
Abstract
The hematopoietic system consists of well-characterized lineages that provide a diverse repertoire of cell subsets and highly specialized immune functions. This review summarizes the key characteristics and criteria that define hematopoietic cell fate conversion. Perturbation of lineage-specific transcription factors can enable cell fate reprogramming through transdifferentiation, dedifferentiation, or lineage diversion. Members of the CCAAT/enhancer binding protein family of transcription factors, which efficiently convert B cells into macrophages and neutrophils, serve here as a prime example for illustrating the underlying molecular processes. In addition, this review explores disease-associated phenomena, such as lineage infidelity in leukemia, and highlights novel therapeutic strategies that employ fate conversion. Controlled reprogramming systems can also serve as scalable and robust in vitro platforms for screening approaches and for dissecting functional mechanisms. Thus, hematopoietic cell fate reprogramming provides valuable insights into the specification of cellular identity and offers translational opportunities in immunity and cancer.
Introduction
Over the last several decades, research in experimental hematology and immunology has systematically refined our understanding of the architecture of immune cell compartments and their cellular subsets, including distinct progenitor cells (Jacobsen and Nerlov, 2019). This progress has been enabled by the availability of core experimental methodologies, such as fluorescence-activated flow cytometry, allowing the precise definition of cell subsets by surface marker combinations and their prospective isolation. More recent technological advances, including high-throughput single-cell transcriptomic profiling, have further increased the capability to resolve cell states, revealing additional subsets, intermediates and rare populations. As a result, models of the hematopoietic landscape continue to expand and shift, partially replacing long-standing maps of hematopoiesis (Laurenti and Gottgens, 2018). Despite these ongoing revisions, hematopoiesis remains one of the best-understood mammalian differentiation systems and offers an unprecedented experimental toolkit. It therefore serves as a paradigm system to uncover general principles of tissue specification, homeostasis, and, last but not least, fate conversion that may apply to tissues beyond the blood system.
Notably, the establishment and stability of hematopoietic cell identity are not as rigid as classical models of the hematopoietic system may suggest. Traditional “sticks-and-balls” representations implicate one-dimensional differentiation trajectories, discrete endpoints, and a “vacuum” of fate potential between defined cell types (Guilliams et al., 2018). However, experimental studies in developmental biology have long indicated that somatic cell identity can be reversed under defined conditions, enabling the acquisition of alternative fate potential, including, in extreme cases, pluripotency in fully differentiated cell types (Takahashi et al., 2007; Takahashi and Yamanaka, 2006).
Mechanistically, cell fate conversion typically involves activation of lineage-defining transcription factors, rewiring of gene regulatory states and networks, and remodeling of the epigenetic landscape (Cedar and Bergman, 2011). In the hematopoietic system, these principles can manifest as direct cell fate conversion (transdifferentiation), as well as through processes involving dedifferentiation toward progenitor-like states and lineage diversion in less committed subsets (Cobaleda and Busslinger, 2008; Graf, 2011; Pires et al., 2019).
This review highlights key principles and mechanisms of hematopoietic cell fate reprogramming, with a major focus on direct cell fate conversion, dedifferentiation, and lineage diversion within the hematopoietic system. Readers interested in induced pluripotent stem cell generation and subsequent directed differentiation toward mature somatic cells are referred to recent overviews (Cerneckis et al., 2024; Morris, 2026; Takahashi and Yamanaka, 2016). Hematopoietic cell fate reprogramming can serve as a highly valuable perspective beyond refining our understanding of the regulatory logic and architecture of the hematopoietic system. Fate conversion programs may contribute to pathological mechanisms and can be leveraged for disease modeling. In addition, reprogramming-based systems can provide rapid and reductionistic platforms for immune cell engineering. Finally, emerging therapeutic strategies that aim to harness immune effector functions through hematopoietic cell reprogramming are discussed. Direct cell conversion involving members of the CCAAT/enhancer binding protein (CEBP) transcription factor family is a well-studied system and will be used as a prime example throughout the sections to illustrate the principles of the above-mentioned concepts (Cirovic et al., 2017; Xie et al., 2004).
A Primer in Cell Plasticity and Fate Reprogramming
Immune cells undergo complex differentiation steps until they reach a state of full immune effector potential. Along this line, the progenitor cell commits to the differentiated cell states and becomes more and more restricted, thereby prohibiting alternative fates (Adolfsson et al., 2005; Jacobsen and Nerlov, 2019). It is important to emphasize that this progressive process results from the stabilization of a gene regulatory state and not by irreversible loss of genetic information (with the exception of red blood cells undergoing enucleation or lymphocytes during rearrangement of antigen receptor segments) (Huang, 2012). This implies that an enforced transition of the gene regulatory state by perturbation is possible and can reset or redirect the identity of a cell and potentially unlock regenerative or immune-modulatory functions.
Throughout the last century, impressive experimental examples demonstrated that a somatic cell (nucleus) can indeed be reprogrammed, challenging our foundational understanding of cellular identity (Graf, 2011). Nuclear transfer experiments indicated that differentiated nuclei retain stem cell-like potential when placed in a permissive environment (Gurdon, 1962). This approach ultimately resulted in the cloning of Dolly the sheep, representing probably the most impressive textbook entry from this field of science (Wilmut et al., 1997). Moreover, cell fusions, the generation of heterokaryons, showed rapid reactivation of silent lineage genes without DNA replication pointing to the importance of soluble factors acting in trans (Blau et al., 1983,1985). The discovery of fibroblast-to-muscle cell conversion represented another major advancement in the field and led to the identification of the transcription factor Myod1, the first defined single-factor component inducing lineage conversion (Davis et al., 1987; Tapscott et al., 1988).
Most interestingly, two decades after the discovery of Myod1, Shinya Yamanaka and colleagues reported that a simple combination of four transcription factors, SOX2, OCT4, MYC, and KLF4, can induce a pluripotent, embryonic stem cell-like state in fibroblasts (Takahashi et al., 2007; Takahashi and Yamanaka, 2006).
This finding sparked strong interest to identify alternative and additional reprogramming factors and combinations to induce different lineages. To date, numerous additional transcription factors and combinations have been described to convert fibroblasts into various other cell types, such as neurons, cardiomyocytes, hepatocytes, myoblasts, leukocyte subsets and endothelial-like precursor cells. The list of achievable reprogramming cell types continues to grow and includes the conversion towards distant cell fates across germ layers (Pires et al., 2019; Wang et al., 2021).
One can define at least three trajectories of hematopoietic cell fate reprogramming (Fig. 1). First, committed and mature cells can directly convert into another committed cell type through transdifferentiation, resulting in a direct switch of cellular identity (but allowing transient, intermediate states). Second, during dedifferentiation, cells first enter a less mature and uncommitted state before subsequently differentiating into an alternative, non-canonical cell type. Third, lineage diversion describes differentiation into an alternative cell type through the redirection of the developmental route of an uncommitted or partially committed progenitor cell (Cobaleda and Busslinger, 2008).
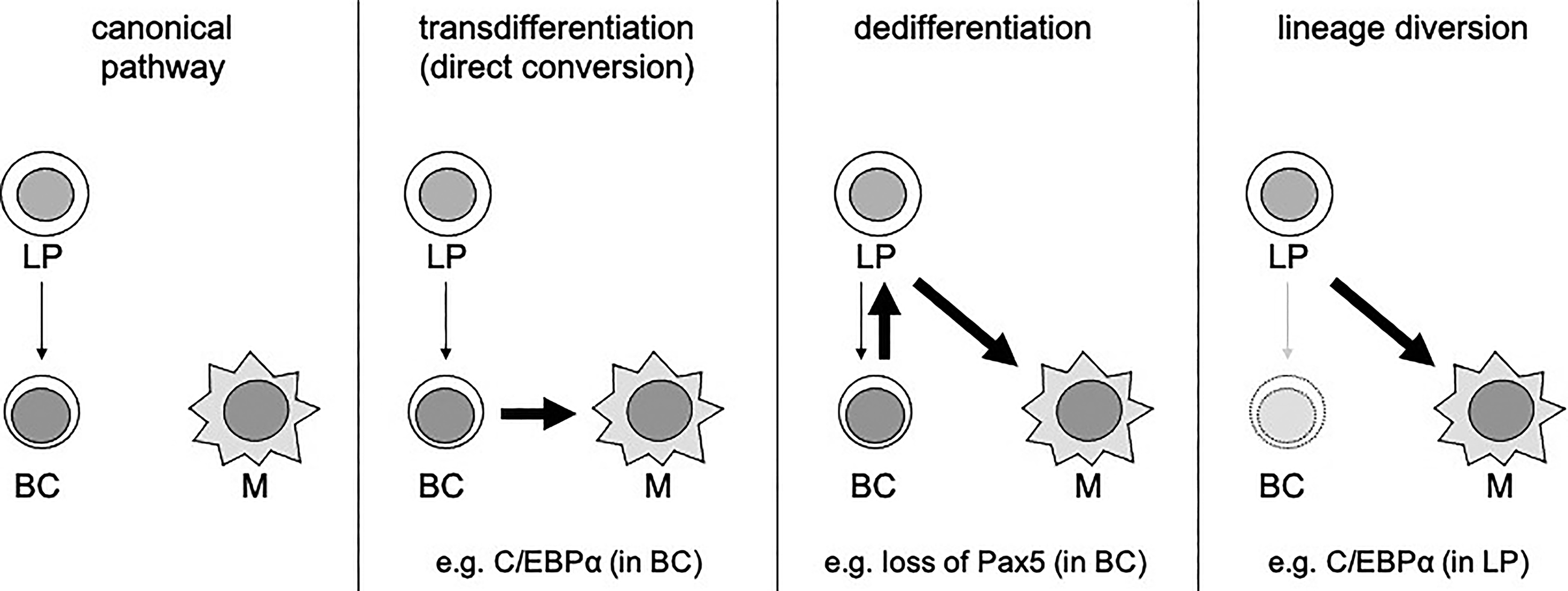
Modes of hematopoietic cell fate reprogramming. Comparison of the different operational modes for fate reprogramming in relation to the canonical pathway. Arrows indicate (fate) transitions. Bold arrows point to reprogramming re-routing events. Note, that a macrophage progenitor is omitted in this illustration for clarity. LP, lymphoid progenitor; BC, B cell; M, macrophage.
For all of these operational modes, examples can be found based on the manipulation or enforced expression of lineage-specific transcription factors in the hematopoietic system. A prototypical and simple example is the lymphoid-to-myeloid transdifferentiation instructed by ectopic activation of the CEBP family member, C/EBPα, as will be further discussed below (Xie et al., 2004).
Besides C/EBPα, around 1600 transcription factors are encoded in the human genome according to a recent estimation (Lambert et al., 2018). This fact alone implies that there are many more reprogramming factors and combinations yet to be discovered. Transcription factors share the ability to bind chromatin in a sequence-dependent manner and regulate gene expression. Besides the defining DNA-binding domain (DBD), transcription factors can harbor transactivation domains and additional less well-defined regulatory domains (RD) controlling protein domain-interaction behavior, activity, or stability of the transcription factor.
Specifically, the proteins encoded by the six genes of the CEBP-family (CEBPA, CEBPB, CEBPD, CEBPG, CEBPE, and DDIT3) all contain a characteristic carboxy-terminal region (bZIP domain), representing a designated DBD enriched for a basic amino-acid sequence, nuclear localization signal, leucine zipper domain and tail region facilitating protein–protein interactions (Tsukada et al., 2011). C/EBPs act in a pleiotropic fashion participating in fundamental cell biological processes, such as cell differentiation, metabolic regulation, inflammation, and cell proliferation. In this way, they play a crucial role in the development and function of cell types, including adipocytes, hepatocytes, and several leukocyte subsets. The family members are expressed in a lineage-specific manner. The founding member C/EBPα, for example, is indispensable for myeloid cell development most strongly affecting granulocyte–monocyte progenitor (GMP) formation, whereas deletion at a subsequent differentiation step remains largely ineffective (Heath et al., 2004; Zhang et al., 1997). Importantly, CEBPA is one of the most affected genes in acute myeloid leukemia (AML). Mutations of CEBPA are found in ∼10% of AML patients and the involvement in the disease mechanism is associated with aberrant interaction with components of the cell cycle machinery and differentiation block (Nerlov, 2007; Pabst et al., 2001; Pabst and Mueller, 2009). The strong phenotype in these settings of defective C/EBPα function is one reason for the implication of a potent lineage-instructive role in a heterologous expression system. There are multiple other transcription factors that fulfill lineage-specific roles during hematopoiesis. GATA factors belong to the earliest found examples for lineage regulation and display instructive capacity in multiple hematopoietic contexts, primarily toward a myelo-erythroid and eosinophilic/basophilic identity; forced expression can redirect progenitor trajectories depending on developmental stage, dose, order of expression, and niche signals (Iwasaki et al., 2003, 2006; Iwasaki and Akashi, 2007; Taghon et al., 2007). In the lymphoid compartment, PAX5 and EBF1 serve as master regulators in the B-cell lineage, whereas BCL11B is strongly associated with T-cell identity (Li et al., 2010; Nechanitzky et al., 2013; Nutt et al., 1999; Rolink et al., 1999). PU.1 (SPI1) functions as a context- and dose-dependent regulator across hematopoietic cell types and participates particularly in myeloid and dendritic cell programs (Barozzi et al., 2014; Carotta et al., 2010).
Prominent Examples of Hematopoietic Cell Fate Reprogramming
Notably, all of the transcription factors introduced above belong to a comparatively small set of master regulators whose single perturbation can already be sufficient to trigger drastic changes in cellular identity under permissive conditions. This includes C/EBP or GATA factors as strong lineage instructors, or PAX5 and BCL11B, which can also serve as “guardians” or “keepers” of differentiated cell states (Cirovic et al., 2017; Kulessa et al., 1995; Li et al., 2010; Nutt et al., 1999; Pevny et al., 1991; Rolink et al., 1999; Xie et al., 2004) (Table 1). The detection of bona fide conversion events is not trivial and should involve thorough evaluation at multiple levels, including altered phenotypic identity, function, and, most importantly, a clarification of the cell developmental history (Fig. 2). First, identity should be assessed by concordant (multi-parameter) phenotypic profiling. At a minimum, this includes alteration in lineage-specific surface marker expression, moreover transcriptome-wide changes toward the target lineage, and if possible, consistent epigenetic remodeling. Second, a shift in functional properties should be demonstrated using lineage-appropriate assays that cover key characteristic behaviors of the target (and/or starting) cell. Third, lineage relationship and history must be clarified to rule out the outgrowth of rare contaminants, selective survival of preexisting intermediates, or, in general, mixed cultures. Genetic lineage tracing (e.g., conditional Cre-based ancestry labeling) and permanent genetic scars (e.g., unique immunoreceptor rearrangements or mutations) are particularly valuable in this context (Laiosa et al., 2006; Xie et al., 2004).
Examples for Hematopoietic Cell Fate Reprogramming Induced by Manipulation of Single Transcription Factors

CLP, common lymphoid progenitors; GMP, granulocyte–monocyte progenitor.
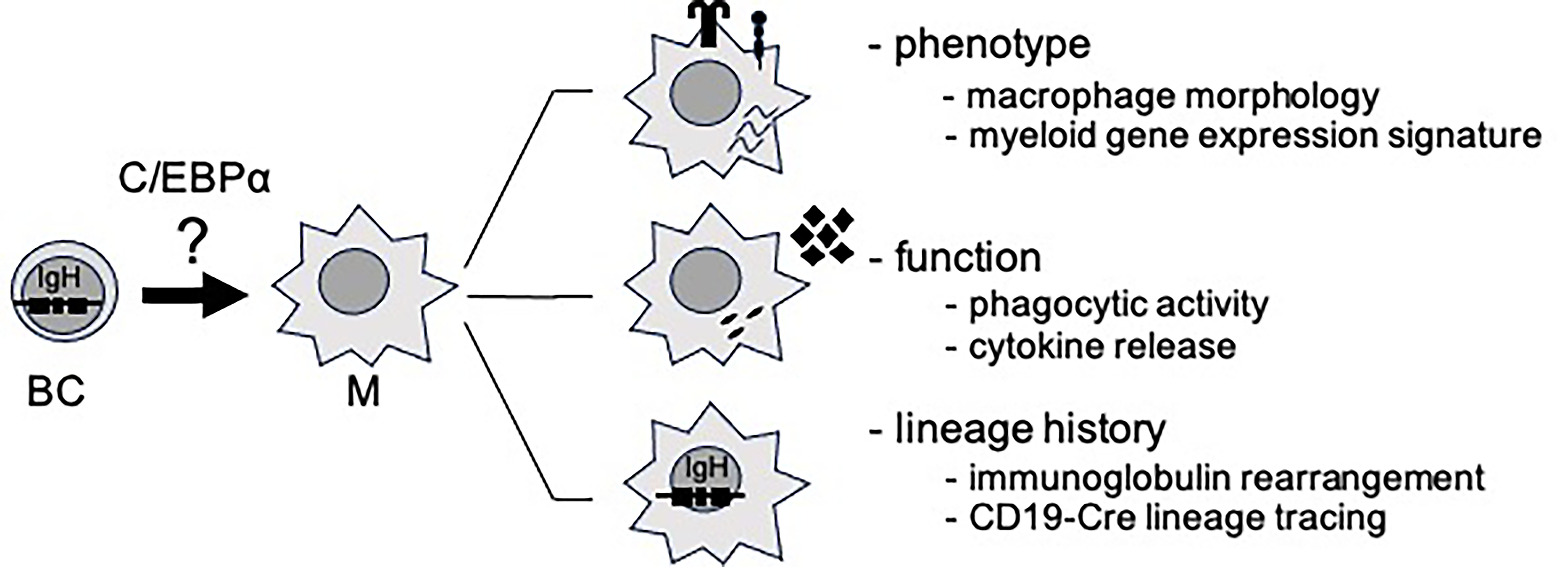
Criteria for confirmation of bona fide reprogramming. Validation level of criteria (phenotype, function, lineage history) and exemplary readouts for each level illustrated for C/EBPα-induced lympho-myeloid transdifferentiation. BC, B cell; M, macrophage; IgH, immunoglobulin heavy chain locus.
Following these conditions, several underlying mechanistic principles across the operational modes can be observed upon enforced expression of transcription factors and lineage conversion. This includes the induction of target cell-specific gene expression patterns and establishment of gene regulatory networks (Bussmann et al., 2009). Concomitant suppression of the original program corroborates the transition, which is then consolidated by autocrine, autoregulatory feedback loops and epigenetic changes (Hsu et al., 2006; Xie et al., 2004).
Indeed, most of these conceptual aspects can be observed in the paradigmatic example of C/EBP-induced cell fate reprogramming. Graf et al. found that enforced, single expression of C/EBPα can reprogram murine committed B cells into macrophage-like cells with substantial efficacy and speed (Bussmann et al., 2009; Xie et al., 2004). Notably, the speed of conversion can be regulated by a posttranslational modification of C/EBPα at Arginine 35, induced by the methyltransferase Carm1. Mechanistic studies revealed that blocking this modification results in increased affinity to the binding partner PU.1, leading to both accelerated transition and a lineage bias toward macrophages during cell specification (Torcal Garcia et al., 2023). Furthermore, kinetic analyses of gene expression patterns in transitioning pre-B cells did not reveal the induction of a progenitor state, suggesting bona fide transdifferentiation (and not dedifferentiation), as the underlying operational mode following enforced C/EBPα expression (Di Tullio et al., 2011).
Considering the introduced requirements for the cell conversion, enforced expression of C/EBPα checks all the boxes. This includes overt phenotypic changes, including macrophage-like morphology and a shift in lineage-specific surface markers (phenotypic requirement). In addition, converted cells acquired typical functionality of innate immune cells, specifically phagocytic activity, migratory behavior, and cytokine-release upon inflammatory stimulation (functional requirement). Importantly, the lymphoid developmental origin of the macrophage-like cells was confirmed in two ways—the presence of irreversible rearrangement events within the immunoglobulin locus and reporter positivity in a B cell lineage-specific tracing system (origin requirement) (Bussmann et al., 2009; Xie et al., 2004).
From a mechanistic perspective, several follow-up studies have pointed to multimodal reprogramming of cellular identity (Bussmann et al., 2009; van Oevelen et al., 2015). This process is characterized by the rapid induction of a myeloid gene expression signature and the simultaneous destabilization of the B cell program. Cross-antagonism between C/EBPα and the master regulator of B cell commitment, PAX5, is indeed an important and known feature in this lineage switch (Hsu et al., 2006). At the epigenetic level, the newly acquired state is stabilized by the involvement of histone modifiers and broad remodeling of the chromatin landscape (Barneda-Zahonero et al., 2013; van Oevelen et al., 2015). On a larger scale, transdifferentiation is accompanied by topological changes in genome organization. Although topologically associated domain organization per se was dispensable for transdifferentiation, induced macrophages lacking the re-organisation were partially impaired to mount an acute inflammatory response after endotoxin challenge (Stik et al., 2020). Surprisingly, promoter DNA methylation remained largely unchanged during cell conversion, indicating that myeloid cell identity is also compatible with a lymphoid methylation pattern (Rodriguez-Ubreva et al., 2012). As an additional layer of fate fine-tuning, posttranscriptional regulation by microRNA programs stabilizes the newly acquired program (Rodriguez-Ubreva et al., 2014).
Hematopoietic cell fate conversion based on the dedifferentiation operational mode is evident in the B cell lineage after inactivation of the master regulator Pax5 and the subsequent erosion of B cell identity (Nutt et al., 1999; Rolink et al., 1999). In comparison to transdifferentiation, this mode of action clearly proceeds through a progenitor stage before the acquisition of alternative states, including myeloid cells and T cells (Schaniel et al., 2002). Notably, also C/EBPβ was able to convert B cells toward a GMP-like state constantly giving rise to monocytes and neutrophils on a clonal level pointing to a dedifferentiation-based mode of action (Cirovic et al., 2017).
Uncommitted hematopoietic progenitors are susceptible to lineage diversion through multiple mechanisms. For example, Nakajima and colleagues demonstrated that controlled activation of C/EBPα in lymphoid progenitors can drive myeloid fate in vitro and in vivo (Fukuchi et al., 2006). Similarly, GATA1 diverts progenitors toward megakaryocytic-erythroid programs (Iwasaki et al., 2003). Interestingly, fate diversion can also be induced by ectopic cytokine signaling. Introduction of transgenic cytokine receptor components, such as IL-2RB or CSF2R, and stimulation with cognate ligands, shifts the differentiation output of common lymphoid progenitors (CLP) toward myeloid cells. Early T cell progenitors are also susceptible using this approach, but cease to respond once they become more committed (Iwasaki-Arai et al., 2003; King et al., 2002; Kondo et al., 2000). This could indicate the existence of a narrow window for a cellular state along the differentiation axis that is vulnerable to signaling pathway distortion. It was also demonstrated that endogenous signaling pathway activation can induce fate diversion in CLP toward myeloid cells. Notably, Toll-like receptor signaling and the IL-6-C/EBPβ axis were involved in this trajectory, implicating a potent mechanism and purpose for lineage diversion by enforcing the innate immune response after pathogen encounter (Kawano et al., 2025; Nagai et al., 2006). This variant of fate reprogramming strongly resembles well-described adaptive mechanisms in the innate immune system, termed as “emergency hematopoiesis” or “trained immunity” (Swann et al., 2024). It was recently demonstrated that microbial cues can indeed provoke such a long-term effect in mice and humans in vivo (Cirovic et al., 2020; de Laval et al., 2020; Kaufmann et al., 2018). Strikingly, in one of these studies, the effect strictly depended on the presence of C/EBPβ, further demonstrating the marked potential of C/EBP family members in fate specification (de Laval et al., 2020).
Disease Association of Cell Fate Reprogramming
Hematopoietic cell fate conversion is not merely an artificial experimental curiosity but may also constitute an integral part of disease manifestation. Several members of the C/EBP family, in fact, are targeted by immunoglobulin translocation events in B-cell precursor acute lymphoblastic leukemia (BCP-ALL) (Akasaka et al., 2007; Chapiro et al., 2006). The exact consequences for disease remain elusive. However, in one case, CEBPA was found to be overexpressed (Chapiro et al., 2006). In this context, it is tempting to speculate that C/EBP-mediated fate reprogramming is a component of disease etiology or progression (Akasaka et al., 2007; Chapiro et al., 2006). Indeed, rare lineage switch and lineage ambiguity events have been described in hematological malignancies and are clinically relevant, as these patients have poor prognosis and are difficult to diagnose (Hu et al., 2017; Silbert et al., 2025). Mejstrikova and colleagues reported interesting cases of lineage switching BCP-ALL without KMT2A involvement that displayed early monocytosis during treatment. Intriguingly, the conspicuous monocytes shared immunoreceptor gene rearrangements with the transformed B cell blast, pointing to a clonal relationship. Moreover, lineage switch was associated with increased CSF2R levels and CEBPA expression, including hypomethylation of the CEBPA promoter (Slamova et al., 2014). Similarly, malignant B cells in Hodgkin’s lymphoma patients upregulate the myeloid CEBP target gene CSF1R, thereby providing access to additional growth factor resources and further promoting tumor cell proliferation and survival (Lamprecht et al., 2010). In a similar scenario, it was shown that lineage infidelity can be coupled to oncogenic signaling programs. Concurrent NF-κB and Notch activation in committed B cells can induce transformation and prime conversion toward myeloid lineage through a dedifferentiation route (Xiu et al., 2020). In addition, clonally related transitions from lymphoid malignancies to histiocytic/dendritic neoplasms provide evidence that fate-conversion programs can accompany tumor progression (Feldman et al., 2008; Fraser et al., 2009; Mansfield et al., 2026). Further investigations implicated rewiring of master transcription factors, consistent with the loss of B-lineage identity and the gain of myeloid regulators, i.e., downregulation of PAX5 and upregulation of PU.1 and C/EBPβ in follicular lymphoma-associated histiocytic/dendritic neoplasms (Feldman et al., 2008). Together, these studies highlight a potential role of hematopoietic fate conversion in malignancy and support the inclusion of phenotypic plasticity as an additional hallmark of cancer (Hanahan, 2022).
Apart from mechanisms that intrinsically boost tumor survival and progression, fate reprogramming may also contribute to immune evasion under the selective pressure during targeted immune therapy. Results from clinical observation already documented clonally related myeloid relapse following administration of CD19-specific chimeric antigen receptor-modified T cells in B-ALL (Gardner et al., 2016). More recently, 75 cases qualifying as lineage switch were reported, the majority of which represented a B-ALL to AML conversion, with most occurring within the first six months after immunotherapy. General treatment outcomes and overall survival rates in these patients were poor in comparison (Silbert et al., 2025).
In addition, Biragyn and colleagues provided evidence that fate conversion may serve as an indirect mechanism of immune evasion (Chen et al., 2022). They reported the conversion of nonmalignant B-cell precursors toward macrophage-like cells. This process was enforced by cancer cells across different tumor entities, resulting in a protumorigenic microenvironment characterized by dampened T-cell-mediated effector functions.
Clearly, all these findings advocate for a better understanding of the direct and indirect roles and mechanisms of cell fate reprogramming during tumor progression.
Besides malignancy, there are few examples involving hematopoietic dedifferentiation or transdifferentiation events following pathogenic challenge. Macrophages that are directly infected with murine cytomegaly virus compared to bystander cells lose key characteristics of innate immune cell identity, including morphological, phenotypic, and functional features such as cytokine production, phagocytic activity, and antigen-presentation. This compromised immune function and, as a consequence, increased susceptibility toward secondary infection. Notably, infected macrophages showed significant reduction of key myeloid transcription factors, such as Cebpa, Cebpb, Spi1 (PU.1), and Irf8. Therefore, the dedifferentiated-like state can potentially be explained by a virus-mediated breakdown of the lineage-defining gene regulatory network (Baasch et al., 2021). Moreover, signs for transdifferentiation of class-switched B cells toward activated neutrophils were found in peripheral blood of severe COVID-19 patients and was strongly associated with CEBPD and CEBPE expression (Wilk et al., 2020). Importantly, ectopic expression of human CEBPE in B cells indeed leads to efficient transdifferentiation to neutrophils in vitro (Cirovic et al., 2017). However, whether the postulated conversion in the COVID-19 patients represents true events or analytical artifacts is under debate (Alquicira-Hernandez et al., 2021). Addressing the criteria for lineage conversion as described above may help to further validate these findings.
Translational Applications of Hematopoietic Cell Fate Reprogramming
All the examples mentioned in the previous chapter may impose a negative connotation to hematopoietic cell fate programming. One may therefore ask whether there is also a positive twist to fate conversion. Indeed, the same logic for conversion that fuels malignancy can be applied for the benefit of therapeutic approaches. Differentiation therapy applied in acute promyelocytic leukemia involves therapy-induced terminal differentiation of malignant blast cells leading to disease remission (Huang et al., 1988; Nowak et al., 2009). Analogously, fate conversion has been suggested to bear therapeutic potential within a transdifferentiation-based framework. As proof of concept, a report showed that transdifferentiation by enforced expression of C/EBPα in vivo is able to convert B cell-derived malignancies into nontumorigenic cells in a mouse model (Rapino et al., 2013). Consistently, Majeti and colleagues demonstrated the successful conversion of human B cell-derived ALL into nonleukemic macrophage-like cells by transient expression of PU.1 and C/EBPα in the xenograft mouse model (McClellan et al., 2015). In another example, conversion of B cell leukemia toward the T cell lineage by activation of Notch signaling was achieved. In this case, however, the malignant state was transferred to the new fate (Somasundaram et al., 2016). This outcome illustrated that fate conversion towards an alternative differentiated state alone does not necessarily eliminate tumorigenicity. One caveat for such an approach, in general, is that virtually all tumor cells have to be hit, in order to prevent relapse due to refractory or missed cells (Rapino et al., 2013). Transdifferentiation of cancer into potent antigen-presenting cells was developed as novel immunotherapy and elegant solution to address this issue (Ascic et al., 2024; Linde et al., 2023; Zimmermannova et al., 2023). Indeed, transdifferentiated cancer cells possessed unlocked myeloid functionality and were able to activate presentation of tumor-associated antigens and subsequently anti-tumor immunogenicity. This represents a unique immunotherapeutic approach with the potential to develop, de facto, a universal and personalized vaccination therapy against cancer. Majeti and colleagues used a protocol involving C/EBPα and PU.1, as shown previously, to induce a macrophage-like state in tumor cells (Linde et al., 2023; McClellan et al., 2015). It was demonstrated that the induction of the three transcription factors PU.1, IRF8, and BATF3 efficiently converted fibroblasts into conventional type 1 dendritic cells (cDC1) (Rosa et al., 2018,2022). Pereira and colleagues applied this conversion protocol to achieve highly potent antigen-presenting activity in cancer cells and anti-tumorigenicity (Ascic et al., 2024; Zimmermannova et al., 2023). This state could be induced in (tumor) cells from a wide range of tissues and germ layers. Strikingly, mice that were able to eradicate tumor cells in a melanoma model, were also able to do so after a secondary challenge indicating potent induction of adaptive immune memory (Ascic et al., 2024). Reprogramming cDC1 in the tumor model in vivo resulted in de novo formation of tertiary lymphoid structures and a shift from a pro- toward an anti-tumorigenic T cell population in a poorly immunogenic or immune-suppressive tumor microenvironment. In light of potential therapeutic use in future, the authors validated a number vector systems for transcription factor delivery and confirmed successful transduction using non-integrative, nonreplicative adenoviral and adeno-associated viral vectors in situ. Notably, while overall transduction efficiency was low, converted cDC1 cells could be found after nine days in the tumor and a minor fraction of under 0.1% of converted tumor cells was sufficient to significantly improve median survival in the xenograft animal model (Ascic et al., 2024). This low requirement for cell conversion potentially represents also an advantageous feature to reduce off-target effects (e.g., by a small fraction of converted healthy tissue cells).
Besides this interesting approach to myeloid reprogramming with high translational potential, there are also examples how to efficiently use direct cell conversion for mechanistic studies in vitro. Graf and colleagues developed an inducible human B cell line model (BLaER1) that allows activation of transdifferentiation after addition of tamoxifen/estradiol with high efficiency (Rapino et al., 2013). After induction, converted cells showed high resemblance with primary monocytes (Gaidt et al., 2016). This model system was used to robustly study human inflammasome pathway components (Andreeva et al., 2017; Gaidt et al., 2016,2017). Moreover, a screening approach based on a CRISPR-Cas9 library was developed to identify regulators involved in transdifferentiation (Arnan et al., 2022). More than one thousand genes, that could be involved in the transdifferentiation of BLaER1 cells were investigated leading to a list of known players and putative regulators of lympho-myeloid conversion. These examples showcase the potential of in vitro transdifferentiation systems to robustly and systematically study myeloid cell biology and innate immunity at scale.
Outlook
Studying and harnessing cell fate reprogramming is a promising approach for understanding the mechanisms underlying hematopoiesis and its associated defects. Considering the complex diversity of the system and the sheer number of transcription factors encoded in the genome, we can expect a growing number of reported transcription factor combinations to target a hematopoietic cell of interest in the future. Elegant screening approaches to efficiently identify new combinations have already been successfully established both within and outside the hematopoietic system (Henriques-Oliveira et al., 2025; Joung et al., 2023; Kurochkin et al., 2026; Liu et al., 2018). Computational tools to predict conditions will further aid to narrow down potential candidates (Hersbach et al., 2022; Jung et al., 2021; Rackham et al., 2016; Sadria and Bury, 2024). Definition of the exact order and stoichiometry could represent a challenge in protocols with more complex combinations of transcription factors (Iwasaki et al., 2006; Kurochkin et al., 2026). In addition, replacement of experimental protocols requiring genetic manipulation with chemical cocktails would represent a major improvement in the field (E et al., 2025; Wang et al., 2023). Notably, induction of functional neutrophils from mouse embryonic fibroblasts was recently reported by the supplementation of thymidine in basic culture medium (Ye et al., 2022). After a decade of significant progress in the field of genetic engineering and genome editing technologies, hematopoietic cell fate reprogramming has the potential to drive and join an era of tailored immune cell engineering.
Author’s Contributions
B.C.: Conceptualization, Writing—original draft, and writing—review and editing.
Footnotes
Acknowledgments
The author thanks the members of the Cirovic laboratory for helpful discussions and critical feedback on the article.
Author Disclosure Statement
The author declared no potential conflicts of interest.
Funding Information
This work was supported by institutional core funding [Institute for Immunodeficiency, Center for Chronic Immunodeficiency (CCI), Medical Center-University of Freiburg, University of Freiburg].