
Abstract
Antimicrobial resistance (AMR) is a critical global health challenge, as it reduces the effectiveness of current therapies and demands novel integrative approaches. This narrative, integrative review analyzes how physical activity and nutrition interact with the pharmacological design of antimicrobial agents, influencing their absorption, metabolism, distribution, efficacy, and resistance development, drawing on studies published between 2015 and 2025 across microbiology, pharmacology, nutrition, and exercise physiology. Available evidence indicates that physical activity enhances immune competence, modifies pharmacokinetics, and promotes microbiome diversity, whereas nutrition influences bioavailability, micronutrient support, and nutrient–drug interactions. Conditions such as obesity, malnutrition, and metabolic disorders can critically alter drug disposition and therapeutic outcomes. Nutraceuticals and functional foods may act synergistically with antimicrobials, although antagonistic effects can impair their absorption or potency, and lifestyle-driven modulation of the microbiota and host metabolism appears to play an important role in resistance pathways. Emerging strategies, including prodrugs, nanocarriers, and personalized dosing algorithms, have the potential to optimize therapy according to lifestyle and metabolic profiles. Overall, incorporating lifestyle determinants into antimicrobial research and stewardship may improve therapeutic efficacy, reduce resistance, support precision medicine, and position diet and physical activity as key modulators of infection management.
Introduction
Antimicrobial resistance (AMR) has emerged as one of the most formidable public health challenges of the 21st century, with an estimated 4.95 million deaths associated with drug-resistant infections in 2019 alone.
1
The global proliferation of multidrug-resistant bacteria has significantly compromised the efficacy of existing antimicrobial therapies, threatening the successful treatment of common infections and surgical procedures.
2
Although advances in medicinal chemistry and microbial genomics have driven efforts to develop next-generation antibiotics, these strategies have not kept pace with the rapid evolution of resistance mechanisms.
3
Notably, most current approaches to antimicrobial stewardship and drug design focus on pathogen-targeted interventions, often neglecting the broader host-related and environmental determinants that modulate drug efficacy and resistance development.
4
Increasing evidence suggests that lifestyle factors, particularly diet and physical activity, play pivotal roles in shaping the host immune response, influencing gut microbial ecology, and modulating the pharmacokinetics and pharmacodynamics of antimicrobial agents (Figure 1).5,6 Interplay between lifestyle factors (physical activity, nutrition) and antimicrobial drug design in resistance management.
Despite their biological relevance, these extrinsic modulators remain largely overlooked in conventional antimicrobial research, highlighting a critical gap in our understanding of how behavioral and physiological variables affect therapeutic success and resistance trajectories.5,7
Emerging research has increasingly highlighted the link between host lifestyle factors and antimicrobial efficacy, particularly through their impact on the gut microbiome, immune regulation, and systemic metabolism (Figure 1). Regular physical activity has been shown to enhance mucosal immunity and microbial diversity, which may influence host resistance to infections and responsiveness to antibiotic treatment. 8 Exercise-induced shifts in microbial composition are associated with increased levels of beneficial taxa, for instance, in an experimental animal model, Faecalibacterium and Akkermansia, and elevated production of short-chain fatty acids, both of which play a role in immune modulation and pathogen defense.8,9 Concurrently, dietary patterns have demonstrated a profound effect on microbial ecology and metabolic pathways, with diets rich in fiber and polyphenols supporting microbial homeostasis and anti-inflammatory profiles. Western diets high in fat and sugar have been associated with dysbiosis and systemic inflammation, 10 which may influence microbial composition and functional potential. This microbial configuration has been linked to changes in gene expression and the relative abundance of resistance-associated taxa, although direct causal pathways remain incompletely understood. In this context, Western dietary patterns may contribute to a microbiome environment that could favor resistance-related processes, although evidence remains heterogeneous and further studies are required to establish causality. 11 These changes are relevant not only to infection risk but also to drug metabolism: alterations in microbial enzymatic activity can influence the biotransformation and bioavailability of orally administered antimicrobial agents, thereby affecting therapeutic outcomes. Moreover, nutritional deficiencies such as iron, zinc, and vitamin A depletion have been associated with impaired immune responses and altered pharmacokinetics of various antimicrobial agents.12–14 Together, these findings suggest that diet and exercise modulate host‒pathogen dynamics and antibiotic function through intertwined physiological and microbial mechanisms. 15
Despite growing evidence linking lifestyle factors with immune regulation, microbiome composition, and drug metabolism, current antimicrobial research has largely overlooked the potential consequences of these interactions on therapeutic efficacy and resistance evolution. Most pharmacological studies are conducted under standardized metabolic conditions, frequently neglecting interindividual variability related to physical fitness, nutritional status, or microbiome diversity. 16 This gap is particularly relevant given that sedentary behavior, poor dietary habits, and metabolic disorders such as obesity or malnutrition are widespread in both clinical and general populations—conditions that can significantly alter drug pharmacokinetics and host susceptibility to infection. 17 Furthermore, the design and testing of antimicrobial agents have not yet incorporated metabolic phenotypes as a variable of interest, nor have lifestyle-related factors been systematically considered in the development of therapeutic guidelines or dosing regimens.18,19 As such, a multidisciplinary framework that integrates insights from microbiology, pharmacology, nutrition, and exercise physiology to better understand how behavioral and physiological variables influence antimicrobial efficacy and resistance mechanisms is urgently needed.
This review addresses a critical gap by examining how physical activity and nutrition interact with antimicrobial pharmacology, with particular emphasis on pharmacokinetic and pharmacodynamic processes and their implications for drug design and therapeutic optimization.
Given the growing burden of antimicrobial resistance, this work focuses on how lifestyle-related factors influence drug absorption, metabolism, distribution, efficacy, and resistance development. It further explores how associated physiological and metabolic variations—such as changes in body composition, tissue perfusion, pH, and enzymatic activity—may inform antimicrobial drug design at the molecular level. By integrating evidence from microbiology, pharmacokinetics, immunology, and lifestyle sciences, this review aims to identify key mechanisms linking modifiable behaviors with antimicrobial performance and to support more precise and context-sensitive therapeutic strategies.
Ultimately, this integrative perspective may contribute to more personalized, context-sensitive therapeutic approaches that enhance treatment efficacy and reduce the emergence of resistance, aligning with the principles of precision medicine and translational pharmacology. 19
Methods
This review was conducted via a narrative, integrative approach to synthesize current scientific literature exploring the interactions between physical activity, nutrition, and the pharmacological design and efficacy of antimicrobial agents following previous authors. This narrative approach allows for the integration of evidence from diverse disciplines, including microbiology, pharmacology, nutrition, and exercise physiology, facilitating the synthesis of complex interactions between lifestyle factors and antimicrobial pharmacology.20,21 Given the multifactorial nature of antimicrobial resistance and its modulation by lifestyle variables such as diet, physical activity, and metabolic status, a narrative synthesis allows for a more integrative and interpretive approach than traditional systematic reviews. This method enables the contextualization of emerging mechanisms, theoretical models, and translational implications that may not yet be captured in homogeneous or quantitatively comparable datasets.22,23
A comprehensive literature search was conducted between January 2015 and May 2025 using the electronic databases PubMed, Scopus, and Web of Science, aiming to capture recent advances in the interdisciplinary fields of antimicrobial pharmacology, nutrition, and exercise science. However, it is important to note that earlier foundational studies, particularly those establishing the effects of physical activity on renal blood flow, hepatic metabolism, and drug pharmacokinetics, were considered for conceptual background and interpretation.The search strategy combined controlled vocabulary (MeSH terms) and free-text keywords related to antimicrobial resistance, drug efficacy, physical activity, nutrition, microbiome, pharmacokinetics, nutraceuticals, and personalized medicine, using Boolean operators (AND, OR) to optimize retrieval across disciplines.
The inclusion criteria were: (1) peer-reviewed articles published in English; (2) studies involving human or relevant animal models; and (3) studies explicitly addressing the interaction between lifestyle factors (physical activity and/or nutrition) and antimicrobial pharmacokinetics, pharmacodynamics, immune response, microbiome–drug interactions, or resistance-related outcomes. Exclusion criteria comprised non-English publications, review articles (used only for background), conference abstracts without full-text availability, and studies not directly examining the interaction between lifestyle factors and antimicrobial-related outcomes.
A total of 274 records were initially identified. After removing 83 duplicates and 8 records for other reasons (e.g., conference abstracts without full text), 183 records were screened by title and abstract, of which 90 were excluded due to lack of relevance. A total of 93 reports were sought for retrieval, and 7 could not be accessed. Consequently, 86 full-text articles were assessed for eligibility. Of these, 52 were excluded for the following reasons: lack of direct relevance to the interaction between lifestyle factors and antimicrobial pharmacokinetics, pharmacodynamics, or resistance-related outcomes (n = 25); review articles used only as background (n = 15); and insufficient data on pharmacokinetics, pharmacodynamics, or resistance mechanisms (n = 12). Finally, 34 studies were included in the review. The study selection process is summarized in Figure 2. It should be emphasized that this article is a narrative (integrative) review and not a systematic review. A PRISMA-style flow diagram has been included solely to enhance the transparency and reproducibility of the literature-screening process and does not imply adherence to the full methodological requirements of a systematic review (such as a registered protocol, formal risk-of-bias assessment, or quantitative synthesis). PRISMA-style flow diagram of the study selection process used to document literature screening for this narrative review (the diagram is presented for transparency and does not denote a systematic review).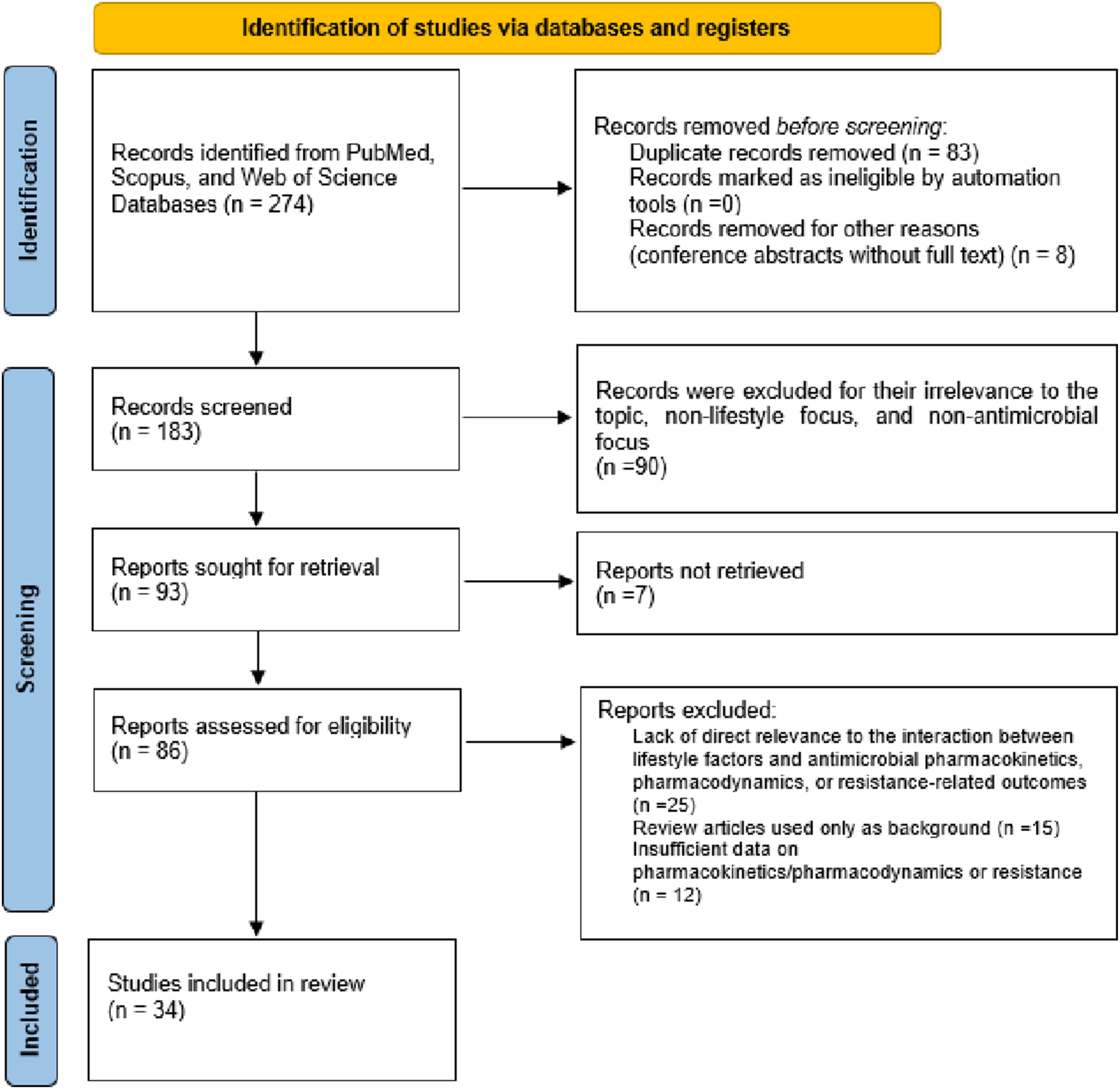
Data extraction was performed manually by the lead author, and studies were thematically grouped into the following categories: (1) modulation of the microbiome by lifestyle factors; (2) nutrition and drug metabolism; (3) exercise and immune response; (4) drug–lifestyle interactions in resistance development; and (5) personalized antimicrobial pharmacology. The relatively limited number of included studies reflects the emerging and highly specific nature of research at the intersection of physical activity, nutrition, and antimicrobial pharmacology, as well as the intentional application of restrictive inclusion criteria to ensure conceptual coherence and relevance.
Given the narrative and integrative design of this review, a formal standardized quality appraisal tool was not applied. Instead, studies were evaluated based on their methodological clarity, relevance to the research question, and contribution to the interdisciplinary framework. Particular consideration was given to study design (clinical, animal, or in vitro), consistency of findings, and the extent to which studies directly addressed interactions between lifestyle factors and antimicrobial pharmacology. When synthesizing the evidence, conflicting findings were interpreted in light of differences in study design, population characteristics, and methodological approaches, allowing for a contextual and critical integration of the available literature.
Discussion
Conceptual and evidence-based integration of lifestyle–drug interactions
Class-specific pharmacokinetics and resistance mechanisms of antimicrobial agents
Antimicrobial agents encompass pharmacologically distinct classes, including antibiotics, antifungals, and antivirals, each characterized by fundamentally different pharmacokinetic profiles, mechanisms of action, and resistance pathways. Antibiotics primarily target bacterial structures or processes (e.g., cell wall synthesis, protein synthesis) and often exhibit rapid distribution and elimination, with many hydrophilic agents relying on renal clearance. In contrast, antifungal drugs, such as azoles and echinocandins, typically act on eukaryotic cell components (e.g., ergosterol synthesis or cell membrane integrity) and frequently undergo extensive hepatic metabolism, resulting in different susceptibility to host metabolic and nutritional states. Antiviral agents, on the other hand, often require intracellular activation and are highly dependent on host cellular machinery, with pharmacokinetics influenced by factors such as cellular uptake, enzymatic activation, and tissue-specific distribution. Moreover, resistance mechanisms vary considerably across these classes, ranging from enzymatic degradation and efflux pumps in bacteria to target mutations in fungi and rapid genomic variability in viruses. These differences are critical when considering the influence of lifestyle factors, as exercise, nutrition, and metabolic status may differentially affect drug absorption, distribution, metabolism, and efficacy depending on the antimicrobial class. Therefore, lifestyle–drug interactions should be interpreted within a class-specific pharmacological framework rather than under a generalized concept of “antimicrobials”.1–4 It should be noted that, given the comparatively larger body of evidence available for antibacterial agents, the present review focuses predominantly on antibiotics, whereas the influence of lifestyle factors on antifungal and antiviral pharmacology is addressed more briefly and should be regarded as an area requiring dedicated future investigation.
Impact of lifestyle factors on antimicrobial resistance
AMR represents a growing global health threat, driven not only by antibiotic misuse but also by lifestyle-related factors that modulate host–microbe interactions and influence microbial evolution. Among these factors, physical inactivity, poor dietary patterns, chronic stress, and metabolic dysregulation play underappreciated roles in shaping microbial ecology, immune resilience, and the horizontal transfer of antibiotic resistance genes (ARGs). These behaviors modify both the human microbiome and systemic physiological states in ways that may compromise drug efficacy and increase microbial virulence.
Sedentary behavior has been associated with systemic inflammation, insulin resistance, and gut dysbiosis, all of which create a permissive environment for the selection of resistant bacterial strains. Physical inactivity has been reported to contribute to reduced microbial diversity, and some studies describe a relative decrease in beneficial genera such as Faecalibacterium prausnitzii and Akkermansia muciniphila, which have anti-inflammatory and colonization-resistant properties. 22 Low physical activity also impairs gut epithelial barrier function, increasing lipopolysaccharide (LPS) translocation and systemic endotoxemia. This persistent immune activation fosters immune exhaustion and disrupts mucosal immunity—conditions that favor chronic infections, increase antibiotic use, and potentially accelerate AMR propagation. 23
Modern dietary habits, characterized by high intake of saturated fats, refined carbohydrates, and ultra-processed foods, profoundly shape the gut microbiome and its resistome. Evidence suggests that diets rich in animal protein and fat may promote the expansion of Bacteroides and Proteobacteria, taxa that have been associated with a higher prevalence of antimicrobial resistance genes (ARGs). 24 Moreover, these dietary patterns reduce short-chain fatty acid (SCFA) production—particularly that of butyrate and acetate—by diminishing the number of fiber-fermenting bacteria. SCFAs are essential for mucosal immunity and limiting pathogen growth. It has been proposed that their deficiency may be associated with increased gut pH and impaired immune exclusion, conditions that could favour the persistence of opportunistic pathogens and resistance gene carriers, although this remains a largely hypothetical pathway requiring further confirmation. 25 Some processed foods also contain trace amounts of antibiotics and antibiotic-like preservatives that exert subtherapeutic antimicrobial pressure, further selecting for resistant organisms. 24 In this sense, it has been hypothesized that the Western diet might act as a selective pressure analogous to low-dose antibiotic exposure, although direct causal evidence in humans remains limited.
Obesity and metabolic syndrome, which are largely driven by poor diet and inactivity, have been proposed as indirect but potent enhancers of AMR. These conditions are linked to increased infection susceptibility, delayed immune responses, and impaired vaccine efficacy. Observational data suggest that individuals with metabolic dysfunction may carry a higher burden of multidrug-resistant enterobacteria, although such associations are correlational and do not establish a direct causal relationship. 26 Mechanistically, and largely on the basis of preclinical and associative evidence, metabolic dysfunction affects bile acid metabolism, alters microbial signaling, and reduces antimicrobial peptide production. Furthermore, obesity promotes chronic low-grade inflammation (“metaflammation”), which not only compromises mucosal defenses but also facilitates biofilm formation—a critical factor in antibiotic evasion.
Chronic stress, poor sleep, and psychological dysregulation influence the hypothalamic‒pituitary‒adrenal (HPA) axis and the autonomic nervous system, which in turn modulate mucosal immunity and the gut microbial structure. These alterations may indirectly affect AMR by increasing colonization by Enterobacteriaceae and Clostridioides difficile, reducing the production of immunoglobulin A (IgA) and promoting antibiotic use due to recurrent subclinical infections.
A key mechanism potentially linking lifestyle factors and antimicrobial resistance is the modulation of horizontal gene transfer (HGT) within the gut microbiome.27,28 Dysbiotic microbial communities have been shown—primarily in in vitro studies and animal models—to harbor increased densities of mobile genetic elements such as plasmids, integrons, and transposons, which facilitate the dissemination of antibiotic resistance genes (ARGs). 27 Lifestyle-induced alterations in microbial composition may increase the relative abundance and proximity of commensal and opportunistic bacteria, creating conditions that could favor gene exchange. However, it is important to note that most of the mechanistic evidence supporting these processes derives from controlled experimental models,29,30 and direct clinical evidence in humans remains limited.29,30 Therefore, while these findings suggest a plausible link between lifestyle-modulated microbiota and HGT dynamics, further studies are required to establish their clinical relevance. 29
Despite these limitations, emerging evidence indicates that lifestyle interventions may modulate the microbiome in ways that are associated with reduced ARG abundance and improved microbial resilience. Moderate physical activity, plant-rich diets, and stress management protocols have been linked to the reversal of dysbiosis and a lower prevalence of resistance-associated taxa. Similarly, the intake of fermented foods rich in bacteriocin-producing lactobacilli may competitively exclude ARG carriers and restore colonization resistance. 28 These findings highlight the potential role of preventive lifestyle strategies within the antimicrobial stewardship continuum, although their direct impact on HGT dynamics in humans remains to be fully elucidated.
Lifestyle factors, including physical activity, dietary composition, stress levels, and metabolic health, shape both the structure and functional resistome of the human microbiota. Through effects on microbial ecology, immune function, and gene transfer mechanisms, these variables play decisive roles in AMR evolution. While antibiotic misuse remains a central driver, addressing behavioral and metabolic contributors offers a promising, underutilized strategy for curbing resistance. Future clinical frameworks may incorporate lifestyle screening into AMR risk stratification, aligning antibiotic therapy with individualized metabolic and microbiome profiles. In the following section, we explore how diet and physical activity comodulate the microbiome in more detail, laying the foundation for integrated resistance management.
An often overlooked contributor to lifestyle-associated AMR is environmental exposure in urban settings. Urban dwellers are frequently exposed to environmental reservoirs of ARGs, including wastewater, contaminated surfaces, and air particulates in public transport and healthcare settings. Physical inactivity and indoor-dominant lifestyles reduce natural immunological priming and the diversity of environmental microbiota exposure, contributing to immune dysregulation and a less competitive microbiota that cannot efficiently prevent colonization by resistant strains. 31 Furthermore, individuals with limited outdoor activity may be more frequently exposed to indoor antimicrobial cleaning agents, which exert selective pressure on the skin and gut microbiota. Over time, this may promote the enrichment of efflux pump-containing commensals or bacteria carrying multidrug resistance (MDR) plasmids.
Emerging research suggests that lifestyle factors may be associated with epigenetic modifications, including DNA methylation, histone acetylation, and miRNA expression, that could influence not only host immunity but also microbiome composition and ARG carriage. For example, sedentary lifestyles and ultraprocessed diets have been linked to hypermethylation of genes involved in mucosal defense and xenobiotic metabolism, which indirectly affects microbial colonization and resilience. 29 Additionally, host–microbe coregulation of gene expression means that epigenetic environments influenced by diet and physical activity can select for microbial populations that are either more cooperative (commensal-dominant) or more virulent/resistant (pathobiont-dominant). This relationship suggests that resistance risk is not only microbial but also host programmed and therefore potentially reversible through lifestyle modulation.
Recognizing lifestyle behaviors as modifiable risk factors for AMR paves the way for precision prevention strategies. Just as pharmacogenomics is used to tailor medications, a lifestyle-based resistance risk profile could inform prophylactic measures or antimicrobial prescribing practices. For example, patients with sedentary, dysmetabolic, or highly urbanized profiles may benefit from microbiome-preserving antibiotic selection, increased probiotic support, or closer resistance monitoring during therapy. Moreover, public health policies could integrate AMR considerations into urban planning, dietary guidelines, and exercise promotion campaigns. Such approaches acknowledge AMR not just as a clinical outcome of drug misuse but also as a systems-level reflection of human–environment–microbiota interactions.
Modulation of the human microbiome through diet and exercise
The human gut microbiome plays a critical role in maintaining immune homeostasis, metabolic balance, and host defense. Both dietary composition and physical activity are recognized as primary modulators of microbial composition, diversity, and function. These lifestyle factors shape not only microbial abundance but also the production of bioactive metabolites such as short-chain fatty acids (SCFAs), which directly influence inflammation, barrier integrity, and antimicrobial resistance (AMR). In this section, we explore the molecular pathways through which diet and exercise comodulate the microbiome, highlighting the implications for host–microbe interactions and therapeutic resistance.
Diet is one of the most potent and rapid influences on the composition of the gut microbiome. Diets high in fiber, polyphenols, and complex carbohydrates increase the relative abundance of Bacteroidetes, Lactobacilli, and Prevotella, increasing the production of beneficial SCFAs, such as butyrate, propionate, and acetate. These metabolites improve colonic barrier function, suppress inflammatory signaling, and enhance mucosal immunity, collectively reducing pathogen colonization and ARG dissemination. 32 Conversely, Western diets rich in fat and refined sugars favor microbial shifts toward Firmicutes and Proteobacteria, increasing gut permeability and low-grade endotoxemia. This microbial configuration is associated with enhanced quorum sensing, biofilm formation, and the upregulation of resistance genes in commensals such as Escherichia coli and Enterococcus faecalis. 33
Physical activity exerts a bidirectional influence on the microbiome, promoting diversity, enhancing SCFA synthesis, and enriching commensal species involved in immune regulation and anti-inflammatory pathways.34–36 Both acute and chronic exercise have been shown to increase the abundance of Akkermansia muciniphila, a mucin-degrading bacterium that maintains barrier integrity and reduces intestinal inflammation. 37 Moreover, endurance exercise has been shown to increase the abundance of butyrate-producing genera, such as Roseburia and Faecalibacterium, which act as critical suppressors of proinflammatory cytokine production. These microbes also indirectly reduce antibiotic resistance propagation by outcompeting ARG-harboring strains and maintaining ecological balance. 38
SCFAs are fermentation products of dietary fibers and act as post-biotic messengers between the microbiota and the host. Butyrate inhibits histone deacetylase (HDAC) activity, inducing anti-inflammatory gene expression and promoting Treg cell differentiation. 39 It also enhances the production of antimicrobial peptides (e.g., β-defensins) and tight junction proteins (occludin, claudin-1), which strengthen the intestinal barrier against pathogenic infiltration. Notably, SCFAs influence bacterial resistance by modulating the intracellular pH, interfering with efflux pump activity, and quenching virulence gene expression in enteropathogens. Thus, fiber-rich diets and exercise-derived SCFA profiles may indirectly reduce AMR risk beyond simply modulating immune responses.
Diet and exercise interact to shape the gut resistome—the collection of all resistance genes in the microbiota. For example, dietary restriction of antibiotics and processed additives reduces the selective pressure that is associated with ARG carriage. Concurrently, regular aerobic exercise enhances microbial gene richness and may contribute to horizontal gene transfer suppression by enriching taxa that secrete quorum-sensing inhibitors. 35 In one clinical study, individuals adhering to a Mediterranean diet with moderate physical activity presented a 30% reduction in fecal ARGs over 12 weeks, alongside increased microbial diversity and SCFA levels. 36 These findings suggest that microbiome-targeted lifestyle interventions can reverse dysbiotic and resistogenic states in metabolically compromised individuals.
Baseline metabolic status significantly influences the responsiveness of the microbiota to lifestyle changes. Obese and diabetic individuals often exhibit blunted SCFA production, increased bile acid deconjugation, and colonization by resistance-prone pathobionts. Exercise and prebiotic intervention can partially restore these deficits, although the magnitude of response varies with age, sex, and epigenetic background. 40 Epigenetic interactions between SCFAs and host cells, such as butyrate-driven HDAC inhibition, highlight a host–microbe codependency that could be leveraged for therapeutic precision. These findings emphasize the potential for integrated lifestyle-microbiome therapeutics in future antimicrobial stewardship strategies.
The human microbiome is highly plastic and responds dynamically to dietary inputs and physical activity patterns. These lifestyle elements not only influence microbial ecology but also affect antimicrobial resistance potential via metabolite signaling, barrier regulation, and competition dynamics. Diet and exercise act synergistically to cultivate a microbial ecosystem that is more resilient, less susceptible to pathogenic colonization, and less prone to resistance propagation. Future antimicrobial strategies may include microbiome-responsiveness screening before antibiotic therapy, incorporation of SCFA-producing probiotic strains and targeted prebiotic–exercise protocols to support host–microbiome cooperation during infection or antibiotic treatment. Recognizing lifestyle as a modulatory axis in resistance evolution opens new frontiers for nonpharmacological resistance mitigation.
In addition to macronutrient composition, the inclusion of fermented foods such as yogurt, kefir, kimchi, and miso provides direct microbial supplementation and enhances gut ecosystem resilience. These foods deliver live lactic acid bacteria and bacteriocin-producing strains that suppress pathogens and outcompete ARG-harboring organisms. 28 Furthermore, the postbiotic metabolites of fermentation, such as lactic acid, conjugated linoleic acid (CLA), and bacteriocins, modulate the gut pH, mucin production, and immune signaling. In clinical studies, the regular intake of fermented foods has been linked to a lower fecal ARG burden and greater microbial diversity, particularly when fermented foods are combined with physical activity. 41 These findings highlight the promising role of symbiotic strategies (diet + live organisms) in resistance control.
Prebiotics, which are nondigestible food components such as inulin, fructooligosaccharides (FOSs), and resistant starches, selectively stimulate the growth of beneficial microbes. When coupled with physical activity, prebiotics support the enrichment of SCFA-producing taxa and inhibit colonization by proteobacteria, which are known carriers of mobile ARGs. Resistant starches (e.g., from cooked-cooled potatoes or underripe bananas) act as delayed-fermenting substrates, promoting SCFA release along the distal colon. This not only enhances mucosal immunity but also lowers the luminal pH, creating an inhospitable environment for resistance-prone pathogens. These dietary fibers synergize with exercise-induced microbial changes, enhancing systemic metabolic and immune responses.
Emerging evidence suggests that early-life exposure to diet and exercise (or their absence) has long-term effects on microbiome plasticity and resistance outcomes. For example, childhood sedentary behavior and ultra-processed diets impair microbiome maturation, resulting in reduced microbial diversity and increased ARG susceptibility in adulthood. 39 Conversely, physical activity and dietary fiber during developmental windows increase microbial adaptability, promote epithelial tolerance, and reduce long-term colonization by opportunistic pathogens. This developmental programming underscores the importance of lifestyle intervention across the lifespan, not merely during therapeutic windows.
The gut microbiome is a metabolically and immunologically active ecosystem that responds dynamically to lifestyle inputs. Diet and exercise not only remodel taxonomic diversity but also shift microbial function toward anti-inflammatory, barrier-stabilizing, and resistance-suppressing profiles. Strategies incorporating fiber-rich diets, fermented foods, prebiotics, and regular physical activity represent powerful, nonpharmacologic tools for shaping a microbiome that is less permissive to resistance proliferation. Translationally, future approaches may include lifestyle-based AMR risk screening, coadministration of SCFA-boosting dietary plans with antibiotic regimens, and personalized symbiotic protocols tailored to the baseline microbiota and metabolic status. These strategies will be particularly relevant in high-risk populations, such as elderly individuals, immunocompromised individuals, or individuals with recurrent infections, where microbiome-targeted therapy may enhance both drug efficacy and resistance prevention.
Nutritional status and its role in drug absorption, metabolism, and efficacy
Nutritional status is a critical determinant of the pharmacokinetics and pharmacodynamics of antimicrobial drugs. Nutrient deficiencies or excesses can alter the physiological environment in which drugs are absorbed, distributed, metabolized, and excreted.42,43 For example, malnutrition may compromise the integrity of the gastrointestinal tract, reduce plasma protein levels, and impair hepatic and renal function, all of which are essential for proper drug metabolism and clearance. 44 Specifically, according to Koziolek et al. (2019), nutritional imbalances modulate hepatic enzyme activity and plasma protein levels, directly affecting the pharmacokinetics of various antimicrobial agents. 45 On the other hand, obesity is associated with increased adipose tissue and altered enzyme expression, which may lead to unpredictable drug distribution and efficacy. 46 These variations necessitate a more personalized approach to antimicrobial dosing on the basis of nutritional profiling.
Micronutrients as modulators of antimicrobial pharmacokinetics and the immune response
The micronutrient status also plays a pivotal role in modulating the enzymatic systems involved in drug metabolism. Vitamins such as B-complexes (e.g., B6, B12, and folate) are crucial cofactors in hepatic cytochrome P450 enzyme activity and govern the biotransformation of many antibiotics. 47 Deficiencies in these micronutrients can lead to suboptimal drug activation or prolonged half-life, thereby increasing the risk of toxicity or therapeutic failure. 47 Similarly, minerals such as zinc and magnesium are essential for maintaining cellular membrane integrity and drug transporter function, influencing the absorption of orally administered antimicrobials. Andrews (2001) discussed the role of zinc in regulating membrane transport proteins, highlighting its relevance in modulating the absorption and efficacy of orally administered drugs. 48
In addition to absorption and metabolism, nutrition affects drug efficacy through immunonutrition pathways. A well-nourished individual is more likely to mount a robust immune response, thereby enhancing the synergy between host defenses and antimicrobial therapy. 49 Conversely, undernourished patients may experience immunosuppression, reducing the clinical effectiveness of even potent antibiotics. This relationship highlights the importance of integrating nutritional support in infection management protocols, especially in vulnerable populations such as elderly individuals, hospitalized patients, and individuals with chronic diseases. 50 Calder (2013) noted that inadequate nutrition in elderly populations is closely linked to impaired immune responses, which can compromise the effectiveness of antimicrobial treatments. 51
For example, multiple micronutrients are known to modulate immune cell activity and cytokine production. Specifically, zinc deficiency impairs T-cell proliferation and natural killer cell activity, increasing susceptibility to infections and diminishing the immune system’s ability to work synergistically with antibiotics. 52 Similarly, selenium plays a crucial role in redox regulation and the function of neutrophils and macrophages, which are key in bacterial clearance.53,54 Vitamin D deficiency, which is highly prevalent in older adults and chronically ill patients, has also been associated with impaired innate immunity and increased risk of respiratory tract infections. 55
Clinical evidence supports the use of immunonutrition strategies—including supplementation with arginine, glutamine, omega-3 fatty acids, and nucleotides—to improve infection outcomes in surgical and intensive care unit (ICU) patients. These nutrients can enhance mucosal barrier integrity, modulate inflammatory responses, and support lymphocyte proliferation, thereby reinforcing the host response to antimicrobial agents. Despite promising results, their application must be individualized on the basis of patient condition, type of infection, and nutritional status. 56 Proactive assessment and correction of nutritional deficiencies could optimize immune function and, by extension, improve drug efficacy and clinical outcomes.
Nutrient–drug interactions and food-mediated bioavailability
Moreover, nutrient–drug interactions can either potentiate or inhibit antimicrobial action. For example, high-fat meals can increase the bioavailability of lipophilic drugs such as rifampin but may delay gastric emptying, affecting peak plasma concentrations. Yasuji et al. (2012) reported that high-fat meals can enhance the absorption of lipophilic antibiotics such as rifampin but also alter pharmacokinetic parameters such as the time to peak concentration due to delayed gastric emptying. 57 Moreover, calcium and iron supplements are known to chelate with tetracyclines and fluoroquinolones, reducing their absorption. Such interactions underscore the need for timing and dietary guidelines when prescribing antimicrobials, particularly in populations with high supplement usage.58,59
Furthermore, dietary fiber and certain plant-derived compounds can also bind to antimicrobial agents or modify the gut transit time, influencing drug dissolution and mucosal absorption.60,61 For example, soluble fibers such as pectin and guar gum can form viscous gels in the intestine that slow gastric emptying and impede the absorption of antibiotics such as amoxicillin or ciprofloxacin.62,63 Similarly, polyphenols found in foods such as green tea, grapes, and certain herbal supplements have been shown to interfere with drug-metabolizing enzymes and transporters, potentially reducing the bioavailability of various antimicrobial agents. 64 For example, Knop et al. reported that epigallocatechin gallate (EGCG), the major catechin in green tea, can inhibit key drug transporters and modify first-pass metabolism, particularly antibiotics with narrow therapeutic windows. 65
Finally, the timing and composition of meals can impact the dissolution environment in the stomach and duodenum, thereby affecting the solubility of pH-dependent drugs. Proton pump inhibitors (PPIs), often used alongside antimicrobials in hospitalized patients, further complicate absorption dynamics by increasing the gastric pH.66,67 These multifactorial influences underscore the need for individualized dietary guidelines accompanying antimicrobial prescriptions, especially in patients with complex nutritional profiles or polypharmacy.
Beyond their effects on drug absorption and bioavailability, nutritional factors may also contribute to antimicrobial resistance through indirect selective mechanisms. Diet-induced alterations in drug pharmacokinetics can lead to subtherapeutic antimicrobial concentrations at the site of infection, a well-established driver of resistance selection. For example, reduced absorption due to nutrient–drug interactions (e.g., chelation with minerals or delayed gastric emptying associated with high-fat meals) may result in drug levels that fall below the minimum inhibitory concentration (MIC), thereby allowing partially resistant bacterial populations to survive and proliferate. High-fat meals have been shown to significantly alter antimicrobial pharmacokinetics by modifying drug absorption and systemic exposure. 45 When such alterations lead to subinhibitory concentrations, they may promote adaptive responses such as increased expression of efflux pumps and stress-response pathways, ultimately favoring the selection of resistant mutants. 68 Therefore, nutritional status and dietary patterns may not only influence antimicrobial efficacy but also modulate the evolutionary pressure that is associated with resistance development.
Contraindications and antagonistic interactions between functional foods and antimicrobials
In addition to potential synergistic effects, certain foods and dietary components may exert clinically significant antagonistic interactions with antimicrobial agents, directly affecting their pharmacokinetics and therapeutic efficacy. 43
A well-established example is the interaction between tetracyclines or fluoroquinolones and calcium-rich foods or mineral supplements. Divalent cations such as calcium, magnesium, and iron can form insoluble complexes with these antibiotics in the gastrointestinal tract, significantly reducing their absorption and systemic bioavailability. This reduction may lead to subtherapeutic plasma concentrations, increasing the risk of treatment failure and promoting the selection of resistant bacterial strains. 63
Similarly, grapefruit juice and other dietary components can interfere with drug metabolism through inhibition of cytochrome P450 enzymes, particularly CYP3A4. 69 This interaction may alter antimicrobial exposure by increasing or unpredictably modifying drug plasma levels, potentially leading to toxicity or inconsistent therapeutic effects.46,63
These food–drug interactions highlight the importance of considering dietary intake when prescribing antimicrobial therapy. Failure to account for such interactions may compromise drug efficacy and contribute to antimicrobial resistance through inadequate exposure. Therefore, clinical guidance regarding meal composition and timing should be integrated into antimicrobial treatment strategies to ensure optimal pharmacological outcomes. 55
Clinical implications and perspectives
In clinical practice, the evaluation of nutritional status should be an integral part of antimicrobial stewardship programs. 70 Tools such as body mass index (BMI), serum ALB levels, and dietary assessments can aid in tailoring drug regimens. Moreover, emerging technologies in metabolomics and nutrigenomics offer promising avenues to predict individual responses to drugs on the basis of nutritional biomarkers. In summary, optimizing nutritional status is not only a supportive measure but also a strategic component in enhancing antimicrobial efficacy and minimizing resistance development. 71
Metabolomics and nutrigenomics offer promising opportunities for individualized treatment. By analyzing metabolic profiles or gene–nutrient interactions, clinicians can predict how a patient’s nutritional context may influence drug response. 72 For example, the identification of polymorphisms affecting vitamin B metabolism or cytochrome P450 activity could guide the selection and dosing of specific antibiotics. 73 Additionally, single nucleotide polymorphisms (SNPs) in genes such as CYP2C19 or CYP3A4 can result in altered metabolism of macrolides and azole antifungals, requiring adjusted dosing or alternative therapies.74,75 According to Berno et al. (2014), genetic polymorphisms in CYP3A4 and CYP3A5—enzymes responsible for metabolizing more than 60% of clinically used drugs—may explain interindividual variability in the pharmacokinetics of CYP-metabolized antimicrobials. 76 Genetic variants in folate transporters (SLC19A1) may also influence the efficacy of antifolate agents, especially in patients with underlying nutritional deficiencies. Furthermore, metabolomic signatures indicating deficiencies in essential amino acids or antioxidant status have been linked to poor responses to antimicrobial therapy, particularly in critical care settings.77,78
In parallel, antimicrobial stewardship programs should broaden their scope to incorporate nutritional monitoring and intervention as standard practices. Nutritional optimization through tailored diets, micronutrient repletion, or enteral/parenteral support could enhance drug efficacy and reduce resistance by ensuring proper therapeutic exposure and immunocompetence. 20
From a practical standpoint, the integration of lifestyle-informed strategies into antimicrobial therapy requires consideration of both patient-level and system-level factors. Routine clinical assessment could incorporate simple indicators such as physical activity level, dietary patterns, and metabolic status to guide antimicrobial selection and dosing. However, implementation may be constrained by variability in healthcare resources, time limitations, and access to specialized tools such as microbiome profiling or metabolomic analysis, particularly in resource-limited settings. Socioeconomic factors may further influence the feasibility of lifestyle interventions, affecting adherence to nutritional recommendations or engagement in physical activity. Therefore, pragmatic and scalable approaches—such as standardized lifestyle screening, basic nutritional guidance, and incorporation of digital health monitoring—may represent feasible first steps toward integrating lifestyle determinants into antimicrobial stewardship.
Physical activity as a modulator of the immune response and susceptibility to infection
Immune adaptations to physical activity: From protection to vulnerability
Physical activity profoundly influences the human immune system and is capable of enhancing or suppressing immune surveillance, depending on the intensity, duration, and frequency of exercise. 79 Regular moderate-intensity exercise has been consistently associated with beneficial adaptations in both the innate and adaptive immune responses. For example, moderate aerobic training stimulates the circulation of immunoglobulins, anti-inflammatory cytokines (e.g., IL-10), and natural killer (NK) cells, thereby improving host defense mechanisms and reducing systemic inflammation. Koelwyn et al. (2015) highlighted that exercise modulates multiple components of the inflammation–immune axis, suggesting that physical activity may serve as an adjunctive strategy to enhance immune function, not only in cancer prevention and control but also potentially in the context of infection management.80,81
Additionally, this immunoenhancing effect is supported by clinical and epidemiological studies. Nieman et al. (1994), in a landmark study, reported that individuals engaging in regular moderate exercise experienced up to a 40–50% reduction in upper respiratory tract infections (URTIs) compared with sedentary controls. 82 Similarly, a study by Woods et al. (1999) revealed that physically active older adults had a more robust T-cell response and higher vaccine efficacy rates than their inactive peers did, highlighting the protective effects of exercise on immune aging (immunosenescence). 83 More recently, a randomized trial by Şevgin and Özer et al. demonstrated that a 12-week program of light to moderate exercise significantly increased post–COVID-19 vaccine antibody levels in older adults (65+ years), confirming that physical activity is an effective behavioral adjuvant to improve the vaccine response in elderly individuals. 84
However, excessive or prolonged high-intensity exercise, particularly when combined with inadequate recovery, can lead to transient immunosuppression—a phenomenon often referred to as the “open window” theory. During this period, which lasts from 3 to 72 hours post-exercise, several components of the immune system are temporarily downregulated, including reduced NK cell activity, suppressed salivary IgA levels, and decreased neutrophil function. 85 Marathon runners and endurance athletes, for example, often report an increased incidence of URTI in the days following competition or intense training bouts. 86 Nevertheless, Campbell and Turner (2025) challenge the long-standing “open window” hypothesis by demonstrating that acute bouts of vigorous exercise do not suppress but rather transiently redistribute immune cells to peripheral sites, enhancing immune surveillance and overall immunocompetence across the lifespan. 87
The immunological outcomes of physical activity are therefore described by a J-shaped curve, where sedentary behavior and excessive exercise are associated with increased infection risk, whereas moderate activity is protective. 88 This model underscores the need for dose‒response precision in exercise prescriptions, especially for individuals with underlying immune dysregulation, such as elderly individuals, immunocompromised individuals, or those recovering from illness. According to Gleeson (2013), overtraining syndrome significantly impairs immune function in athletes, increasing vulnerability to infections and reducing overall performance, thus necessitating proactive clinical strategies to preserve immune health during periods of intensive training. 89 Moreover, factors such as nutrition, sleep, psychological stress, and the training environment modulate the immune effects of exercise. 90 Athletes experiencing energy deficiency, poor diet, or chronic stress may exhibit heightened vulnerability to infections despite high fitness levels. Thus, physical activity should be considered within a broader biopsychosocial context when evaluating its effects on immunity and infection susceptibility. 91 Thus, physical activity should be considered within a broader biopsychosocial context when evaluating its effects on immunity and infection susceptibility. In this regard, the immunological benefits of exercise are primarily associated with regular moderate-intensity activity, whereas excessive or prolonged high-intensity training may induce transient immunosuppression and increase infection risk.
Implications for infection risk and antimicrobial therapy
The immunomodulatory effects of physical activity have direct implications for infection susceptibility and, consequently, for the use and efficacy of antimicrobial therapies. Individuals who engage in regular moderate exercise may not only experience fewer infections but also require shorter or less aggressive antimicrobial treatment owing to improved immune competence. Conversely, immunosuppression resulting from overtraining could necessitate earlier or more potent antimicrobial interventions.92,93
Exercise-induced changes in immune function may also affect the pharmacodynamics of antimicrobial agents. Enhanced circulation, improved tissue perfusion, and altered expression of drug transporters in active individuals could theoretically influence drug distribution and efficacy. 94 For example, physical activity has been shown to increase lymphatic flow and microvascular density, potentially facilitating better drug delivery to infected tissues. 95 Although clinical studies on this topic are still emerging, animal models suggest that physical conditioning may improve the effectiveness of antibiotics in systemic infections. 96
Moreover, physical activity can modulate inflammatory pathways that intersect with the immune response to infection. Chronic low-grade inflammation, often observed in sedentary individuals or those with metabolic syndrome, can blunt the immune system’s ability to respond to pathogens and interfere with antimicrobial efficacy. A 2025 meta-analysis of interventional studies confirmed that regular aerobic exercise significantly reduces systemic inflammatory markers—such as CRP, IL-6, and TNF-α—in individuals with metabolic syndrome, thereby potentially increasing the efficacy of antimicrobial treatments for infections such as pneumonia or cellulitis. 97
In special populations—such as hospitalized patients, individuals with chronic respiratory diseases, or elderly individuals—integrating physical activity into the recovery plan may serve as a nonpharmacological adjunct to antimicrobial therapy. For example, pulmonary rehabilitation programs combining exercise and physiotherapy have been shown to reduce respiratory infections and antibiotic use in patients with chronic obstructive pulmonary disease (COPD). Similarly, mobilization strategies in hospital wards have been linked to shorter lengths of stay and a lower incidence of hospital-acquired infections. 98
Given these findings, clinicians should consider a patient’s physical activity level when designing infection management strategies. Physical inactivity may be a modifiable risk factor for both infection occurrence and treatment failure. Moreover, excessive exercise should be approached cautiously in clinical contexts involving acute or unresolved infections. Personalized recommendations that balance immune stimulation with adequate recovery may contribute to better therapeutic outcomes and reduced antimicrobial burden.
These immunological adaptations have direct implications for antimicrobial therapy outcomes. Enhanced innate and adaptive immune responses associated with regular moderate physical activity may improve pathogen clearance, thereby reducing infection severity, shortening treatment duration, and potentially lowering the required antimicrobial dose. Conversely, impaired immune function resulting from physical inactivity or excessive training may contribute to prolonged infections, increased reliance on antimicrobial therapy, and greater variability in drug response. In this context, host immune competence should be considered a key modifier of antimicrobial effectiveness, highlighting the relevance of physical activity as a complementary factor in optimizing therapeutic outcomes and reducing the risk of treatment failure.
Interference of nutraceuticals and functional foods with antimicrobial agents
Nutraceuticals and functional foods can markedly influence the efficacy of antibiotic therapies through a variety of mechanisms. These dietary substances, ranging from vitamins, minerals, and herbal extracts to probiotic foods, may exert synergistic effects that enhance antimicrobial action or antagonistic interactions that impair drug effectiveness or alter pharmacokinetics. Understanding these interactions is crucial for optimizing treatment outcomes and avoiding unintended reductions in antibiotic efficacy.
It is important to note that the evidence supporting interactions between nutraceuticals, functional foods, and antimicrobial agents is derived from heterogeneous sources, including in vitro studies, animal models, and a more limited number of clinical investigations. While experimental studies provide mechanistic insights into potential synergistic or antagonistic effects, their direct translation to clinical outcomes remains uncertain. Clinical evidence, although growing, is still limited and often context-dependent, particularly with respect to dosage, bioavailability, and host-specific factors. Therefore, distinctions between experimentally observed effects, clinically validated interactions, and theoretical or mechanistic hypotheses should be carefully considered when interpreting the impact of nutraceuticals on antimicrobial therapy.
Synergistic effects and therapeutic benefits
Many nutraceuticals have been investigated as adjuvants to conventional antibiotics, with evidence of improved therapeutic outcomes. For example, vitamin E supplementation has demonstrated notable benefits when combined with antibiotics in infection management. In a clinical study of pediatric urinary tract infections, the addition of vitamin E to antibiotic therapy significantly reduced fever and urinary symptoms compared with the use of antibiotics alone. 99 This effect is attributed to vitamin E’s antioxidant and immune-supportive properties, which help alleviate local inflammation and oxidative stress in the urinary tract, thereby facilitating recovery.99,100 In addition to urinary infections, vitamin E has shown broad synergy with antibiotics against drug-resistant pathogens. In vitro studies indicate that vitamin E combined with various antibiotics can restore sensitivity in multidrug-resistant bacteria such as Pseudomonas aeruginosa and MRSA, increasing bacterial killing compared with antibiotics alone. 101 Mechanistically, vitamin E may inhibit bacterial proteins that neutralize antibiotics; for example, it interferes with a bacterial lipocalin that binds and sequesters antibiotics. Vitamin E also modulates host immunity; supplemental vitamin E has been found to increase the ability of neutrophils to kill Streptococcus pneumoniae, suggesting a nutritional strategy to bolster host defense during antibiotic treatment. 102 Collectively, these findings highlight that certain vitamins as nutraceuticals can serve as adjunct therapies that increase antimicrobial efficacy and help combat resistant infections.
Polyphenols and phytochemicals
Similarly, polyphenols and other phytochemicals in functional foods exhibit synergistic interactions with antibiotics. Dietary flavonoids are prominent examples: compounds such as quercetin and rutin, which are abundant in fruits and vegetables, have intrinsic antimicrobial properties and can work in tandem with antibiotic agents. Quercetin or rutin combined with conventional antibiotics significantly improved bacterial killing of clinical multidrug-resistant strains. In their study, the addition of these flavonoids restored the efficacy of gentamicin and ceftriaxone against resistant E. coli and Proteus mirabilis isolates, whereas antibiotics alone had limited effects.103–105 Quercetin in particular has a strong synergistic effect with ceftriaxone, presumably due to its ability to disrupt bacterial membranes or inhibit efflux pumps, thereby allowing the antibiotic to accumulate at the infection site. Since quercetin and rutin are common dietary components, their use as nutraceutical adjuvants is a promising and accessible strategy to enhance antibiotic therapy and possibly reduce the required dosage of conventional drugs. Other plant-derived nutraceuticals under investigation include garlic allicin, tea catechins, curcumin, and propolis, many of which have been shown to potentiate various antibiotics in laboratory studies.103–105 For example, green tea epigallocatechin (EGCG) can synergize with β-lactam antibiotics against Staphylococcus by damaging bacterial cell walls and inhibiting β-lactamase enzymes, according to some reports. 106 These synergistic interactions suggest that diets rich in polyphenols or supplemented with specific nutraceutical extracts could improve clinical outcomes in infections, especially those caused by resistant microbes, by increasing antimicrobial potency.
Microbiome and immune modulation
In addition to direct antimicrobial synergy, nutraceuticals may benefit antibiotic therapy through immune modulation and microbiome support. Probiotic functional foods are a case in point: while not antibiotics themselves, probiotics (live beneficial microbes in yogurt, fermented foods, etc.) can enhance treatment by maintaining the gut microbiota balance and preventing opportunistic infections such as C. difficile. Clinical trials have shown that the coadministration of probiotics with antibiotics can reduce antibiotic-associated diarrhea and yeast overgrowth, thereby improving patient comfort and compliance. 107 In one randomized trial, patients with recurrent tonsillitis who received Lactobacillus probiotics alongside antibiotics experienced faster symptom resolution and fewer secondary infections than did those who received antibiotics alone. These benefits likely stem from probiotics inhibiting pathogenic bacteria and modulating the host immune response. Similarly, prebiotic and nutraceutical fibers in functional foods (e.g., inulin) may feed protective gut bacteria during antibiotic treatment, helping to mitigate dysbiosis and speed recovery. 108 While probiotics do not directly attack the infectious agent, their interference is beneficial in that they counteract the collateral damage caused by antibiotics, highlighting a positive relationship between diet and antimicrobial therapy.
Antagonistic interactions and impaired efficacy
Not all nutraceutical interactions are beneficial for certain foods, and supplements can negatively interfere with antibiotics, reducing their absorption or activity. One well-known example is the interaction between calcium-rich foods and tetracycline antibiotics. Divalent minerals such as calcium, magnesium, and iron can chelate tetracyclines and fluoroquinolones in the gut, forming insoluble complexes that the body cannot absorb. Even a small amount of dairy can drastically impair tetracycline bioavailability. The addition of milk (containing minimal calcium) to coffee or tea severely reduces tetracycline absorption in healthy volunteers, decreasing drug availability in the bloodstream. The presence of calcium ions needs to be carefully controlled to avoid >50% loss of active drug levels. Similarly, taking oral antibiotics with supplements such as calcium tablets, iron tonics, or antacids has been reported to significantly reduce absorption; one pharmacokinetic study revealed that milk coadministration reduced tetracycline absorption by ∼30% and iron supplementation by over 75%. 109 Thus, patients are usually advised to avoid dairy or mineral supplements around the time of dosing for antibiotics susceptible to chelation. This food‒drug interaction exemplifies how a functional food (milk) can inadvertently cause treatment failure by preventing adequate antibiotic uptake. 110
Herbal nutraceuticals and botanical supplements can also alter antibiotic effectiveness via metabolic interactions. Certain herbs modulate the same liver enzymes that metabolize drugs, leading to changes in antibiotic clearance. For example, St. John’s Wort, a popular herbal antidepressant, is a potent inducer of cytochrome P450 (CYP3A4) enzymes and P-glycoprotein transporters. Coadministration of St. John’s Wort has been shown to lower the plasma levels of various drugs; in the context of antibiotics, it could reduce the concentrations of macrolides or antitubercular drugs that are CYP3A4 substrates, risking subtherapeutic exposure. 111 Conversely, grapefruit and certain citrus fruits act as CYP3A4 inhibitors in the intestine. Drinking grapefruit juice can increase the systemic levels of some antibiotics by preventing their first-pass metabolism. A classic case is the interaction with erythromycin (a macrolide antibiotic): grapefruit juice can increase erythromycin blood levels, which may exacerbate side effects such as QT prolongation. 112 Although not all antibiotics are affected, clinicians caution against grapefruit when patients take medications such as erythromycin or clarithromycin. Similarly, traditional Chinese and Ayurvedic herbal formulations containing compounds such as berberine have been shown to influence antibiotic pharmacokinetics. In a rat study, the herbal remedy Huang Lian (rich in berberine) significantly altered the oral absorption of ciprofloxacin, likely by inhibiting metabolic enzymes and efflux pumps in the gut. 113 These findings underscore the need for awareness of herb‒drug interactions during antibiotic therapy, while nutraceuticals are “natural,” they can behave pharmacologically like drugs and must be considered in the regimen to avoid unexpected under- or over-exposure to the antibiotic.
In some cases, nutraceutical interference may protect bacteria and promote antimicrobial resistance. Antioxidant supplements are a double-edged sword in this context. Bactericidal antibiotics often kill microbes by generating reactive oxygen species (ROS) inside bacteria, contributing to oxidative damage to bacterial proteins and DNA. High doses of exogenous antioxidants (e.g., vitamin C and N-acetylcysteine) might quench these ROS, unintentionally shielding bacteria from antibiotic-induced oxidative stress. Laboratory experiments have shown that the addition of antioxidants can significantly reduce the killing ability of several bactericidal antibiotics.114,115 In one study, antioxidant compounds and even bacterial antioxidant enzymes were found to reduce antibiotic lethality, resulting in increased survival of bacteria in the presence of drugs such as quinolones. This effect could allow some bacteria to persist and possibly develop resistance mutations. In one study, antioxidant compounds—and even bacterial antioxidant enzymes—were shown to reduce antibiotic lethality, resulting in increased bacterial survival in the presence of drugs such as quinolones. This effect could allow some bacteria to persist and potentially develop resistance mutations. However, the role of reactive oxygen species (ROS) in antibiotic-mediated killing remains debated, and its relative contribution to bacterial lethality is not fully established. While some experimental studies suggest that exogenous antioxidants may attenuate bactericidal activity by reducing antibiotic-induced oxidative stress, others report context-dependent or minimal effects. These discrepancies highlight the complexity of ROS-mediated mechanisms and suggest that the impact of antioxidant supplementation on antibiotic efficacy may vary depending on the drug class, bacterial species, and experimental conditions.114,116
Interestingly, a paradoxical effect has also been reported, as certain antioxidants may reduce antibiotic-induced oxidative DNA damage in bacteria, thereby lowering mutation rates and potentially slowing the development of resistance. Therefore, caution is warranted when interpreting the interaction between antioxidants and antimicrobial therapy.114,116 These nuanced outcomes illustrate that the timing, type, and dose of nutraceutical antioxidants relative to antibiotic therapy are critical. In general, it is advisable to avoid indiscriminate high-dose antioxidant use during antibiotic treatment unless supported by evidence, as it may undermine the antibiotic’s mode of action. This example highlights how metabolic context matters: the same nutraceutical can have either antagonistic or beneficial interference with antimicrobials depending on the scenario.
Interactions between nutraceuticals/functional foods and antimicrobial agents.
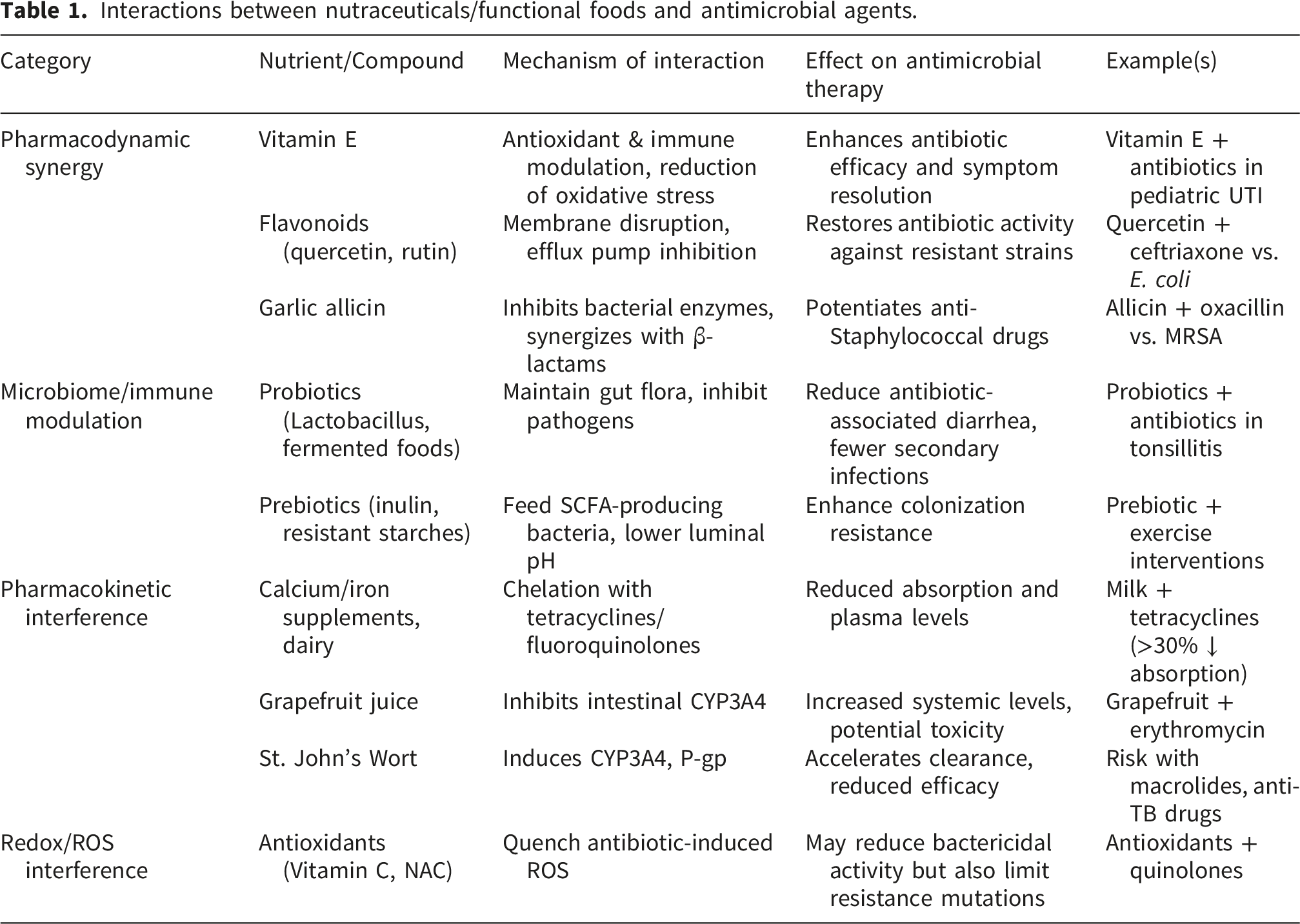
Moving forward, clinicians and researchers emphasize the importance of considering a patient’s diet and supplement use as part of antimicrobial stewardship. By recognizing both synergistic opportunities (e.g., combining antibiotics with specific nutraceuticals to overcome resistance) and antagonistic risks (e.g., nutrient‒drug binding or metabolism interference), healthcare providers can tailor treatments better. In practice, this means advising patients on the proper timing of meals/supplements relative to antibiotic dosing, exploring evidence-based nutraceutical adjuncts in difficult infections, and monitoring for any unexpected changes in drug efficacy that could arise from dietary habits. Ultimately, integrating nutrition science with pharmacology offers a more holistic approach to infection management, one that could improve outcomes and help curb antibiotic resistance through complementary strategies.
Design of antimicrobials considering the metabolic profiles of physically active vs. sedentary individuals
Differences between physically active and sedentary individuals and implications for antimicrobial pharmacology.

In this context, emerging nanocarrier systems with stimuli-responsive properties may offer potential for adaptive drug delivery. For example, pH-sensitive or temperature-responsive nanocarriers could theoretically modulate drug release in response to physiological changes associated with physical activity, such as transient acidosis or increased tissue temperature during exercise. 91
Body composition and drug distribution
One key difference between active and sedentary individuals is body composition. Regular exercise tends to increase fat-free mass (muscle, bone, water) and reduce adipose tissue. This shift influences the volume of distribution (Vd) of drugs. Hydrophilic antibiotics (such as β-lactams or aminoglycosides) are largely distributed in body water compartments; an active person with more total body water and plasma volume may have a larger distribution space for such drugs, potentially lowering peak concentrations. 117 In contrast, lipophilic antibiotics (e.g., macrolides and fluoroquinolones) tend to accumulate in fatty tissues; sedentary individuals with greater body fat might sequester more lipophilic drugs in adipose stores, effectively increasing Vd and prolonging the drug’s half-life in the body. Conversely, a lean athlete might have less reservoir for a lipophilic drug, leading to higher circulating levels for a given dose. Studies have shown that weight loss and reduced adiposity can increase the bioavailability and distribution of certain drugs that otherwise partition into fat. 118 This implies that an antibiotic’s dosing may need adjustment based on body composition; for example, in an obese sedentary patient, a higher mg/kg dose or loading dose might be required for hydrophilic antimicrobials to adequately penetrate tissues, whereas for lipophilic drugs, caution is needed to avoid accumulation. 119 Indeed, clinical guidance now often recommends using adjusted body weight dosing for obese patients for drugs such as aminoglycosides to balance underdosing (if total weight is used) and overdosing (if lean weight alone) drugs are used. Recent studies have provided updated antimicrobial dosing recommendations specifically for obese (often sedentary) individuals, acknowledging that standard doses may not achieve the same concentrations owing to altered pharmacokinetics. In drug design, this could translate to developing formulations that are effective across a range of body compositions, such as longer-acting depot injections that bypass extreme distribution differences or nanocarriers that target drugs to specific tissues regardless of fat content. 120
From a clinical pharmacology perspective, these physiological differences may have practical implications for antimicrobial dosing. For example, hydrophilic antibiotics such as β-lactams and aminoglycosides, which depend on renal clearance and extracellular fluid distribution, may exhibit altered peak concentrations and clearance rates in physically active individuals with increased plasma volume and renal perfusion. 117 Conversely, in sedentary or obese patients, increased adiposity and reduced tissue perfusion may lead to suboptimal drug penetration, particularly in poorly vascularized tissues. Clinical observations in obese populations have already demonstrated variability in antimicrobial exposure and an increased risk of underdosing when standard regimens are applied. These findings suggest that patient-specific factors such as physical activity level and body composition should be considered when optimizing dosing strategies, particularly for drugs with narrow therapeutic windows or concentration-dependent activity.
Protein binding and free drug levels
Physical activity can acutely alter plasma biochemistry, which in turn affects drug binding. During exercise, there is an increase in circulating free fatty acids (FFAs) as the body mobilizes fat for energy. FFAs can displace drugs from their binding sites on serum albumin and other proteins because FFAs themselves have a high affinity for albumin. This is especially relevant for antibiotics that are moderately to highly protein-bound (e.g., oxacillin, ceftriaxone, doxycycline). 121 An elegant animal experiment demonstrated that exercise-induced stress (repeated cold-water swimming in rats) led to elevated FFAs and was associated with significantly increased free levels of ampicillin in the bloodstream. Compared with sedentary controls, rats subjected to daily exercise presented increased serum and tissue ampicillin concentrations, despite receiving the same dosage. 122 This was attributed to FFAs competitively displacing ampicillin from protein binding sites, effectively increasing the active unbound fraction of the drug.
In human terms, an athletic person who engages in exercise around the time of dosing might experience transiently higher free antibiotic levels, which could increase bacterial killing but also risk toxicity if the spike is significant. Drug designers could consider these dynamics by, for example, formulating antibiotics with binding characteristics that are less susceptible to displacement or by advising dosing schedules (timing doses during rest periods) in physically active patients. 123 Additionally, chronic exercise tends to lower baseline triglyceride and possibly free fatty acid levels at rest (due to improved metabolic efficiency), which might mean that chronically active individuals have lower interference with protein binding outside of acute exercise bouts. Tailoring antibiotic dosing or delivery (such as sustained-release formulations) could help maintain stable free drug levels in the face of these fluctuations. 124
Cardiovascular function and tissue perfusion
Another distinction is that physically fit individuals usually have greater cardiac output reserve and muscle perfusion, especially during activity, whereas sedentary individuals may have poorer peripheral circulation. Enhanced blood flow in active persons can aid in the distribution of antibiotics to infection sites. During exercise, blood flow to skeletal muscles, skin, and lungs increases, whereas splanchnic (gut) and renal blood flow may decrease.125,126 These changes can have immediate effects on drug kinetics. An early clinical pharmacology study by Ylitalo et al. (1977) demonstrated that intense physical exercise beginning shortly after oral antibiotic intake significantly altered the pharmacokinetic profile of the drug in humans. In that crossover trial, volunteers who performed vigorous exercise after taking tetracycline or doxycycline had higher serum concentrations and area under the curve (AUC) values for the antibiotic than did those at rest (Ylitalo et al., 1977). Exercise strongly suppresses renal blood flow and thus renal excretion of drugs, leading to their accumulation in the plasma. Although total urinary recovery of the dose was unchanged (implying that overall absorption was similar), the rate of absorption appeared faster and peak levels were higher during exercise, likely due to increased gastrointestinal motility and perhaps quicker transit to absorption sites. 127
This study highlighted that a person’s physical activity at dosing time can impact antibiotic exposure; consequently, dosing strategies might be adjusted (e.g., avoiding immediate intense exercise after taking an antibiotic that is heavily reliant on renal clearance to prevent excessive levels). For drug development, one might consider designing antimicrobials with robust pharmacokinetics that are less sensitive to transient changes in perfusion. For example, antibiotics eliminated primarily by the liver (hepatic clearance) with low extraction ratios might be less affected by exercise-induced liver blood flow reduction than high-extraction drugs are. Recognizing these differences, modern physiologically based pharmacokinetic (PBPK) models sometimes include scenarios for exercise vs. rest to predict drug levels under various conditions. 127
Chronic differences in perfusion and organ function also play a role. Sedentary individuals, especially if they have underlying conditions such as diabetes or vascular disease, may have impaired microcirculation in tissues (slower antibiotic penetration into, say, peripheral limb infections). In contrast, an athlete’s well-vascularized muscle might achieve higher antibiotic tissue concentrations, aiding infection control in those tissues. Drug designers might respond to this by targeting drug delivery for at-risk populations. For example, for a predominantly sedentary population with peripheral artery disease, developing an antibiotic with better tissue penetration or using drug carriers that home to poorly perfused tissue could improve outcomes.128,129 Nanocarrier antibiotics could be engineered to exploit alternate transport pathways (e.g., lymphatic delivery) to reach infection sites in patients with compromised circulation. Moreover, in highly active patients, one might worry about antibiotics being distributed so well into muscle that serum levels drop faster—as was observed with digoxin in athletes—although for most antibiotics, this is less of an issue given regular dosing. 130 Nonetheless, exercise-induced changes in Vd and clearance underscore the importance of confirming dosing in both active and sedentary subjects during drug development rather than only testing drugs in resting individuals. Historically, drug trials often assume a resting baseline; Basso (2017) argues that failing to account for exercise can miss important kinetic interactions. 131 The incorporation of exercise conditions in preapproval studies or at least postmarketing surveillance can ensure that dosing guidelines are adequate for the physically active segment of patients. 132
Metabolic enzyme activity
Perhaps the most significant impact of lifestyle on pharmacology is through metabolic enzyme modulation. Long-term physical activity (exercise training) has been associated with changes in the expression of drug-metabolizing enzymes in the liver and other tissues. Regular endurance exercise can increase liver volume and increase hepatic blood flow at rest, potentially increasing the content of cytochrome P450 (CYP) enzymes, which are responsible for drug metabolism. Niederberger and Parnham (2021) noted that specific types of chronic exercise have been linked to increased clearance of drugs, likely due to increased CYP450 expression (and concomitant increases in mitochondrial density that can support metabolic processes). 133 For antibiotics that are metabolized primarily by the liver, an active individual might clear the drug more rapidly, shortening its half-life and reducing exposure unless doses are adjusted.
For example, if an antibiotic is metabolized by CYP3A4, one could speculate that a marathon runner might have slightly higher 3A4 activity (akin to how other inducers work), necessitating a higher dose or more frequent dosing to maintain therapeutic levels, although concrete clinical data on specific antibiotics are still limited. On the other hand, acute strenuous exercise can transiently suppress certain metabolic functions. Intense exercise triggers the release of inflammatory cytokines and stress hormones, which, in some studies, lead to temporary downregulation of the mRNAs of some CYP enzymes.134,135 There is also an exercise-induced drop in blood pH (lactic acidosis) during heavy exertion, which can alter enzyme kinetics and drug ionization states. For drugs with pH-sensitive absorption or distribution (e.g., weakly basic antibiotics such as aminoglycosides), acidosis can increase the unbound fraction in plasma and tissues. These complex effects indicate that the timing of exercise relative to dosing could momentarily change how fast a drug is metabolized or eliminated. 136
From a drug design perspective, one might mitigate these variances by developing prodrugs or formulations activated by specific metabolic pathways that are more consistent between individuals. For example, an antimicrobial prodrug that relies on ubiquitous esterases (which are less likely to be affected by lifestyle) might achieve more predictable activation in both sedentary and active bodies than one that depends on a particular CYP enzyme level. Alternatively, developers might create dosing algorithms that incorporate patient activity level as a factor — akin to how renal dosing is adjusted for kidney function. As pharmacogenomics and digital health monitoring advance, it is conceivable that future antibiotic prescriptions could use wearable data (such as activity trackers) to inform dosing schedules (e.g., “take this dose in the evening when at rest” for drugs affected by exercise or use higher nightly doses if daytime exercise increases clearance). While this level of personalization is still emergent, it underscores that precision medicine for infections could extend beyond genetic factors to include lifestyle factors such as exercise. 137
Immune response and inflammation
In addition to pharmacokinetics, physical activity versus sedentariness can influence the host’s baseline immune competence and inflammatory status, which in turn affects infection outcomes and possibly the required potency of an antimicrobial. Regular moderate exercise is known to enhance immune surveillance and reduce chronic inflammation, whereas a sedentary lifestyle (especially obesity) is often associated with a state of chronic low-grade inflammation and immune dysregulation. An active individual might mount a faster or stronger immune response to infection, effectively assisting the antibiotic in clearing the pathogen. In such cases, the therapeutic efficacy of a bacteriostatic antibiotic, for example, might be greater in an active person because their immune system can quickly eliminate growth-inhibiting bacteria.138,139
On the other hand, sedentary patients, particularly those with metabolic syndrome or frailty, might have impaired leukocyte function and a sluggish immune response, placing greater onus on the antibiotic to perform. This suggests that when new antimicrobials are designed or antibiotic classes are chosen, one could consider the typical patient profile. For example, for an antibiotic intended primarily for hospital-associated infections in immobile, critically ill patients (very sedentary by circumstance), it might be advantageous to design the drug to have some immunomodulatory or anti-inflammatory properties to counteract the host’s weak immune contribution. 140 Some newer antimicrobial and host-directed therapies aim to modulate the immune system (e.g., boosting macrophage activity) in tandem with direct antibacterial effects, which could be particularly useful for sedentary or immunocompromised populations. Conversely, if an antibiotic is developed for generally healthy outpatients (who may include young athletic individuals), the priority might be maximizing pharmacokinetic stability during varied daily activities and minimizing any deleterious effects on exercise capacity (for example, avoiding mitochondrial toxicity that could hamper muscle function or avoiding agents that carry the risk of tendon injury, which is exacerbated in active people). 141
A salient example of tailoring drug design to active vs. sedentary users is the case of fluoroquinolone antibiotics and tendon toxicity. Fluoroquinolones (such as ciprofloxacin and levofloxacin) are effective broad-spectrum agents, but they are infamous for causing tendinopathy and even tendon rupture in some patients, especially those who are physically active. Athletes and military trainees on fluoroquinolones have reported that Achilles tendon injuries are likely due to the effects of drugs on collagen turnover and the blood supply to tendons. 142 This side effect profile has prompted caution in prescribing these drugs to younger, athletic individuals unless absolutely necessary. This finding also suggests the need to develop safer antimicrobial alternatives for active populations, such as the design of new quinolone derivatives without the chemical substructures implicated in tendon damage or the use of nanocarrier delivery to infected tissues to reduce systemic exposure to joints and tendons. Thus, the “physically active” metabolic profile concerns not only kinetic differences but also unique adverse effect susceptibilities. Drug development pipelines sometimes assess musculoskeletal toxicity in animal models under exercise conditions to address issues that might only manifest with mechanical strain.
Thus, while an antibiotic’s primary design goals are usually centered on broad-spectrum efficacy and minimizing resistance and toxicity, considering the patient’s metabolic profile is an emerging aspect of personalized antimicrobial therapy. Physically active and sedentary individuals represent two ends of a spectrum, and optimal drug design strives for efficacy across this range. By accounting for differences in distribution (body composition), metabolism (enzyme levels, organ perfusion), excretion (renal function variations), and host factors (immune response, tissue environment), we can develop antimicrobials that are robust to these variables or even tailored to specific subpopulations. Such foresight in design and testing can improve clinical outcomes—ensuring that, whether a patient is an athlete or couch-bound, they receive the full benefit of the antimicrobial with minimal side effects, ultimately aiding in better infection control and reducing the chance of resistance stemming from underdosing or erratic drug levels. The relationship of lifestyle and drug action is a reminder that infectious disease treatment is not one-size-fits-all, and future antibiotics may come with the nuance and flexibility required to treat patients as whole persons with unique physiological contexts. 143
Influence of macronutrient composition on drug bioavailability and distribution
The oral bioavailability of an antimicrobial drug—defined as the proportion of the administered dose that reaches systemic circulation without undergoing alteration—is a pivotal factor in attaining therapeutic plasma concentrations that are sufficient to eradicate pathogens without fostering resistance owing to underdosing. This pharmacokinetic parameter is contingent upon drug-specific and host-related variables, including first-pass hepatic metabolism, the activity of intestinal enzymes (CYP3A4, UGT), and the expression of transporters (P-gp, OATP, PepT1) in the gastrointestinal epithelium. 144 The concurrent ingestion of macronutrients (lipids, proteins, carbohydrates) has been demonstrated to exert a substantial influence on the solubility, dissolution, and permeability of orally administered antimicrobials. 145 These alterations have the capacity to impact pivotal pharmacokinetic parameters, including C_(max) (maximum plasma concentration), AUC (area under the concentration‒time curve), and T_(max) (time to reach C_(max)), which may in turn affect the clinical efficacy of the pharmaceutical agent under examination. 146
The absorption of lipophilic drugs is contingent upon their incorporation into micelles that are formed by bile salts. 147 The incorporation of dietary lipids has been demonstrated to increase micellar solubilization, thereby increasing the dissolved drug fraction and promoting membrane permeability. 148 Conversely, proteins may impede the uptake of hydrophilic drugs, such as cephalosporins, via PepT1 or antibiotics utilizing amino acid transporters, thereby potentially diminishing absorption rates. 149 Carbohydrates, particularly those with a high glycemic index, have a significant effect on the motility of the stomach and the pH of its contents. This influence results in alterations to the dissolution and ionization states of pH-sensitive compounds.
Acidic gastric conditions have been shown to facilitate the maintenance of drugs with pKa values near gastric pH in nonionized forms, thereby reducing permeability. 150 Conversely, premature ionization has the potential to compromise solubility in the proximal intestine. 151 These dynamics are relevant for antimicrobials characterized by narrow therapeutic windows or time-dependent efficacy parameters (T>MIC, C_max/MIC). 152 Alterations in macronutrient composition have been demonstrated to exert a substantial influence on antimicrobial pharmacokinetics, as evidenced by both experimental models and population studies. 153 High-fat environments enhance the bioavailability of lipophilic drugs, such as posaconazole and albendazole. Conversely, fluctuations in protein and carbohydrate intake have been demonstrated to influence the distribution and clearance of hydrophilic drugs, such as cephalexin and flucloxacillin154. It is imperative to consider these drug–nutrient interactions when formulating dosage regimens with the objectives of optimizing therapeutic concentrations, minimizing toxicity, and preventing resistance. 155
The clinical implications extend beyond the mere act of dosing, whether with or without food, and necessitate the incorporation of nutritional considerations into therapeutic guidelines. 156 This integration is associated with the adjustment of pharmaceutical forms (tablet, capsule, and suspension) according to the patient’s nutritional status. 157 In the domain of antimicrobial therapy, it is essential to examine both the pathogen’s susceptibility to treatment and the potential for nutrient‒drug interactions. This consideration assumes particular significance within the framework of personalized medicine.
The process of drug absorption within the gastrointestinal tract is governed by physiological mechanisms that regulate solubility, transport, and permeability. Lipid ingestion serves as a paradigm: high-fat meals stimulate bile secretion, leading to micelle formation that solubilizes lipophilic drugs that are otherwise poorly soluble in aqueous conditions. 158 Compared with fasting micelles, these micelles facilitate transcellular transport, increasing both C_(max) and AUC. The presence of lipids has been demonstrated to delay gastric emptying, thereby prolonging the drug’s presence in regions conducive to absorption, such as the duodenum and jejunum. This prolongation in the absorption process has been shown to enhance the overall uptake of the drug. 159
This mechanism is highly relevant to azole antifungals. 160 In controlled pharmacokinetic studies, compared with fasting, posaconazole administered with a high-fat meal increased C_(max) by 16% and AUC_(0–72 h) by 51%. Another crossover study demonstrated that a 300 mg dose administered with a high-fat breakfast nearly doubled systemic exposure (AUC and C_(max)) compared with the fasted state. The observed effects are attributed to enhanced micellar solubilization and delayed gastric clearance. 161 Therefore, it is imperative that posaconazole be administered concomitantly with a fatty meal to ensure the attainment of therapeutic levels and to mitigate variability, which has the potential to engender resistance.
However, notably, not all lipophilic drugs benefit from lipid coadministration. A recent quantitative analysis of 19 quinolones revealed that delafloxacin led to a >40% C_(max) reduction and a delayed T_(max) (1.5–2 h) following a high-fat meal, without a significant change in the AUC.162,163 A recent systematic review of fluorinated drugs revealed that, while the rate of absorption is reduced, the extent of systemic exposure remains unaltered.
Albendazole, a benzimidazole carbamate anthelmintic, undergoes first-pass metabolism to its active sulfoxide. Its systemic exposure markedly increases in response to a high-fat meal. In a randomized crossover study, a 400 mg dose resulted in a twofold increase in both the C_(max) and the AUC of the parent drug and sulfoxide metabolite compared with a low-fat meal (p < 0.05). 164 A meta-analysis of 91 time series studies demonstrated a 14% increase in sulfoxide bioavailability and a 2.8-fold increase in AUC400 with fatty meals (both p < 0.01). 165
The hypothesis that peptides derived from protein-rich meals may compete with cephalexin for PepT1-mediated transport is both mechanistically sound and supported by the known substrate specificity of the transporter. 166 Although the prevailing focus of extant pharmacokinetic studies has focused on factors such as gastric pH, divalent cation chelation, and renal clearance, there is an ongoing absence of systematic examinations of the influence of dietary protein on cephalexin’s C_(max) and AUC. 167 Further research is necessary to better define the role of protein intake in modulating transporter-mediated absorption and its potential implications for therapeutic efficacy. Such research should be conducted in in vitro and clinical settings.
Chelation with divalent cations (e.g., Ca2+) remains a classic mechanism for tetracyclines. The extent of this phenomenon is contingent upon the formulation and type of salt utilized. In controlled studies, the administration of 100 mg of doxycycline hyclate tablets following high-fat, dairy-rich meals resulted in 24% and 13% reductions in C_(max) and AUC, respectively. 168 This effect was attributed to the formation of insoluble doxycycline–calcium complexes. A comparable investigation involving DFD-09 capsules (40 mg) revealed 11% and 14% decreases in C_(max) and AUC0-24, respectively. In contrast, doxycycline monohydrate (200 mg immediate-release tablets) demonstrated comparable bioavailability irrespective of whether the subject was in a fasted or fed state. This finding indicates that complexation is reduced under the conditions of the pH of the stomach. 45
It has been demonstrated that meals with a high glycemic index elicit a robust insulin response, concomitant with an elevation in peptide hormones such as cholecystokinin (CCK) and gastrin. In a randomized trial, high-GI meals resulted in elevated insulin indices compared with low-GI meals, indicative of significant postprandial hyperinsulinemia. 169 This state has been shown to prolong gastric acid exposure, thereby affecting pH-sensitive drugs in the gastrointestinal tract. Recent findings have shown that delayed gastric emptying and lower pH—mediated by CCK and gastrin—hinder the dissolution and absorption of drugs with pKa ≈ 3, such as penicillin derivatives. For example, the administration of flucloxacillin (pKa ≈ 2.7) following a high-carbohydrate breakfast resulted in a 15–20% reduction in C_(max), a 0.5–1 h delay in T_(max), and an ∼10% decrease in the AUC compared with fasting. 170
Noninsorbed complex carbohydrates reach the colon, where microbiota fermentation produces short-chain fatty acids (SCFAs) (acetate, propionate, butyrate), which reach high luminal concentrations. SCFAs have been shown to modulate colonic physiology by inducing the expression of transporters such as MCT1 and GPR41/43. 171 This affects the disposition of nutrients and metabolites within the colon. Although most orally administered antimicrobials are absorbed in the upper segments of the gastrointestinal tract, drugs subject to enterohepatic recirculation may undergo secondary absorption in the colon. 172 This process has the potential to prolong systemic exposure and give rise to multiple plasma concentration peaks. Pharmacokinetic simulations have demonstrated that such recycling increases the area under the curve (AUC) for high-hepatic-extraction drugs. 173 This finding underscores the necessity of considering the modulation of SCFA-mediated processes in relevant models.
The interactions between macronutrients and oral antimicrobials can be categorized into several domains, including micellization, transporter competition, complex formation, and changes in motility and luminal pH. Clinical guidelines ought to comprise recommendations founded upon patient nutritional profiles, antimicrobial properties, and formulation characteristics. To reduce pharmacokinetic variability and the development of resistance, interdisciplinary strategies grounded in pharmacology, nutrition, and gastrointestinal physiology must be employed.
Recent findings support the adaptation of antimicrobial dosing according to meal composition. For drugs such as albendazole and posaconazole, whose efficacy is contingent upon surpassing MIC thresholds, coadministration with lipid-rich foods is recommended. Conversely, for delafloxacin, the formulation chosen to mitigate food interactions is of paramount importance. The extent of pharmacokinetic alterations is determined by the presence of calcium and the formulation type of tetracyclines. The optimization of therapeutic regimens and the minimization of variability and resistance are contingent upon the integration of clinical nutrition into antimicrobial stewardship.
Oxidative stress, antioxidants, and synergy or antagonism with antimicrobial therapies
Reactive oxygen species (ROS) are continuously produced as byproducts of cellular metabolism, particularly within the mitochondrial electron transport chain, and play a central role in immunity. 174 These radical and nonradical species, including superoxide (O2·−), hydrogen peroxide (H2O2), and peroxynitrite (ONOO−), serve as key signaling molecules in the innate immune response, promoting inflammasome activation, phagocyte recruitment, and phagosome‒lysosome fusion. 175 Furthermore, they contribute to microbial clearance by facilitating transition metal efflux and disrupting microbial membranes. The production of ROS has been shown to exceed the host’s antioxidant defenses, leading to an occurrence referred to as oxidative stress. 176 This process results in damage to lipids, proteins, and nucleic acids. This duality signifies that, while ROS enhance antimicrobial defense, their dysregulation can exacerbate inflammation and tissue injury, potentially prolonging infection or triggering systemic dysfunction. 177
In the context of bacterial and fungal infections, phagocytic cells—mostly neutrophils and macrophages—are responsible for generating ROS through the process of activating the nicotinamide adenine dinucleotide phosphate (NADPH) oxidase complex. 178 ROS, including hydrogen peroxide (H2O2), superoxide (O2·−), and hypochlorite (HOCI), have been shown to exert direct antimicrobial effects by disrupting microbial membranes, denaturing proteins, and oxidizing intracellular targets. In addition to their microbicidal role, ROS modulate adaptive immunity. H2O2 has been demonstrated to play a pivotal role in the maturation of T cells, the process of antigen presentation, and the polarization of T-helper subsets. These functions are critical for the resolution of persistent infections. Nevertheless, elevated levels of ROS have been reported to induce lipid peroxidation in host tissues. 179 This process has been shown to promote the release of proteolytic enzymes, such as myeloperoxidase and lysozyme, which contribute to the amplification of inflammatory cascades. The culmination of these cascades may result in severe outcomes, including sepsis and irreversible organ damage. 180
The efficacy of certain antimicrobial agents is modulated by the host’s oxidative environment. For example, some bactericidal antibiotics—such as fluoroquinolones and β-lactams—exert part of their activity through the generation of ROS. 181 These drugs have been shown to induce oxidative stress within bacterial cells, thereby enhancing DNA damage and activating repair systems. Recent studies have demonstrated that supplementation with exogenous antioxidants, such as N-acetylcysteine (NAC) or vitamin C, can reduce antibiotic-induced ROS production and increase bacterial survival. 182 These findings indicate a possible antagonistic interaction between potent antioxidants and antimicrobials that is contingent upon ROS-mediated bactericidal mechanisms.
Antioxidants do not merely function as ROS scavengers. Some antibiotics also exhibit intrinsic antimicrobial properties or synergize with conventional antibiotics to enhance bacterial eradication. For example, the concomitant administration of vitamin C with β-lactams or aminoglycosides has been shown to enhance bactericidal activity against strains of Escherichia coli and Klebsiella pneumoniae that are resistant to β-lactams. 183 This increase is attributed primarily to an increase in membrane permeability. Research in this area has demonstrated that persister cells—metabolically quiescent bacteria refractory to treatment—exhibit a paradoxical elevation in intracellular ROS levels in response to vitamin C, resulting in increased efficacy of ciprofloxacin and gentamicin184.
In a similar manner, NAC has demonstrated synergy with gentamicin in eradicating Pseudomonas aeruginosa biofilms through a reduction in exopolysaccharide production and an increase in antibiotic penetration into the biofilm matrix. 185 These findings indicate that certain antioxidants may exert a regulatory influence on bacterial physiology that extends beyond their traditional function as redox buffers. 186
The antioxidant properties of compounds such as vitamin C, NAC, and polyphenols offer protection against oxidative tissue damage. However, these compounds may counteract the efficacy of antimicrobials whose bactericidal action relies on ROS generation. 187 Several studies have demonstrated that coadministration of high-dose antioxidants significantly reduces antibiotic-induced ROS production, resulting in an increase in bacterial survival in survival curve assays. Similarly, models of Staphylococcus aureus infection demonstrated diminished activity of oxacillin, fosfomycin, and vancomycin when they were administered concomitantly with polyphenol-rich supplements. 188 These findings suggest the potential of antioxidants to prevent the eradication of pathogens, particularly when used concurrently with ROS-dependent antibiotics.
These interactions between nutraceuticals and pharmaceutical drugs are of significant therapeutic importance, particularly in patients with chronic comorbidities such as diabetes mellitus or cardiovascular disease, where antioxidant supplementation (e.g., vitamin C, NAC, polyphenols) is frequently utilized to reduce oxidative stress. 188 In such cases, the utilization of bactericidal antibiotics, whose efficacy is contingent upon ROS production, necessitates meticulous adjustment. 189 To address this potential for antagonism, clinical strategies have been developed. These strategies include the administration of antioxidants a minimum of two hours before or after the antibiotic dose, or alternatively, the selection of bacteriostatic agents whose mechanisms do not depend on oxidative stress. The objective of these approaches is twofold: first, to prevent pharmacodynamic interference, and second, to preserve antimicrobial effectiveness.
In the context of chronic infections and biofilm-associated diseases, where pathogens frequently demonstrate elevated levels of resistance and necessitate intensified treatment regimens, the concurrent administration of antioxidants and antimicrobials has been demonstrated to have a synergistic effect. 190 In different models of Mycobacterium tuberculosis, the combination of oral vitamin C with rifampicin and isoniazid resulted in an increase in pulmonary bacterial clearance compared with monotherapy. 191 This effect was attributable to localized hydrogen peroxide generation within the alveolar lumen, which selectively increased the oxidative pressure on mycobacteria without amplifying tissue injury. 192 In addition, in models of multidrug-resistantEscherichia coli urinary tract infections, NAC enhances the efficacy of ciprofloxacin by reducing persister cell populations and improving antibiotic penetration across the urethral mucosa. 193 These findings underscore the importance of evaluating redox conditions and antioxidant bioavailability at infection sites to ascertain combinations that leverage the antimicrobial properties of antioxidants while circumventing antagonism with ROS-dependent drugs.
In patients suffering from critical illnesses or immunosuppression, systemic oxidative stress is frequently exacerbated. Antioxidant administration is therefore employed to protect target organs from inflammatory damage. 194 However, in this same population, antimicrobial selection must consider that some fungicidal or bactericidal drugs (e.g., liposomal amphotericin B, newer-generation fluoroquinolones) exert some of their effects through the generation of microenvironmental ROS. 195 In such cases, monitoring redox status via biomarkers, such as reduced/oxidized glutathione (GSH/GSSG) or serum malondialdehyde levels, can serve as a guide for the optimization of antioxidant doses and administration sequences. 196 This approach is designed to avert suboptimal levels of antibiotics. Additionally, liposomal formulations of certain drugs, such as amphotericin B, have been demonstrated to facilitate the release of the active agent within the phagosomes of macrophages. 197 In this environment, the local production of ROS has been shown to favor lipid ester activation and the generation of cytotoxic species. This process effectively minimizes the impact of systemic antioxidants.
The adaptation of antimicrobial therapy to a patient’s redox characteristics constitutes a component of precision medicine, a multifaceted field that integrates pharmacokinetics, pharmacodynamics, and nutritional status. The implementation of clinical guidelines that encompass recommendations concerning the temporal separation between the administration of antioxidants and antimicrobials, the selection of drugs whose efficacy is not predicated on ROS production, and the utilization of protective formulations (e.g., liposomes, nanoparticles) to preserve oxidative action at the site of infection has the potential to markedly improve clinical outcomes.
Gut–brain axis: Impact of exercise and nutrition on microbial–host interaction and infection outcomes
The gut‒brain axis is a bidirectional communication network that connects the gastrointestinal tract, including its microbiota, with the central nervous system. This connection involves neural pathways (for example, the vagus nerve), endocrine routes (stress hormones such as cortisol), and immune interactions (cytokines and microbial metabolites) that allow the gut and the brain to influence each other.198,199 The intestinal microbiota plays a crucial role in this communication by producing neurotransmitters (e.g., GABA, serotonin) and metabolites such as short-chain fatty acids that can modulate brain and immune function. In turn, psychological or neurological states—such as stress—can alter intestinal motility, mucus secretion, and epithelial permeability, affecting the habitat of microorganisms and host homeostasis.200,201 Under healthy conditions, the gut‒brain axis contributes to immune tolerance and inflammatory balance: a diverse and balanced microbiota educates the intestinal immune system and is associated with appropriate responses to pathogens while sending neurochemical signals that influence mood and neuronal function. In contrast, dysbiosis or microbial imbalance can have systemic consequences, including gastrointestinal and psychological disorders. Thus, microbe‒host interactions through the gut‒brain axis are important determinants of overall health. 202
Numerous studies have shown that deliberate alterations in the intestinal microbiota can impact the gut‒brain axis. For example, the administration of probiotics has been associated with improvements in both intestinal disorders and neuropsychiatric symptoms, underscoring the integrative nature of this axis. 203 At the molecular level, certain components of commensal bacteria can stimulate the vagus nerve or influence immune cells, modulating the release of cytokines that in turn affect brain function. In this way, the gut‒brain axis is a continuous feedback circuit: the microbiota influences the host (including the brain), and the host’s physiological state modulates the intestinal environment. 204
Influence of physical exercise on the gut‒brain axis
Regular physical exercise is recognized as a positive modulator of the intestinal microbiota and, therefore, of the gut‒brain axis. In general, moderate-intensity exercise has been shown to increase microbial diversity and the abundance of beneficial bacteria, particularly within the phylum Firmicutes. 205 This enrichment of the “friendly” microflora contributes to greater production of anti-inflammatory metabolites such as short-chain fatty acids, which strengthen the intestinal barrier and exert systemic neuromodulatory and anti-inflammatory effects. Additionally, exercise stimulates the secretion of immunoglobulin A in the gastrointestinal mucosa, reinforcing protection against pathogens in the intestinal lumen. These changes indicate that physical activity can train the intestinal immune system to keep infections at bay, acting through the microbiota and components of the gut‒brain axis. 206
Recent studies have explored the effects of exercise on microbiota-mediated infection resistance. Notably, moderate exercise can increase host resistance to colonization by multidrug-resistant pathogens. Xu et al. demonstrated in a murine model that moderate exercise significantly reduced intestinal colonization by methicillin-resistantStaphylococcus aureus (MRSA) compared with sedentary animals. This protective effect was attributed to specific changes in the microbiota: physical activity increased an intestinal probiotic strain (Dubosiella newyorkensis, strain L8) that hinders MRSA growth by depleting key nutrients such as fucose, which is necessary for the virulence of this pathogen. 34 This study exemplifies how exercise, which acts through the gut–microbiota axis, improves resistance to serious bacterial infections by strengthening host defenses. Likewise, regular exercise has been shown to decrease chronic systemic inflammation associated with dysbiosis, which could translate into better infection outcomes by preventing unbalanced immune responses. Therefore, exercise acts as a modulatory factor of the gut‒brain axis, promoting a favorable microbial and immune environment that can improve the host’s ability to prevent and control infections.40,207
Influence of nutrition on the dynamics of the gut–brain axis
Diet is one of the most powerful determinants of intestinal microbial composition and, therefore, of the signals flowing along the gut‒brain axis. Adequate and balanced nutrition (rich in dietary fiber, fermentables, and essential nutrients) tends to foster a diverse microbiota with a predominance of beneficial bacteria (for example, butyrate-producing genera such as Faecalibacterium), which strengthens intestinal barrier integrity and reduces systemic inflammation. In contrast, Western diets high in saturated fats and simple sugars have been linked to proinflammatory dysbiosis. Agus et al. (2016) reported that a fat/sugar-rich Western diet in mice induced an inflammatory intestinal environment with overgrowth of Proteobacteria (including pathogenic E. coli strains) and decreased the number of protective bacteria. This microbial change was associated with reduced concentrations of short-chain fatty acids (SCFAs) and lower expression of the intestinal SCFA receptor (GPR43). Consequently, the transfer of the microbiota from Western diet-fed mice to germ-free mice increased their susceptibility to infection by adherent-invasiveE. coli, demonstrating that an unhealthy diet alone can select for a microbiota that exacerbates vulnerability to enteric infections.208,209 Furthermore, SCFA deficiency and insufficient GPR43 activation under these conditions have been linked to dysregulated immune responses and a greater predisposition to colitis, illustrating how nutrition influences microbe‒host interactions at the intestinal mucosal level.
Host nutritional states, such as obesity, malnutrition, and fasting, also modify the gut‒brain axis and the response to pathogens. Obesity, for example, is often associated with dysbiosis with lower diversity and predominance of proinflammatory bacteria, along with a state of chronic low-grade inflammation in the host. This inflammatory environment can compromise intestinal barrier function and alter vagal signaling, affecting both neuroendocrine homeostasis and innate immunity. As a result, individuals with obesity may exhibit less efficient responses to certain infections and greater severity in outcomes (for example, increased risk of complications in respiratory viral infections), partly owing to this altered gut–brain–immune axis.210,211 On the other hand, malnutrition and micronutrient deficiencies compromise the development and maintenance of beneficial microbiota and weaken the intestinal barrier. Children with severe malnutrition exhibit immature and less diverse microbiota, which contributes to local immunosuppression, villous atrophy, and increased intestinal permeability. This creates a vicious cycle in which bacterial translocation and systemic inflammation worsen the clinical state. From the perspective of the gut‒brain axis, malnutrition can limit the production of neurotransmitters derived from microbes and alter enteric neural signaling, which even affects appetite and eating behaviors in a negative feedback loop.210,212
A special case of nutritional influence is fasting or acute caloric restriction, which triggers systemic metabolic changes (ketogenesis, increased ketone bodies) and modulates both the microbiota and the immune response. It was traditionally thought that a lack of intake during an infection could be harmful; however, recent evidence suggests that controlled fasting could have beneficial effects in certain contexts of gastrointestinal infection. A study in mice revealed that 48 hours of fasting before and during oral infection with Salmonella enterica serotype Typhimurium almost completely suppressed bacterial invasion of the intestinal mucosa and attenuated the intestinal inflammatory response.213,214 This effect was due to fasting inhibiting the expression of virulence genes (SPI-1) in Salmonella through microbiota-mediated changes in the luminal environment, preventing the pathogen from crossing the epithelium. In gnotobiotic mice without a microbiota, a protective effect of fasting was not observed, underscoring the importance of the microbial community in mediating how a lack of food affects the host‒pathogen‒microbiota triad. Furthermore, even when Salmonella virulence was restored in the absence of microbiota, fasting induced a reduction in the host’s proinflammatory signals, reducing tissue damage. These findings indicate that the anorexigenic response during infection (loss of appetite mediated by the brain) could be an evolutionary mechanism beneficial to the host, modulating the gut‒brain axis in a way that favors the commensal microbiota and hinders the establishment of invasive pathogens. 215
Consequences for susceptibility and infection outcomes
Modulation of the gut‒brain axis through exercise and nutrition has direct implications for how the body deals with infections. In general, a balanced gut‒brain axis, supported by diverse microbiota, regular physical activity, and a healthy diet, tends to prepare the immune system to respond efficiently to pathogens without incurring excessive inflammation. For example, physically active individuals not only present lower baseline inflammation but also may exhibit a lower incidence and severity of certain gastrointestinal infections, possibly owing to a robust microbiota and greater intestinal IgA. Likewise, high-fiber diets that promote butyrate production have been associated with lower susceptibility to infections by Clostridioides difficile or other enteric pathogens, since SCFAs strengthen epithelial tight junctions and energize colonocytes, increasing resistance to colonization. 216
In contrast, disruptions in the gut‒brain axis, whether due to sedentarism, inadequate diet, or chronic stress, can predispose individuals to worse infectious outcomes. Psychological stress, in particular, has been implicated in increased risk of intestinal infections: brain–gut axis–mediated mechanisms, such as the release of catecholamines and cortisol, can alter microbial composition and increase the permeability of the gut. It has been shown that social stress in mice induces thinning of the mucus layer and increased expression of proinflammatory genes in epithelial cells, accompanied by evidence of bacterial translocation (i.e.,bacteria crossing the intestinal barrier). These changes, which are dependent on the presence of microbiota, suggest that stress primes intestinal cells into a pro-oxidative and pro-inflammatory state that could facilitate infections or chronic inflammation. Such a predisposition under stress translates clinically into a greater incidence of diseases such as colitis or gastroenteritis in people subjected to prolonged stress, confirming that the neuroimmune state influences pathogen interactions.217,218
Thus, the gut‒brain axis acts as an integrative bridge where lifestyle factors (exercise, diet, stress) and intestinal microbiology converge to determine the host’s susceptibility to infections and their outcomes. Maintaining a balanced microbiota through adequate nutrition and physical activity can strengthen innate and adaptive defenses, optimizing microbe‒host interactions in favor of the organism. On the other hand, ignoring these aspects can destabilize this axis, decreasing resistance to pathogens or exacerbating inflammation during infection. Understanding these links opens opportunities for adjunctive interventions in infectious diseases—for example, exercise programmes in vulnerable populations or modulation of diet and microbiota—taking advantage of the plasticity of the gut–brain axis to improve host health.
Designing prodrugs and nanocarriers for nutritional contexts (fasting, obesity, etc.)
The fasting state significantly affects the pharmacokinetics (PK) of many drugs, mainly by altering oral absorption and presystemic metabolism. In the absence of food, biliary and pancreatic secretion decreases, gastrointestinal pH changes, and gastric emptying accelerates, which can reduce the dissolution and absorption of lipophilic or slow-release drugs. 219 For example, some drugs show much lower bioavailability during fasting because they lack the stimulus of dietary fats necessary for their solubilization; this fasted vs. fed variability phenomenon complicates optimal dosing. Additionally, prolonged fasting induces metabolic adaptations such as ketogenesis and changes in hepatic blood flow that can modify the rate of drug metabolism in some cases, reducing hepatic clearance due to lower enzymatic activity or portal flow.219,220 Pharmacodynamically, fasting can increase sensitivity to certain drugs (e.g., sedatives or antihypertensives) owing to lower energy reserves and changes in receptor distribution.
To address these variations, pharmaceutical scientists are exploring the design of prodrugs and delivery systems that minimize dependence on food intake. One approach is to develop prodrugs that improve aqueous solubility, for example, by conjugating the active drug with hydrophilic groups or promoieties that allow efficient absorption even in the fasting state. These promoieties can be designed to be cleaved by intestinal or hepatic enzymes regardless of the presence of food. In this way, the active drug is released in a controlled manner after absorption, avoiding peaks or abrupt drops associated with the fed state. Similarly, lipid-based nanocarriers (such as solid lipid nanoparticles or self-emulsifying systems) are used to simulate the solubilizing effect of a meal. These vehicles contain digestible lipids that, when administered with the drug, stimulate bile secretion and form fine microemulsions that solubilize the active ingredient in the patient’s GI tract during fasting.221,222 The evidence indicates that lipid nanoparticles reduce pharmacokinetic variability between fed and fasted states, providing more consistent absorption profiles. For example, compared with conventional oral formulations, nanoemulsion formulations of poorly soluble drugs (such as certain antivirals) have shown >2-fold increases in bioavailability during fasting and smaller differences in bioavailability compared with the postprandial state. In addition, micellar mixtures with phospholipids (e.g., phosphatidylcholine micelles) have been developed to encapsulate the drug and facilitate its absorption even without food, a strategy that has reduced interindividual variability in the systemic exposure of molecules such as vilazodone between fasting and fed subjects. 223
Another tactic is the design of modified-release systems that retain the drug until it reaches regions of the intestine where conditions are more stable between fasting and feeding. For example, capsules with enteric coatings or gastroretentive floating devices can prevent rapid gastric evacuation during fasting, releasing the drug gradually and improving absorption. The use of pH-sensitive polymers is also being investigated to adjust drug release according to the luminal environment: in the absence of food, when the intestinal pH is relatively high, the polymer can swell and release more slowly, whereas in the postprandial state with a relatively high buffering pH, release is faster. These innovations aim to mitigate the need to synchronize medication with meals, increasing the safety and efficacy of drugs in patients who, by necessity or medical indications, must remain in intermittent or prolonged fasting.224,225
Pharmacological alterations associated with obesity and design strategies
Obesity involves profound physiological changes that influence drug pharmacokinetics, affecting all phases: absorption, distribution, metabolism, and excretion. In individuals with obesity, oral absorption may be modified by accelerated gastrointestinal transit and faster-than-normal gastric emptying, potentially reducing the absorption of prolonged-release formulations or drugs absorbed in specific intestinal segments. Distribution is particularly altered by the increase in adipose mass: lipophilic drugs tend to have a markedly greater volume of distribution in obese people, as they accumulate in fatty tissue, prolonging their half-life. In contrast, hydrophilic drugs show moderate or minimal increases in volume of distribution, remaining mainly in aqueous compartments. 226 In addition, obesity is often associated with nonalcoholic fatty liver and increased kidney size, factors that can alter metabolism and clearance: some hepatic enzymes (CYP450s) may have increased or reduced activity depending on the isoform, and hepatorenal blood flow is increased owing to increased cardiac output. Overall, evidence suggests that metabolism and elimination in obese individuals may be similar or slightly lower per kilogram of body weight than in nonobese individuals, implying that dosing on the basis solely of total body weight can lead to overdosing. All these alterations highlight the need to consider obesity in the design of personalized pharmacological therapies.
To optimize pharmacotherapy in obesity contexts, prodrugs and nanocarriers are being developed that are specifically targeted to adipose tissues or adapted to the metabolic environment of obesity. 227 An innovative approach is the use of “lipidomimetic” prodrugs designed to exploit dietary lipid absorption pathways and direct drugs to fat tissue. Mattarei et al. (2017) presented a strategy of linking an active compound (pterostilbene, with antiobesity potential) to a synthetic triglyceride glycerol molecule, creating a prodrug that mimics the structure of a triglyceride. This prodrug, administered orally, is processed by pancreatic lipases as if it were dietary fat, preferentially entering the intestinal lymphatic circulation in the form of chylomicrons. As a result, the drug travels directly to fat deposits where it is locally released after hydrolysis, achieving high concentrations in adipose tissue and minimizing unwanted systemic exposure. This adipocyte-targeted delivery method demonstrates the feasibility of designing prodrugs that activate specifically in the target organ affected by obesity, improving efficacy (by delivering the drug “where it is needed”) and reducing systemic adverse effects. 228
In parallel, nanocarrier systems offer solutions to address the particularities of distribution in obesity. For example, polymeric nanoparticles or functionalized liposomes can be used to modulate tissue distribution: researchers are exploring the possibility of decorating the nanoparticle surface with ligands that recognize receptors overexpressed in adipose tissue or in organs affected by obesity comorbidities (such as the liver in fatty liver). In this way, after systemic administration, the nanocarrier preferentially accumulated the drug in those tissues. Likewise, in cases where obesity reduces the free fraction of a drug owing to high adiposity or altered plasma proteins, nanocarriers can serve as “reservoirs” that release the drug in a controlled manner, maintaining stable therapeutic concentrations. A concrete example is the use of nanoemulsions for oncological drugs in obese patients: studies suggest that nanoparticulate formulations of chemotherapeutics achieve more uniform tumor tissue penetration and reduce uptake by adipose tissue than conventional formulations do, avoiding tumor underdosing.229,230
In addition, obesity is often accompanied by chronic inflammation and changes in the tissue microenvironment (slightly acidic pH in inflamed adipose tissue, local hypoxia). This makes it possible to conceive prodrugs that are activatable by stimuli characteristic of obesity. For example, a prodrug could be designed whose activation (release of the active drug) occurs through the action of inflammatory enzymes elevated in obesity (e.g., metalloproteinases) or by the lower pH in dysfunctional adipocytes. Similarly, nanocarriers sensitive to pH or reactive oxygen species could release more drugs in inflamed environments typical of obesity, increasing therapy specificity. These smart design approaches aim to ensure that obese patients receive the right effective dose, overcoming altered PK challenges, while also directing therapy to avoid cumulative toxicity. 231
Implications of malnutrition in drug disposition
Protein-caloric malnutrition and profound nutritional deficiencies cause pathophysiological changes that markedly affect pharmacokinetics and pharmacodynamics. In severe malnutrition, there is a decrease in lean body mass, organ tissues (liver, kidney, and musculature), and plasma proteins, especially albumin. As a consequence, many drugs present a reduced volume of distribution (due to lower muscle and fat mass in cases of emaciation) and an increased free fraction due to hypoalbuminemia. The latter implies that a greater proportion of the drug remains unbound in the plasma, enhancing its effect or toxicity at normal doses. For example, in children with severe malnutrition, a higher incidence of digoxin and anticonvulsant toxicity has been documented, attributed partly to low albumin and electrolyte alterations.232,233 In terms of metabolism, malnutrition reduces the expression and activity of various phase I and II hepatic enzymes, as well as transporters, due to hepatic atrophy and vitamin cofactor deficiencies. This can prolong the elimination half-life of drugs that are normally cleared quickly, requiring dose or interval adjustments. In addition, renal perfusion can also decrease malnutrition, reducing glomerular filtration; in fact, studies indicate that in malnourished patients, renal clearance of aminoglycoside antibiotics is reduced, increasing the risk of accumulation.
In light of these alterations, the design of pharmaceutical interventions for malnourished patients seeks to achieve a balance between efficacy and safety adapted to their physiology. A crucial strategy is to develop prodrugs that do not depend on intense hepatic metabolism for activation. For example, instead of a drug requiring conversion by CYP450 (which is potentially deficient in malnutrition), one can opt for a prodrug activated by gastrointestinal or blood enzymes that are more preserved or even by slow chemical hydrolysis that does not require enzymes.232,233 This ensures that the malnourished patient can sufficiently convert the prodrug into the active drug. Relatedly, prodrugs that release toxic byproducts that the weakened body cannot adequately clear should be avoided. The emphasis in malnutrition contexts is on metabolic simplicity: molecules whose PK profile is predictable even with low metabolic capacity. Additionally, controlled-release systems can be beneficial for preventing high plasma concentration peaks in susceptible patients. For example, formulating a drug in a sustained-release nanoparticle (e.g., a biodegradable polymeric matrix) can maintain more stable levels and prevent acute toxicity from high free plasma concentrations. These formulations can also protect the drug from an adverse gastrointestinal environment common in malnutrition (altered pH, dysbiotic microbiota) until it reaches the optimal absorption site. 234
Another design approach focuses on combining nutritional therapy with drug delivery through nanocarriers. Since malnutrition is often accompanied by vitamin and mineral deficiencies that affect pharmacodynamics (e.g., zinc deficiency compromises immunity or vitamin K deficiency alters coagulation and the response to anticoagulants), nanocapsules are being explored for their ability to coencapsulate essential nutrients along with drugs. In this way, the nanocarrier could gradually overcome certain deficiencies while releasing the therapeutic drug, improving the patient’s overall response. A conceptual example is a lipid nanoparticle carrying both vitamin D and an antibiotic for children with malnutrition and tuberculosis, improving the absorption of both and boosting the child’s immune system. Similarly, in malabsorption situations associated with malnutrition, nanovehicles can facilitate transcellular drug transport through the damaged intestinal wall. Some nanoparticles (e.g., mucoadhesive polymers) can prolong contact with the atrophic mucosa, increasing the absorbed drug fraction even when villi are diminished. 235
From a pharmacodynamic perspective, malnutrition often increases drug sensitivity (due to receptor alterations and general frailty). Therefore, targeted and slow-release formulations are doubly useful: they not only ensure that enough drug reaches the therapeutic target but also prevent excessive doses from impacting other vulnerable tissues. For example, in malnourished animal models, a standard dose of a nonsteroidal anti-inflammatory drug can cause severe lesions; however, a nanoparticulate formulation with targeted colonic release minimizes systemic absorption at nontarget sites, reducing gastric and renal damage. This finding illustrates how adapting pharmaceutics to the nutritional context improves tolerability.
Integrated design proposals and case studies
The development of prodrugs and nanocarriers adapted to specific nutritional contexts benefits from an interdisciplinary approach that considers molecular and physiological changes in these states. An emerging general principle is to formulate with knowledge of the context: for example, in patients alternating between fasting and feeding periods (intermittent fasting), an “intelligent” system could be devised to release more drug during fasting hours and reduce release after meals to compensate for absorption variability. This could be achieved with an enzyme-sensitive coating whose level fluctuates with intake, releasing the drug preferably when such enzymes (associated with feeding) are low. In obesity, an interesting concept is to employ sequestrant therapies: nanopolymers that trap endogenous molecules altered by obesity (for example, excess proinflammatory bile acids) in combination with the release of an anti-inflammatory drug.235,236 A relevant case study is the development of oral polymeric resins that bind bile acids in obese patients, reducing metabolic inflammation and improving insulin sensitivity; integrating an antidiabetic agent into such resins could simultaneously treat the cause and the symptoms.
In the context of malnutrition, clinical trials with adapted formulations are still scarce, but an illustrative case is ready-to-use therapeutic foods (RUTFs) enriched with antiparasitic or antibiotic drugs. These nutritional pastes designed for the rehabilitation of children with severe malnutrition have incorporated drugs uniformly, ensuring caloric intake along with treatment. Although not “nanotechnological,” they represent an integrated drug‒nutrition approach that has improved adherence and clinical response in malnourished populations. 237 In the future, we could see edible nanoparticles included in therapeutic foods that carry antimicrobial agents that are gradually released as the child gains weight, thus synchronizing nutritional recovery with infection eradication.
Therefore, personalizing the design of prodrugs and nanocarriers according to the patient’s nutritional context promises to optimize the efficacy of pharmacological treatments. Fasting, obesity, and malnutrition profoundly affect drug disposition; ignoring these factors can lead to undertreatment or toxicity. Through strategies such as prodrugs activatable by alternative routes, tissue-targeted nanocarriers, or formulations mitigating food-related variability, therapy can be adapted to patient needs. The case studies mentioned demonstrate tangible improvements—from better bioavailability in fasting to selective delivery in adipose tissue—indicating that these innovations are feasible. Future research should strengthen the safety of these systems in vulnerable populations and establish clear dosing guidelines. Integrating knowledge from pharmacology, nutrition, and materials science will be key to designing the next generation of “nutrition-sensitive” treatments, ensuring that patients receive the right dose in the right place at the right time, regardless of their dietary or nutritional state. 238
Role of exercise-induced inflammation in antimicrobial drug efficacy and resistance mechanisms
Acute and chronic forms of physical exercise, particularly those involving prolonged or high-intensity efforts or long durations, elicit a systemic inflammatory response characterized by increased circulating cytokines (e.g., IL-6, TNF-α), activation of immune cells, and elevated expression of endothelial adhesion molecules.239,240 While this type of inflammation is typically transient and part of a coordinated physiological adaptation, its magnitude and duration can vary markedly depending on the exercise modality. However, well-regulated exercise is associated with anti-inflammatory effects, promoting a systemic environment that counterbalances chronic low-grade inflammation. 241 These effects are mediated both by a reduction in visceral fat mass, with a subsequent decrease in pro-inflammatory adipokine release, and by the acute induction of anti-inflammatory mediators (e.g., IL-10 and IL-1ra) with each exercise bout.
Moreover, well-regulated exercise-induced inflammation can meaningfully influence the pharmacokinetics and pharmacodynamics of antimicrobial drugs. Exercise alters regional blood flow, modulates hepatic and renal clearance, and affects gastrointestinal absorption, all of which are key determinants of drug bioavailability, metabolism, and excretion. 242 For example, splanchnic blood flow can be reduced by up to 80% during intense exercise, 243 delaying gastric emptying and modifying drug absorption rates. Exercise also decreases renal blood flow and hepatic perfusion, which may increase plasma concentrations and alter the elimination kinetics of drugs predominantly cleared by the kidneys or liver. 244 These physiological shifts can affect antimicrobial plasma levels and half-lives, with potential implications for both efficacy and toxicity. 245 Although direct evidence for antimicrobials is limited, the mechanisms described suggest that cytokine-mediated modulation of cytochrome P450 activity and drug transporters (e.g., P-glycoprotein)242,243 may contribute to altered drug disposition during or after exercise.
Exercise-induced inflammatory responses can significantly affect the tissue-specific distribution and local bioavailability of antimicrobial agents. 242 Inflammation-driven changes in capillary permeability, plasma protein binding, and immune cell trafficking affect drug delivery and efficacy. 246 These changes may enhance immune‒drug synergy or, conversely, hinder antimicrobial action in poorly perfused or edematous tissues. Elevated acute-phase proteins can reduce the free fraction of active drug by increasing plasma binding, thereby limiting its availability at infection sites.246–248 Understanding these dynamics is key for optimizing dosing in physically active or recovering patients.
This inflammatory milieu also modulates hepatic and intestinal expression of drug-metabolizing enzymes and transporters, notably cytochrome P450 isoforms and P-glycoprotein249. For antibiotics with narrow therapeutic windows or high hepatic clearance, such modulation may enhance or impair clinical efficacy. Moreover, transient increases in vascular permeability during exercise may facilitate drug penetration into infected tissues, which is potentially beneficial in acute infections.
From a resistance perspective, chronic low-grade inflammation, as observed in overtrained athletes or individuals exposed to sustained high-intensity training, may promote conditions favorable for antimicrobial resistance. 250 Mechanisms include persistent immune activation, microbiota dysbiosis, and impaired mucosal barriers. Inflammatory oxidative stress can also increase horizontal gene transfer via plasmid mobilization, accelerating resistance gene dissemination. 251
In conclusion, exercise-induced inflammation has dual effects: acutely enhancing immune‒drug interactions and tissue penetration but potentially facilitating resistance under chronic or excessive conditions. These effects highlight the need for individualized exercise prescriptions in patients receiving antimicrobial therapy.
Clinical evidence linking lifestyle interventions with improved antimicrobial outcomes
Emerging clinical and translational evidence suggests that integrated lifestyle interventions, particularly structured physical activity and dietary modifications, can increase antimicrobial efficacy and reduce the risk of treatment failure or relapse. 133 These effects are mediated through improvements in immune competence, metabolic regulation, and the gut microbial composition, all of which intersect with drug action and host‒pathogen dynamics.252,253
Specifically, moderate aerobic exercise has been shown to improve immune surveillance and vaccine responsiveness, especially in older adults, suggesting an increased capacity for pathogen clearance. 254 For example, in patients with chronic respiratory infections—such as bronchiectasis or pneumonia—structured exercise interventions (e.g., pulmonary rehabilitation) have demonstrated improved antibiotic efficacy, faster resolution of symptoms, and reduced systemic inflammation when combined with nutritional support. 255 These findings point to a synergistic effect between physical activity256,257 and host-directed therapies in infections.
Moreover, lifestyle interventions also modulate the composition of the gut microbiota, which plays a critical role in mucosal immunity and the metabolism of orally administered antibiotics. 258 Regular physical activity enhances microbial diversity and may contribute to the abundance of short-chain fatty acid–producing taxa, such as Faecalibacterium and Roseburia, which support epithelial integrity and suppress opportunistic pathogens. 8 This microbiota-mediated resilience may mitigate dysbiosis-related resistance mechanisms, including impaired drug absorption and altered transporter expression.
Furthermore, metabolic status and systemic inflammation, both of which are influenced by lifestyle, directly affect antimicrobial pharmacokinetics.119,259 Obesity, for example, alters the volume of distribution and clearance of lipophilic antibiotics, frequently resulting in underdosing and therapeutic failure. 260 Weight loss and improved glycemic control can normalize these pharmacokinetic parameters, increasing drug-target engagement and potentially reducing the risk of resistance. 261
Despite encouraging observational and interventional data, randomized controlled trials specifically designed to evaluate lifestyle and antimicrobial interactions remain scarce. Future studies should stratify participants by infection type, comorbidity profile, and baseline functional status to identify causal mechanisms and define optimal cointervention strategies.
Practical applications, future research and limitations
This review has several limitations that should be acknowledged. First, the narrative and integrative design, while appropriate for addressing a complex and multidisciplinary topic, does not follow the strict methodological framework of a systematic review, which may limit reproducibility and introduce a degree of subjectivity in study selection and interpretation. Second, the possibility of publication bias cannot be excluded, as studies reporting significant or positive findings are more likely to be published and included. Third, the included studies are characterized by considerable heterogeneity in terms of design, population (human vs. animal models), and methodological approaches, particularly in microbiome and pharmacokinetic assessments. This variability limits direct comparability and the ability to draw definitive conclusions. Fourth, in keeping with its narrative nature, no formal standardized quality appraisal or risk-of-bias assessment of the included studies was performed, and study selection was carried out by a single reviewer; consequently, the potential for selection bias and for the subjective weighting of evidence cannot be ruled out. Fifth, a substantial proportion of the mechanistic claims discussed—particularly those concerning microbiome-mediated resistance pathways and horizontal gene transfer—are based on preclinical, in vitro, or associative data, and should therefore be regarded as emerging or hypothetical rather than causally established. Therefore, the findings of this review should be interpreted with caution, and further well-designed, controlled studies are needed to strengthen the evidence base in this emerging field. More specifically, future priorities include: • Personalized prescribing – Incorporating physical activity level, body composition, and nutritional status into antibiotic dosing and selection. • Integration into stewardship programs – Embedding exercise and nutritional interventions alongside pharmacological treatment to reduce relapse and treatment failure • Drug design innovation – Development of prodrugs, nanocarriers, and pH/ROS-sensitive formulations tailored to metabolic conditions such as obesity or malnutrition • Clinical monitoring – Using wearable devices and digital health data to adjust dosing on the basis of activity patterns and metabolic fluctuations • Public health strategies – Promotion of healthy lifestyle habits as complementary measures to slow resistance development and enhance host resilience. • Lifestyle-informed prescribing – Incorporating lifestyle factors (diet, physical activity, metabolic status) into antimicrobial decision-making to optimize efficacy and reduce variability in drug response • Dietary co-management– Providing targeted dietary guidance during antimicrobial therapy to prevent food–drug interactions and ensure adequate drug absorption and exposure
The relationship between physical activity, nutrition, and antimicrobial pharmacology spans diverse physiological and biochemical systems that cannot yet be captured through uniform experimental models. Nonetheless, the existing evidence is heterogeneous in scope and methodology, and many findings remain preliminary or preclinical. These factors should be considered when concluding research in clinical contexts.
Future studies should integrate exercise physiology, nutritional science, and pharmacology within unified experimental and clinical frameworks. Multidimensional approaches combining metabolomics, microbiome analysis, and pharmacokinetics will be essential to clarify how lifestyle factors shape antimicrobial efficacy and resistance. Translational and precision medicine perspectives are particularly promising, aiming to design therapeutic strategies and novel drug compounds responsive to individual metabolic and behavioral profiles.
Conclusion
Lifestyle factors, especially diet and physical activity, have significant effects on antimicrobial pharmacology, shaping microbiome composition, immune competence, and drug kinetics. Neglecting these influences may compromise therapeutic efficacy and contribute to resistance development. Current evidence suggests that appropriately prescribed moderate physical activity and nutritional optimization can support antimicrobial therapy by improving host response and reducing variability in drug exposure.
For drug development, these findings highlight the importance of designing formulations that are robust to metabolic variability and of incorporating host-related determinants into pharmacological evaluation. Emerging strategies, such as prodrugs and nanocarrier-based systems, may offer potential for more personalized antimicrobial delivery; however, their application in lifestyle-adapted therapy remains largely conceptual. Advances in stimuli-responsive systems (e.g., pH- or temperature-sensitive delivery platforms) suggest possible mechanisms through which drug release could be influenced by physiological changes associated with physical activity, although direct clinical evidence is still limited.
Clinically, an integrative model combining pharmacology with lifestyle medicine may contribute to more context-sensitive antimicrobial strategies, supporting precision medicine approaches and helping to mitigate the global antimicrobial resistance (AMR) challenge.
Footnotes
Acknowledgements
The authors have no acknowledgements to disclose.
Ethical considerations
This article does not contain any studies with human participants or animals performed by any of the authors.
Author contributions
Conceptualization, V.J.C.-S.; methodology, J.F.T.-A. and A.I.B.-V.; investigation, A.R.-Z., A.M.-R.; writing—original draft preparation, all authors; writing—review and editing, all authors; visualization, all authors; supervision, V.J.C.-S.; project administration, V.J.C.-S. All authors have read and agreed to the published version of the manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
{kind=link}