
Abstract
Background
MicroRNAs (miRNAs) are small non-coding molecules regulating gene expression. Recently circulating miRNAs could be detected in the plasma, serving as novel biomarkers. Different forms of exercise mobilize progenitor cells from the bone marrow, helping in tissue repair. Data of different forms of exercise on endothelial cell damage are lacking. The aim of the study was to evaluate the impact of different exercise modalities on the plasma concentration of miRNA-126, as a marker for endothelial damage.
Methods
The plasma concentration of miRNA-126 and miRNA-133 (marker for muscle damage) was assessed by qRT-PCR analysis in plasma samples from healthy individuals performing one of the following exercise tests: (1) maximal symptom-limited exercise test, (2) bicycling for 4 h, (3) running a marathon, and (4) resistance exercise.
Results
A maximal symptom-limited exercise test resulted in a significant increase of circulating miRNA-126 at maximum power (2.1-fold versus begin), whereas the concentration of miRNA-133 remained unchanged. In line, four hours of cycling increased plasma concentration of miRNA-126 with a maximum 30 minutes after begin (4.6-fold versus begin) without an impact on miRNA-133 concentration. Finishing a marathon race resulted in an increase of miRNA-126 and miRNA-133. In contrast, eccentric resistance training led to an isolated increase of miRNA-133 level (2.1-fold versus begin) with unchanged miRNA-126.
Conclusion
Different endurance exercise protocols lead to damage of the endothelial cell layer as evident by an increase in miRNA-126. On the other hand, resistance exercise has no impact on the endothelial cells, but leads to a destruction of muscular cells.
Introduction
MicroRNAs (miRNAs) are short (∼20–30 nucleotides), highly conserved, functional, non-coding regulatory molecules that play a key role in regulating gene expression by inhibiting protein translation or enhancing messenger RNA degradation (reviewed in Small and Olson 1 ). The human genome consists of more than 1000 miRNAs 2 and their participation in cardiovascular disease was recognized during recent years.3,4 Besides the localization of miRNAs in the cytoplasm, recent studies reported circulating miRNAs in the serum and plasma in a variety of diseases including cancer, stroke and heart disease. 5 It has even been suggested that these circulating miRNAs can be used as minimally-invasive biomarkers. 6 For example, miRNA-499, which is produced almost exclusively in the heart, was significantly elevated in individuals with acute myocardial infarction. 7
In recent years it became evident that exercise training is a very powerful therapeutic strategy to counteract the development and progression of cardiovascular disease (for review see Lavie and Milani 8 ). One important target of increased physical activity is, besides the peripheral skeletal muscle, 9 the endothelial cell layer.10,11 It could be clearly documented that especially in patients with endothelial dysfunction, an exercise training programme leads to an improvement of nitric oxide bioavailability 12 and increased endogenous repair via bone-marrow derived progenitor cells.13,14 Performing not only an exercise training programme over several weeks, but also a much shorter exercise test, like bicycling for 4 h 15 or a single cardiopulmonary exercise test, 16 resulted in an increased number of circulating endothelial progenitor cells. Based on the actual concept, that endothelial progenitor cells are mobilized from the bone marrow, helping to repair a damaged endothelial cell layer, 17 we have to assume that increased physical activity leads to a damage of the endothelial cell layer. Measuring endothelial cell derived microparticles as a marker for endothelial damage after three maximal exercise tests, no significant rise in the plasma was evident. 18 In contrast, in measuring circulating mature endothelial cells (CECs) as another marker for endothelial damage 19 it became evident that an exercise stress test (Bruce protocol) in patients with coronary artery disease 20 or a 4-h bicycling test in healthy individuals 15 resulted in a damage of the endothelial cell layer.
To shed some light onto these discrepancies concerning the influence of exercise on the endothelial cell layer and its damage, we analysed the concentration of miRNA-126, which is highly expressed in endothelial cells, in the situation of four different exercise protocols in healthy individuals. In addition, the plasma concentration of miRNA-133 as a marker for muscle damage was assessed in all individuals.
Methods
Participants and study design
To address the issue of exercise-induced damage of the endothelial cell layer four different studies with different exercise forms and intensities/duration were conducted. None of the healthy individuals had a history of somatic diseases and none of them were taking medications.
Study 1: single symptom-limited exercise test
A total of 13 healthy individuals were included into this study. All participants performed a maximal spiroergometry starting at 50 W with an increase of 25 W every 2 min. Blood samples were taken at baseline, at the end of each interval and 5 min after finishing the exercise test. Plasma was prepared by centrifugation (10 min at 2400 × g) and stored at −80℃ until used.
Study 2: bicycling for 4 h below the anaerobic threshold
Twelve healthy well trained men performed a cycling test on a bicycle ergometer for 4 h at 70% of the individual anaerobic threshold. Blood samples were taken at baseline and at pre-specified time points during the race (5, 10, 15, 30, 60, 120, 180, 240 min) as well as 1 and 24 h after finishing, as recently described. 15 Plasma was prepared by centrifugation (10 min at 2400 × g) and stored at −80℃ until used.
Study 3: marathon race
This study included 22 male middle-aged marathon runners with no history of coronary artery disease, as recently described. 21 Participants were enrolled in the study only if they had successfully finished at least five marathon races in the past three years and had no history of structural heart disease. Exclusion criteria comprised history of established heart disease, diabetes mellitus, angina pectoris, renal failure, musculo-skeletal disease at inclusion preventing future regular marathon running, psychiatric disease, and unwillingness to give informed consent. At baseline (just before the race) and directly after finishing the marathon blood was drawn and plasma was collected by centrifugation (10 min at 2400 × g) and stored at −80℃ until used
Study 4: singular resistance training with additional eccentric loads
Eleven trained subjects performed three different resistance exercises (lat pulldown, leg press and butterfly) on electronically controlled Milon training machines. All exercises were performed with three sets of 15 repetitions and a 1-min pause between sets. All subjects trained with an increased eccentric load of 25% of the concentric load. One week before the actual test the training load for each of the three exercises was determined in a trial. The participants rated the load analogue to the Borg-RPE-Scale (17–18) as ‘very hard’ in the concentric work. On the day of the actual test blood samples were taken before the resistance training, immediately after and 1 h after the training session. Plasma was prepared by centrifugation (10 min at 2400 × g) and stored at −80℃ until used.
Quantification of miRNA by RT-PCR
For all experiments total RNA was isolated from 100 µl of plasma (Qiagen miRNAeasy, Hilden, Germany). To allow for normalization of sample-to-sample variation in RNA isolation and reverse transcription, we supplemented the samples with Caenorhabditis elegans miRNA-39 (cel-miRNA-39) as previously described.22,23 An aliquot of the isolated RNA was reverse transcribed using the TaqMan microRNA reverse transcription kit and the respective primer (miRNA-126, miRNA-133, cel-miRNA-39, ABI, Foster City, CA, USA) according to the manufacturer’s instructions. Subsequently 2 µl of the product was used for detecting specific miRNAs by quantitative PCR using TagMan microRNA assay kits (ABI, Foster City, CA, USA) for the corresponding miRNA. All samples were analysed in duplicate.
Assessment of plasma phosphocreatine kinase activity
The activity of phosphocreatine kinase (CPK) activity in the plasma was assessed by a standard spectrophotometric method as described elsewhere. 24
Statistical analysis
Data are expressed as mean ± SEM. Intragroup comparisons were performed with Student’s t-test or repeated measures ANOVA followed by Tukey post test, where appropriate. A probability value of <0.05 was considered statistically significant.
Results
Characteristics of individuals included into the different studies
Baseline characteristics of individuals included into the different studies
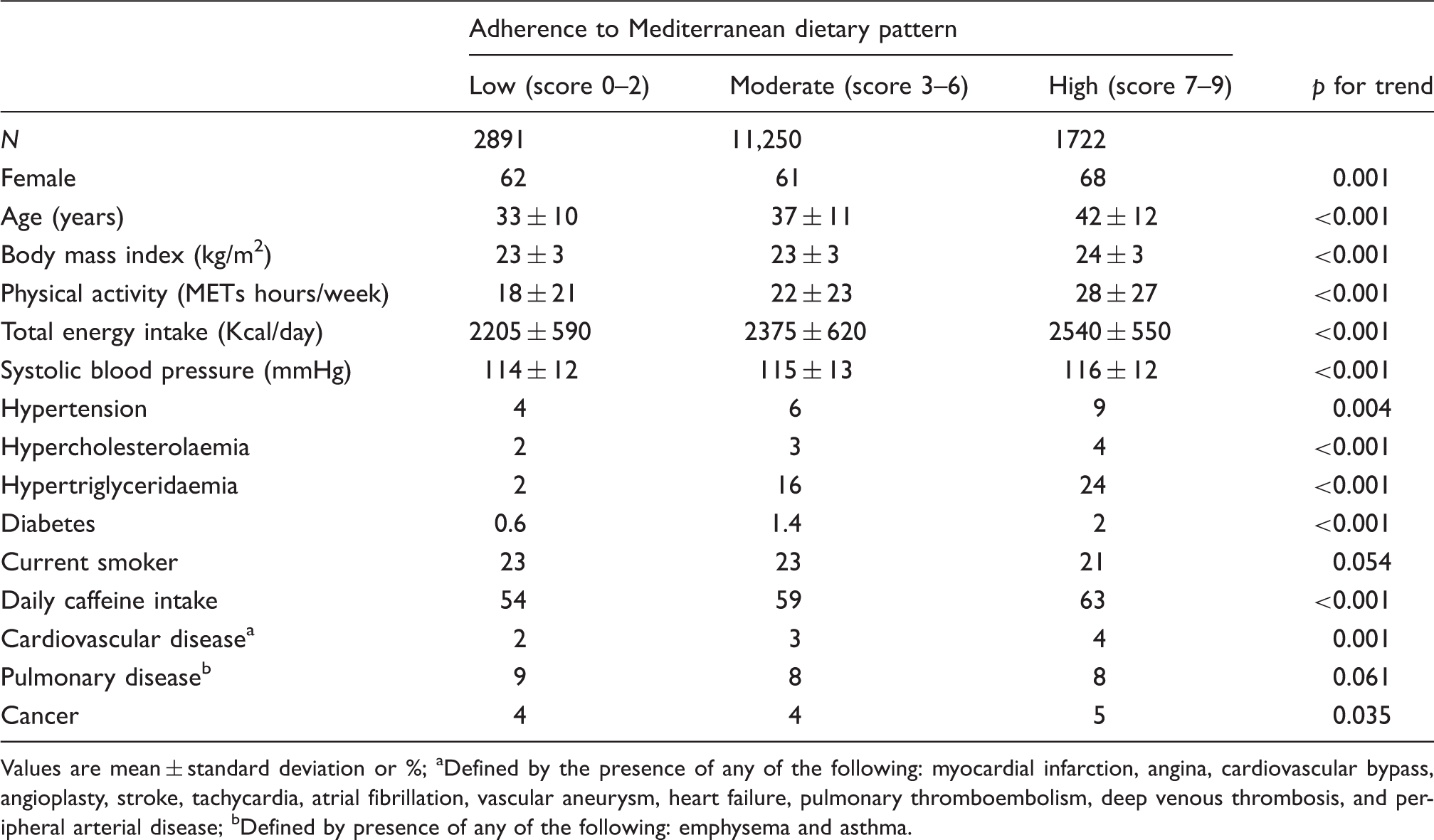
IAT: individual anaerobic threshold; MET: standard metabolic equivalent.
Impact of a single maximal exercise test on circulating miRNA-126 and miRNA-133
In healthy subjects performing a single symptom-limited exercise test, the circulating miRNA-126 level increased significantly at the highest work load compared with baseline measurement (2.1 ± 0.3-fold versus begin, p < 0.05) (Figure 1). This difference was present even 5 min after finishing the test (2.0 ± 0.2-fold vs. begin, p < 0.05). In contrast, miRNA-133 levels and CPK activity did not change significantly during and 5 min after the maximal exercise test.
Quantification of circulating miRNA-126, miRNA-133 and CPK activity in healthy individuals during a maximal exercise test. Plasma concentration of miRNAs and CPK activity was evaluated at begin, at 50% of the maximal performance (50% max.), at the ventilatory threshold (VT), at maximum (Max.) and 5 min after the test in the recovery phase. Values are depicted as mean ± SEM as relative increase versus begin. *p < 0.05 vs. begin.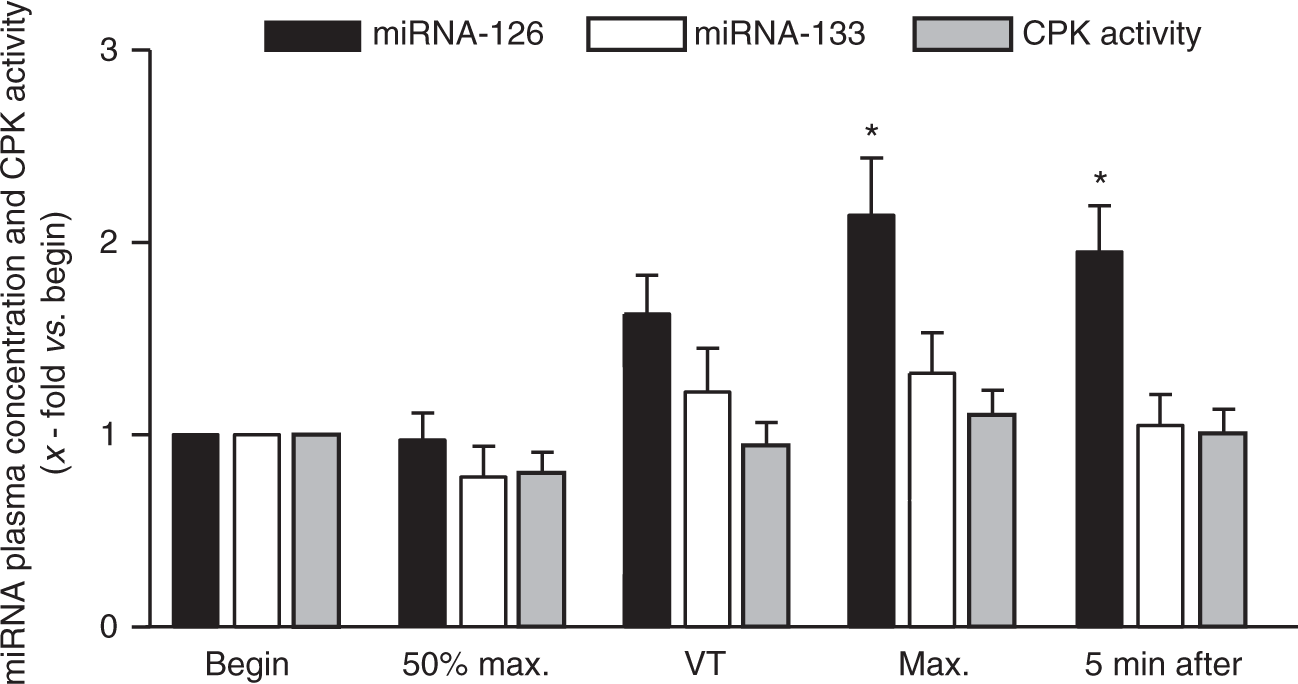
Impact of bicycling for 4 h on circulating miRNA-126 and miRNA-133
In healthy sportive individuals undergoing a bicycle race for 4 h at 70% of the individual anaerobic threshold, the plasma concentration of miRNA-126 significantly increased 30 min after the start (4.6 ± 1.1-fold vs. begin, p < 0.05) and remained elevated until the end of the test (4.0 ± 0.8-fold vs. begin, p < 0.05) (Figure 2). Plasma concentration of miRNA-133 and CPK activity did not display a bicycle-dependent pattern (p = not significant). Measurement of miRNA-126 and -133 as well CPK activity in the recovery period (1 and 24 h after finishing the exercise intervention), the serum concentrations/activity did not differ significantly compared with baseline measurements.
Quantification of circulating miRNA-126 and miRNA-133 as well as CPK activity in healthy individuals during a 4-h cycling test at 70% of the individual anaerobic threshold. Plasma samples were analysed at different time points during the test and at 1 h and 24 h after the test. Values are depicted as mean ± SEM as relative increase vs. begin. *p < 0.05 vs. begin.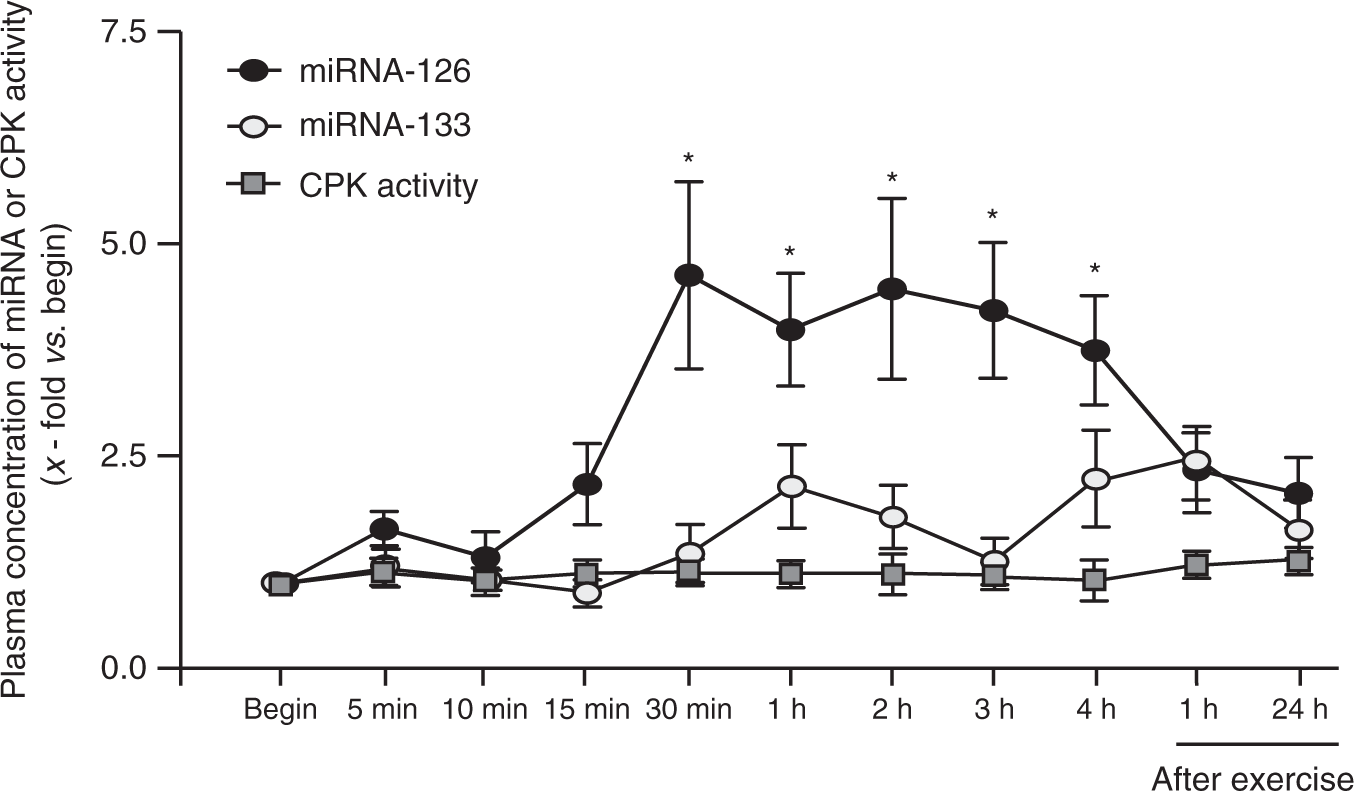
Impact of a marathon race on circulating miRNA-126 and miRNA-133
In the participants of a marathon race, the plasma concentration of miRNA-126 and miRNA-133 were significantly increased immediately after finishing the marathon race (miRNA-126: 3.42 ± 0.48-fold vs. begin; miRNA-133: 8.9 ± 2.63-fold vs. begin; p < 0.001; Figure 3). In addition, measurement of CPK activity in the plasma revealed a significant increase immediately after the race (3.78 ± 0.31 fold vs. begin).
Quantification of circulating miRNA-126 and miRNA-133 (a) as well as CPK activity (b) in healthy individuals before and immediately after performing a marathon race. Values are depicted as mean ± SEM as relative increase vs. begin. *p < 0.05 vs. begin.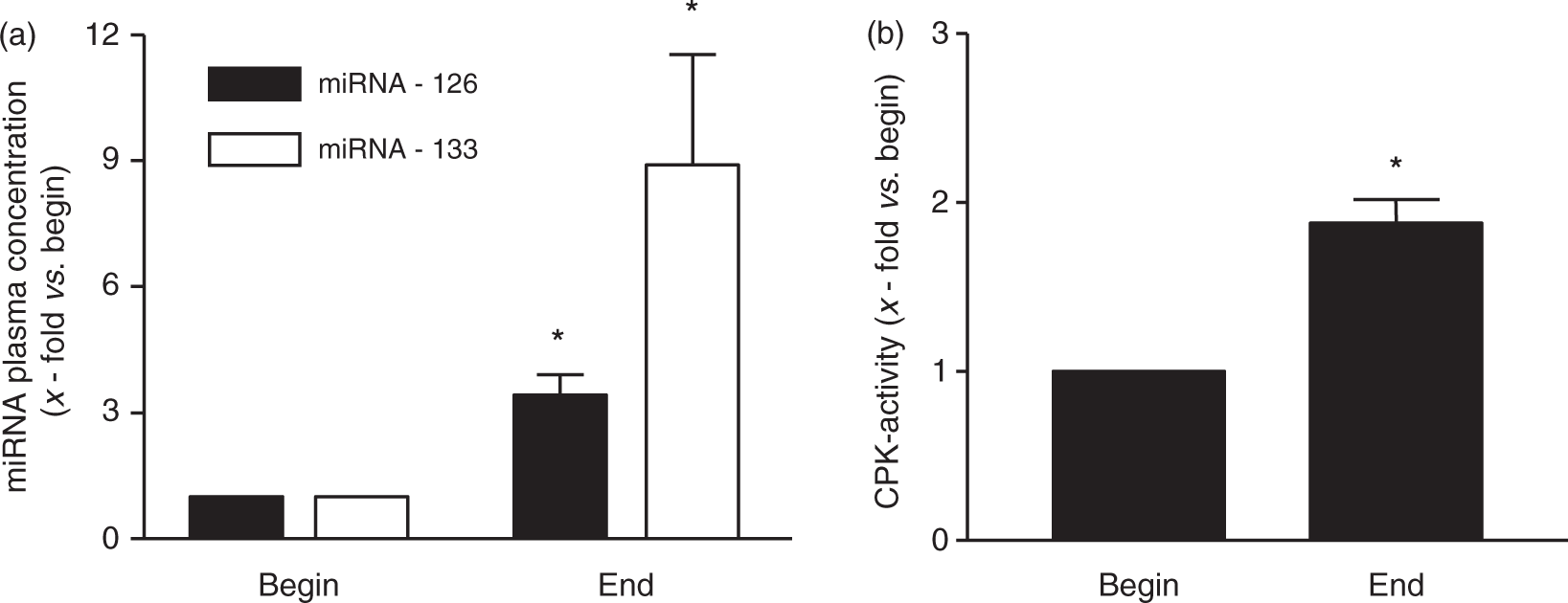
Impact of singular resistance training with additional eccentric loads on circulating miRNA-126 and miRNA-133
Interestingly, in healthy subjects performing an isolated resistance training with additional eccentric workloads, we did not find a change in circulating miRNA-126 level, whereas the serum concentration of miRNA-133 was significantly increased immediately after the training session (2.1 ± 0.4-fold vs. begin, p < 0.05) (Figure 4). At 1 h after finishing the test, the concentration of circulating miRNA-133 was back to baseline values. The increase of miRNA-133 goes along with a significant increase in CPK activity in the plasma after the training sessions (pre: 7.83 ± 0.6 arb. unit/ml; post: 10.47 ± 1.27 arb. unit/ml; 1 h post: 7.31 ± 1.05 arb. unit/ml).
Quantification of circulating miRNA-133 (a) and miRNA-126 (b) as well as CPK activity (b) in healthy individuals before, immediately after and 1 h after performing a singular resistance exercise programme with additional eccentric loads. Values are depicted as mean ± SEM as relative increase vs. begin.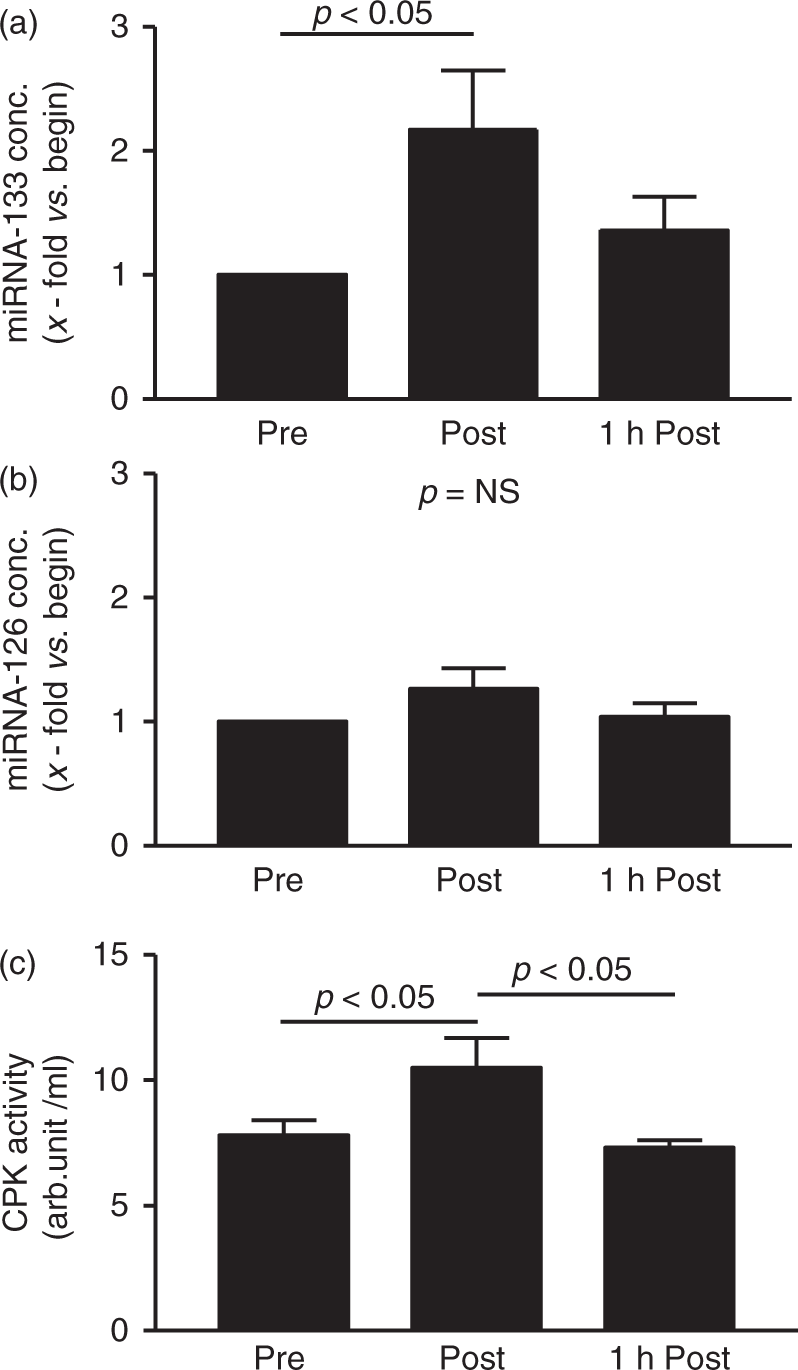
Discussion
In measuring plasma concentrations of circulating miRNAs (miRNA-126 as marker for endothelial cell damage and miRNA-133 as marker for muscular damage) in relation to four different exercise regimes in healthy individuals, three important messages emerge.
First, in line with previous reports,25,26 our data show that circulating miRNA-126 (endothelial marker) and miRNA-133 (muscular marker) can be reliably measured in human blood plasma. Second, without influencing miRNA-133 plasma concentrations, a maximal exercise test as well as 4 h of bicycling below the anaerobic threshold clearly demonstrated an increase of circulating miRNA-126 in healthy subjects. Third, running a marathon race led to a significant increase of both miRNA-126 and miRNA-133. Fourth, in contrast, a singular resistance exercise with additional eccentric loads resulted in an increase of miRNA-133 and is not associated with a change of circulating miRNA-126 level.
Based on these results one may conclude that either performing a single spiroergometry test or cycling 4 h below the anaerobic threshold results in damage to the endothelial cell layer, without damaging the musculature. On the other hand, running a marathon results in damage to the endothelial cell layer as well as to the musculature, whereas a resistance training influences only the musculature and not the endothelium.
Micro-RNA as marker for cell damage
Up to now several studies investigating different diseases could clearly demonstrate that miRNAs are detectable in the plasma.22,26–29 Since the blood contains high levels of RNase activity that will degrade exogenous added RNA, 30 we have to assume that the miRNAs are not present in a free, unprotected state. Numerous studies described an association between circulating miRNA and secreted membrane vesicles such as microvesicles, exosomes or apoptotic and senescent bodies.31–34 Nevertheless, the inclusion of circulating miRNA into vesicles has still to be questioned. In cell culture experiments Wang and colleagues studied the export of miRNAs in different cell types. 35 They could clearly demonstrate that exported miRNAs are found both within and without cell derived vesicles, and that nucleophosmin 1 seems to be a protein protecting vesicle-free miRNA from degradation.
The rapid appearance (within 1–4 h after tissue injury) of miRNAs in the circulation 27 is not completely understood, but they may derive from lysed or necrotic cells. In a myocardial injury model, rats were injected with isoproterenol and the presence of miRNA-208 in the circulation was as rapid as cTnI – what is regarded as the gold standard for diagnosing myocardial infarction. 36 In the discussion the authors argue that these results clearly support the hypothesis that miRNAs leak out of injured cells. This hypothesis of miRNA leakage from injured cells is furthermore supported by observations in acute myocardial infarction models26,27 as well as in drug-induced liver injury. 28
Based on these considerations, it is reasonable to consider the rise in circulating miRNAs as an indirect marker of cell damage.
Endothelial damage elicited by exercise
Despite the observation that at least strenuous exercise like running a marathon affects endothelial function, 37 the exact mechanism remains speculative. Whether a damage/lysis of endothelial cells is one of the possible mechanisms remains uncertain. Using either endothelial derived microparticles 18 or mature circulating endothelial cells 19 the results are inconclusive. Based on the results described in the present study, where the circulating concentration of miRNA-126 was used as marker for endothelial cell destruction, a damage to endothelial cells occurs even in the case of a single maximal exercise test. In the case of longer exercise tests being performed, such as bicycling for 4 h below the individual anaerobic threshold or running a marathon, a clear increase of miRNA-126 in the plasma was evident, arguing for the lysis of endothelial cells. In the case of an exercise test of just resistance exercise being performed, no injury of endothelial cells, using plasma miRNA-126 as marker, was noted. The significant rise of circulating miRNA-126 was already detectable after 30 min of bicycling (Figure 2). This points to an early disruption of endothelial cells elicited by exercise. As mentioned above this early increase is also seen in patients after a myocardial infarction, where an elevation of circulating miRNA-208a became detectable within 1 h, whereas cardiac troponin I was still negative. 27
Is miRNA-126 a specific marker for endothelial cells? Expression studies using a real-time PCR approach clearly documented that miRNA-126 is highly expressed in microvascular and macrovascular endothelial cells. 38 This specificity for endothelial cells was furthermore confirmed by in-situ hybridization on human renal sections. 38 In addition miRNA-126 knockout animals displayed severely delayed retinal development 39 and targeted deletion of miRNA-126 is reported to result in vascular leakage, progressive haemorrhages and defects in endothelial cell proliferation.40,41
Exercise and muscle damage measured by miRNA-133
In contrast to miRNA-126, miRNA-133 is highly expressed in skeletal and cardiac muscle.42,43 At least in skeletal muscle miRNA-133 is required for proper muscle development, and regulation of proliferation and differentiation by repression of serum response factors. 42 In recent reports circulating levels of miRNA-133 were used as a new biomarker for acute myocardial infarction. 27 In the present study a significant rise of miRNA-133 in the circulation was seen after a marathon run and after resistance training with additional eccentric loads. No elevation was detected in less strenuous forms of exercise such as a single maximal exercise test or cycling for 4 h at 70% of the individual aerobic threshold. These data are in accordance with the measurement of CPK activity, which has been used to detect cell damage after a marathon race. 44 In addition electronmicroscopic images from muscle biopsies taken shortly after a marathon run showed myofibrillar lysis, dilatation and disruption of the T-tubular system as well as extracellular oedema with endothelial injury. 45
In conclusion, the data presented in this exercise study investigating the impact of different exercise modalities on the concentration of circulating miRNA-126 and miRNA-133 support the notion that exercise-induced damage to the endothelial cell layer as well as to muscular tissue is evident depending on the exercise format. Very strenuous exercise like running a marathon has an impact on endothelial cells and muscular cells, whereas less stringent forms affect only the endothelium or the muscle.
Study limitations
Several limitations of the present study should be mentioned.
First, with the data presented in the manuscript we are not able to discriminate between different mechanisms leading to an increase in circulating miRNA-126. Nevertheless, the validity for using miRNA-133 as a marker for cell damage is confirmed by the measurement of CPK activity, an established marker for cell damage. Unfortunately, no such easily assessable specific maker exists for endothelial cells. Second, the sampling of the plasma after finishing the exercise is different in the various exercise groups, ranging from 5 min to 24 h. Therefore, an exact timing for the recovery is not possible for the different interventions, especially with the knowledge that already 5 min after the maximal exercise test the values are back to normal. Third, a possible gender specific effect cannot be ruled out, since the groups are too small. Therefore, further studies have to clarify whether a gender effect with respect to an exercise-induced endothelial damage can be observed.
Footnotes
Acknowledgements
The authors would like to thank Angela Kricke for her excellent technical assistance.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Conflict of interest
None declared.