
Abstract
Stem cells may be manipulated in vitro to induce hepatic differentiation. We investigated the effect of hypoxia and photobiomodulation therapy (PBMT) on the hepatogenic differentiation of human adipose-derived stem cells (hASCs). hASCs were exposed to different carbon dioxide concentrations with photobiomodulation (PBM) using low-level light. Cell survival and secretion of hepatocyte growth factor (HGF) of the hASCs were evaluated by immunostaining and Western blot analyses. Hepatic differentiation was assessed via immunocytochemical staining, fluorescence-activated cell sorting, and Western blot analysis for liver-specific genes and proteins, including albumin (ALB), cytokeratins 8/18, and alpha-fetoprotein (AFP). PBM therapy has been shown to enhance proliferation and cytokine secretion of a number of cells. The expression profiles of ALB, AFP, and cytokeratin 8/18 demonstrated that when HGF, hypoxia, or PBMT were treated individually, incomplete hepatocyte differentiation was achieved. In contrast, quantitative analysis of ALB, cytokeratins 8/18, and AFP showed that HGF was enhanced significantly by hypoxia+PBM treatment. The obtained cell populations contained progenitors that expressed both hepatic ALB and cytokeratin 8/18 markers, as well as AFP. These data suggest that PBMT and hypoxia are effective biostimulators of hASCs in hepatogenic differentiation, which enhances the survival of hASCs and stimulates the secretion of growth factors.
Introduction
Because the availability of suitable human livers for transplantation falls short of the number of potential patients, liver failure is a global medical problem (Becker and Elias, 2007; Rauchfuss et al., 2016). In addition, the availability of primary human hepatocytes for cell therapy and drug development applications is significantly limited (Goldman and Gouon-Evans, 2016; Kopp et al., 2016; Takebe et al., 2017; Tsolaki and Yannaki, 2015; Kerkhove et al., 2004). Thus, there is a compelling need for an abundant and accessible supply of cells that can safely and effectively improve liver function in these patients (Becker and Elias, 2007; Rauchfuss et al., 2016). Recent findings show that mesenchymal stromal cell (MSC) can differentiate into a cell exhibiting a hepatocyte phenotype (Aurich et al., 2009; Bhogal et al., 2011).
The photobiomodulation (PBM) has been implemented for various purposes for some time, such as to provide pain relief, to reduce inflammation, and to improve local circulation. Moreover, many studies have demonstrated that PBM has positive biostimulatory effects on stem cells (Choi et al., 2013). It is well known that the proliferation and growth factor secretion (such as hepatocyte growth factor [HGF], fibroblast growth factor [FGF], and vascular endothelial growth factor [VEGF]) of human adipose-derived stem cells (hASCs) was also enhanced by PBM irradiation (Choi et al., 2013; Park et al., 2014).
Many studies have indicated that hypoxia influences the development of the embryo by regulating the differentiation and self-renewal (including the maintenance of stemness and proliferation) of stem cells. In some organs, such as the lung, nervous system, and heart, hypoxia induces the differentiation of stem cells into mature cells (Park et al., 2013a). Hypoxic preconditioning has been reported to enhance the survival of human mesenchymal stem cells (Bhang et al., 2011). In addition, hypoxia stimulates the production of growth factors, such as HGF, endothelial growth factor, and bFGF that induce hepatocyte differentiation (Park et al., 2013a).
This study was designed to further optimize the conditions of differentiation in an effort to produce hASC-derived hepatocytes that closely recapitulate the human hepatic phenotype.
Materials and Methods
Culture of hASCs
hASCs supplied from CEFO (Seoul, South Korea) were cultured in low-glucose Dulbecco’s modified Eagle’s medium F-12 (Welgene, South Korea) supplemented with 10% fetal bovine serum (Welgene), 100 units/mL penicillin, and 100 µg/mL streptomycin at 37.0°C in a 5% CO2 incubator. hASCs between passages 5 and 8 were used for all experiments. Adherent cells obtained from human adipose tissue were expanded in vitro. The cells were positive for human MSC markers CD29 (β1 integrin), CD90 (Thy-1), and CD105 (endoglin). However, the cells were negative for human endothelial cell markers CD34, CD31, KDR (VEGF receptor) and hematopoietic cell marker CD45 on flow cytometry analyses (Supplementary Fig. S1). These results indicated that the expanded cells included a large population of hASCs and were not contaminated with endothelial cells.
In vitro hepatic differentiation
hASCs were plated on collagen type I-coated dishes at a concentration of 4 × 104 cells/cm2. When the cells reached confluency, hepatogenic induction was carried out over a period of 21 days under normoxic (21% oxygen) or hypoxic (5% oxygen) conditions. First, the cells were treated for 7 days with hepatogenic differentiation basal medium (HDBM) serum free supplemented with EGF and bFGF (PeproTech EC, UK). Afterwards, the cells were cultured for 14 days in an HDBM, containing HGF, FGF, and nicotinamide (Supplementary Table S2).
PBM irradiation
Light-emitting diode (LED; WON Technology, Daejeon, South Korea) was applied for 10 minutes daily (Table 1). The light source used was an LED (WON Technology Co., Ltd., South Korea) designed to fit over a standard multiwell plate (12.5 × 8.5 cm) for cell culture. The LED had an emission wavelength peaked at 660 nm. The irradiance at the surface of the cell monolayer was measured by a power meter (Orion, Ophir Optronics Ltd., UT). To obtain the energy dose of 6 J/cm2, the exposure time for LED array was 10 minutes under a power density of 10 mW/cm2 (1 milliwatt × second = 0.001 joules).
Dosimetric Parameters PBM Infrared
LED, light-emitting diode; PBM, photobiomodulation.
Cell viability assay
The cell viability of spheroids was analyzed by using a live/dead viability cytotoxicity assay kit after 10 days in culture (Molecular Probes, CA). Briefly, 1 mL of HEPES-buffered saline solution containing 2 µL of green (SYTO 10 fluorescent nucleic acid stain solution) and 2 µL of red (ethidium homodimer-2) nucleic acid stain solution was added to plates. The percentage of live/dead cells was scored by counting number of pixels per image.
Fluorescence-activated cell sorting
Cells were washed with phosphate-buffered saline (PBS) containing 0.5% bovine serum albumin (BSA; Sigma Aldrich, MO). The cells were stained in PBS containing 1% BSA with either isotype controls or antigen-specific antibodies for 60 minutes. The cells were washed thrice with PBS containing 0.5% BSA and resuspended in PBS for flow cytometry using an Accuri device (BD Biosciences).
Immunofluorescence staining
Indirect immunofluorescence staining was performed using a standard procedure. In brief, cells were fixed with 4% paraformaldehyde, blocked with 5% BSA/PBS (1 hour, 24°C), washed twice with PBS, treated with 0.1% Triton X-100/PBS for 1 minute, and washed extensively in PBS. The sections were stained with specific primary antibodies and fluorescent-conjugated secondary antibodies (Supplementary Table S1). The cells were counterstained with 4,6-diamino-2-phenylindole dihydrochloride (Vector Laboratories). Processed images were analyzed for fluorescence intensity using ImageJ software (NIH).
Western blot
Samples were solubilized in lysis buffer for 1 hour at 4°C. Proteins were separated by Sodium dodecyl sulfate polyacrylamide gel electrophoresis using 10% or 15% resolving gels followed by transfer to nitrocellulose membranes (Bio-Rad, CA). The membranes were incubated with the primary antibody for 1 hour at room temperature. For detection, peroxidase-conjugated anti-mouse IgG or anti-rabbit IgG and enhanced chemiluminescence (Amersham Pharmacia Biotech, NJ) were used as described by the manufacturer. Membranes were scanned to create chemiluminescent images and to quantify with an image analyzer (Kodak, NY).
Statistical analyses
All the quantitative results were obtained from triplicate samples. Data were expressed as a mean ± standard deviation. Statistical analysis was carried out using a two-sample t test for comparing two groups of samples and a one-way analysis of variance for three groups. A value of p < 0.05 was considered to be statistically significant.
Results
Inhibition of hASC apoptosis
To verify the cell viability of hASCs cultured as hypoxia and PBM, a dead cell assay of fluorescent dyes was carried out. Nonviable cells were stained red. Dead cells were significantly less (p < 0.05) in hypoxia- and PBM-treated hASC as opposed to a hypoxia culture (Fig. 1A). This indicated that apoptosis induced by a lack of cell–matrix interaction (anoikis) in vitro culture environment was prevented in hASCs cultured as hypoxia and PBM. Furthermore, hASCs had reduced proapoptotic factors Poly(ADP-ribose)polymerase-1, Bcl-2-associated X protein (PARP, Caspase-3, and BAX) and enhanced antiapoptotic factor B-cell lymphoma 2 and Phosphorylated AKT (Bcl-2 and pAKT) at the protein level (Fig. 1B). This indicated that hypoxia- and PBM-treated hASC secrete higher levels of antiapoptotic factors relative to hASCs in normoxia culture. Therefore, PBM–hypoxia-cultured cells seemed to be more adaptable and more resistant to hypoxia compared with hASCs in normal cell culture conditions.
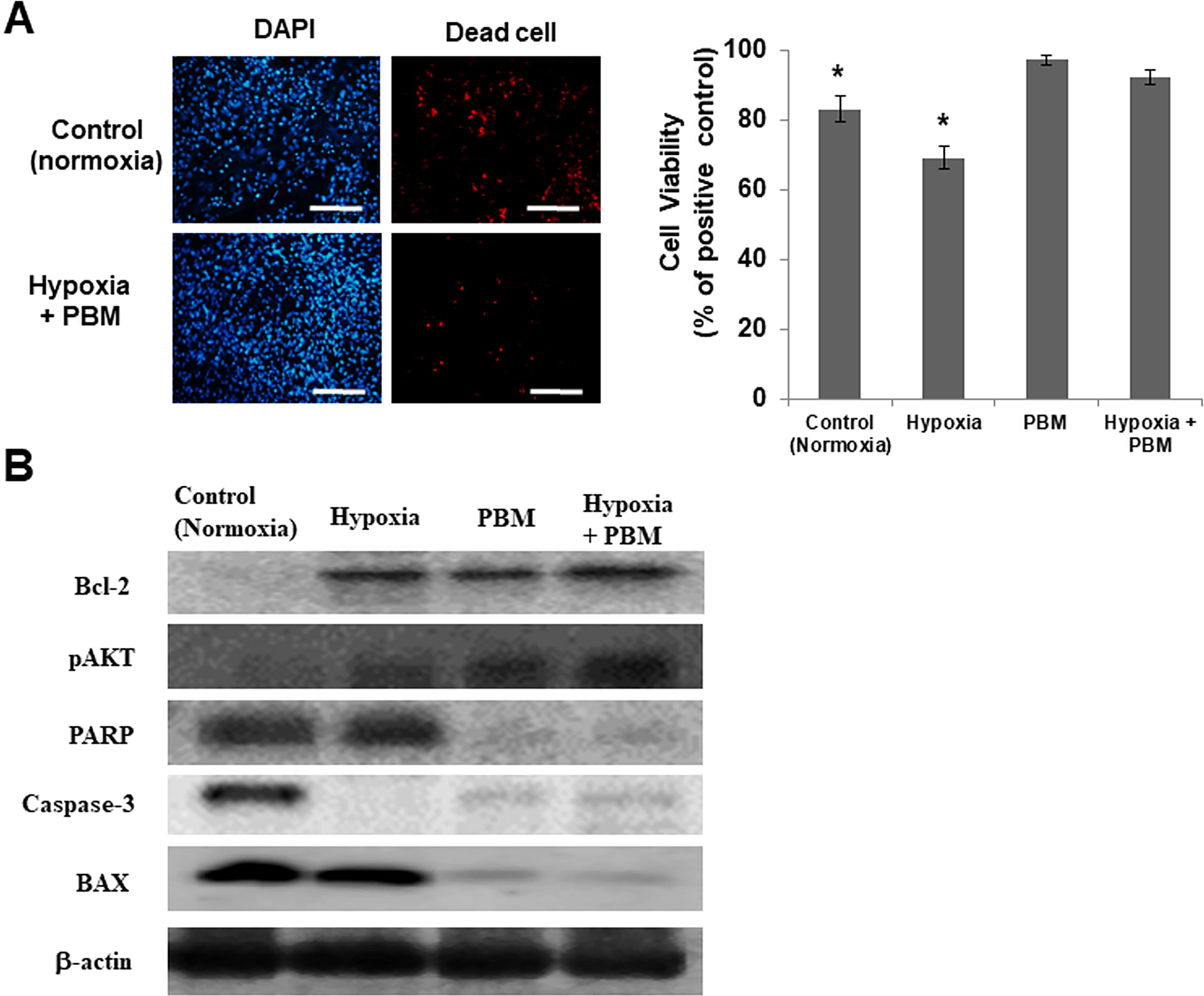
Inhibition of apoptosis in hASCs by PBM–hypoxia. hASCs were cultured for 21 days under normoxic (21% oxygen) or hypoxic (5% oxygen) conditions.
Production of growth factors by cells in PBM–hypoxia culture
Hypoxia condition was confirmed by the elevated expression of hypoxia-inducible factors 1(HIF1α) under 5% O2 condition (Fig. 2A). Additionally, PBM–hypoxia cultures showed a dramatic increase in the expression of hypoxia-induced survival factors, such as HIF1α, relative to cells in a normal cell culture condition (5% CO2, non-PBM; Fig. 2A). PBM regulates the expression of growth factors, and PBM–hypoxia cultured cells showed considerable expression of HGF and FGF2; Fig. 2B). The expression of growth factors in PBM–hypoxia-cultured cells was much greater than that of PBM-treated cells or hypoxia-treated cells (Fig. 2C).
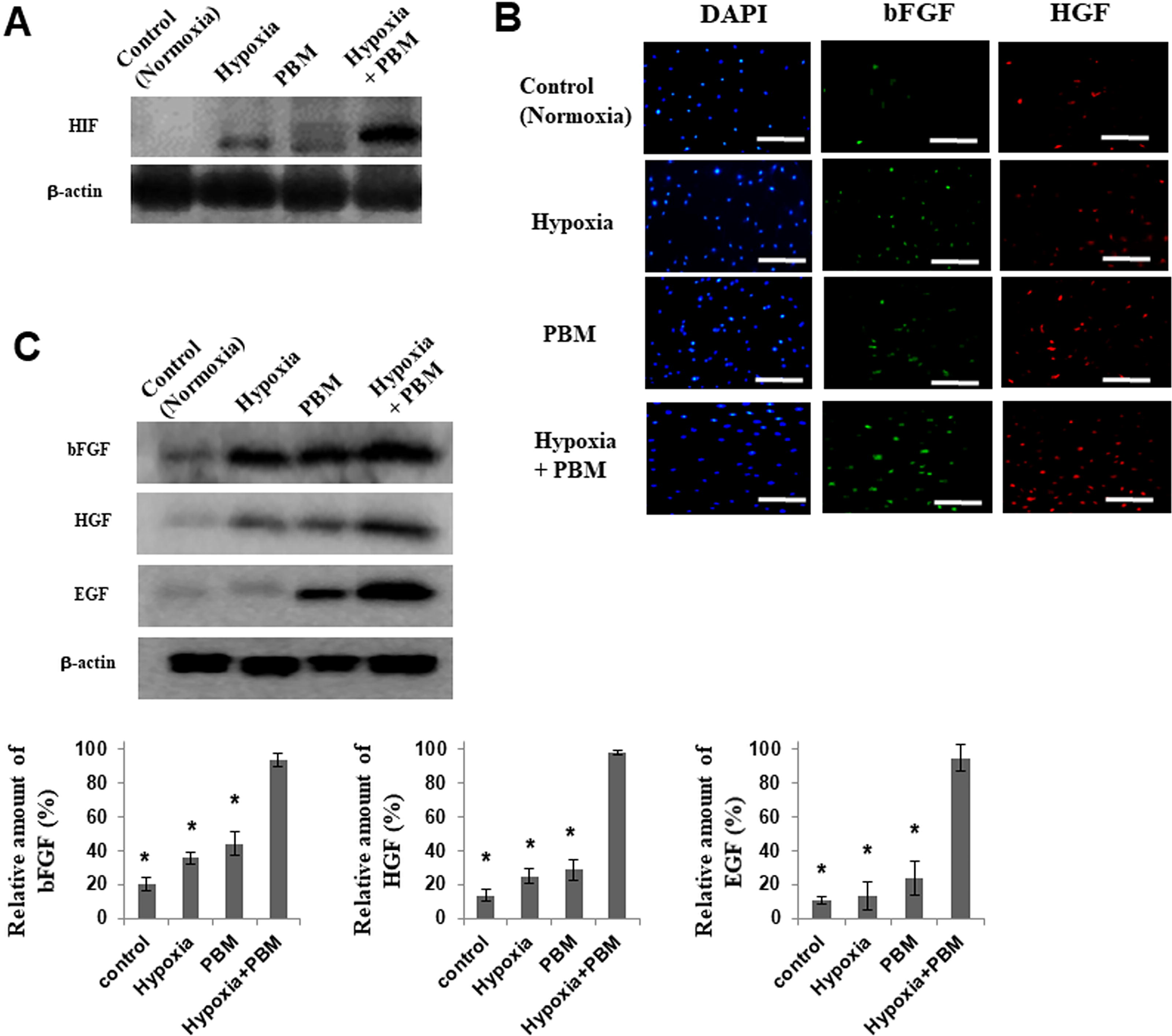
Production of growth factors by cells in PBM–hypoxia culture.
Synergy of effect of PBM and hypoxia on the hepatic differentiation of the hASCs
To investigate the effect of PBM on the hepatic differentiation of the hASCs, we cultured the hASCs under PBM–hypoxia conditions for 21 days, respectively. The expression of alpha-fetoprotein (AFP) and albumin (ALB) was increased by PBM culture conditions on 21 days (Fig. 3A). These results confirm that PBM augments hepatic differentiation in MSC in general. In particular, the expression of human AFP and ALB in PBM–hypoxia-cultured cells was much greater than that of PBM-treated cells or hypoxia-treated cells (Fig. 3A).
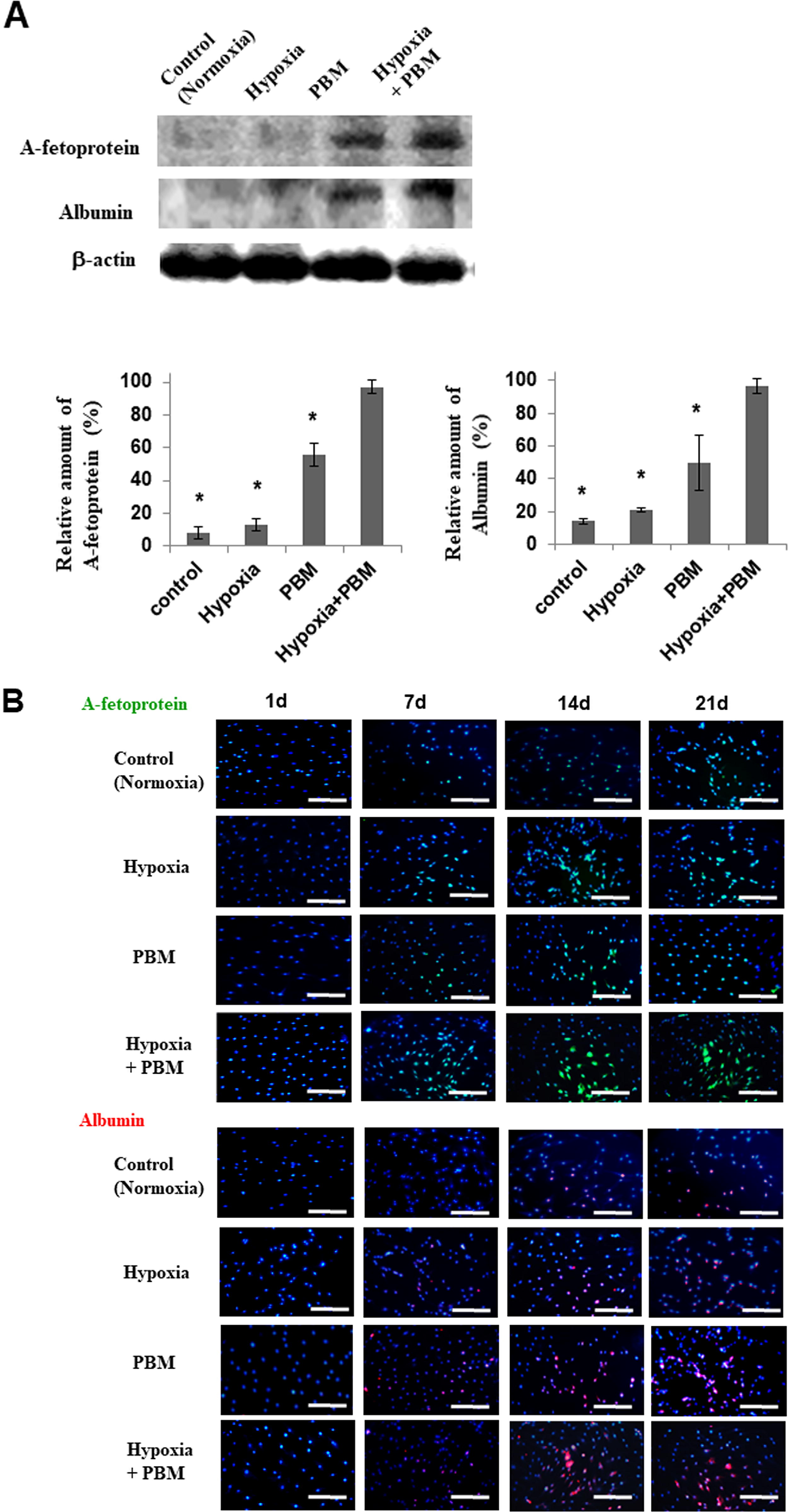
In vitro differentiation.
Many more AFP (early liver-specific markers)- or ALB (late liver-specific markers)-positive cells were observed among cells exposed to hypoxia (5% O2) compared with those subjected to normoxic conditions (21% O2) by immunofluorescence staining (Fig. 3B). The protein level of AFP (early liver-specific markers) was increased by PBM–hypoxia culture for 14 days. However, the protein level of ALB (late liver-specific markers) was increased by PBM–hypoxia culture for 21 days (Fig. 3B). These data suggest that PBM–hypoxia induces upregulation of genes that are predominantly involved in hepatic differentiation and metabolism.
Discussion
As multipotent stem cells residing in the stromal fraction of adipose tissue, hASCs are an abundant source of the cells required for transplantation (Gimble and Guilak, 2003; Huang et al., 2004). hASCs can be easily isolated from a small volume of adipose tissue and subsequently harvested by a simple, minimally invasive method (Park et al., 2013b). In addition, their numbers can be expanded rapidly in vitro using standard cell culture techniques. Most importantly, PBM increases the gene expression and release of several hepatic differentiation-related growth factors, including FGF, EGF, and HGF, from stem cells (Park et al., 2014; Park et al., 2017). In this case, the induction does not depend on the oxygen tension and involves the activation of a different regulatory mechanism possibly mediated by mitogen-activated protein kinase and phosphatidylinositol 3-kinase/Akt signaling pathway (Hu et al., 2007).
Hypoxia is a critical microenvironmental factor for normal embryonic development (Imanirad and Dzierzak, 2013). Mammalian embryonic development occurs at low intrauterine O2 levels ranging from 2% to 9%. Also, hypoxia is known to promote cell differentiation in many tissues (Park et al., 2013a). However, the effect of hypoxia on the differentiation of stem cells varies among individual tissues (Park et al., 2013b). Most importantly, the effect of hypoxia on the hepatic differentiation of hASCs was previously unclear. Our results revealed that hypoxia (5% O2) induced the differentiation of mesenchymal cell progenitors into hepatic cells, as evidenced by an increased expression of the hepatic cell marker ALB and an increased number of AFP- and CK8/18-positive cells.
In addition, the metabolic/molecular biology studies performed on hASCs show that mitochondrial respiration can provide a sufficient amount of energy under PBM–hypoxia culturing to permit cell proliferation (Park et al., 2017). Thus, we applied 5% O2 treated with PBM to research the effect of PBM–hypoxia condition on the hASCs. We discovered that differentiation and secretion of EGF, HGF, and FGF were also enhanced by PBM. It has been reported that hypoxia promotes the differentiation and attenuates the stemness of hASCs through blocking of the Wnt/β-catenin pathway (Sengbusch et al., 2005). These data suggest that the PBM–hypoxia had effects on either hepatic differentiation or self-renewal of the hASCs. In PBM–hypoxia cultures, if growth factors are secreted from stem cells after PBM and hypoxia, the factors might stimulate the hepatic differentiation of stem cells in an autocrine or paracrine manner (Park et al., 2013b).
Footnotes
Author’s Contributions
I.-S.P.: Conceptualization, data curation, formal analysis, funding acquisition, investigation, methodology, project administration, resources, supervision, validation, visualization, writing—original draft, and writing—review and editing.
Ethics Statement
An ethics statement is not applicable because this study does not involve animals or humans.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by the Ajou University Research Fund and National Medical Center (grant number: NMC I-2024-002), South Korea.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.