
Abstract
Small interfering RNA (siRNA) represents a transformative therapeutic class that enables precise gene silencing through RNA interference (RNAi). N-acetyl galactosamine (GalNAc)-conjugated siRNAs have achieved remarkable clinical success with six FDA-approved therapeutics targeting liver diseases. Following subcutaneous administration, GalNAc–siRNAs rapidly accumulate in hepatocytes via asialoglycoprotein receptor (ASGPR)-mediated endocytosis and are sequestered within endolysosomal compartments, creating an intracellular depot that sustains therapeutic effects for weeks to months. While RNAi-mediated degradation and nuclease metabolism contribute to siRNA clearance, the detection of full-length, active siRNA in circulation long after dosing suggests alternative clearance mechanisms exist. This review examines the pharmacokinetic properties and intracellular trafficking of GalNAc–siRNA, with particular focus on underexplored clearance pathways from hepatocytes. We discuss potential mechanisms including endosomal recycling, efflux via exosomes and lysosomal exocytosis that may facilitate siRNA redistribution from tissues to circulation. Understanding these clearance mechanisms could enable correlation of plasma and tissue drug concentrations, reduce preclinical animal use, inform clinical dosing strategies and guide the design of next-generation siRNA therapeutics with optimized tissue residence time and enhanced therapeutic durability.
Keywords
Introduction
Small interfering RNA (siRNA) has emerged as a powerful class of therapeutics, enabling precise post-transcriptional gene silencing and opening new avenues for treating a wide range of diseases. siRNAs are short double-stranded RNA molecules, typically 20–25 nucleotides, that can downregulate mRNA expression through RNA interference (RNAi). This mechanism requires cytosolic siRNA to incorporate into RNA-induced silencing complexes (RISC) to bind their target mRNA with high specificity and mediate degradation. This leads to reduced target mRNA and protein levels, resulting in the intended downstream biochemical and physiological changes.1,2 RNAi-based therapeutics leverage this mechanism to transiently modulate disease-associated targets. The evidence for the efficacy of this approach is highlighted by seven FDA-approved siRNA therapeutics and many ongoing clinical trials.2–5
Since their initial development, siRNA therapeutics have continued to improve through intentional incorporation of chemical modifications to the oligonucleotides and identification of a wide range of delivery modalities. These enhancements have helped overcome several extracellular and intracellular barriers, improving stability, uptake, specificity and durability of therapeutic effect while reducing renal clearance and toxicity.1,6–8 Among these advances, liver-targeted delivery has emerged as particularly successful, with six of the seven FDA-approved siRNA therapeutics, and many in clinical development, being N-acetyl galactosamine (GalNAc)-siRNA conjugates. 2 Current GalNAc ligands for siRNA delivery are synthetic forms of N-acetylated galactosamine-terminated glycans, which can bind to the asialoglycoprotein receptor (ASGPR), highly and specifically expressed on liver hepatocytes (Fig. 1). 9
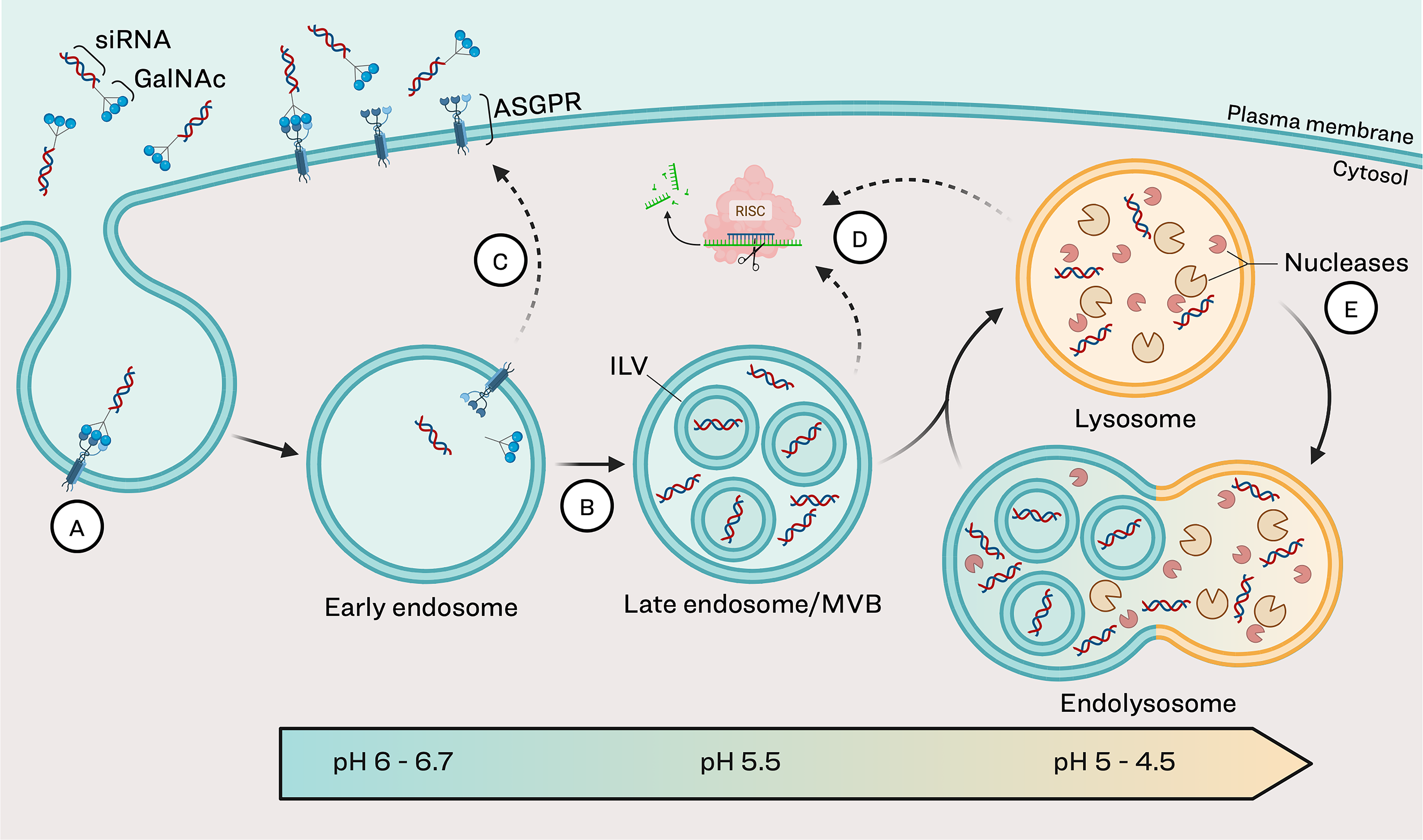
Endolysosomal maturation and siRNA trafficking.
Following binding, the ligand–receptor pair is internalized via endocytosis in clathrin-coated vesicles and trafficked to endosomes. 10 Within the endosomes, GalNAc dissociates from ASGPR, which is quickly recycled back to the cell surface.11–13 This high rate of receptor recycling allows for efficient hepatic uptake of GalNAc–siRNA from peripheral circulation. The bulk of the siRNA sequesters in subcellular compartments within the endolysosomal pathway, 7 mostly in therapeutically active forms (full-length antisense and 3′ n-1), creating a durable intracellular depot. 14 Over time, siRNA is slowly released from the endolysosomal compartments into the cytosol where it is available for RISC loading and subsequent RNAi for sustained mRNA silencing (Fig. 1).10,14–16
While GalNAc–siRNAs have had clinical success, the intracellular mechanisms influencing their pharmacokinetic (PK) behavior and efficacy remain poorly understood. One critical knowledge gap is the process of siRNA clearance from the endolysosomal compartments within hepatocytes. Specifically, there is little known about the pathways that contribute to siRNA redistribution back into circulation. To date, research has largely focused on nuclease-mediated metabolic clearance of siRNA. However, modern chemical modification patterns have markedly stabilized siRNA against degradation. Modifications have evolved from standard template chemistry (STC) through enhanced stability chemistry (ESC), advanced ESC and ESC+, which have progressively increased stability and improved target specificity. 2 The prolonged detection of intact siRNA in both target tissues and circulation suggests that non-metabolic clearance pathways may contribute to elimination of siRNA from the liver. This review examines the PK and intracellular trafficking of GalNAc–siRNA, highlighting known and proposed pathways of clearance from tissue.
PK Properties of GalNAc–siRNA
PK studies quantify how a drug moves through the body over time, focusing on parameters like absorption rate, plasma concentration, half-life, clearance, and volume of distribution. This information is essential for establishing dosing strategies that maximize efficacy and minimize toxicity in patients. Applying insights from PK studies with siRNA compounds of varied chemistries and delivery methods improves understanding of absorption, distribution, metabolism, and excretion (ADME) properties. This characterization facilitates the design of predictive preclinical studies to better inform clinical study design, ultimately accelerating drug development and improving patient outcomes. FDA-approved GalNAc-siRNAs givosiran (Givlaari), lumasiran (Oxlumo), inclisiran (Leqvio), vutrisiran (Amvuttra), nedosiran (Rivfloza), and Qfitlia (fitusiran) have shown similar plasma PK in clinical studies (Table 1),17–28 owing to the GalNAc-driven disposition of these molecules as detailed below.
Clinical Plasma PK Parameters of FDA-Approved GalNAc-siRNAs
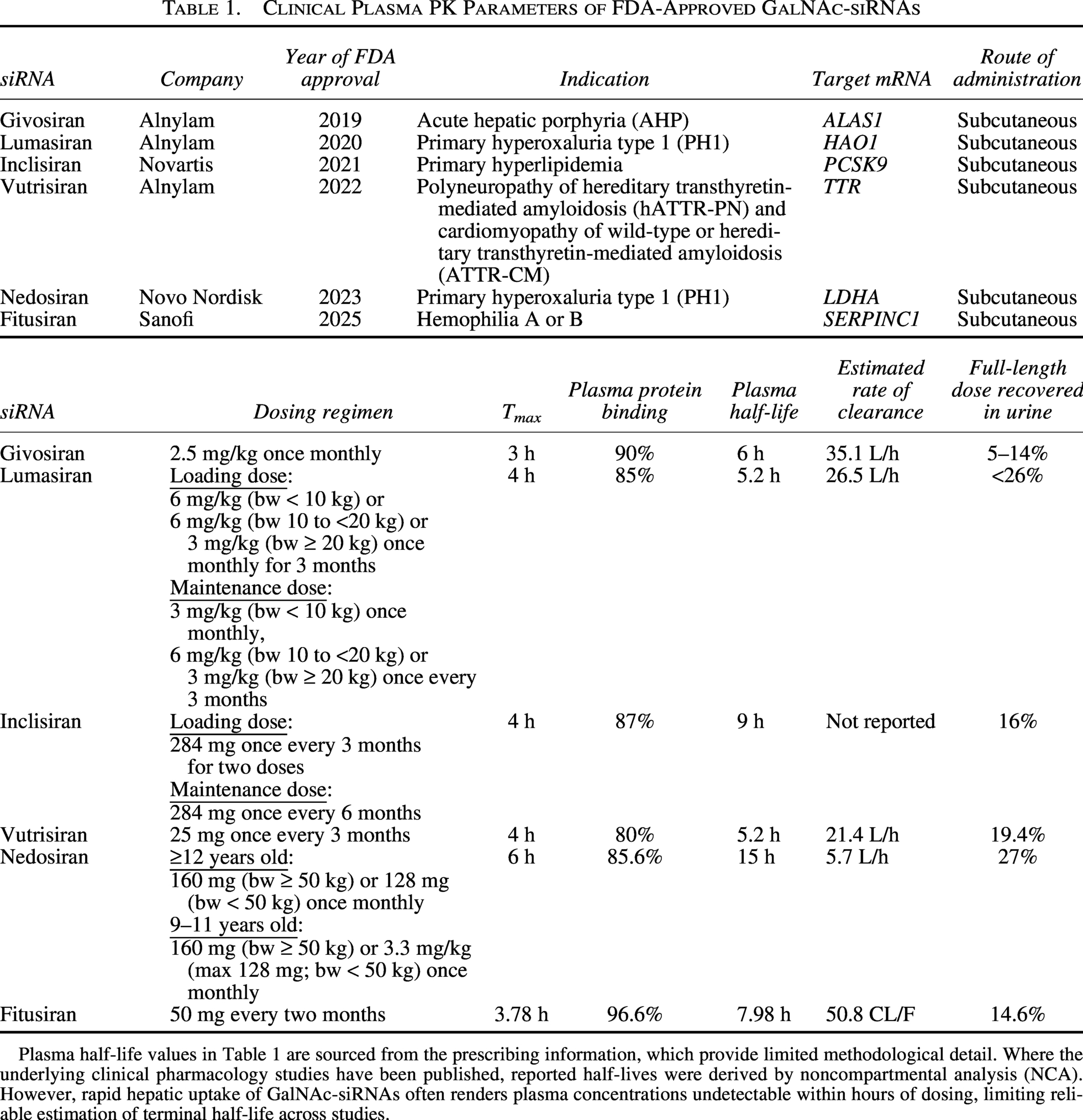
Plasma half-life values in Table 1 are sourced from the prescribing information, which provide limited methodological detail. Where the underlying clinical pharmacology studies have been published, reported half-lives were derived by noncompartmental analysis (NCA). However, rapid hepatic uptake of GalNAc-siRNAs often renders plasma concentrations undetectable within hours of dosing, limiting reliable estimation of terminal half-life across studies.
ADME properties of GalNAc–siRNA
ADME studies follow the fate of a drug from administration to elimination from the body. While route of administration significantly influences ADME characteristics of a drug, all currently approved GalNAc–siRNAs are administered subcutaneously (Table 1). Following subcutaneous injection, GalNAc–siRNAs are quickly absorbed into the peripheral circulation, where they typically undergo minimal metabolism.29,30 In preclinical species, plasma half-lives range from ∼1 to 6 h in rats and ∼2 to 9 h in nonhuman primates.5,29 Half-lives observed in clinical studies are similar to preclinical findings, with half-lives of ∼5–15 h (Table 1). Additionally, allometric scaling approaches have proven reliable for predicting human plasma PK parameters from preclinical data.29,30
Following absorption, GalNAc–siRNAs are rapidly cleared from the bloodstream and primarily accumulate in the liver, with some distribution to the kidneys.5,29,31 Nair et al. demonstrated in mice that the maximum concentration (Cmax) was more than 10 times higher in liver than in the kidney, with liver time to Cmax (Tmax) occurring in half the time. 31 While liver samples are not typically collected in clinical studies, liver half-life can be estimated using PK modeling based on clinical and preclinical plasma biomarker data, siRNA dosing and siRNA elimination profiles. One PK study estimated liver half-life for 11 clinical stage GalNAc–siRNAs where the predicted liver half-lives ranged from 0.6 to 3 weeks in mice, 1 to 8 weeks in monkeys, and 1.5 to 14 weeks in humans. 32 The authors determined that the liver half-life in humans is 1–8 and 1–3 times longer than in mice and monkeys, respectively. Their model with fitusiran predicted a liver half-life in humans of around 3 weeks, which is similar to previous predictions. The paper found no clear relationship between chemical modification type and liver half-life across STC, ESC, ESC + and Arrowhead chemistry. However, the conclusions are limited by data availability, which may have prevented the identification of chemistry-dependent trends. In contrast, other studies have established a relationship between chemistry and metabolic stability. Work in preclinical species shows that more chemically stable siRNA persists in the liver longer, even at lower doses, compared to a less stable version dosed >3-fold higher.7,33siRNA elimination can occur through metabolism within tissues and through excretion. While much of the siRNA is likely excreted as metabolites, preclinical and clinical studies show that a portion of the siRNA recovered in urine is intact. Preclinical data show that 9–25% of the administered dose is recovered as full-length in urine across lumasiran, givosiran, and vutrisiran. 29 Clinical studies across the FDA-approved GalNAc–siRNAs show similar recovery (Table 1). Nedosiran, the only molecule that uses an ESC+ chemical modification pattern, has the highest recovery in urine at 27%. However, the remaining molecules all use ESC chemistry and fall within a comparable range of 5–26%. This suggests that additional factors outside of chemistry influence metabolic stability.
PK basis for proposed intracellular clearance pathways
As noted above, GalNAc–siRNAs have much longer half-lives in liver than in plasma. Plasma concentrations often fall below detectable levels within hours of dosing using methods like HPLC and mass spectrometry.5,29–31 This rapid plasma clearance limits the utility of using plasma PK measurements to predict trends such as steady-state liver concentrations, where the tissue and plasma concentration ratio stabilizes. However, highly sensitive detection methods have revealed a more complete picture of siRNA disposition. In preclinical studies, PCR-based assays can capture plasma siRNA concentrations through extended timepoints postdose and demonstrate this steady-state of elimination. 33 Using these sensitive methods, siRNA delivered through GalNAc conjugation can be detected within the peripheral circulation in minimally metabolized forms for >3 months following a single dose. 33 Together, these observations suggest a disposition profile that cannot be explained by plasma-phase kinetics alone. The short plasma half-life reflects efficient hepatic uptake driven by ASGPR-mediated endocytosis rather than systemic elimination. The prolonged hepatic retention implies that once sequestered within endolysosomal compartments, siRNA is eliminated from the tissue slowly through mechanisms that are largely not understood. Critically, the recovery of full-length and minimally metabolized siRNA in plasma suggests that a portion of the tissue depot re-enters circulation in active forms, which is inconsistent with purely degradative clearance. A closer examination of intracellular trafficking pathways is needed to identify factors that influence the rate and form of siRNA recirculation.
Intracellular Kinetics of GalNAc–siRNA
The endolysosomal pathway is the primary intracellular route through which GalNAc-siRNA is trafficked following receptor-mediated endocytosis. In this pathway, internalized cargo progresses through increasingly acidic membrane-bound compartments where it is sorted, processed, and either recycled or degraded. Understanding the kinetics of GalNAc–siRNA movement through this pathway is critical for defining the intracellular determinants of its PK behavior and therapeutic activity.
Endolysosomal pathway and uptake of GalNAc–siRNA
The endolysosomal pathway allows transient storage and recycling of cellular cargo with eventual degradation or extracellular release (Fig. 1). Extracellular material enters this pathway through endocytic vesicles that form through invagination of the plasma membrane. These vesicles fuse to form early endosomes, where sorting of internalized cargo occurs. Early endosomes undergo subsequent maturation steps, eventually becoming late endosomes that fuse with lysosomes, the primary digestive compartment of the cell.34–37 A key feature of endosomal maturation is a gradual reduction in pH, dropping from 6.0–6.7 in early endosomes to 5.5 in late endosomes and 4.5–5 in lysosomes. 5 This acidification is critical for processing internalized GalNAc–siRNA: the low pH facilitates dissociation of GalNAc–siRNA from ASGPR, freeing up ASGPR to be recycled back to the cell surface, where it can bind additional GalNAc–siRNA.11,13,38 Within the acidic endosomal environment, β-N-acetyl glucosaminidase cleaves GalNAc moieties from oligonucleotides.29,39 Studies with GalNAc-conjugated antisense oligonucleotides (ASOs) in mice demonstrate that cleavage occurs rapidly, with substantial GalNAc cleavage within 1 h and complete cleavage by 72 h in liver samples. 40
Endosomal entrapment and metabolic stability in intracellular depots
Following its initial processing, siRNA accumulates in the endolysosomal pathway, a phenomenon referred to as endosomal entrapment. Live imaging studies in mouse primary hepatocytes demonstrated rapid accumulation, where GalNAc–siRNA colocalized with endolysosomal compartments in <90 min. 7 While the exact identity of the compartments that sequester oligonucleotides has not been fully defined, late endosomes and lysosomes are thought to serve as primary intracellular depots.7,29,41 Metabolic stability conferred by chemical modifications is thought to preserve siRNA functionality during gradual release from the endolysosomal depots into the cytosol for sustained therapeutic effect. 14
Endosomal escape
For siRNA to mediate transcript silencing, it must leave the endolysosomal storage compartments and enter the cytosol where it can access RISC machinery.42,43 This process of release, known as endosomal escape, is thought to occur through brief, localized disruptions in the lipid bilayer of endosomes. 14 These disruptions may arise from localized membrane instabilities or from fusion events along the endolysosomal pathway, such as when maturing endosomes fuse with lysosomes. 16 Brief perturbations in endosomal membrane continuity are thought to allow nearby siRNA to escape into the cytoplasm. However, release into the cytosol is thought to occur infrequently and in extremely small amounts, with cytosolic bioavailability of GalNAc-delivered siRNA estimated at 0.01–1% of total endocytosed siRNA, which is why endosomal escape is considered the rate-limiting step for RNA therapeutics.10,14–16 While beyond the scope of this review, strategies to enhance endosomal escape are an active area of research aimed at improving siRNA durability and efficacy.44,45
Current Understanding of siRNA Clearance from Target Tissue
Once internalized, siRNA can be cleared through degradation, resulting in loss of therapeutic activity. The two most widely accepted clearance mechanisms include nuclease-mediated degradation and Argonaute2 (Ago2)-mediated turnover, both of which are generally understood to result in breakdown of the siRNA molecule itself. While grounded in RNA biology, there are gaps in the mechanistic understanding of each, and their precise contributions to GalNAc–siRNA clearance in a therapeutic context are incompletely defined.
Nuclease-mediated degradation of siRNA
Nuclease-mediated degradation is the clearance mechanism that is most supported through the wide reporting of siRNA degradation in the liver in both in vitro and in vivo studies.29,46,47 Nucleases are a subclass of hydrolases with a preference for oligonucleotides as their substrate. Nucleases are further characterized by where within the oligonucleotide they cleave preferentially. Endonucleases tend to cleave phosphodiester linkages within the strand to generate shortmers, while exonucleases typically remove single nucleotides from the 3′ or 5′ end. 48 Within the endolysosomal compartments, lysosomal enzymes are the most well characterized, containing over 60 different hydrolases that can degrade a diverse range of molecules, including RNAs.49–51 While specific nucleases that metabolize siRNA within different cellular compartments have not been defined, a variety of nucleases have been identified that may play a role.39,52–55 Comprehensive characterization of intracellular nuclease populations and systematic profiling of their activity against chemically modified siRNAs are needed to address this gap.
siRNA clearance associated with Ago2 turnover
Another commonly recognized clearance mechanism involves the engagement of siRNA with Ago2, the catalytic component of RISC. 56 RNAi with siRNA begins when the antisense strand is loaded into Ago2, which unwinds the siRNA, cleaves the sense strand and retains the antisense strand. Antisense-loaded Ago2 then binds target mRNA through sequence complementarity and mediates transcript degradation. 57 The antisense-loaded complex can catalyze multiple rounds of mRNA cleavage until Ago2 itself undergoes degradation, 58 with a half-life exceeding 5 days in vitro 59 and approximately 3.8 days in mice. 7
The fate of the Ago2-loaded antisense strand is not well-defined but is thought to be degraded along with Ago2. Measuring Ago2-bound siRNA provides a functional readout of the therapeutically active cytosolic pool. Several studies have demonstrated that Ago2-loaded siRNA concentrations correlate more closely with mRNA knockdown than total liver siRNA levels.2,7,31,33 Ago2-bound siRNA levels specifically capture the cytosolic fraction that is actively engaged in target suppression. Total liver measurements not only include the large, inactive endolysosomal depot, but also some amount of uptake into nonparenchymal cells within the liver that may have varying levels of target expression. 40
While Ago2 half-life in mice is on the order of days, clinical and preclinical studies have shown that GalNAc–siRNAs have a duration of effect ranging from weeks to months. 2 The limited siRNA cytosolic bioavailability described earlier puts constraints on the magnitude and durability of knockdown. It has been proposed that the siRNA released from the endolysosomal depots replaces cleared antisense strands and is loaded into new Ago2 at a rate dependent on the cytosolic availability of the siRNA and Ago2. 7
Proposed Mechanisms of siRNA Clearance from Target Tissue
While clearance via Ago2 turnover and nuclease-mediated degradation involve the breakdown of siRNA, rendering it therapeutically inactive, evidence for minimally metabolized siRNA in systemic circulation 33 suggests that additional elimination pathways may exist. Given that the vast majority of internalized siRNA resides within the endolysosomal pathway, these compartments represent a plausible route by which intact siRNA may be redistributed to systemic circulation. The following sections speculate on potential exit points for siRNA from the endolysosomal pathway, drawing on established cell biology in the absence of direct mechanistic evidence. These proposed mechanisms may facilitate clearance of both therapeutically active and degraded siRNA, though the relative contributions of each remain unclear.
Efflux of intracellular cargo from endosomes
As described earlier, early endosomes are the initial sorting site for internalized cargo. While some reports define “recycling endosomes” as a specific subclass, reliable molecular markers to distinguish these from other early endosomes have not been well defined. 60 For the purpose of this review, they will be referred to as early endosomes. Within early endosomes, internalized cargo can either be retained as the endosome matures or recycled back to the cell surface. 34 This raises the possibility that recycling may contribute to siRNA redistribution (Fig. 2). Extensive recycling from early endosomes would reduce intracellular siRNA retention, potentially limiting therapeutic activity. Detection of nonvesicular siRNA relatively soon after uptake into cells would provide evidence to support this as a redistribution pathway.
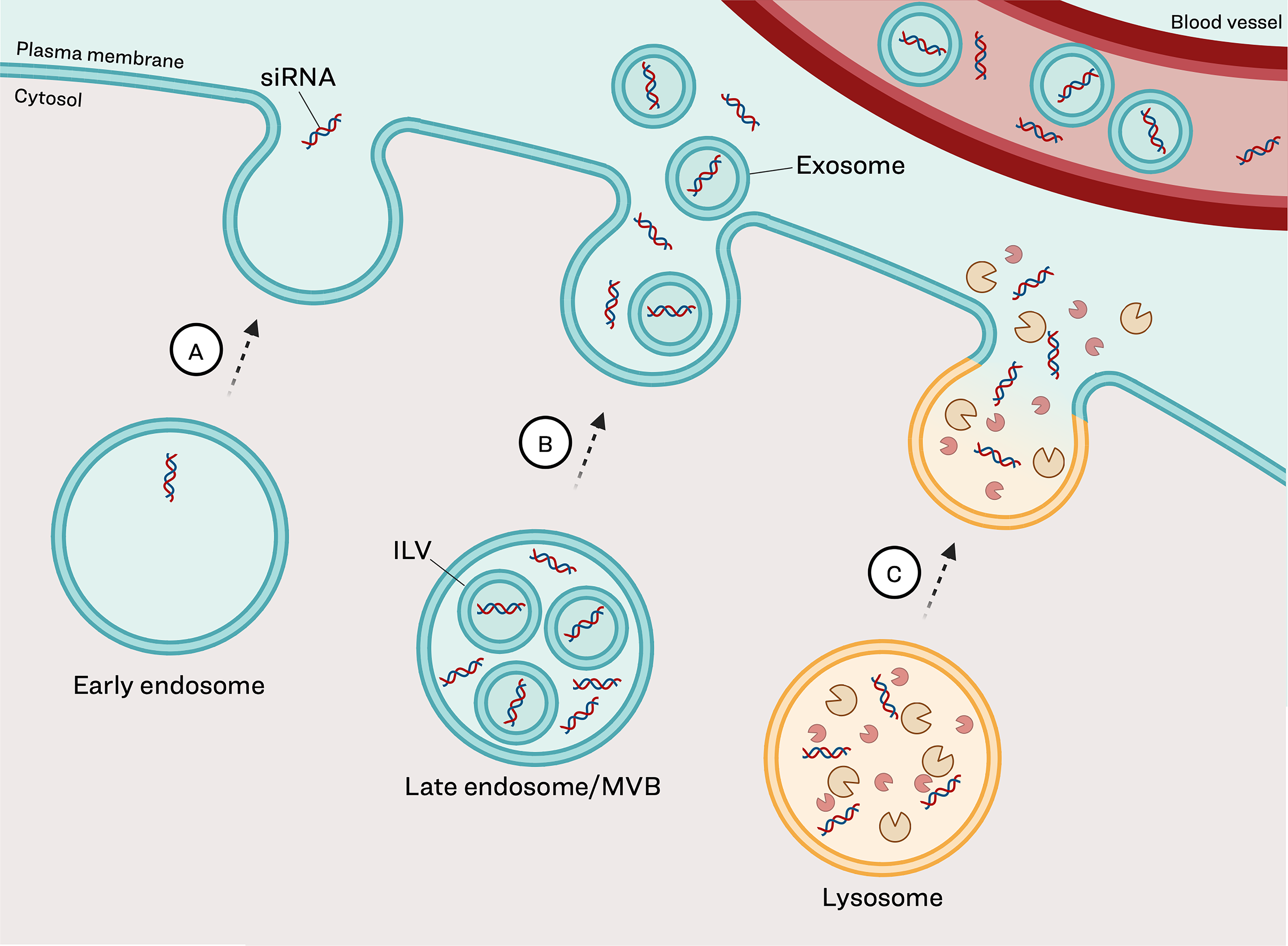
Potential mechanisms of siRNA redistribution from the endolysosomal pathway. Following receptor-mediated endocytosis, a fraction of full-length siRNA can be recirculated from tissues back into systemic circulation.
In addition to direct recycling, cargo can be expelled from cells within extracellular vesicles (EVs). Under physiological conditions, this process is an important cellular mechanism necessary for timely elimination of intracellular cargo. EVs are typically classified based on their biogenesis, as the currently identified markers are insufficient to uniquely distinguish different classes of EVs. 61 EVs originating from endosomes are termed exosomes and form during the endosomal maturation pathway (Fig. 2). Traditionally, new cargo would enter the cell in early endosomes, which mature to late endosomes, also called multivesicular bodies (MVB), which then fuse with lysosomes for degradation. Alternatively, MVBs, which contain intralumenal vesicles (ILVs), can recycle back to the cell membrane where the ILVs can be released as exosomes into the extracellular space. There is evidence through both in vitro and in vivo studies that endogenous RNAs traffic through the endolysosomal pathway and can be detected in exosomes.62–64 There is little understanding or consensus on how endogenous miRNAs come to be found in exosomes but both nonselective and selective active sorting via the specific sequence motifs have been proposed.61,65,66 The presence of similar mechanisms to influence siRNA sorting into exosomes remains to be determined. Detection of therapeutically active siRNA within exosomes would provide compelling evidence that exosome-mediated export may contribute to GalNAc–siRNA clearance from target tissue.
Clearance of intracellular cargo from lysosomes
Lysosomes represent another intracellular depot for siRNA and a potential source of cellular efflux. 51 A proposed siRNA clearance mechanism from lysosomes is lysosomal exocytosis, which is the bulk release of cargo into the extracellular space via fusion of lysosomal and plasma membranes (Fig. 2). This is a normal physiological process that occurs broadly across all cell types to eliminate accumulated cargo and maintain homeostasis. 67 For siRNA therapeutics, lysosomal exocytosis would result in direct release of siRNA into the extracellular space, potentially enabling reuptake into systemic circulation. A key question is whether accumulation of siRNA therapeutics can specifically trigger or increase this efflux pathway. Characterizing the role of lysosomal exocytosis in siRNA clearance could enhance our understanding of hepatocyte redistribution kinetics and inform strategies to modulate tissue retention.
Efflux of siRNA from the cytosol
There are additional efflux pathways that may facilitate clearance of cytosolic siRNA. One hypothesis is that nucleic acid therapeutics could be cleared from the cytosol through ectosomes—EVs that form from outward budding of the plasma membrane and contain cytosolic contents.36,68 While not well understood, Ago2 complexed with endogenous miRNA and siRNA has been detected in EVs69,70; however, a much larger portion of Ago-RNA has been detected in bodily fluids outside of vesicles from unknown origins.71,72 Additionally, siRNA may be cleared from the cytoplasm through secretory autophagy, a process in which cytosolic contents are engulfed and, rather than being targeted for degradation in the lysosome, cargo is released into extracellular space.73,74 Finally, if siRNA is released from lysosomes into the cytosol, it could either re-enter the endolysosomal pathway or undergo exocytosis. Given the limited cytosolic bioavailability discussed earlier, cytosolic clearance pathways likely contribute minimally to siRNA redistribution.
Methodological Considerations and Limitations
The evidence reviewed here relies heavily on analytical methods, some of which have inherent limitations that should be considered when making mechanistic conclusions about redistribution of siRNA. First, while PCR-based detection methods are highly sensitive, there is still some level of detection beyond full-length and AS 3′ n-1, albeit with reduced sensitivity. 75 Given that AS 3′ n-1 is often the predominant metabolite detected,22,40,41 the assay generally captures the therapeutically relevant siRNA fraction. However, it is worth noting that the structural identity of the detected species is typically inferred rather than directly confirmed in PCR-based studies. Fluorescence-based imaging has provided valuable spatial information about GalNAc-siRNA colocalization with endolysosomal markers, supporting models of intracellular depot formation. 7 However, limitations include altered uptake and cellular trafficking from dye conjugation, false-positive signals from fluorophore detachment and difficulty distinguishing cytosolic from endolysosomally located siRNA. 43 Finally, PK modeling is inherently an indirect measurement used to estimate liver half-life. Model-derived liver half-life values depend on assumptions embedded in the model, including the relationship between plasma and tissue kinetics. 76 These estimates should therefore be understood as model-dependent approximations and not interpreted as direct measurements.
Significance of Understanding siRNA Cellular Efflux Pathways
The characterization of siRNA efflux pathways carries significant translational value. A better understanding of how siRNA redistributes from tissues to circulation could improve both preclinical study design and clinical monitoring while informing the engineering of more durable therapeutics. This section examines two key applications: leveraging steady-state plasma-tissue relationships for PK monitoring and exploiting clearance mechanisms to design more durable RNA therapeutics.
Correlating plasma and tissue concentrations of siRNA at steady state
The redistribution of the largely unmetabolized, and therefore active, siRNA molecules from target tissues to plasma at steady state of elimination could allow for indirect measurement of siRNA tissue concentrations. The ability to extrapolate tissue concentrations from plasma would enable the use of fewer animals in preclinical testing due to the reduced reliance on terminal tissue collections. Additionally, this methodology could enable monitoring of tissue concentrations in the clinic, where plasma is often the most readily available biomatrix. Achieving this correlation would require clinical studies with plasma sampling extended beyond the distribution phase to capture steady-state elimination kinetics with relevant sensitive detection methods. Pharmacodynamic biomarker data could provide an indirect estimate of tissue drug levels, but direct validation may require liver biopsy samples, where available, to confirm the plasma-tissue relationship. Together, these capabilities would strengthen both preclinical and clinical study design for RNA therapeutics.
Leveraging efflux pathways to improve durability of exposure and effect
Understanding the different factors that influence siRNA tissue clearance could inform the design of more durable therapeutics. Currently there is a significant focus on improving siRNA uptake through novel ligand conjugation, increasing siRNA stability and durability against metabolism, and enhancing cytoplasmic availability through manipulation of endosomal escape mechanisms.44,45,77,78 However, these enhancements often fail to improve tissue residence time, which may play a role in the durability of therapeutic effect. The existing approach to achieve more durable effects is often to use higher dose levels, which may have consequent tolerability issues. With better characterization of intracellular kinetics and tissue clearance mechanisms, it may be possible to engineer siRNA that minimizes tissue clearance and extends duration of effect without increasing dose. Mechanistic knowledge would therefore help identify the factors that can be leveraged to modulate siRNA behavior and build the next generation of RNA therapeutics.
Conclusion
Understanding siRNA clearance represents a critical frontier for RNA therapeutic development. Future research should aim to characterize mechanisms of siRNA efflux with an emphasis on the molecular determinants that govern siRNA sorting into EVs and the capacity for siRNA to induce lysosomal exocytosis. Integrating mechanistic insights into PK models has the potential to improve capabilities for dose projections, enabling safer and more efficient study designs. Ultimately, the ability to engineer siRNA molecules with optimized clearance profiles could enhance therapeutic durability and expand the scope of RNAi-based therapeutics.
Footnotes
Acknowledgments
The authors would like to thank Christopher M. Wiethoff, Michael Moazami, and John J. Rhoden for taking the time to review this article and for providing insightful feedback that improved the final work. The authors are also grateful to Bradley L. Ackermann for his guidance throughout the process of publication of this article.
Author Disclosure Statement
M.C.T., K.C., and S.D. are employees of Eli Lilly and Company and may own stock or stock options in the company. The authors declare no other competing interests.
Funding Information
This work was funded by Eli Lilly and Company. All authors are employees of Eli Lilly and Company.