
Abstract
Blastoids are stem cell-based cellular models that resemble preimplantation embryos and have the potential to ease the investigation of developmental processes and associated diseases. Despite the fast-paced progress of blastoid technology in mice and humans, there was limited evidence of its potential for nonhuman primates. Wu et al. developed a highly efficient protocol for blastoid production in rhesus monkeys and performed standard assays for blastoid characterization at the cellular and molecular levels. By applying cellular reprogramming, Wu et al. generated blastoids from young and aged monkeys. The integration of blastoid protocol with microfluidics allowed their production at scale. If follow-up reports describe blastoid differentiation to peri- and post-implantation stages, it may place rhesus monkeys as an attractive model species for exploring mammalian development ex vivo.
The time-window from fertilization to gastrulation is one of the most dynamic periods in mammalian ontogeny (Rossant, 2018; Tam and Loebel, 2007). Despite the importance of understanding post-implantation development, these developmental processes remain less understood due to ethical issues with harvesting post-implantation embryos, most notably in nonhuman primates and humans. The development of stem-cell-based embryo models (blastoids thereafter) arrived as an alternative route for dissecting late preimplantation development, implantation, and post-implantation development (Harrison et al., 2017; Rivron et al., 2018). The fast adoption of this technology by several research groups and the accelerated development of such models (Liu and Polo, 2024) reinforce their potential. In sharp contrast, the progress of blastoid technology has been slow for nonhuman primates. The sole report on blastoids was done in cynomolgus monkeys (Li et al., 2023), although it allowed the production of blastoids capable of implantation in vivo and early pregnancy development.
Wu and co-authors crafted a blastoid protocol for rhesus monkeys by stepwise adaptation of those developed for mouse, macaques, and humans (Wu et al., 2025). The initial production of blastoids requires high survival of stem cells after dissociation and adequate cellular aggregation. Wu and co-authors tested chemicals to enhance the survival of rhesus monkey embryonic stem cells (rmESCs) after single-cell dissociation. The experiment revealed that well-known small molecules routinely used for ESC passaging in humans and nonhuman primates (Aksoy et al., 2021; Chen et al., 2021; Watanabe et al., 2007) led to high survival of dissociated rmESCs and accelerated spheroid formation (Fig. 1). Since rmESC survival reached ∼80% after dissociation, it would be interesting to prospect additional conditions for higher cell survival.
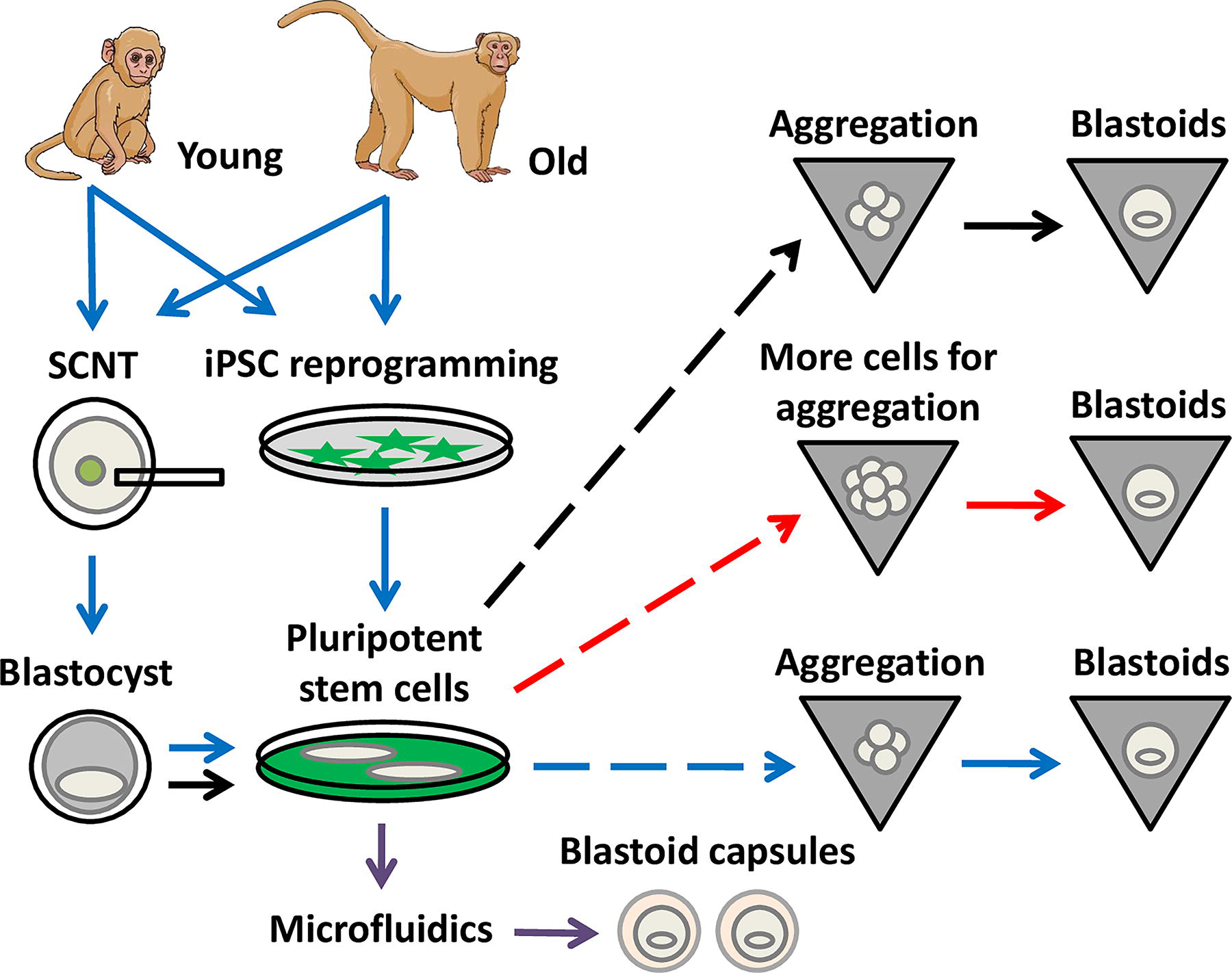
Blastoid production from young and aged rhesus monkeys. Blastoid production was crafted by high survival of pluripotent cells after dissociation (dashed lines), and use of hypoblast- and trophectoderm-inducing media. Late-passage pluripotent stem cells can give rise to blastoids by increasing the number of starting cells and extending the culture in hypoblast-inducting medium (red arrows). Cellular reprogramming allowed blastoid production from young and aged donors, albeit at lower efficiency (blue arrows). Microfluidics with hydrogel microspheres led to large-scale production of blastoid capsules (purple arrows). iPSC: induced pluripotent stem cells. SCNT, somatic cell nuclear transfer.
Blastoids require adequate differentiation conditions to establish the three cell lineages found in blastocysts (Liu and Polo, 2024). Wu and co-authors used a two-step approach that relies on trophectoderm (TE) and hypoblast from rmESCs during aggregation. The initial blastoid medium was for inducing hypoblast differentiation, while the later was for TE formation. The assessment of a few lineage-specifying transcription factors was initial proof for the differentiation of hypoblast and TE cell lineages (i.e., GATA3 for TE, GATA4 for hypoblast). For high-passage rmESCs (>25 cell passages), blastoid production was tweaked by increasing starting cell numbers and extending their culture in hypoblast-inducing medium (Fig. 1). Collectively, these adaptations in the protocol increased the efficiency of blastoid production from ∼30 to ∼80%, as determined by the rate of blastoid cavitation (Wu et al., 2025). Since TE stem cells can be derived from rhesus embryos (rmTSCs) (Keding et al., 2025; Midic et al., 2018), it remains unknown if aggregation of rmESCs with rmTSCs could make the protocol simpler and more efficient.
The validation of blastoids requires extensive cellular and molecular characterization. Wu and co-authors explored the identity of cell types in blastoids by deriving cell lines. These cell lines displayed the expression of cell-lineage transcription factors (e.g., POU5F1, NR2F2, and TEAD4), thus suggesting the stability of their cellular identities. To further explore these cell types, Wu and co-authors performed single-cell RNA sequencing of monkey blastoids. The comparison with published datasets led to the clustering of nonhuman primate and human blastoids versus monkey blastocysts, which revealed that their source was the main factor of transcriptional variation. Due to the limited exploration of these datasets, additional transcriptional analyses between monkey and humans blastoids could be informative about peri-implantation molecular processes, estimate cellular viability, infer differentiation potential, and potentially the gene regulatory networks governing key cell types.
Embryos are the main source of stem cells for blastoid production, although they limit their production to unknown genomic genotypes. Cellular reprogramming generates tailored stem cells for blastoid production in mice and humans (Li et al., 2019; Liu et al., 2021). To reprogram somatic cells from aged rhesus monkeys, Wu and co-authors relied on somatic cell nuclear transfer (SCNT) and reprogramming by defined factors (i.e., iPSC reprogramming) using the non-integrating Sendai virus approach (Wu et al., 2025). Both SCNT-rmESCs and induced pluripotent stem cells (rm-iPSCs) were derived from young (1–5 years old) and aged (15–25 years old) rhesus monkeys (Fig. 1). The reprogrammed rmESCs were characterized at the cellular level by blastoid formation and at the molecular level by expression of pluripotency markers and karyotyping. The limited description of reprogramming assays did not reveal their efficiencies (cloned blastocysts per oocytes or rm-iPSC colonies from starting fibroblasts) or the impact of donor rhesus monkeys (genotype or age) on reprogramming efficiencies. Blastoid production from SCNT-rmESCs and rm-iPSCs was lower than rmESCs (∼60% vs. 80%, respectively; Fig. 1). Nonetheless, these analyses suggest that both young and aged somatic cells were amenable to reprogramming to a pluripotent state and gave rise to morphologically similar blastoids. An interesting follow-up analysis would dissect the molecular differences among embryo-derived and reprogramming-based blastoids, since it was expected that stem cell derivation would erase most, if not all, epigenetic memory of the donor somatic cells (Brambrink et al., 2006; Johannesson et al., 2014; Moura, 2023). The derivation of rmESCs, rmTSC, and extraembryonic endoderm cells reinforced the similarities between blastocysts and blastoids (Wu et al., 2025), although there was a limited characterization of such cell lines.
A fundamental goal of blastoid technology in model organisms is to demonstrate their ability to recapitulate peri- and post-implantation development both in vitro and in vivo. Wu and co-authors began this validation using an extended in vitro culture model that partially recapitulates implantation (Li et al., 2023). The blastoids attached to the culture dish in a similar fashion to blastocysts. Further, these attached blastoids expanded as 2D cell culture and secreted monkey chorionic gonadotropin. Unlike the previous study in macaques (Li et al., 2023), the rhesus blastoids were not reported to develop any further in vitro and had a short-lived fate after transfer to recipient females. Therefore, it remains paramount to demonstrate the differentiation of rhesus blastoids to peri- and post-implantation stages.
The scale of blastoid production is directly proportional to its potential application. Hence, Wu and co-authors applied microfluidics technology to generate synthetic hydrogel microspheres for scalable encapsulation of blastoids (Fig. 1). The hydrogel biocompatibility was demonstrated by encapsulating dissociated stem cells and blastoids, which survived and proliferated accordingly. The microfluidics emulsion approach allowed a large-scale production of encapsulated blastoids (∼4,000 blastoids per hour). The capsule was stable for ∼42 hours under in vitro conditions, and released blastoids were able to attach to the culture dish, thus suggesting preserved viability. There was evidence of higher cryotolerance of encapsulated blastoids, possibly due to diminished cell death after thawing. Although these findings are of clear interest, it needs further evidence to demonstrate the advantage of encapsulated blastoids for long-term cryopreservation. Blastoid capsules transferred for 6 hours to the uteri of recipient females were recovered more efficiently than blastoids.
This is the first report on blastoid production in macaques (Macaca spp.) using cellular reprogramming and the first in rhesus monekys (Wu et al., 2025). In comparison to the report on cynomolgus monkeys (Li et al., 2023), cellular reprogramming was used for blastoid production from aged animals, and further technical leaps led to their large-scale production. A major drawback of the report was the limited characterization of their rhesus blastoids and lack of experimental evidence of their later development. It remains paramount to explore the developmental potential of these blastoids in vitro and in vivo. At least partially, these discrepancies could be due to the clear use of naïve ESCs in the cynomolgus monkeys report (Li et al., 2023). If the progress occurs in the foreseeable future, this scalable blastoid system could become useful for evolutionary studies of developmental biology, toxicological studies, genetic screens, more sophisticated disease models, and contributing to the understanding of human development and disease using human-monkey in vitro chimeras (De Los Angeles et al., 2022).
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No information to declare.
Author’s Contributions
M.T.M.: Writing—original draft, review and editing.