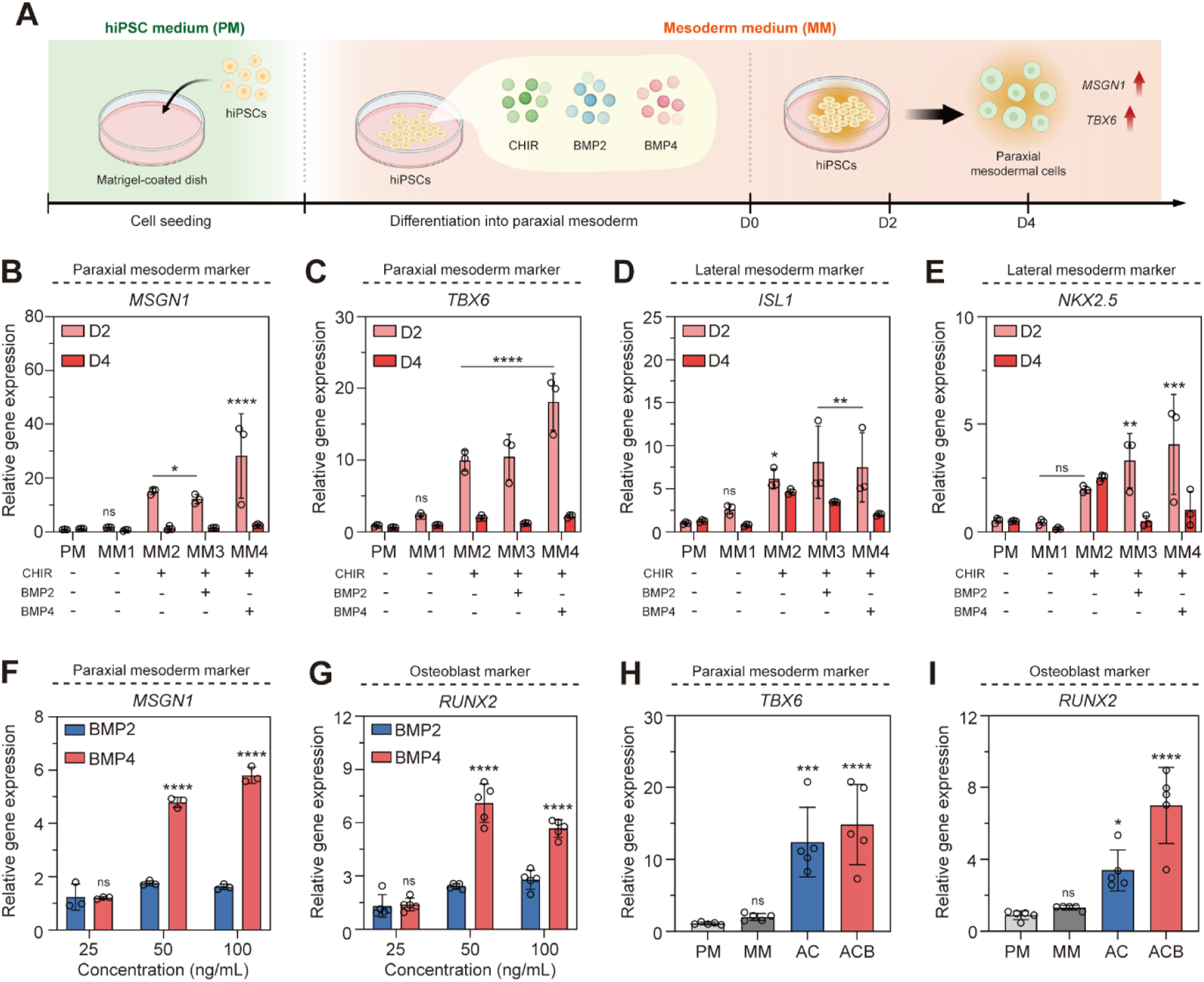
Next, we compared the efficiency of BMP2 and BMP4 at various concentrations (25, 50, and 100 ng/mL) to further optimize PM differentiation. qPCR analysis of the PM marker MSGN1 showed that BMP4 treatment consistently resulted in higher differentiation efficiency than BMP2, with fold differences of 2.72 times at 50 ng/mL and 3.58 times at 100 ng/mL (Figure 2(f) and S1). Furthermore, the expression of RUNX2, a key transcription factor indicative of osteoblast differentiation potential, was also significantly higher with BMP4 treatment: 2.93 times higher at 50 ng/mL and 2.04 times higher at 100 ng/mL compared to BMP2 (Figure 2(g)). Based on these findings, a concentration of 50 ng/mL BMP4 was selected for further analysis. Comparing the control PM group (PM), the CHIR-only group (AC), and the CHIR/BMP4 co-treated group (ACB), the gene expression of TBX6 was 11.45-fold higher in the AC group and 13.7-fold higher in the ACB group compared to PM. Similarly, RUNX2 expression was 3.82-fold and 7.9-fold higher in the AC and ACB groups, respectively (Figures 2(h) and 2(i)). These results confirm the establishment of a robust protocol for inducing paraxial mesoderm differentiation using 50 ng/mL BMP4 in conjunction with CHIR and validate the use of RUNX2 as an early indicator of osteogenic commitment.
Co-activation of Wnt/BMP signals in paraxial mesodermal differentiation promotes osteogenic commitment
To investigate the effects of the 2-day mesodermal induction on subsequent osteogenic fate, the transcriptional profiles of 75 osteogenesis-associated genes were analyzed in hiPSCs, AC, and ACB groups (Figure 3(a)). Heatmap analysis clearly demonstrated distinct gene expression patterns in the AC and ACB groups compared to the undifferentiated hiPSC group, particularly concerning Wnt/BMP signaling-related genes. Twelve key genes were selected for detailed analysis (Figure 3(b)). Notably, the ACB group showed upregulated expression of mesodermal and BMP-responsive transcription factors, including Dickkopf-1 (DKK1), Msh Homeobox 1 (MSX1), and MSGN1. Conversely, the hiPSC group exhibited low overall expression, and the AC group showed only partial increases in select genes. Total RNA Sequencing. (a) Transcription profile of 75 genes relevant to osteogenesis for hiPSC cultured for 2 days in mesoderm differentiation medium. Relative expression: red (high) and blue (low) (b) Heatmaps for markers related to osteogenesis. Relative expression: red (high) and green (low) (c) Gene ontology enrichment dot plot. Relative expression: red (high) and big dot (high). Groups: AC, CHIR; ACB, CHIR+BMP4. hiPSC, Human induced pluripotent stem cells.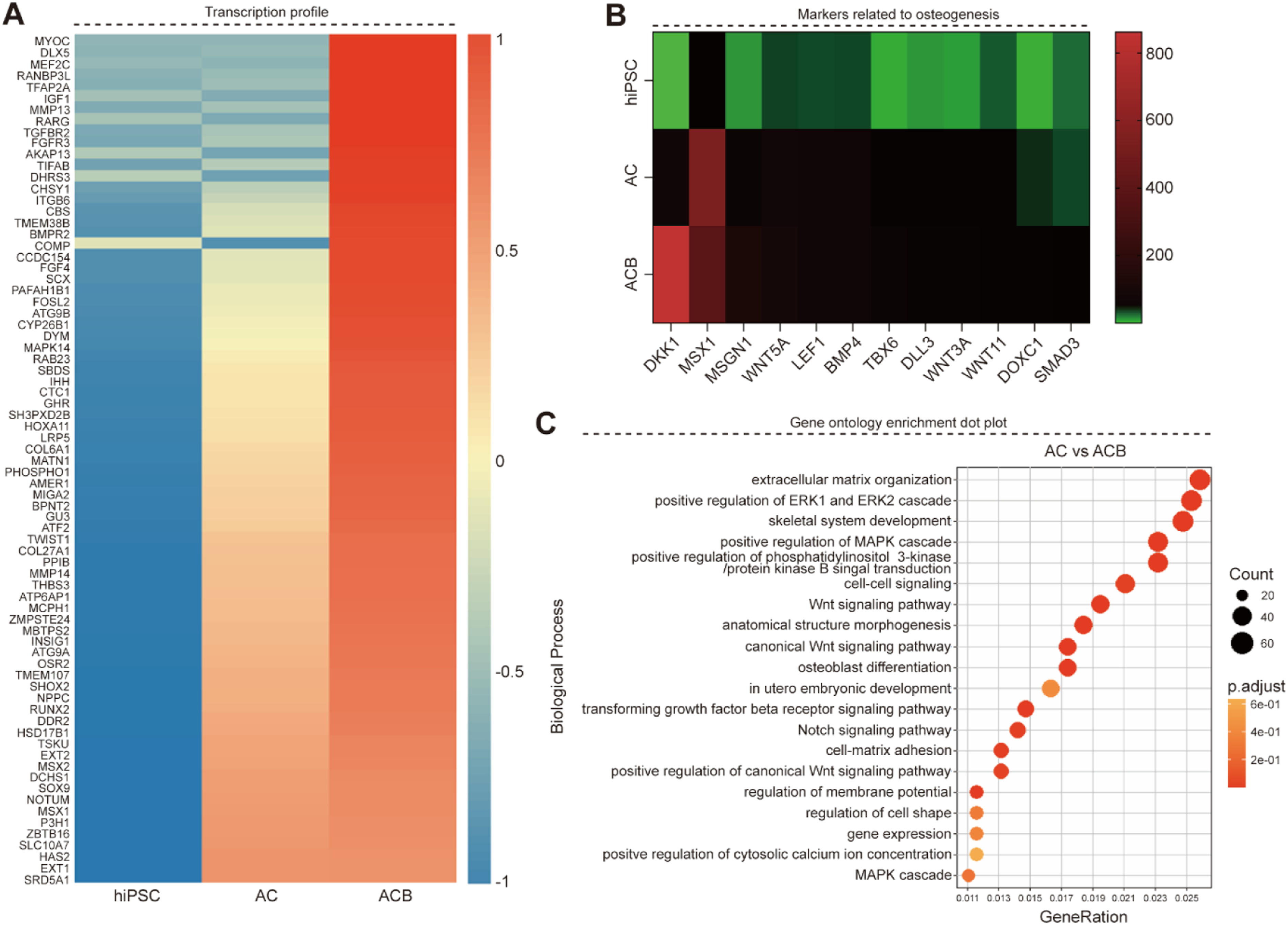
Gene Ontology (GO) analysis of the differentially expressed genes (DEGs) between the AC and ACB groups revealed significant changes in biological processes associated with major signaling cascades, such as the MAPK cascade and Wnt signaling pathway, as well as cell structure and differentiation-related processes (Figure 3(c)). The highest number of DEGs and the greatest proportional changes were observed in GO terms related to extracellular matrix organization, positive regulation of the ERK1/ERK2 cascade, and skeletal system development. These findings collectively suggest that the simultaneous activation of Wnt signaling (CHIR) and stimulation by BMP4 strongly promotes mesodermal transformation and effectively primes the transcription factor network for subsequent entry into the osteogenic lineage. Therefore, the ACB treatment condition demonstrated superior efficacy in promoting both mesoderm induction and downstream osteogenic commitment compared to existing conditions.
Limited BMP4 activation during the mesoderm period enhances osteoblast maturation
Following the successful establishment of the 2-day paraxial mesoderm differentiation protocol (Stage 1), we varied the subsequent BMP4 treatment duration during the 7-day osteoblast differentiation period (Stage 2) (Figure 4(a)). Cellular morphology observation confirmed stable cell growth throughout the 9-day Stage 2 period (Figure 4(b)). RT-PCR analysis of the early osteoblast marker RUNX2 confirmed that the addition of BMP4 (compared to the control group) enhanced osteoblast differentiation by 2.42-fold (Figure 4(c)). Importantly, the co-treatment of CHIR and BMP4 (at both 50 ng/mL and 100 ng/mL) during Stage 1 further improved RUNX2 expression by 2.73 and 3.57 times, respectively, underscoring the benefits of co-activating the Wnt and BMP pathways during the initial mesodermal phase. When evaluating Collagen Type I (COL1) expression, the group co-treated with CHIR and BMP4 at 50 ng/mL showed a 1.6-fold enhancement compared to the BMP4-only group, whereas the 100 ng/mL co-treatment did not yield significant improvement (Figure 4(d)). Limited activation of the BMP signaling in the mesoderm period promotes the maturation of osteoblasts. (a) Schematic protocol for osteoblast differentiation via mesoderm. (b) Optical images of hiPSC according to differentiation (X40). (c-d) Graphs of mRNA gene expression of RUNX2 and COL1 according to BMP4 concentration. (e-g) Graphs of mRNA expression of COL1, ON, and OCN. ON: osteonectin, OCN: osteocalcin. Scale bar: 500 μm. Groups: DAG, Dorsal axial mesoderm; APEL, Albumin, polyvinyl alcohol, essential lipids; AC, CHIR; ACB, CHIR+BMP4. hiPSC, Human induced pluripotent stem cells; CHIR, CHIR99021; BMP4, Bone morphogenetic protein 4. All data represent mean ± SD (n=5). *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001.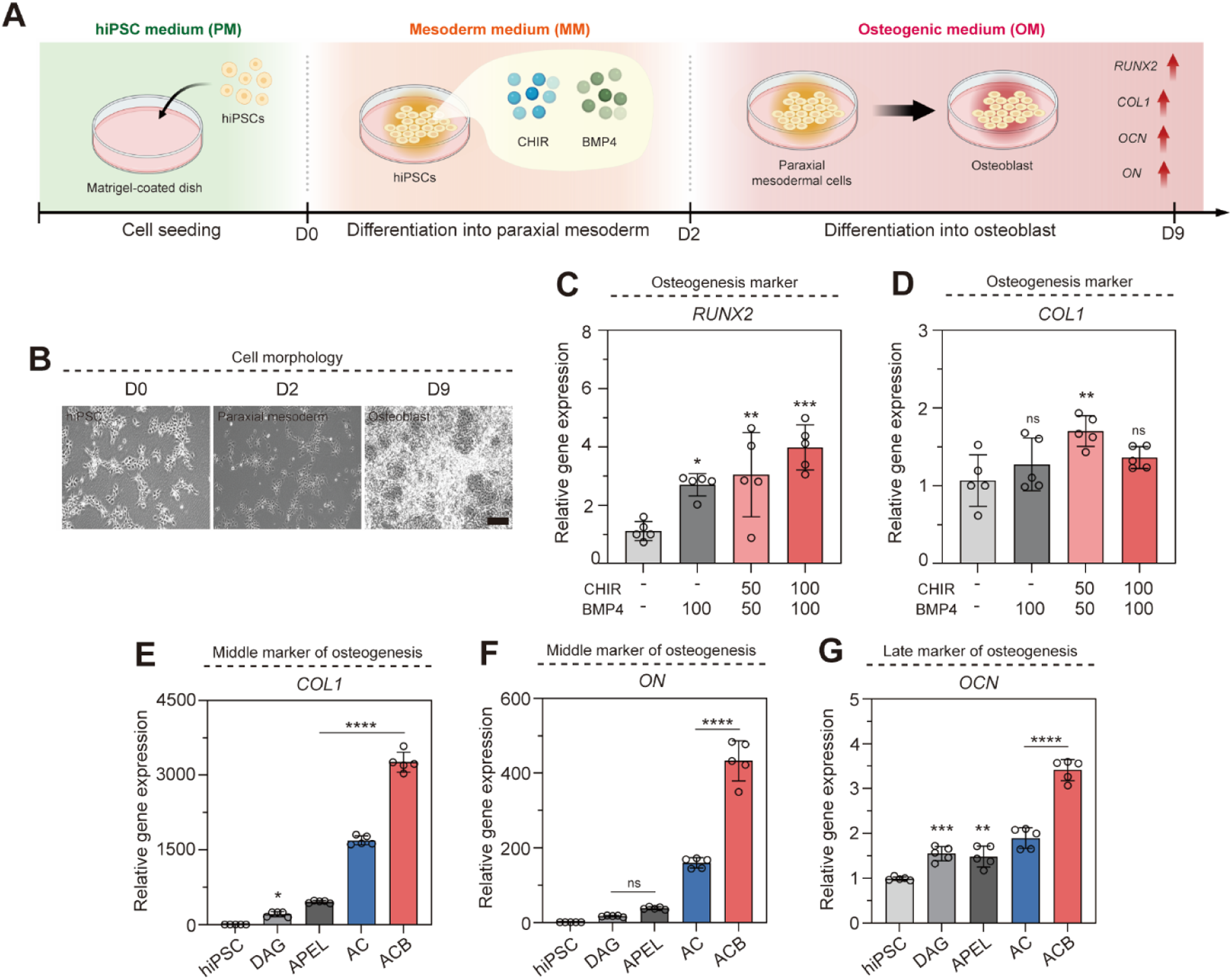
Detailed analysis comparing hiPSC, mesoderm-induced media (DAG; Dorsal axial mesoderm, APEL; Albumin, polyvinyl alcohol, essential lipids), AC, and ACB groups confirmed that the AC and ACB groups exhibited higher expressions of key bone formation markers: COL1, osteonectin (ON), and osteocalcin (OCN) (Figures 4E–4(g)). The ACB group showed a significant advantage over the AC group, with expression increases of 1.93-fold (COL1), 1.8-fold (ON), and 2.71-fold (OCN). These results establish that Wnt/BMP co-signaling during the mesodermal period not only promotes osteoblast differentiation but also significantly enhances osteoblast maturation.
Co-activation of Wnt/BMP signaling promotes osteoblast maturation and mineralization
We assessed the terminal outcome of osteoblast differentiation by analyzing calcium deposition and matrix protein expression. Cellular morphology remained stable across all groups over the 9-days (Figure 5(a)). ARS, a classic measure of calcium deposition, revealed the most intense staining in the ACB group (Figure 5(b)). Quantitative spectrophotometric analysis (absorbance at 540 nm) confirmed that the ACB group exhibited a 1.81-fold higher staining level compared to the AC group (Figure 5(c)). Von Kossa staining, which detects calcium phosphate and other mineral salts, similarly showed a greater area of black staining in the ACB group (Figure 5(d)), indicating superior calcium deposition. Quantification of the stained regions confirmed the ACB group was 3.64-fold higher than the AC group (Figure 5(e)). Coactivation of Wnt/BMP signaling facilitates the maturation of osteoblasts. (a) Optical images of hiPSC for 9 days with or without BMP4 (X40). (b) ARS images of hiPSC on day 9 (X40). (c) Quantification graph of ARS absorbance at 540 nm wavelength (n=4). (d) Von Kossa images of hiPSC on day 9 (X100). (e) Quantification graph of Von Kossa+ area (n=6). (f) Immunofluorescence staining images for hiPSC after osteogenic differentiation (X200). (g) Quantification graph of RUNX2+ and COL1+ area (n=3). Scale bars: (a-b) 500 μm, (d) 200 μm, (f) 100 μm. Groups: AC, CHIR; ACB, CHIR+BMP4. All data represent mean ± SD. *P < 0.05, **P < 0.01, and ****P < 0.0001. The symbol * indicates comparisons with the AC group.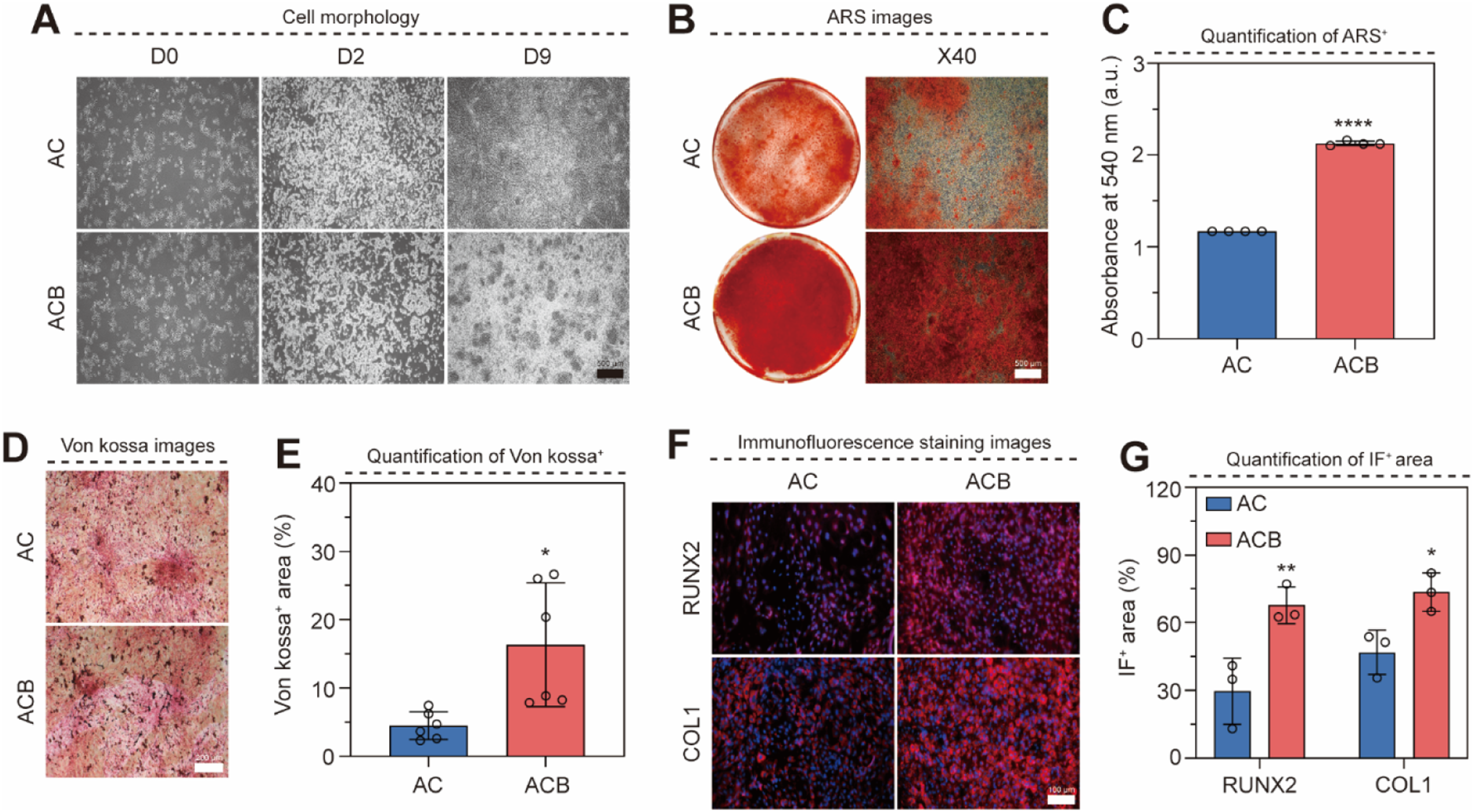
Immunofluorescence staining was used to evaluate osteoblast maturity (Figure 5(f)). The expression of both RUNX2 (early marker) and COL1 (middle/late marker) was more pronounced in the ACB group. Quantification of the expression area showed that the ACB group was 2.29-fold higher in RUNX2 and 1.57-fold higher in COL1 compared to the AC group (Figure 5(g)). These results unequivocally demonstrate that Wnt/BMP co-activation significantly increases calcium deposition and promotes osteoblast differentiation and maturation in vitro.
In vivo bone formation assessment
To validate the in vitro findings, an in vivo bone formation experiment was conducted using a chondroitin sulfate-methacrylate (CSMA) cryogel scaffold (Figure S2) [16]. The CSMA cryogel, known for the biocompatibility of chondroitin sulfate [17], provides a stable microenvironment for cellular activities [18]. In this study, CSMA cryogel scaffold was used as a three-dimensional (3D) cell carrier to help the survival and bone differentiation of hiPSC-derived cells. The scaffold maintains cells at the transplant site and provides an environment suitable for early bone formation, and to evaluate bone formation in vivo, the cells were seeded on a cryogel scaffold and differentiated into osteoblasts that had previously differentiated into osteoblasts through the mesoderm. And the scaffold containing cells was subcutaneously implanted into mice, collected at 1, 4, and 6 weeks, and analyzed (Figure 6(a)). BMSC group served as positive control. The body weight remained overall stable throughout the 6-week observation period, and no statistically significant difference was found between the groups. These results indicate that the scaffold and transplanted cells are well tolerated in vivo, supporting biocompatibility (Figure 6(b)). Observation of the collected scaffolds showed a decrease in size by 6 weeks, confirming the expected biodegradation in vivo (Figure 6(c)). The collected scaffolds were photographed by micro-CT for calciumization (Figure 6(d)). At week 1, it was confirmed that the area of calcium in the ACB group was larger than that of other groups, and the area of calcium compared to the total volume was higher in the ACB at week 6. In the quantification of bone volume to total volume, the ACB group at week 1 was 8.5 times higher than that of the Scaffold group, and at week 6, the bone volume was 4.96 times (Scaffold group), 3.19 times (BMSC group), and 5.28 times (AC group) higher, showing effective calcium (Figure S3). H&E staining at weeks 1, 4, and 6 confirmed that cells were well-localized within the scaffold pores, further demonstrating biocompatibility and cell attachment. Degradation was visibly progressing by week 6 (Figure 6(e)). Differentiation of bone formation in vivo. (a) A schematic diagram of in vivo experiments. (b) Graph of mouse weight change. (c) Images of scaffold incubated for 6 weeks in vivo. (d) 3D rendering images for micro-CT image of scaffold. (e) Images of H&E on scaffold (X200). (f) Images of von Kossa staining on the scaffold (X200). Scale bars: (c) 50 mm, (d) 5 mm, (e-f) 100 μm. Groups: Control, no treatment; Scaffold, scaffold only; BMSC, Scaffold+BMSC; AC, Scaffold+BMSC+CHIR; ACB, Scaffold+BMSC+CHIR+BMP4. All data represent mean ± SD (n=3). *P < 0.05, **P < 0.01, ***P < 0.001, and ****P < 0.0001. The symbol * indicates comparisons with the Scaffold group.
ARS and Von Kossa staining were performed on the collected scaffolds (Figure 6(f) and S4). Calcium salt deposition was first observed via Von Kossa staining at 4 weeks and became more extensive at 6 weeks (Figure S5). Quantification of the Von Kossa-stained area showed no significant difference among groups at Week 1. However, at Week 4, the expression increased compared to the Scaffold-only control in the order: BMSC (2.8 times), AC (3.4 times), and ACB (3.7 times). At Week 6, the trend continued, with expression in BMSC (1.5 times), AC (1.6 times), and ACB (2.3 times), confirming that the ACB treatment group yielded the highest level of in vivo calcium salt deposition and bone matrix formation. The accelerated mineralization observed in immunodeficiency mice was found to be associated with the initial osteogenic activity of transplanted cells.
Analysis of bone formation markers in the scaffold by immunofluorescence staining
Immunofluorescence staining of the in vivo scaffolds analyzed the expression of key osteogenic markers over time (Figure 7(a)). At Week 1, the AC and ACB groups showed clearer expression of Osteopontin (OPN) (a middle marker) than RUNX2 (an early marker), suggesting rapid commitment. OPN expression intensified at Weeks 4 and 6. Importantly, the ACB group consistently showed a wider range and clearer expression of OPN throughout the subcutaneous implantation period, confirming superior osteogenic differentiation. COL1 expression was observed in the BMSC and AC groups at Week 1 (Figure 7(b)). OCN (a late maturation marker) expression, however, was observed in the ACB group starting from Week 1, suggesting accelerated maturation. By Week 6, OCN expression was increased in both AC and ACB groups, indicative of effective mineralization. Analysis of bone formation in the scaffold by immunofluorescence staining. (a-b) Immunofluorescence staining image (X630). (c-f) Fluorescence intensity quantification graph. Scale bars: (a-b) 50 μm. All data represent mean ± SD (n=3). *P < 0.05, **P < 0.01, and ****P < 0.0001. The symbol * indicates comparisons with the Scaffold group.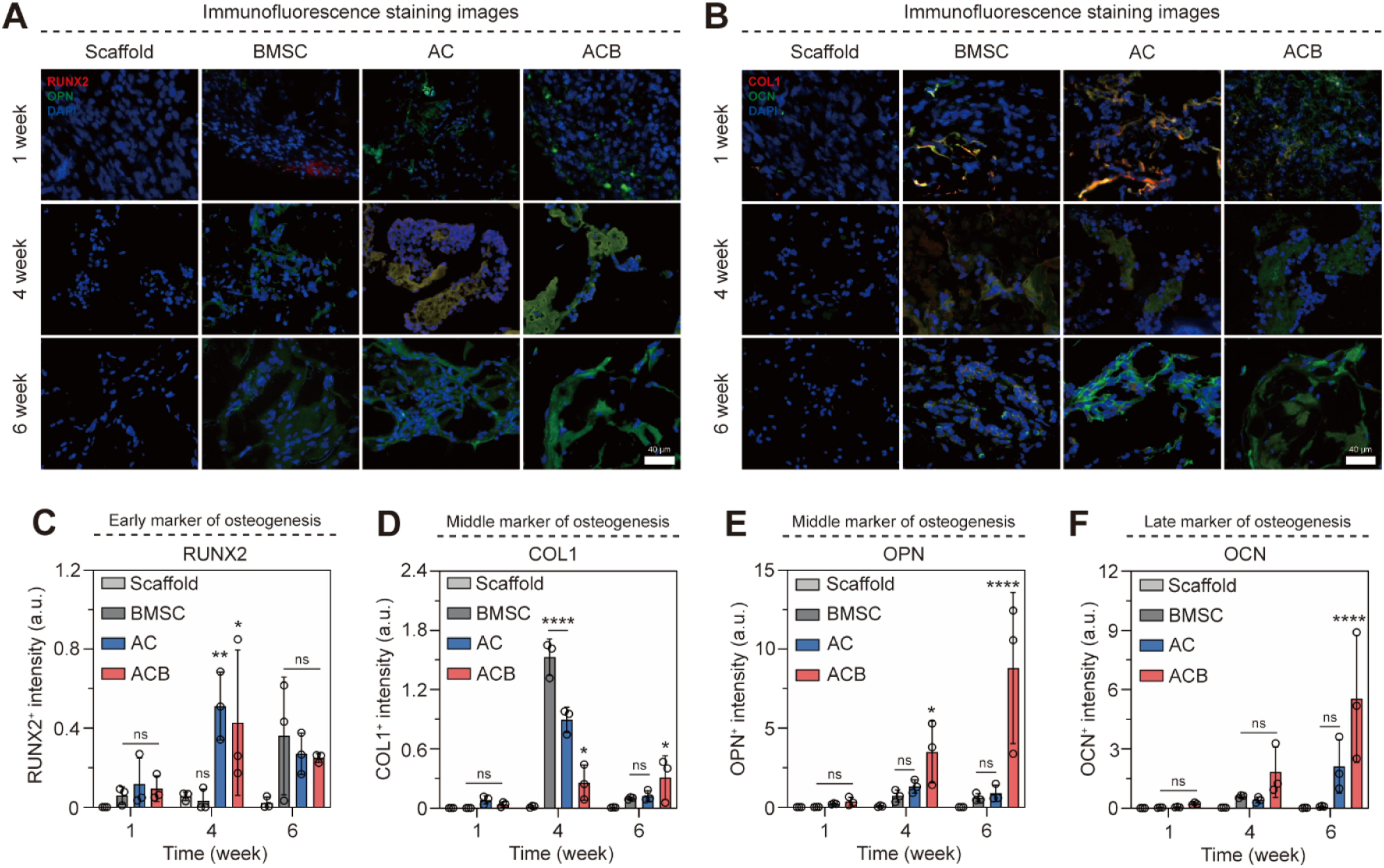
Quantitative analysis of the osteogenic markers (Figure 7(c)) revealed a reduction in early marker expression from Week 4 to Week 6: RUNX2 was reduced in AC (0.53 times) and ACB (0.37 times), and COL1 was reduced in AC (0.12 times) and ACB (0.32 times). This sequential increase followed by a decrease in early/middle markers confirms the expected progression toward terminal bone formation and maturation. Crucially, the ACB group demonstrated a significant increase in late-stage maturation markers from Week 4 to Week 6: OPN (5.01 times) and OCN (5.00 times). These results definitively establish that the Wnt/BMP co-activation protocol (ACB) promotes the most effective and accelerated bone formation differentiation in vivo.
Discussion
The current study critically investigated the sequential induction of hiPSCs to the osteogenic lineage, specifically focusing on the pivotal role of Wnt and BMP signaling co-activation during the initial mesodermal phase. Our findings confirm that while Wnt/BMP signaling is known to be upregulated during standard osteoblast differentiation, their strategic co-activation at early and upstream commitment stages significantly enhances both differentiation efficiency and maturity.26–28 Building on previous studies, we precisely titrated the concentrations of small molecules and growth factors to ensure robust commitment to the paraxial mesoderm.26–28 Specifically, the combination of CHIR and BMP4 provided the necessary signaling threshold to direct progenitor identity toward the paraxial mesoderm rather than the lateral plate mesoderm. Beyond simply enhancing temporal efficiency, the two-stage protocol developed in this study represents a significant advancement by delineating a lineage-specific osteogenic route via the PM. Unlike conventional approaches that primarily focus on terminal osteogenesis, our method more faithfully mimics embryonic skeletal development by orchestrating early signaling pathways—specifically Wnt and BMP—to direct progenitor commitment toward a PM identity.
Although it is already well known that Wnt and BMP signals are involved in bone differentiation, the contribution of this study is not the novelty of these pathways themselves, but the temporal and system-specific activation in the paraxial mesoderm induction phase. In other words, this study showed that by regulating the Wnt and BMP signals together in a specific embryological window, it is possible to effectively induce hiPSC to the paraxial mesoderm-like intermediate stage and then to induce rapid estrogenic progression. We demonstrated quantitatively that activation of the Wnt pathway alone (AC group) was insufficient to achieve the maximal level of osteoblast differentiation and subsequent calcium deposition. Specifically, the combined treatment (ACB group) involving both the Wnt agonist CHIR and BMP4 during the 2-day mesodermal induction phase led to significantly higher outcomes in all measured osteogenic parameters. This superior outcome is mechanistically attributed to the enhanced and accelerated upregulation of the master osteogenic transcription factor, RUNX2.29,30 RUNX2 is indispensable for bone tissue generation, regulating both the proliferation of immature osteoblasts and their terminal maturation by controlling the expression of key matrix proteins, including COL1, ON, and OCN. 27 This highlights that for differentiation protocols starting from pluripotent stem cells, the early, synergistic interplay between Wnt and BMP signaling is essential for rapidly committing cells to the paraxial mesoderm lineage, thereby establishing a robust transcriptional foundation that drives successful, high-level osteoblast maturation. 31 In particular, the observed changes in expression are consistent with the paraxial mesoderm specification and estrogenic priming, suggesting that the differentiation efficiency may be more dependent on the developmental stage and the lineage context than on the simple signal activation itself. Further verification with specific antagonists for Wnt and BMP requires a more sophisticated mechanical interpretation. Therefore, it will be necessary to verify this mechanical interpretation more precisely through inhibitor-based pathway analysis in future studies.
In the early stages of hiPSCs differentiation, qPCR results show a gradual increase in bone formation markers, including RUNX2, suggesting activation of bone formation programs. In addition, the distinct mineral deposits observed in micro-CT and Von Kossa staining confirmed the functional maturation of the differentiated cells and the progression of extracellular matrix mineralization. The subsequent rapid upregulation of RUNX2 by Wnt/BMP stimulation suggests that it may have contributed to the efficient conversion of these cells to the osteogenic family. These results support that hiPSC-based osteogenesis proceeds through a biologically valid pathway, but further studies are needed to determine whether this process involves a transient chondrotic intermediate stage or follows an endochondrotic ossification-like mechanism.32,33
A significant achievement of this work is the development of a rapid and highly efficient in vitro differentiation method. By precisely optimizing the critical commitment phase to 2 days of paraxial mesoderm induction followed by a short 9-day osteogenic maturation, we have substantially decreased the overall differentiation period required to achieve high levels of osteogenic expression and mineralization compared to many conventional protocols. This novel two-stage protocol—which strategically leverages the high inductive capacity of Wnt/BMP co-activation—resulted in: 1. Accelerated Differentiation Markers: OCN expression, a late marker of terminal maturation, was detected in the ACB group in vivo as early as Week 1 (Figure 7(b)), far sooner than in the BMSC or AC groups. 2. Superior In Vivo Performance: In the subcutaneous implantation model, the ACB group consistently demonstrated the highest levels of mineral deposition at both 4 and 6 weeks, culminating in a 2.3-fold increase in calcium salts compared to the scaffold-only control at Week 6 (Figure 6(f)). 3. Terminal Maturation: The significant increase in late markers OPN (5.01-fold) and OCN (5.00-fold) between Weeks 4 and 6 in vivo (Figure 7(c)) confirms that our protocol yields osteoblasts capable of full terminal maturation and functional bone matrix secretion within a clinically relevant timeframe.
In this study, we focused on histological and molecular analyses to evaluate the potential for functional bone formation. Although mechanical strength testing would provide valuable complementary information, the progressive in vivo degradation of the scaffold limited the direct interpretation of mechanical properties over time. Accordingly, further studies including structural and mechanical evaluations will be important for more comprehensive validation of the system. While the subcutaneous implantation model does not fully replicate the biomechanical environment of a load-bearing skeletal site, it serves as a rigorous platform to evaluate the intrinsic osteoinductive capacity of our engineered constructs. Unlike orthotopic models, where the surrounding bone microenvironment provides endogenous signals, the subcutaneous environment is non-permissive for osteogenesis. Therefore, the robust in vivo mineralization achieved here provides definitive evidence that our 2-stage protocol generates cells with autonomous therapeutic efficacy. This ectopic validation is a crucial prerequisite, ensuring that the observed bone formation is inherently driven by the implanted system rather than the host’s innate regenerative response.
Supplemental material
Supplemental material - Synergistic Wnt/BMP Co-activation accelerates osteogenic differentiation of human pluripotent stem cells via paraxial mesoderm induction
Supplemental material for Synergistic Wnt/BMP Co-activation accelerates osteogenic differentiation of human pluripotent stem cells via paraxial mesoderm induction by Hongsik Kim, Sujin Kim, A-Hyeon Kim, Hamyoung Lee, Seongwoo Jeong, Hayoun Kim, Jungbum Kim, Wonhwa Lee, Hwan Drew Kim in Journal of Tissue Engineering
Footnotes
Ethical considerations
The hiPSCs used in this study were purchased from the National Stem Cell Bank in Korea, a commercial supplier that received initial ethical approval. All animal procedures adhered to ethical regulations and were approved by the 4D Biomaterials Animal Care and Use Committee, Korea (KNUT IACUC 2024-02).
Author contributions
H.K.: Investigation, Analysis, Methodology, Writing original draft. S.K.: Investigation, Analysis, Methodology, Writing original draft. A.K., H.L., S.J., H.K., J.K.: Analysis, Methodology. W.L.: Supervision. H.D.K.: Conceptualization, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Korean Fund for Regenerative Medicine (KFRM) grant funded by the Ministry of Science and ICT, the Ministry of Health & Welfare (Code: KFRM 22A0105L1-11) and Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (RS-2025-25400034)
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data Availability Statement
The data are freely available upon request.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.