
Abstract
Plasminogen activator inhibitor-1 (PAI-1), the primary physiological inhibitor of tissue-type plasminogen activator (tPA), is a key regulator of fibrinolysis. Elevated levels of PAI-1 are linked to thrombotic disorders and correlate with poor prognosis across various cancers. In this study, we further characterize the RNA aptamer R10-4, previously shown to bind PAI-1 with high affinity and inhibit its antiproteolytic activity. While R10-4’s role in modulating fibrinolysis is established, its influence on cancer cell behavior remains unclear. Here, we demonstrate that intracellular transfection of R10-4 in triple negative breast cancer cells significantly impairs migration and invasion without affecting proliferation, mirroring the effects observed with other PAI-1–specific RNA aptamers. Moreover, conditioned media from R10-4 transfected cells suppress endothelial tube formation and exhibit reduced secretion of the pro-angiogenic chemokine (C-C) motif ligand 5 (CCL5). Collectively, these findings reveal that R10-4 restores fibrinolytic balance and disrupts PAI-1–mediated tumor progression, positioning it as a promising multifunctional candidate for therapeutic development.
Introduction
Plasminogen activator inhibitor-1 (PAI-1), a member of the serine protease inhibitor (serpin) family, regulates fibrinolysis by binding and inactivating tissue-type plasminogen activator (tPA) and urokinase-type plasminogen activator. While transient PAI-1 expression is necessary for tissue remodeling and wound healing, persistently elevated PAI-1 is linked to thrombotic disease, fibrosis, and cancer progression.1,2 In cancer, PAI-1 supports tumor growth, metastasis, and angiogenesis, in part through effects on extracellular matrix (ECM) remodeling and cell signaling.3–8 High circulating PAI-1 is consistently associated with poor prognosis in breast cancer and other malignancies. Additionally, PAI-1 has been shown to promote angiogenesis, which supply tumors with essential nutrients and oxygen.9–11
RNA aptamers are short, structured oligonucleotides that bind proteins with high affinity and specificity. My laboratory has developed several PAI-1 aptamers, including ones that interact with the vitronectin binding site and influence cell migration and invasion.12–15 Another aptamer, R10-4, blocks PAI-1–tPA interaction and enhances fibrinolysis in vitro. 12 Activity data suggest that R10-4 interacts with the reactive center loop of PAI-1, stabilizing a conformation that inhibits protease inhibition. 12 Here, we extend the functional characterization of R10-4 to assess its effects on cancer progression. Using triple-negative breast cancer (TNBC) cells, we demonstrate that intracellular R10-4 impairs migration and invasion, and conditioned media (CM) from R10-4 transfected cells suppress angiogenesis via reduced Chemokine (C-C motif) ligand 5 (CCL5) secretion. These findings reveal a dual activity of R10-4 in targeting both thrombotic and oncogenic pathways of PAI-1.
Materials and Methods
Transient transfection
TNBCs were seeded into a 6-well plate in Dulbecco's Modified Eagle Medium (DMEM) until approximately 80% confluent. Cultured cells were transiently transfected using Lipofectamine 3000 according to the manufacturer’s protocol as previously described. 14 The R10-4 aptamer (gggaggacgaugcggccaggcgucucacucguuacgcuaucguugcguacuucuggacgacucgcugaggauccgaga) and control scrambled aptamer (gggaggacgaugcgguuggugaagggaggucagcauaucuucccgcggaagcgacgacgacucgcugaggauccgaga) (100 nM) were transfected. Transfection efficiency was determined by isolating total RNA followed by Reverse-transcription-polymerase chain reaction (RT-PCR) analysis and by confocal microscopy.
Cell migration (scratch assay)
Transfected TNBCs (0.5 × 105/well) were seeded in 12-well plates and grown overnight at 37°C. Scratch assays were performed as previously described. 16 Images were taken at 0, 6, and 24 h. Cell migration into the denuded area was analyzed using Image J.
Transwell invasion assays were performed as previously described. 12 Briefly, transfected cells (0.5 × 106) were seeded on ECM-coated Boyden chambers (in triplicates), respectively. After 24 h, invaded cells were fixed, stained, and counted.
In vitro angiogenesis assays were performed as previously described. 14 Briefly, Human umbilical vein endothelial cells (HUVECs) (1.5 × 104) were plated on Matrigel-coated μ-slides with endothelial cell growth media 14 or CM from TNBCs transfected with scrambled or R10-4 aptamer. Tube formation was assessed hourly for 6 h, then every 6 h up to 24 h, stained with Calcein AM, and quantified by fluorescence microscopy. The number of tubes was determined using Image J with an Angiogenesis Analyzer plugin as described previously. 14
Cytokine analysis
Transfected (R10-4 and scApt) and non-transfected cells were incubated in Opti-MEM media for 24 h, after which the CM were collected, centrifuged to remove cellular debris, and analyzed for cytokine expression using a human cytokine array kit (R&D Systems). Spot intensities were quantified with Image J. Although multiple cytokines were included on the membrane, we reported only the cytokines that were detectably expressed. The positive control on each membrane was detected, and each experiment was normalized to the controls provided on the filters, as per the manufacturer’s protocol.
Confocal microscopy
Alexa 647–labeled R10-4 were transiently transfected as described above into TNBCs, fixed, stained with DAPI, and imaged by confocal microscopy.
Statistical analysis
Data are mean ± SD. Statistical analysis was performed using one-way ANOVA with Tukey’s post hoc test (*P < 0.05, **P < 0.001).
Results
R10-4 inhibits cancer cell migration and invasion
To investigate whether R10-4 suppresses PAI-1–mediated cancer progression, we examined its effects on TNBCs. Directly adding R10-4 (100–500 nM) to cells had no effect on migration or invasion, because R10-4 did not internalize into the cells (data not shown). Thus, we transfected R10-4 into the cells and assessed transfection efficiency by Alexa 647 labeling and confocal microscopy. We show that R10-4 is localized to the cytoplasm (Fig. 1A). Since the presence of the label can alter the intracellular distribution of aptamers, we then isolated total RNA from the transfected cells and performed an RT-PCR analysis and detected a cDNA band that corresponds to R10-4 and the control scrambled aptamer (Fig. 1B). No band was detected in non-transfected cells (Fig. 1B). While this confirms intracellular uptake, the exact localization of the aptamers remains unknown. We then assessed cell migration using scratch assays. Control cells closed ∼90%, whereas R10-4 transfected cells achieved on average 20%–30% closure, and cells transfected with the control scrambled aptamer (scApt) closed ∼85% within 24 h (Fig. 1B and C). Transwell assays demonstrated a ∼43% reduction in invasion compared to controls (no aptamer and scApt) (Fig. 1D). Cell proliferation was unaffected (data not shown). These results suggest that intracellular R10-4 significantly impairs PAI-1–mediated migration and invasion of TNBC
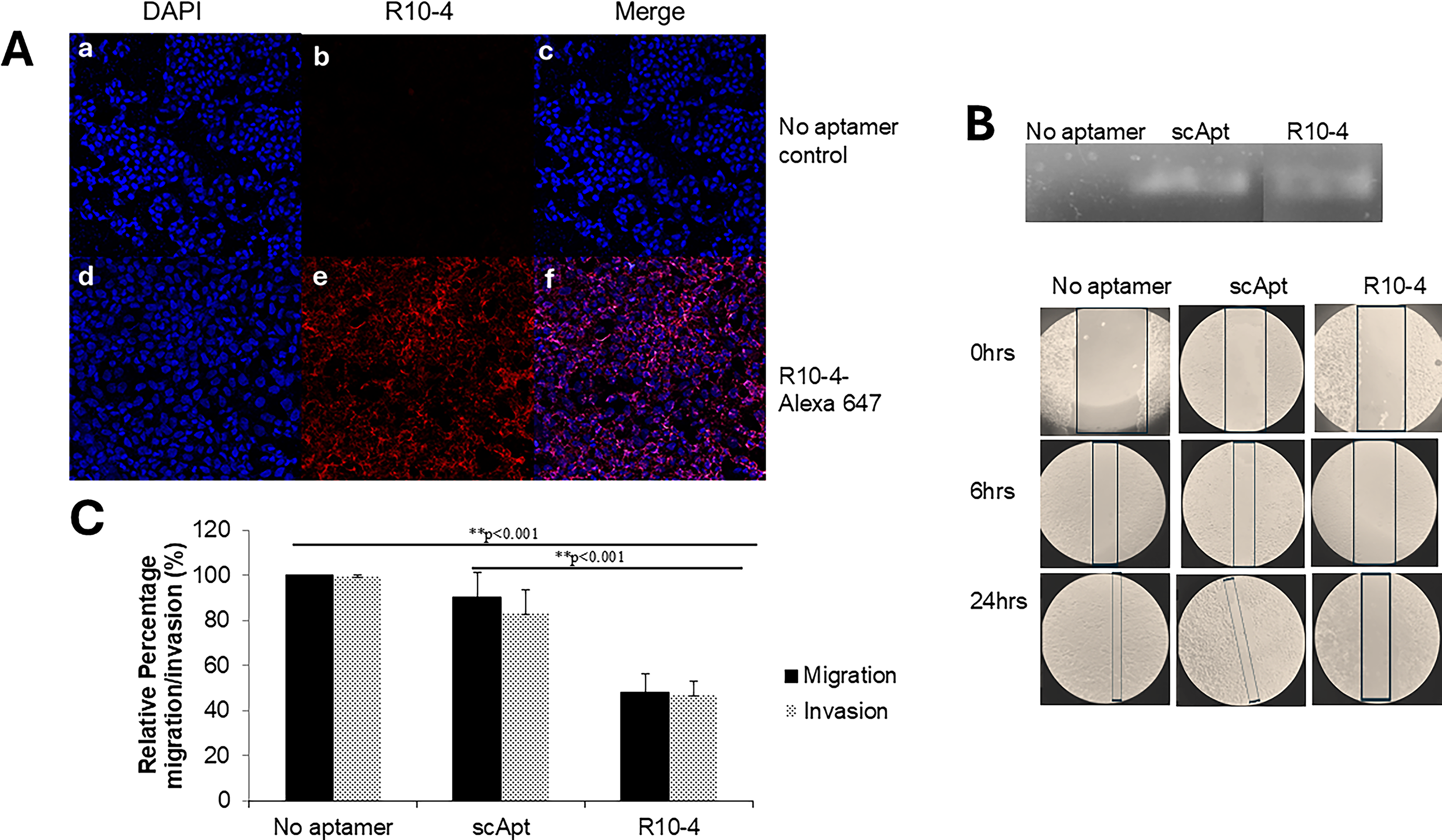
Migration and Invasion of transfected TNBCs:
TNBCs transfected with R10-4 secrete factors that suppress angiogenesis
Because angiogenesis is central to tumor progression, we evaluated whether factors secreted from R10-4 transfected cells alter endothelial tube formation. As expected, HUVECs incubated with normal endothelial cell growth media and with CM from TNBCs transfected with the control scrambled aptamer effectively developed tubes (Fig. 2A). However, incubating HUVECs with CM from cells transfected with 100 nM of R10-4 resulted in a decrease in tube formation (Fig. 2A). Since the CM is Serum Free Medium (SFM: Opti-MEM), we show that SFM alone does not promote tube formation (Fig. 2B and C). CM from cells transfected with R10-4 induced significantly less tubes compared to tubes formed in the presence of CM from cells transfected with scApt. We next assessed the cytokine profiling from transfected cells, which revealed downregulation of CCL5 and MiF in R10-4 transfected cells compared to control cells (Fig. 2D). 17

Analysis of angiogenic tube formation and cytokine expression in response to R10-4 conditioned media.
Discussion
This study identifies R10-4 as a multifunctional RNA aptamer that antagonizes PAI-1, exerting effects on fibrinolysis and tumor progression. Intriguingly, intracellular transfection of R10-4 significantly reduces migration, invasion, and angiogenesis in TNBCs, revealing a previously underappreciated role for PAI-1 within the tumor microenvironment.
Previously, we reported that intracellular transfection of other PAI-1–specific RNA aptamers, SM-20 and WT-15, similarly inhibited breast cancer cell migration and invasion.13,14 However, these aptamers target the VNBD of PAI-1 and do not disrupt the PAI-1/tPA interaction. 13 The observation that aptamers targeting distinct PAI-1 domains produce comparable anti-migratory effects raises questions about the underlying mechanisms. One possibility is that R10-4 engages both the reactive center loop and VNBD of PAI-1, thereby exerting a broader functional inhibition profile.
Our findings also uncover a mechanistic link between PAI-1 and the pro-angiogenic chemokine CCL5, which has been shown previously. 17 CM from R10-4 transfected cells contained markedly reduced levels of CCL5, correlating with impaired angiogenesis. 14 Given CCL5′s role in immune cell recruitment, cancer cell progression, and angiogenesis, suppressing it could represent a downstream consequence of PAI-1 inhibition.18–20 Dissecting PAI-1 and CCL5 synergistic roles in pathological vascularization could lead to the development of targeted strategies to disrupt these processes. Our data show that the intracellular inhibition of PAI-1 by R10-4, resulting in a significant decrease in CCL5, might be the key to mitigating disease progression (Fig. 3). Additionally, we observed downregulation of macrophage migration inhibitory factor (MIF), suggesting that R10-4 may modulate multiple PAI-1–regulated signaling pathways; however, the connection between PAI-1 and MIF is not well established and remains under investigation.
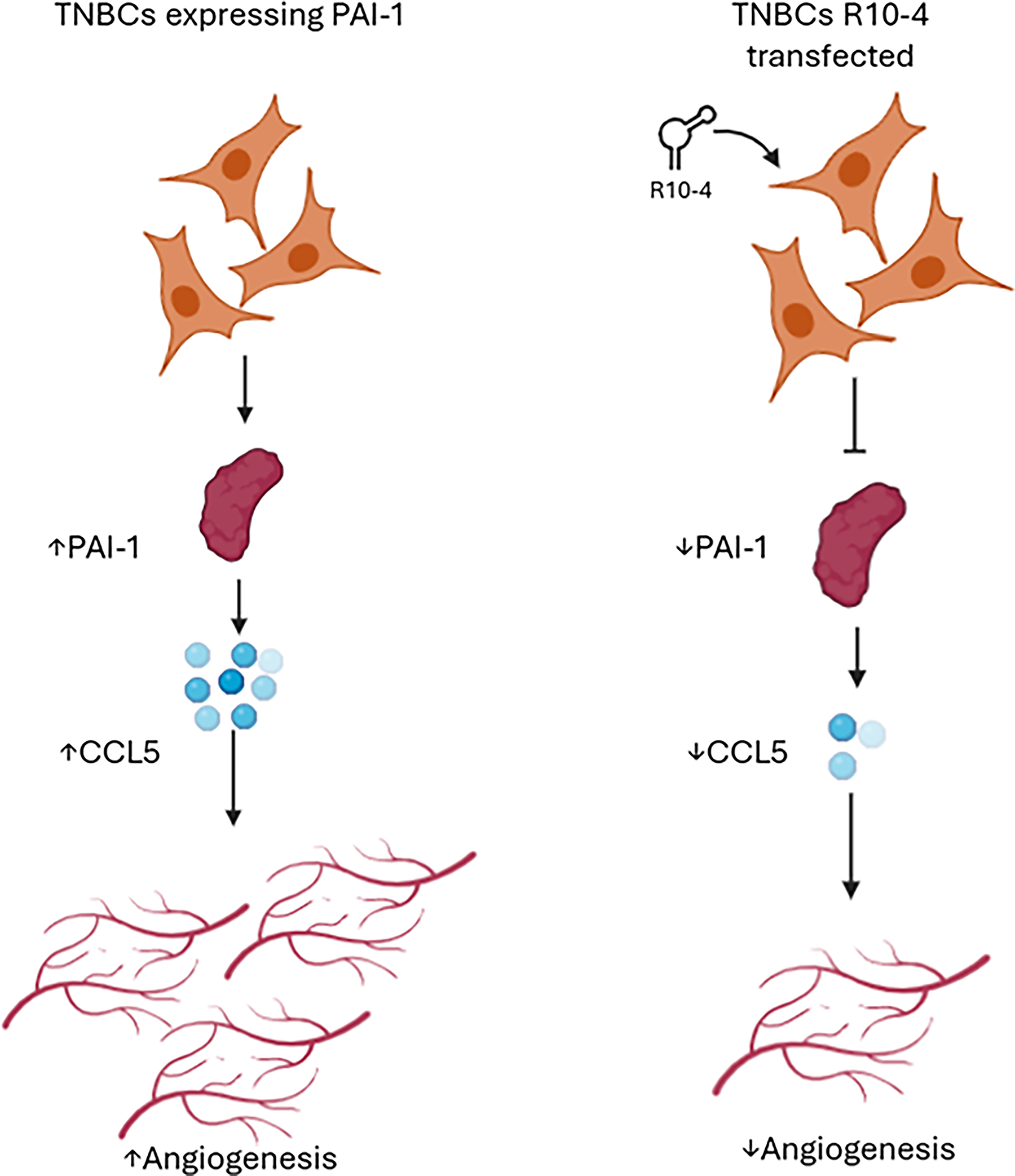
CCL5-mediated angiogenic signaling in TNBCs. Schematic representation of TNBC populations expressing either PAI-1 alone or both PAI-1 and R10-4. These cells secrete the chemokine CCL5 (blue dots), which act on surrounding endothelial cells to promote angiogenesis, illustrated by the formation of new blood vessels. In the presence of R10-4, CCL5 secretion is decreased with an overall decrease in angiogenesis. The diagram highlights a potential mechanism by which TNBCs enhance vascularization through CCL5 signaling, contributing to tumor progression and metastasis. This process is potentially regulated by PAI-1.
Footnotes
Acknowledgments
The authors thank Adi Breitman, Alicia Matthews, and Stephanie Brandal for their early contributions to this work.
Author Disclosure Statement
No financial competing interests exist.
Funding Information
This work was supported by a grant from the Hartwell Foundation (RES236407) and institutional support from Case Western Reserve University College of Arts and Sciences (OPN261229).